Abstract
Acinetobacter baumannii is an important cause of nosocomial infections worldwide. The elucidation of the carbapenem resistance mechanisms of hospital strains is necessary for the effective treatment and prevention of resistance gene transmission. The main mechanism of carbapenem resistance in A. baumannii is carbapenemases, whose expressions are affected by the presence of insertion sequences (ISs) upstream of blaCHDL genes. In this study, 61 imipenem-nonsusceptible A. baumannii isolates were characterized using phenotypic (drug-susceptibility profile using CarbaAcineto NP) and molecular methods. Pulsed field gel electrophoresis (PFGE) and multi-locus sequence typing (MLST) methods were utilized for the genotyping. The majority of isolates (59/61) carried one of the following acquired blaCHDL genes: blaOXA-24-like (39/59), ISAba1-blaOXA-23-like (14/59) or ISAba3-blaOXA-58-like (6/59). Whole genome sequence analysis of 15 selected isolates identified the following intrinsic blaOXA-66 (OXA-51-like; n = 15) and acquired class D β-lactamases (CHDLs): ISAba1-blaOXA-23 (OXA-23-like; n = 7), ISAba3-blaOXA-58-ISAba3 (OXA-58-like; n = 2) and blaOXA-72 (OXA-24-like; n = 6). The isolates were classified into 21 pulsotypes using PFGE, and the representative 15 isolates were found to belong to sequence type ST2 of the Pasteur MLST scheme from the global IC2 clone. The Oxford MLST scheme revealed the diversity among these studied isolates, and identified five sequence types (ST195, ST208, ST208/ST1806, ST348 and ST425). CHDL-type carbapenemases and insertion elements upstream of the blaCHDL genes were found to be widespread among Polish A. baumannii clinical isolates, and this contributed to their carbapenem resistance.
1. Introduction
In February 2017, the World Health Organization (WHO) published a list of the antibiotic-resistant “priority pathogens” that pose the greatest threat to human health [1]. Carbapenem-resistant Acinetobacter baumannii was classified as the most pressing threat in “the critical group”. This non-fermentative, Gram-negative coccobacillus is one of the main causes of severe nosocomial infections, such as ventilator-associated pneumonia, bloodstream infections, bacteremia, urinary tract infections and wound infections, especially in burn patients and post-surgical procedures [2,3]. The increased incidence of carbapenem resistance among clinical isolates, which has risen dramatically over the last twenty years, is of major concern. Resistance to carbapenems in A. baumannii is mediated mainly by the production of various carbapenem-hydrolyzing β-lactamases belonging to Ambler classes A (KPC and GES groups), B (IMP, VIM, SIM and NDM groups) and D (OXA-51-like, OXA-23-like, OXA-24-like, OXA-58-like, OXA-143-like and OXA-235-like) [4]. Among the clinical isolates of A. baumannii, carbapenem-hydrolyzing class D β-lactamases (CHDLs), also called serine oxacillinases, are the most prevalent. Although they exhibit a weak hydrolysis of carbapenems [3], CHDLs may confer high levels of carbapenem resistance when an insertion sequence (IS) is located upstream of the gene, providing a strong promoter that leads to the overexpression of blaCHDL genes [5]. Genes coding for OXA-51-like enzymes, unlike other CHDL enzymes, occur naturally in all A. baumannii strains [6].
In this study, the contribution of different carbapenemases to carbapenem resistance among clinical A. baumannii strains isolated in one tertiary hospital in Warsaw, Poland, was determined, and 61 imipenem-nonsusceptible isolates were characterized using phenotypic and molecular methods, including genotyping of the isolates.
2. Results
2.1. Distribution of blaCHDL Genes and Insertion Sequences
Among all 61 of the collected non-duplicate, imipenem-nonsusceptible clinical isolates of the Acinetobacter calcoaceticus—A. baumannii complex, the identification of the A. baumannii species was confirmed using gyrB multiplex PCR. PCR analysis of the blaCHDL genes occurrence among these isolates revealed the presence of intrinsic blaOXA-51-like genes and following acquired blaCHDL genes: blaOXA-23-like in 14 of the 61 (23%) isolates, blaOXA-24-like in 39 of the 61 (64%) isolates and blaOXA-58-like in 6 of the 61 (10%) isolates. Furthermore, 59 of the 61 isolates harbored no more than two blaCHDL genes (blaOXA-51-like and one of the acquired blaOXA-23/OXA-24/OXA-58 genes). The blaOXA-143-like genes were not detected in any of the isolates. Two isolates (3%) carried only blaOXA-51-like genes, and in these two cases, ISAba1 was found upstream of the genes encoding the OXA-51-like enzymes. Each of the blaOXA-23-like and blaOXA-58-like genes had an upstream ISAba1 and ISAba3 insertion respectively. Whole genome sequencing was performed on 15 selected A. baumannii isolates, including the representative isolates containing each acquired OXA-possessing group, with differing β-lactam sensitivity profiles and CarbAcineto NP assay results, including the time after which positive carbapenemase activity was detected (Table S1). The whole genome datasets of the 15 strains generated and analyzed during the current study are available at the NCBI BioProject repository (SubmissionID: SUB9082120, BioProject ID: PRJNA701882). Whole genome sequencing (WGS) analysis identified the blaOXA-66 gene encoding intrinsic OXA-66 belonging to the OXA-51-like family, as well as the blaADC-30 gene encoding ADC-30 from the AmpC cephalosporinase family, in all of the isolates. Among the acquired CHDLs and IS elements, the genome analysis revealed the presence of blaOXA-72 (OXA-24-like family) in six isolates (no. 76, 81, 159, 165, 176 and 195), ISAba1 upstream of the blaOXA-23 gene (OXA-23-like family) in five isolates (no. 96, 113, 118, 129 and 185) and ISAba3 upstream and downstream of the blaOXA-58 (OXA-58-like family) in two isolates (no. 43 and 52;Figure 1). Additionally, in the case of five isolates (no. 86, 87, 96, 129 and 185), the presence of a blaTEM-1 gene encoding a β-lactamase with a narrow spectrum of activity was demonstrated.
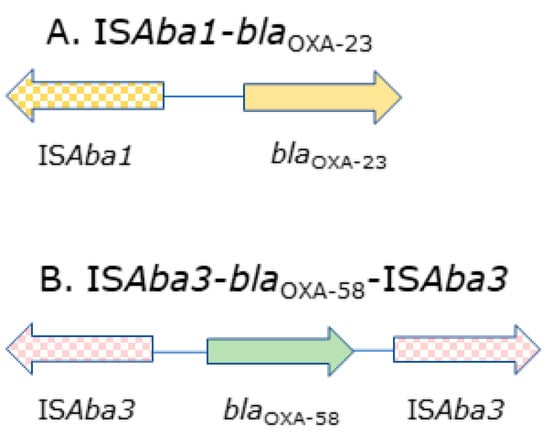
Figure 1.
The intrinsic blaCHDL genes with insertion sequence (IS) element clusters found in the following clinical isolates: (A) no. 86, 87, 96, 113, 118, 129 and 185; (B) no. 43 and 52.
2.2. Antimicrobial Agent Resistance Profiles in Relation to blaCHDL Genes Presence
Table 1 shows the susceptibility pattern of 61 A. baumannii isolates for 13 antibacterial agents. The minimal inhibitory concentration (MIC) distribution for the carbapenems, imipenem and meropenem, is shown in Table 2. All 61 A. baumannii isolates were determined to be colistin-sensitive, and resistant to ciprofloxacin, piperacillin and piperacillin with tazobactam, regardless of the blaCHDL gene carriage. The majority of isolates were resistant to trimethoprim–sulfamethoxazole, with the exception of 5 out of the 39 blaOXA-24-like-carrying isolates. Of the isolates carrying the acquired blaCHDL genes, all isolates containing the ISAba1-blaOXA-23-like and blaOXA-24-like genes were resistant to both of the carbapenems tested. The highest imipenem and meropenem MIC values were obtained for blaOXA-24-likecarrying isolates (64 mg/L and 128 mg/L, respectively).

Table 1.
The susceptibility profiles of A. baumannii isolates (n = 61) carrying blaCHDL genes.
Table 2.
The carbapenem minimal inhibitory concentration (MIC) distribution of the blaCHDL-carrying A. baumannii isolates (n = 61).
2.3. Phenotypic Detection of Carbapenemases
Positive carbapenemase activity results using the CarbAcineto NP Test were obtained for the majority of ISAba1-blaOXA-23-like- (n = 12/14) and blaOXA-24-like-carrying (n = 34/39) isolates. For the other blaOXA-23-like- and blaOXA-24-like-carrying isolates, as well as all of the isolates containing ISAba3-blaOXA-58-like (n = 6) and ISAba1-blaOXA-51-like genes (n = 2), uninterpretable (the optical reading indicated only a slight change in color for the phenol red solution in the test tube) results were observed. In these cases, compared to the internal control (red color tube), after the required incubation time (maximum 2 h), only the development of a red-orange color was observed in the test tube, but not the expected yellow or orange color. The test was performed in duplicate.
Moreover, none of the 61 studied isolates showed MBL-type enzymes based on the results obtained from the double disc diffusion method (DDST) using ethylenediaminetetraacetic acid (EDTA) assay, imipenem and ceftazidime discs.
2.4. Molecular Typing and Genome Analysis of the Isolates
To understand how closely related all 61 A. baumannii isolates are, they were genotyped using pulsed field gel electrophoresis (PFGE) (Figure 2). Twenty-one pulsotypes (PTs) were determined. The isolates were considered to be a cluster when their similarity was at least 80%. Isolates harboring the blaOXA-24-like, ISAba1-blaOXA-23-like and ISAba3-blaOXA-58-like genes were clustered into 12 PTs, 5 PTs and 3 PTs, respectively. Two isolates carrying only the ISAba3-blaOXA-51-like gene were clustered into two different PTs. Among isolates harboring the blaOXA-24-like gene, the majority were isolated in 2012 and 2013 (31/39, 80%) and belonged to 11 different clusters. Most isolates obtained in 2010 and 2011with the blaOXA-24-like gene formed a separate cluster (PT A) and for five of these isolates a similarity of over 98% was observed. One isolate from 2012 was also mapped to the PT A pulsotype.
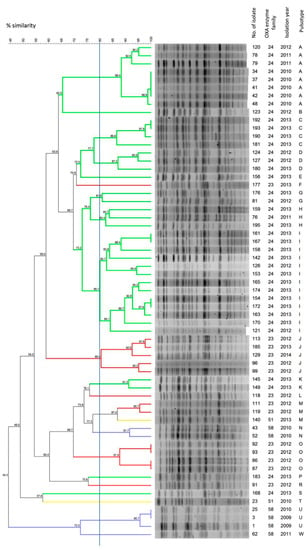
Figure 2.
Analysis of pulsed field gel electrophoresis (PFGE) patterns. The dendrogram presents PFGE profiles’ percentage similarity, the presence of the OXA enzyme family and the isolation year of the A. baumannii clinical isolates. The solid line indicates 80% similarity and is used to define the pulsotypes. Isolates carrying different groups of the acquired blaCHDL genes were marked with the following colors: red, blaOXA-23-like; green, blaOXA-24-like; and blue, blaOXA-58-like. Isolates harboring only intrinsic blaOXA-58-like gene were marked in yellow.
Whole genome analysis of the subsection of isolates (no. 43, 52, 76, 81, 86, 87, 96, 113, 118, 129, 159, 165, 176, 185 and 195) containing the acquired blaCHDL genes and from a variety of the pulsotypes revealed that, when multi-locus sequence typing (MLST) typed, they all belonged to the Pasteur sequence type 2 (ST2). The Oxford MLST scheme distinguished five STs among the whole genome sequenced isolates: ST195 (n = 3), ST208 (n = 1), ST208/ST1806 (n = 2), ST348 (n = 7) and ST425 (n = 2). Information regarding the whole genome sequenced isolates is presented in Table 3.
Table 3.
Sequence types, β-lactamases and epidemiological data of the whole genome sequenced A. baumannii isolates (n = 15).
3. Discussion
An increase in the number of A. baumannii strains that are resistant to carbapenems is one of the major therapeutic problems worldwide [3,7,8]. Most of these carbapenem-resistant strains exhibit multidrug-resistance (MDR) or extensively drug-resistance (XDR) profiles. According to the European Centre for Disease Prevention and Control (ECDC) data, more than 50% of invasive Acinetobacter spp. strains isolated in Poland in 2015 were defined as carbapenem resistant [9]. Among the 61 carbapenem-nonsusceptible isolates analyzed in our study, all were classified as MDR strains, as defined by Magiorakos et al. [10]. More worryingly, several isolates carrying the genes ISAba1-blaOXA-23-like (10 isolates), blaOXA-24-like (6 isolates) and ISAba1-blaOXA-51-like (1 isolate), showed an XDR resistance profile. An in-depth understanding of the mechanisms causing this widespread resistance is one of the main challenges in the fight to limit the occurrence and spread of carbapenem-resistant strains.
An analysis of the occurrence of blaCHDL genes among the studied isolates revealed the presence of the intrinsic blaOXA-51-like gene in all of the isolates. The majority of isolates also carried one of the following acquired blaCHDL genes: blaOXA-24-like, ISAba1-blaOXA-23-like or ISAba3-blaOXA-58-like. In our study, the ISAba1 sequence was also detected upstream of blaOXA-51-like genes in two isolates in which no acquired genes encoding the remaining CHDL enzymes were found. The presence of an insertion sequence located upstream of blaCHDL leads to an overexpression of this gene, and contributes to the resistance of A. baumannii strains to β-lactams, including carbapenems [5]. So far, the occurrence of the following IS elements ISAba1, ISAba2, ISAba3, ISAba4, ISAba9, ISAba10, IS18 and ISAba825, associated with carbapenem resistance in A. baumannii clinical strains, has been described [11,12,13,14]. ISAba1 is the most common insertion sequence identified in A. baumannii, as well as in other Acinetobacter species, worldwide [12,14]. In clinical A. baumannii strains, the ISAba1 element was found primarily upstream of the blaOXA-23-like genes and, less often, upstream of the blaOXA-51-like and blaOXA-58-like genes. The group of OXA-23-like enzymes is the most commonly acquired resistance CHDL carbapenemase worldwide [3,15,16,17,18,19]. In our study, all of the isolates with the blaOXA-23-like genes possessed ISAba1 upstream. This is the first data concerning the frequency of ISAba1 upstream of blaOXA-23-like genes in A. baumannii isolates from Poland. WGS analysis confirmed the presence of ISAba1-blaOXA-23 gene sequences. So far, in isolates studied worldwide, the presence of other insertion sequences such as ISAba4 and ISAba10 upstream of blaOXA-23-like genes has been described [12]. Interestingly, among the strains examined in this study, more were carrying the blaOXA-24-like gene than the ISAba1-blaOXA-23-like gene (64% versus 23%, respectively). A few studies on A. baumannii strains producing OXA-24 in Poland also indicated a high frequency of occurrence among strains isolated from hospitals. Chmielarczyk et al. reported that 79% of the isolates they studied possessed the blaOXA-24 gene, whereas in the study by Nowak et al., only 49% of the isolates studied possessed the blaOXA-24 gene [20,21]. Reports from Poland appear to be contrary to the data from other countries. OXA-24-like carbapenemases produced by A. baumannii have been found in many European (Bulgaria, France, Portugal, Spain and Sweden) and non-European (Israel, United States and Colombia) countries; however, in most reports, OXA-24 gave way to OXA-23, making the latter more frequently occurring [15,17,22,23,24,25,26]. An insertion sequence has never been found upstream of blaOXA-24-like genes. The genome analysis revealed the presence of the blaOXA-72 gene (from blaOXA-24-like family) in six isolates analyzed in this study. This variant retains its carbapenemase activity. According to Liu et al., imipenem MIC values for isolates producing this enzyme is at least 64 mg/L, which is concordant with our results [17]. A third group of the acquired blaCHDL genes identified among the Polish isolates tested was ISAba3-blaOXA-58-like. The blaOXA-58 gene is currently being reported worldwide, and its increased expression is always associated with IS elements upstream of this gene. In A. baumannii strains isolated from different European countries, the following insertion elements have been found in the blaOXA-58-like gene promoters: ISAba1, ISAba2, ISAba3, ISAba3-like, IS18 and ISAba825 [8,13,27,28]. To date, there are only sparse data on the occurrence of blaOXA-58-like genes in A. baumannii strains isolated in Poland, but none contained information about the presence of IS elements upstream of these genes [15,29]. This is the first time the ISAba3 sequence has been detected upstream of the blaOXA-58-like genes in A. baumannii isolates from Poland, as well as in all six of the OXA-58-producing isolates. Moreover, WGS analysis showed the presence of ISAba3 elements both upstream and downstream of the blaOXA-58 gene in the two isolates tested. It has been reported that often the blaOXA-58-like genes in A. baumannii strains are flanked on two sides with the same or different IS elements [4,7,12].
Overall, using PFGE for genotyping all 61 isolates collected over 2010-2014 did not show any long-term epidemiological incidents caused by any carbapenem resistant A. baumannii strains among patients in the tested hospital. Only a few isolates showed a correlation with a 95% similarity, indicating the transmission of strains between several patients (among two, three, and five patients). The largest number (n = 4) of identical (100% similarity in PFGE) isolates (OXA-24-producing) from different patients was received in 2010. The MLST results revealed that all whole genome sequenced isolates belonged to ST2 (by the Pasteur scheme) which is a highly predominant clone in Poland, as well as in other European countries [15,29,30]. In Poland, carbapenem-resistant strains of A. baumannii belonging to other clones, such as ST1, ST5 and ST193, have rarely been isolated [29]. Analysis of the other four out of seven house-keeping genes in the Oxford scheme typing in relation to the Pasteur scheme allows for the differentiation of strains, especially within ST2, that dominate in Europe. The Oxford scheme revealed diversity among studied isolates and distinguished five sequence types (ST195, ST208, ST208/ST1806, ST348 and ST425). So far, genetic typing of Polish strains of A. baumannii has not been carried out using the Oxford scheme. Among the A. baumannii strains isolated in Europe, the following above-mentioned sequence types have been described: ST195 in Croatia, ST208 in Bulgaria and Greece, ST348 in Germany and ST425 in Greece [31,32,33,34,35]. Carbapenem-resistant A. baumannii ST195 and ST208 are widespread in the world, and are dominant mainly in the Far East, e.g., in China [36,37,38,39,40]. Furthermore, the ST208/ST1806 strains have been isolated in South Korea [41].
In our study, the majority of isolates carrying the blaOXA-24-like and ISAba1-blaOXA-23-like genes demonstrated the highest carbapenem MIC values, and were the only isolates that presented a positive CarbAcineto NP test result. According to Dortet et al., this test is capable of detecting the production of various CHDLs, such as OXA-23, OXA-24, OXA-58 and OXA-143 groups, by A. baumannii strains with a high level of resistance to carbapenems, i.e., MICs 8 - >32 mg/L [42]. Positive carbapenemase activity results in the CarbAcineto NP Test were obtained for the majority of the ISAba1-blaOXA-23-like (n = 12/14) and blaOXA-24-like-carrying (n = 34/39) isolates. In this study, the results of the CarbAcineto NP test for several blaOXA-23-like- and blaOXA-24-like-carrying isolates (2/14 and 5/39, respectively), as well as for all isolates with the ISAba3-blaOXA-58-like (n = 6) and ISAba1-blaOXA-51-like genes (n = 2), were uninterpretable. Even though the CarbAcineto NP test is able to detect A. baumannii CHDL-producing isolates, it is still possible that the results would be ambiguous. Literacka et al., in their study, demonstrated that for one OXA-24-like, three OXA-23-like and two OXA-58-like-producing A. baumannii isolates, the results of the CarbAcineto NP Test were uninterpretable [43]. In such cases, expression levels of the genes encoding CHDL enzymes may be low or inhibited, which would suggest that some other resistance mechanism affecting carbapenem activity dominates in those isolates, giving high carbapenem MIC values. Perhaps various resistance mechanisms interact simultaneously in these few isolates, such as efflux pumps, changes in outer membrane proteins and, to a lesser extent, carbapenemases.
4. Materials and Methods
4.1. Bacterial Strains
Here, 61 non-repetitive imipenem-nonsusceptible A. baumannii isolates were collected in 2009 (n = 2), 2010 (n = 9), 2011 (n = 4), 2012 (n = 18), 2013 (n = 27) and 2014 (n = 1) from patients hospitalized in one tertiary hospital in Warsaw, Poland. The isolates were recovered from various clinical specimens, including respiratory tract samples (15, 24.6%), wound swabs (15, 24.6%), urine (15, 24.6%), blood (3, 4.9%) and other specimens (13, 21.3%). The isolates were initially identified using the VITEK 2 system (bioMérieux, Mercy l’Etoile, France) and were assigned to the Acinetobacter calcoaceticus–A. baumannii complex. Identification of A. baumannii was confirmed by gyrB multiplex PCR, as previously described [44,45].
4.2. Antimicrobial Susceptibility Testing
The isolates were first determined to be imipenem-nonsusceptible using a disk diffusion method utilizing an imipenem disk (10 μg, Becton, Dickinson and Company, Franklin Lakes, NJ, USA), according to Clinical and Laboratory Standards Institute (CLSI) recommendations (2018) [46]. Minimal inhibitory concentrations (MICs) of piperacillin, piperacillin/tazobactam, ceftazidime, cefepime, imipenem, meropenem, gentamicin, tobramycin, ciprofloxacin, levofloxacin, colistin and trimethoprim/sulfamethoxazole were determined using the VITEK 2 system (bioMérieux, Mercy l’Etoile, France) and relevant antimicrobial susceptibility testing cards.
The MICs of imipenem and meropenem were also determined with the microdilution method in a Mueller Hinton II (MH II) broth medium (Becton, Dickinson and Company, Franklin Lakes, NJ, USA), according to CLSI guidelines (2012, 2018) [46,47]. Escherichia coli ATCC 25,922 was used as the reference strain for quality control.
4.3. Phenotypic Detection of Carbapenemase Production
4.3.1. Detection of Metallo-β-Lactamases (MBL)
The MBL mechanism of resistance was detected using the double disc diffusion method (DDST) [48]. Disks of 10 μg of imipenem (Becton, Dickinson and Company, Franklin Lakes, NJ, USA) and 30 μg of ceftazidime (Becton, Dickinson and Company, Franklin Lakes, NJ, USA) were placed at a distance of 20 mm (center to center) from a disk containing 0.01 mL of 0.5 mM EDTA solution. The presence of an extended inhibition zone of antibiotics towards the disc with EDTA was interpreted as a positive result for metallo-β-lactamase-producing strains screening.
4.3.2. CarbAcineto NP Test
The CarbAcineto NP test was adapted from the updated version of the Carba NP test, used for the detection of carbapenemase-producing Enterobacteriaceae and Pseudomonas spp. The test was performed as described by Dortet et al. [42].
4.4. Molecular Detection of blaOXA-like Genes and ISs Upstream of blaCHDL Genes
Genomic DNA was extracted using the Genomic Mini isolation kit (A&A Biotechnology, Gdynia, Poland), according to manufacturer’s instructions. Detection of the blaCHDL genes was performed using (1) duplex PCR for blaOXA-51-like and blaOXA-23-like, (2) duplex PCR for blaOXA-24-like and blaOXA-58-like and (3) singleplex for blaOXA-143-like, and (4) singleplex was used for detection of the insertion sequences (ISs) upstream of blaOXA-51-like, blaOXA-23-like, blaOXA-24-like and blaOXA-58-like. The primers used are listed in Table 4. The PCR reactions were performed using Hypernova polymerase (Blirt S.A., Gdańsk, Poland) with the following amplification parameters: (1) all duplex PCRs were performed at 95 °C for 4 min, followed by 25 cycles of 30 s at 95 °C, 30 s at 55 °C, 50 s at 72 °C and a final extension of 3 min at 72 °C, and (2) singleplex PCR for blaOXA-143-like was performed at 95 °C for 4 min, followed by 25 cycles of 30 s at 95 °C, 30 s at 54 °C, 45 s at 72 °C and a final extension of 3 min at 72 °C. In addition, (3) singleplex PCRs for the ISs upstream each blaCHDL were performed at 95 °C for 4 min, followed by 25 cycles of 30 s at 95 °C, 30 s at 56 °C, 75 s at 72 °C and a final extension of 3 min at 72 °C.
Table 4.
Primers used for amplification.
4.5. Pulsed Field Gel Electrophoresis (PFGE)
All 61 A. baumannii isolates were typed by PFGE according to a previous protocol [50] with modifications. An overnight culture of bacteria was suspended in 150 μL of a cell suspension buffer and was mixed with 20 μL of 20 mg/mL Proteinase K (Promega GmbH, Walldorf, Germany) and 170 μL of 1.5% low melting agarose (SeaKem Gold Agarose (Lonza, Basel, Switzerland)) and distributed in a plug mold. The genomic DNA in agarose plugs was lysed in 2.5 mL of cell lysis solution supplemented with 20 μL of 20 mg/mL Proteinase K (Promega GmbH, Walldorf, Germany) and 7.5 μL of 10 mg/mL RNase (Sigma, St. Louis, MO, USA), and was washed and digested with 20 U ApaI restriction enzyme (ABO, Gdańsk, Poland). Electrophoresis was performed using the CHEF DR II system (Bio-Rad, Hercules, CA, USA). The migration conditions were as follows: switch angle 120, voltage 6 V/cm, temperature 14 °C and a two-blocks program with a total run time of 22 h (first-block 1-8 s for 15 h, and second-block 5–20 s for an additional 7 h). The genomic DNA of Salmonella serotype Braenderup strain (H9812) digested with XbaI (ABO, Gdańsk, Poland) and Lambda-DNA Ladder PFG Marker (New England BioLabs, Ipswich, MA, USA) were used as the DNA molecular-weight markers [51]. The PFGE patterns were analyzed using GelCompar II software (Applied Maths, Sint-Martens-Latem, Belgium) with the Dice coefficient and clustering by unweighted pair group method with arithmetic mean (UPGMA) with 1% tolerance. The isolates were clustered in the PFGE pulsotypes (PTs) in accordance with the recommendations of Tenover et al. [52].
4.6. Whole Genome Sequencing (WGS) and Multi-Locus Sequence Typing (MLST)
Several A. baumannii isolate genomes were whole genome sequenced. This was performed using Public Health England - Genomic Services and Development Unit (PHE-GSDU) on the HiSeq 2500 System (Illumina, CambridgeZ, UK) with paired end read lengths of 150 bp. A minimum of 150 Mb of Q30 quality data were obtained for each isolate. FastQ files were quality trimmed using Trimmomatic 0.32 [53]. SPAdes 3.1.1 was used to produce draft chromosomal assemblies, and contigs less than 500 bp were filtered out [54]. All of the above sequencing analyses were performed using PHE Galaxy [55]. Using the WGS data, the sequence types (STs) were determined using both the Pasteur and Oxford MLST databases [56]. In the case of the Pasteur MLST scheme, the cpn60, fusA, gltA, pyrG, recA, rplB and rpoB gene sequences were analyzed, while the Oxford MLST scheme analysis included the gltA, gyrB, gdhB, recA, cpn60, gpi and rpoD gene sequences.
5. Conclusions
From the A. baumannii clinical strains isolated in Poland, the dominating carbapenem resistance mechanism was CHDL enzymes, mainly those from the OXA-24-like family. The presence of insertion sequences flanking blaOXA-58-like (ISAba3) and blaOXA-23-like (ISAba1) genes, which enhance the gene expression, was demonstrated. All tested isolates belong to sequence type ST2 (of the Pasteur MLST scheme) from the global IC2 clone. The Oxford MLST scheme allowed for the differentiation of the sequence types of these isolates.
Supplementary Materials
The following are available online at https://www.mdpi.com/1422-0067/22/5/2486/s1, Table S1: Occurrence of the acquired blaCHDL genes, the β-lactam susceptibility profiles and the carbapenemase activity data of whole genome sequenced A. baumannii isolates (n = 15).
Author Contributions
Conceived and designed the experiments, A.S. and A.E.L.; performed the experiments, A.S. and M.E.W.; analyzed the data, A.S., M.E.W., S.T. and A.E.L.; contributed reagents/materials/analysis tools, A.S. and M.E.W.; wrote the manuscript draft, A.S., M.E.W., S.T. and A.E.L.; final approval of the version to be published, A.S., M.E.W., S.T. and A.E.L. All authors have read and agreed to the published version of the manuscript.
Funding
This study was partially supported by grants from the Medical University of Warsaw (grants no. FW15/PM2/16 and FW15/PM1/18) and by the Foundation for the Development of Diagnostics and Therapy, Warsaw, Poland (REGON: 006220910, NIP: 5262173856 and KRS: 0000195643).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The whole genome datasets of the 15 strains are available at the NCBI BioProject repository (SubmissionID: SUB9082120, BioProject ID: PRJNA701882).
Acknowledgments
Research was carried out with the use of the CePT infrastructure financed by the European Union through the European Regional Development Fund as part of the Operational Program “Innovative Economy” for 2007–2013.
Conflicts of Interest
The authors report no conflict of interest in this work.
References
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Maragakis, L.L.; Perl, T.M. Acinetobacter baumannii: Epidemiology, antimicrobial resistance, and treatment options. Clin. Infect. Dis. 2008, 46, 1254–1263. [Google Scholar] [CrossRef] [PubMed]
- Potron, A.; Poirel, L.; Nordmann, P. Emerging broad-spectrum resistance in Pseudomonas aeruginosa and Acinetobacter baumannii: Mechanisms and epidemiology. Int. J. Antimicrob. Agents 2015, 45, 568–585. [Google Scholar] [CrossRef]
- Poirel, L.; Naas, T.; Nordmann, P. Diversity, epidemiology, and genetics of class D beta-lactamases. Antimicrob. Agents Chemother. 2010, 54, 24–38. [Google Scholar] [CrossRef]
- El-Shazly, S.; Dashti, A.; Vali, L.; Bolaris, M.; Ibrahim, A.S. Molecular epidemiology and characterization of multiple-drug resistant (MDR) clinical isolates of Acinetobacter baumannii. Int. J. Infect. Dis. 2015. [Google Scholar] [CrossRef]
- Evans, B.A.; Amyes, S.G. OXA beta-lactamases. Clin. Microbiol. Rev. 2014, 27, 241–263. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Nordmann, P. Carbapenem resistance in Acinetobacter baumannii: Mechanisms and epidemiology. Clin. Microbiol. Infect. 2006, 12, 826–836. [Google Scholar] [CrossRef]
- Poirel, L.; Nordmann, P. Genetic structures at the origin of acquisition and expression of the carbapenem-hydrolyzing oxacillinase gene bla (OXA-58) in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2006, 50, 1442–1448. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Rapid Risk Assessment: Carbapenem-Resistant Acinetobacter Baumannii in Healthcare Settings—8 December 2016. Available online: https://www.ecdc.europa.eu/en/publications-data/rapid-risk-assessment-carbapenem-resistant-acinetobacter-baumannii-healthcare (accessed on 25 February 2021).
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, S.; Poirel, L.; Papa, A.; Koulourida, V.; Nordmann, P. Overexpression of the naturally occurring blaOXA-51 gene in Acinetobacter baumannii mediated by novel insertion sequence ISAba9. Antimicrob. Agents Chemother. 2009, 53, 4045–4047. [Google Scholar] [CrossRef]
- Pagano, M.; Martins, A.F.; Barth, A.L. Mobile genetic elements related to carbapenem resistance in Acinetobacter baumannii. Braz. J. Microbiol. 2016, 47, 785–792. [Google Scholar] [CrossRef]
- Karah, N.; Giske, C.G.; Sundsfjord, A.; Samuelsen, O. A diversity of OXA-carbapenemases and class 1 integrons among carbapenem-resistant Acinetobacter baumannii clinical isolates from Sweden belonging to different international clonal lineages. Microb. Drug Resist. 2011, 17, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.D.; Bishop, B.; Wright, M.S. Quantitative assessment of insertion sequence impact on bacterial genome architecture. Microb. Genom. 2016, 2, e000062. [Google Scholar] [CrossRef]
- Castanheira, M.; Costello, S.E.; Woosley, L.N.; Deshpande, L.M.; Davies, T.A.; Jones, R.N. Evaluation of clonality and carbapenem resistance mechanisms among Acinetobacter baumannii-Acinetobacter calcoaceticus complex and Enterobacteriaceae isolates collected in European and Mediterranean countries and detection of two novel beta-lactamases, GES-22 and VIM-35. Antimicrob. Agents Chemother. 2014, 58, 7358–7366. [Google Scholar] [CrossRef] [PubMed]
- Mugnier, P.D.; Poirel, L.; Naas, T.; Nordmann, P. Worldwide dissemination of the bla (OXA-23) carbapenemase gene of Acinetobacter baumannii. Emerg. Infect. Dis. 2010, 16, 35–40. [Google Scholar] [CrossRef]
- Liu, C.P.; Lu, H.P.; Luor, T. Clonal relationship and the association of the ST218 strain harboring blaOXA-72 gene to mortality in carbapenem-resistant Acinetobacter baumannii bacteremia. J. Microbiol. Immunol. Infect. 2019, 52, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Hujer, A.M.; Hujer, K.M.; Leonard, D.A.; Powers, R.A.; Wallar, B.J.; Mack, A.R.; Taracila, M.A.; Rather, P.N.; Higgins, P.G.; Prati, F.; et al. A comprehensive and contemporary “snapshot” of beta-lactamases in carbapenem resistant Acinetobacter baumannii. Diagn. Microbiol. Infect. Dis. 2021, 99, 115242. [Google Scholar] [CrossRef] [PubMed]
- Mosqueda, N.; Espinal, P.; Cosgaya, C.; Viota, S.; Plasensia, V.; Alvarez-Lerma, F.; Montero, M.; Gómez, J.; Horcajada, J.P.; Vila, J.; et al. Globally expanding carbapenemase finally appears in Spain: Nosocomial outbreak of Acinetobacter baumannii producing plasmid-encoded OXA-23 in Barcelona, Spain. Antimicrob. Agents Chemother. 2013, 57, 5155–5157. [Google Scholar] [CrossRef] [PubMed]
- Chmielarczyk, A.; Pilarczyk-Zurek, M.; Kaminska, W.; Pobiega, M.; Romaniszyn, D.; Ziolkowski, G.; Wojkowska-Mach, J.; Bulanda, M. Molecular epidemiology and drug resistance of Acinetobacter baumannii isolated from hospitals in southern Poland: ICU as a risk factor for XDR strains. Microb. Drug Resist. 2016, 22, 328–335. [Google Scholar] [CrossRef]
- Nowak, P.; Paluchowska, P.; Budak, A. Distribution of bla (OXA) genes among carbapenem-resistant Acinetobacter baumannii nosocomial strains in Poland. New Microbiol. 2012, 35, 317–325. [Google Scholar] [PubMed]
- Todorova, B.; Velinov, T.; Ivanov, I.; Dobreva, E.; Kantardjiev, T. First detection of OXA-24 carbapenemase-producing Acinetobacter baumannii isolates in Bulgaria. World J. Microbiol. Biotechnol. 2014, 30, 1427–1430. [Google Scholar] [CrossRef]
- Heritier, C.; Poirel, L.; Lambert, T.; Nordmann, P. Contribution of acquired carbapenem-hydrolyzing oxacillinases to carbapenem resistance in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2005, 49, 3198–3202. [Google Scholar] [CrossRef]
- Afzal-Shah, M.; Woodford, N.; Livermore, D.M. Characterization of OXA-25, OXA-26, and OXA-27, molecular class D beta-lactamases associated with carbapenem resistance in clinical isolates of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2001, 45, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.F.; Lan, C.Y. Antimicrobial resistance in Acinetobacter baumannii: From bench to bedside. World J. Clin. Cases 2014, 2, 787–814. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, S.Y.; Cayo, R.; Gales, A.C.; Leal, A.L.; Saavedra, C.H. Early dissemination of OXA-72-producing Acinetobacter baumannii strain in Colombia: A case report. Braz. J. Infect. Dis. 2014, 18, 678–680. [Google Scholar] [CrossRef] [PubMed]
- Villalon, P.; Valdezate, S.; Medina-Pascual, M.J.; Carrasco, G.; Vindel, A.; Saez-Nieto, J.A. Epidemiology of the Acinetobacter-derived cephalosporinase, carbapenem-hydrolysing oxacillinase and metallo-beta-lactamase genes, and of common insertion sequences, in epidemic clones of Acinetobacter baumannii from Spain. J. Antimicrob. Chemother. 2013, 68, 550–553. [Google Scholar] [CrossRef] [PubMed]
- Lopes, B.S.; Evans, B.A.; Amyes, S.G. Disruption of the blaOXA-51-like gene by ISAba16 and activation of the blaOXA-58 gene leading to carbapenem resistance in Acinetobacter baumannii Ab244. J. Antimicrob. Chemother. 2012, 67, 59–63. [Google Scholar] [CrossRef]
- Izdebski, R.; Fiett, J.; Hryniewicz, W.; Gniadkowski, M. Molecular analysis of Acinetobacter baumannii isolates from invasive infections in 2009 in Poland. J. Clin. Microbiol. 2012, 50, 3813–3815. [Google Scholar] [CrossRef] [PubMed]
- Chmielarczyk, A.; Pobiega, M.; Romaniszyn, D.; Wojkowska-Mach, J. Multi-locus sequence typing (MLST) of non-fermentative Gram-negative bacilli isolated from bloodstream infections in southern Poland. Folia Microbiol. 2018, 63, 191–196. [Google Scholar] [CrossRef]
- Palmieri, M.; D’Andrea, M.M.; Pelegrin, A.C.; Perrot, N.; Mirande, C.; Blanc, B.; Legakis, N.; Goossens, H.; Rossolini, G.M.; van Belkum, A. Abundance of colistin-resistant, OXA-23- and ArmA-producing Acinetobacter baumannii belonging to international clone 2 in Greece. Front. Microbiol. 2020, 11, 668. [Google Scholar] [CrossRef] [PubMed]
- Wareth, G.; Brandt, C.; Sprague, L.D.; Neubauer, H.; Pletz, M.W. Spatio-temporal distribution of Acinetobacter baumannii in Germany—a comprehensive systematic review of studies on resistance development in humans (2000–2018). Microorganisms 2020, 8, 375. [Google Scholar] [CrossRef]
- Pfeifer, Y.; Hunfeld, K.P.; Borgmann, S.; Maneg, D.; Blobner, W.; Werner, G.; Higgins, P.G. Carbapenem-resistant Acinetobacter baumannii ST78 with OXA-72 carbapenemase and ESBL gene blaCTX-M-115. J. Antimicrob. Chemother. 2016, 71, 1426–1428. [Google Scholar] [CrossRef]
- Pfeifer, Y.; Trifonova, A.; Pietsch, M.; Brunner, M.; Todorova, I.; Gergova, I.; Wilharm, G.; Werner, G.; Savov, E. Clonal transmission of Gram-negative bacteria with carbapenemases NDM-1, VIM-1, and OXA-23/72 in a Bulgarian hospital. Microb. Drug Resist. 2017, 23, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Ladavac, R.; Bedenic, B.; Vranic-Ladavac, M.; Barisic, N.; Karcic, N.; Pompe, K.; Ferencic, A.; Stojanovic, A.; Seifert, H.; Katic, S.; et al. Emergence of different Acinetobacter baumannii clones in a Croatian hospital and correlation with antibiotic susceptibility. J. Glob. Antimicrob. Resist. 2017, 10, 213–218. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chen, Q.; Zhou, J.W.; Fan, J.Z.; Wu, S.H.; Xu, L.H.; Jiang, Y.; Ruan, Z.; Yu, Y.S.; Yu, D.J.; Wang, X.J. Simultaneous emergence and rapid spread of three OXA-23 producing Acinetobacter baumannii ST208 strains in intensive care units confirmed by whole genome sequencing. Infect. Genet. Evol. 2018, 58, 243–250. [Google Scholar] [CrossRef]
- Leung, E.C.; Leung, P.H.; Lai, R.W. Emergence of carbapenem-resistant Acinetobacter baumannii ST195 harboring blaOXA-23 isolated from bacteremia in Hong Kong. Microb. Drug Resist. 2019, 25, 1199–1203. [Google Scholar] [CrossRef]
- Jia, H.; Chen, Y.; Wang, J.; Xie, X.; Ruan, Z. Emerging challenges of whole-genome-sequencing-powered epidemiological surveillance of globally distributed clonal groups of bacterial infections, giving Acinetobacter baumannii ST195 as an example. Int. J. Med. Microbiol. 2019, 309, 151339. [Google Scholar] [CrossRef]
- Fang, Y.; Quan, J.; Hua, X.; Feng, Y.; Li, X.; Wang, J.; Ruan, Z.; Shang, S.; Yu, Y. Complete genome sequence of Acinetobacter baumannii XH386 (ST208), a multi-drug resistant bacteria isolated from pediatric hospital in China. Genom. Data 2016, 7, 269–274. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, X.; Du, Z.; Huang, W.; Zhang, X.; Zhou, Y. Outbreak of multidrug-resistant Acinetobacter baumannii ST208 producing OXA-23-Like carbapenemase in a children’s hospital in Shanghai, China. Microb. Drug Resist. 2020. [Google Scholar] [CrossRef]
- Kim, M.H.; Jeong, H.; Sim, Y.M.; Lee, S.; Yong, D.; Ryu, C.M.; Choi, J.Y. Using comparative genomics to understand molecular features of carbapenem-resistant Acinetobacter baumannii from South Korea causing invasive infections and their clinical implications. PLoS ONE 2020, 15, e0229416. [Google Scholar] [CrossRef]
- Dortet, L.; Poirel, L.; Errera, C.; Nordmann, P. CarbAcineto NP test for rapid detection of carbapenemase-producing Acinetobacter. spp. J. Clin. Microbiol. 2014, 52, 2359–2364. [Google Scholar] [CrossRef]
- Literacka, E.; Herda, M.; Baraniak, A.; Zabicka, D.; Hryniewicz, W.; Skoczynska, A.; Gniadkowski, M. Evaluation of the Carba NP test for carbapenemase detection in Enterobacteriaceae, Pseudomonas. spp. and Acinetobacter. spp., and its practical use in the routine work of a national reference laboratory for susceptibility testing. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2281–2287. [Google Scholar] [CrossRef]
- Higgins, P.G.; Wisplinghoff, H.; Krut, O.; Seifert, H. A PCR-based method to differentiate between Acinetobacter baumannii and Acinetobacter genomic species 13TU. Clin. Microbiol. Infect. 2007, 13, 1199–1201. [Google Scholar] [CrossRef]
- Higgins, P.G.; Lehmann, M.; Wisplinghoff, H.; Seifert, H. gyrB multiplex PCR to differentiate between Acinetobacter calcoaceticus and Acinetobacter genomic species 3. J. Clin. Microbiol. 2010, 48, 4592–4594. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; supplement M100; CLSI: Wayne, PA, USA, 2018. [Google Scholar]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Test for Bacteria that Grow Aerobically, 9th ed.; approved standard Document M07-A9; CLSI: Wayne, PA, USA, 2012. [Google Scholar]
- Lee, K.; Lim, Y.S.; Yong, D.; Yum, J.H.; Chong, Y. Evaluation of the Hodge test and the imipenem-EDTA double-disk synergy test for differentiating metallo-beta-lactamase-producing isolates of Pseudomonas spp. and Acinetobacter spp. J. Clin. Microbiol. 2003, 41, 4623–4629. [Google Scholar] [CrossRef]
- Woodford, N.; Ellington, M.J.; Coelho, J.M.; Turton, J.F.; Ward, M.E.; Brown, S.; Amyes, S.G.; Livermore, D.M. Multiplex PCR for genes encoding prevalent OXA carbapenemases in Acinetobacter spp. Int. J. Antimicrob. Agents 2006, 27, 351–353. [Google Scholar] [CrossRef]
- Seifert, H.; Dolzani, L.; Bressan, R.; van der Reijden, T.; van Strijen, B.; Stefanik, D.; Heersma, H.; Dijkshoorn, L. Standardization and interlaboratory reproducibility assessment of pulsed-field gel electrophoresis-generated fingerprints of Acinetobacter baumannii. J. Clin. Microbiol. 2005, 43, 4328–4335. [Google Scholar] [CrossRef]
- Hunter, S.B.; Vauterin, P.; Lambert-Fair, M.A.; Van Duyne, M.S.; Kubota, K.; Graves, L.; Wrigley, D.; Barrett, T.; Ribot, E. Establishment of a universal size standard strain for use with the PulseNet standardized pulsed-field gel electrophoresis protocols: Converting the national databases to the new size standard. J. Clin. Microbiol. 2005, 43, 1045–1050. [Google Scholar] [CrossRef]
- Tenover, F.C.; Arbeit, R.D.; Goering, R.V.; Mickelsen, P.A.; Murray, B.E.; Persing, D.H.; Swaminathan, B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: Criteria for bacterial strain typing. J. Clin. Microbiol. 1995, 33, 2233–2239. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Afgan, E.; Baker, D.; van den Beek, M.; Blankenberg, D.; Bouvier, D.; Cech, M.; Chilton, J.; Clements, D.; Coraor, N.; Eberhard, C.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2016 update. Nucleic Acids Res. 2016, 44, W3–W10. [Google Scholar] [CrossRef] [PubMed]
- Public Databases for Molecular Typing and Microbial Genome Diversity. Available online: https://pubmlst.org/organisms/acinetobacter-baumannii (accessed on 25 February 2021).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).