
Surface Topography of Titanium Affects Their Osteogenic Potential through DNA Methylation



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
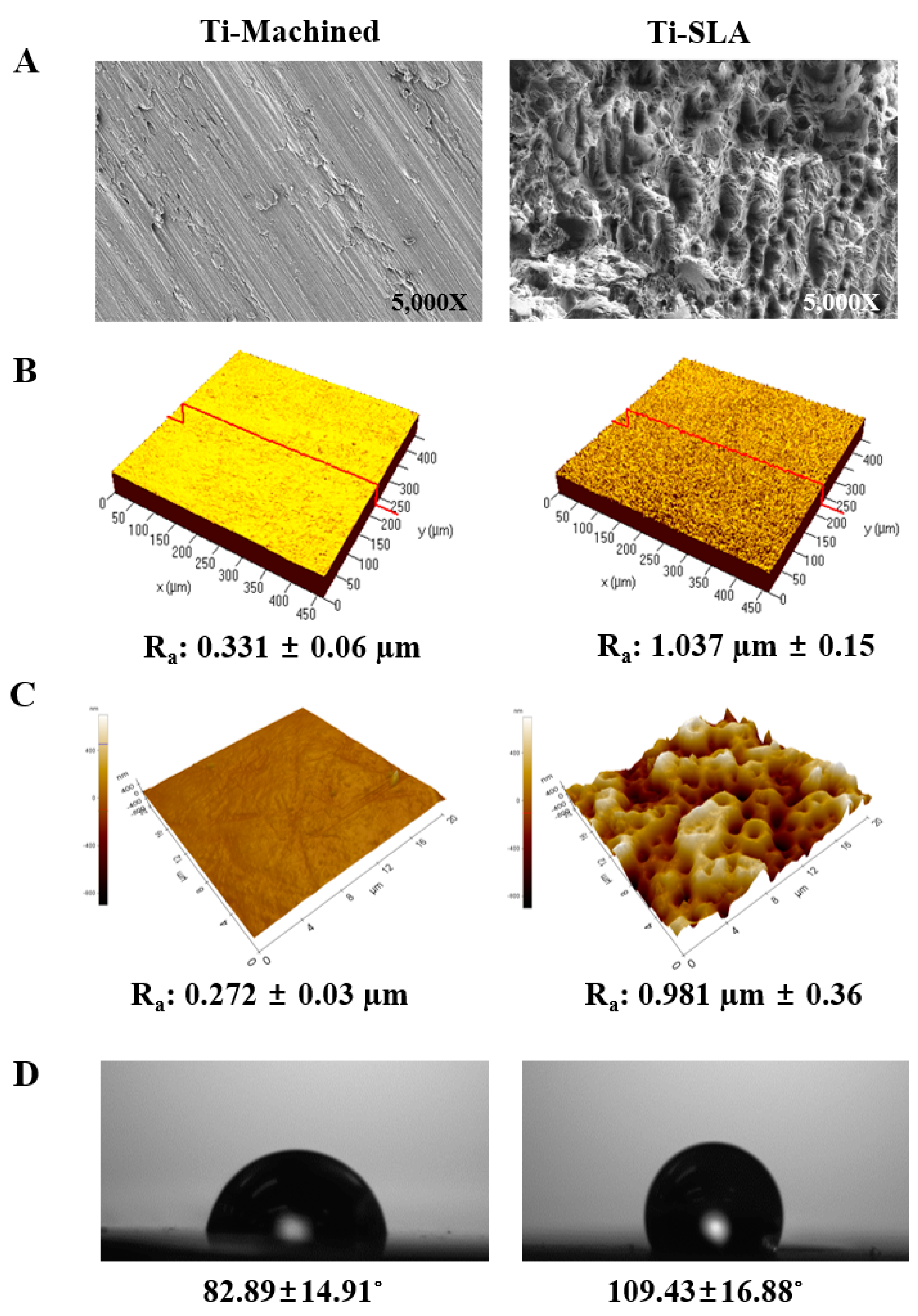
2.1. Characterization and Comparison of Ti Surfaces
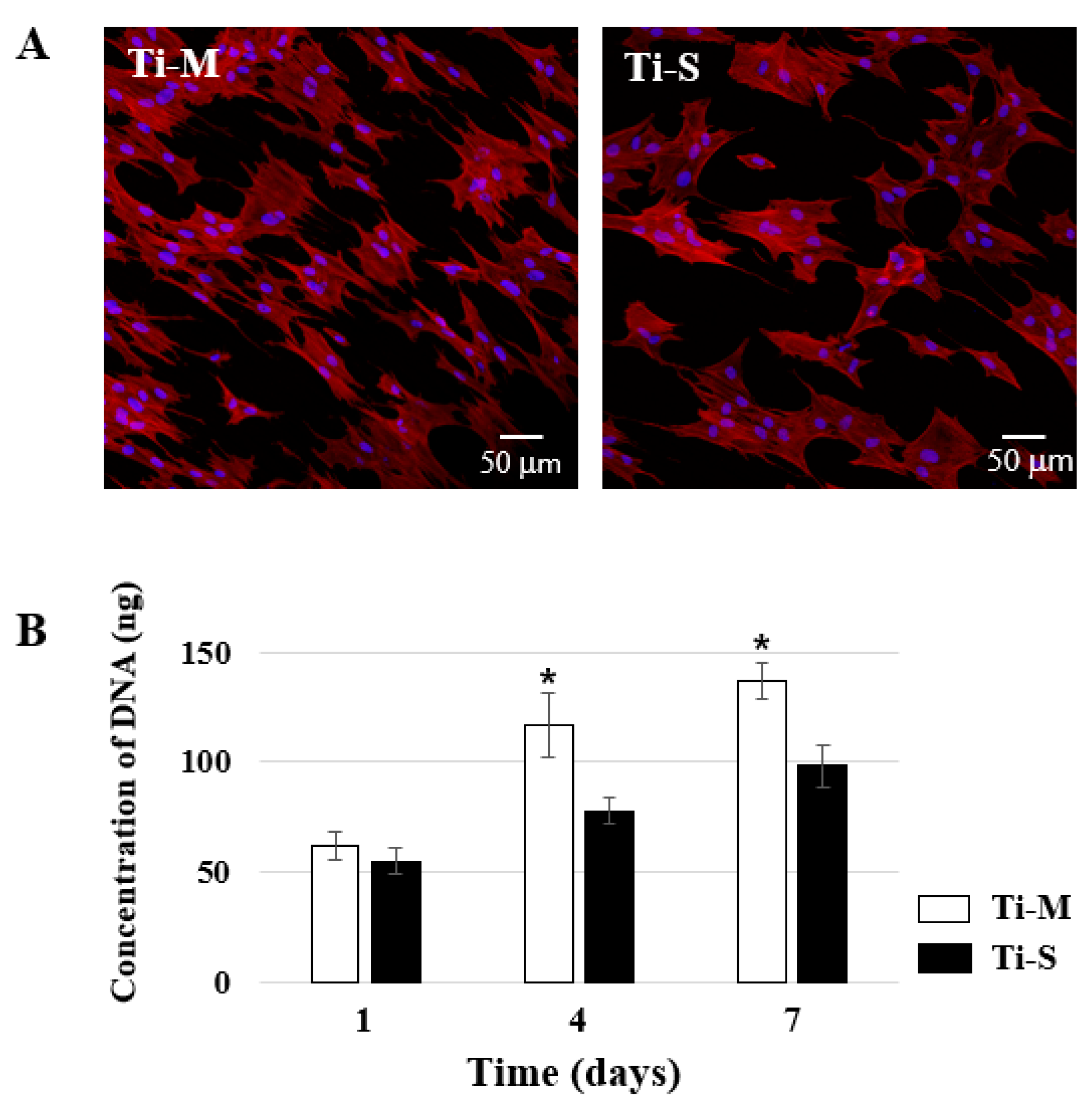
2.2. Cell Morphology
2.3. Cellular Proliferation
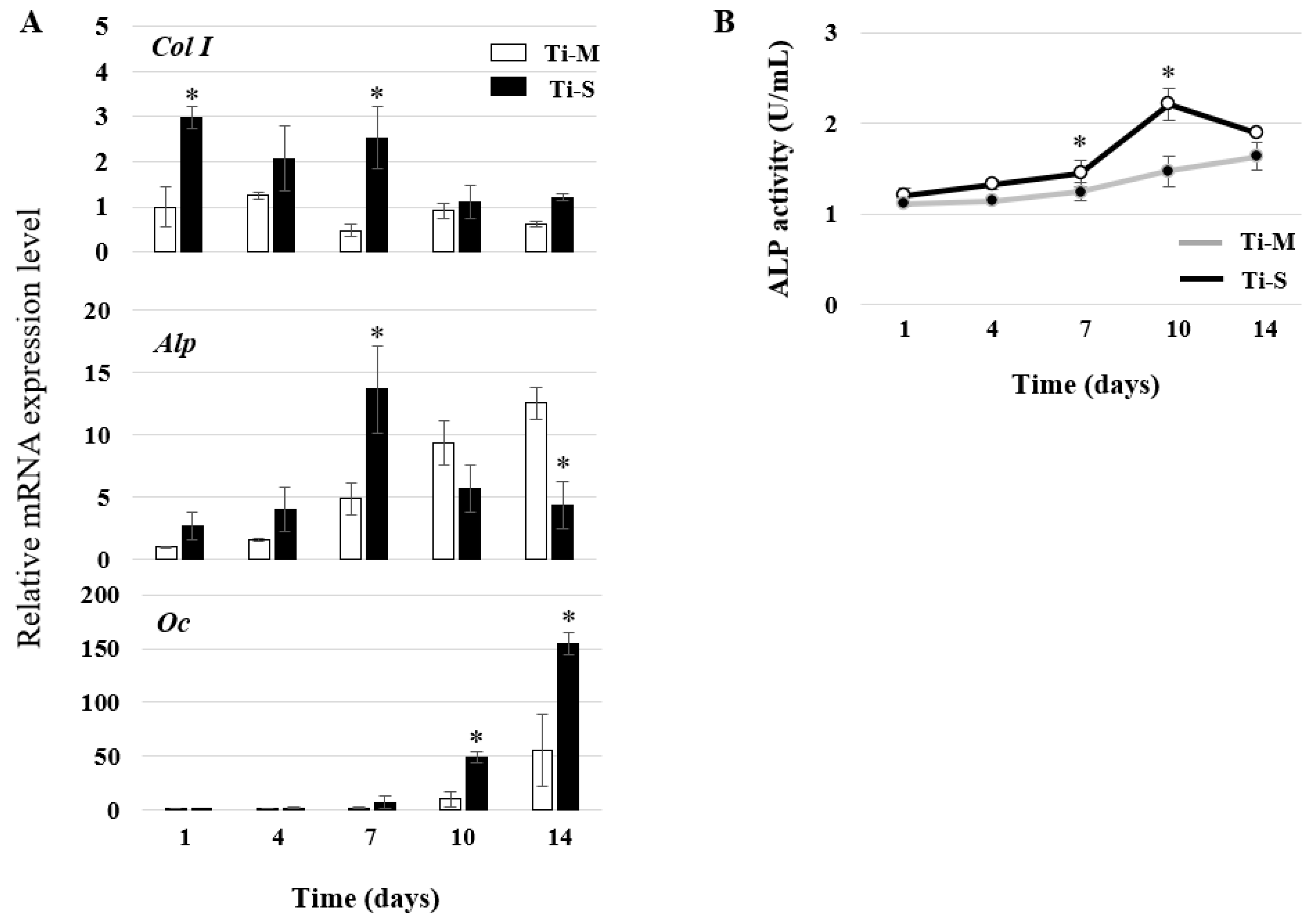
2.4. Osteoblast Differentiation
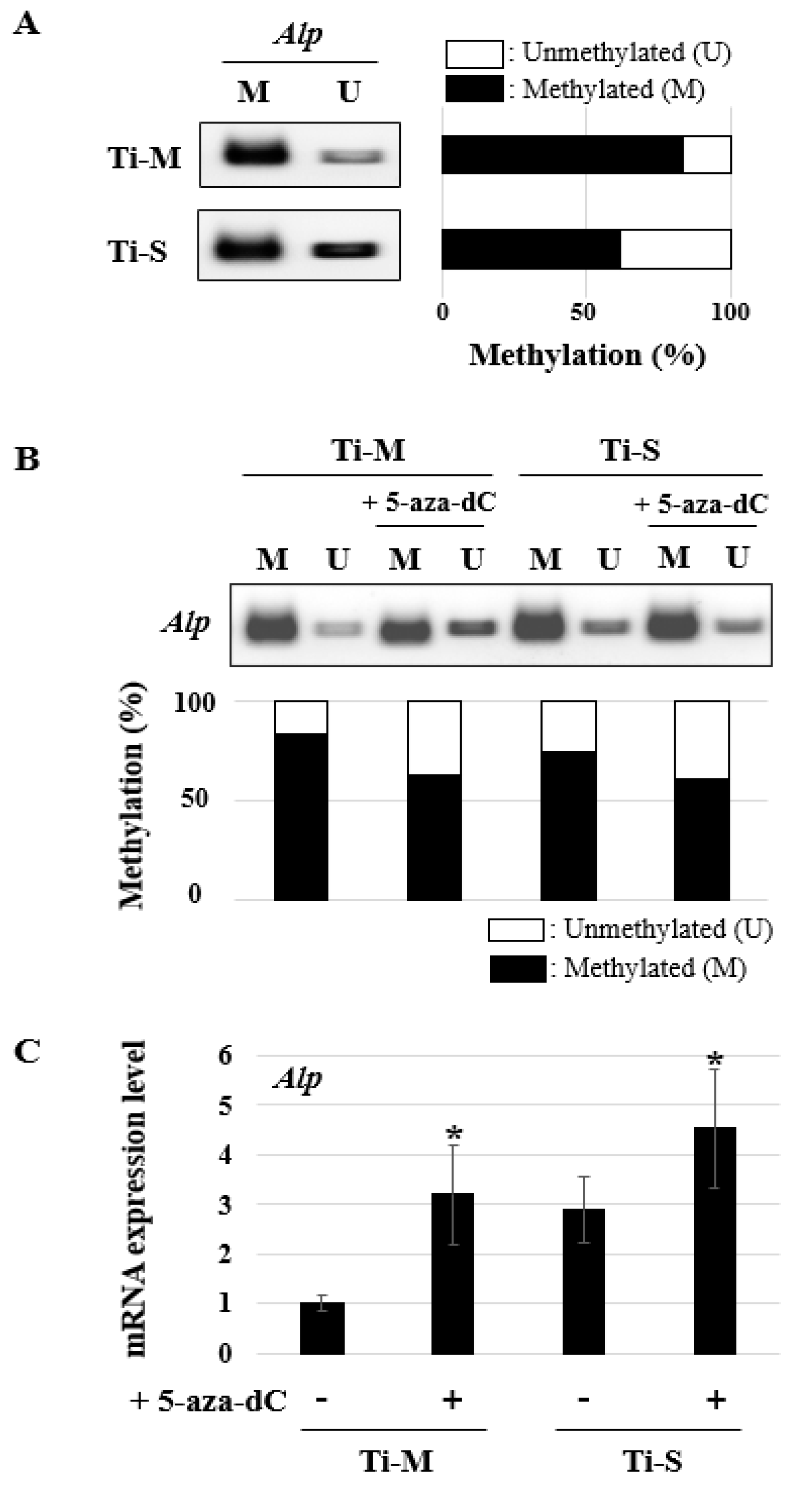
2.5. DNA Methylation Pattern Analysis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Surface Analysis
4.3. Cell Culture
4.4. Cell Morphological Observation
4.5. Cell Proliferation Assay
4.6. Reverse-Transcription PCR and Quantitative Real-Time PCR
4.7. ALP Activity Assay
4.8. Methylation Specific PCR (MSP)
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Branemark, P.I. Osseointegration and its experimental background. J. Prosthet. Dent. 1983, 50, 399–410. [Google Scholar] [CrossRef]
- Albrektsson, T.; Wennerberg, A. On osseointegration in relation to implant surfaces. Clin. Implant. Dent. Relat. Res. 2019, 21 (Suppl. 1), 4–7. [Google Scholar] [CrossRef]
- Novaes, A.B., Jr.; de Souza, S.L.; de Barros, R.R.; Pereira, K.K.; Iezzi, G.; Piattelli, A. Influence of implant surfaces on osseointegration. Braz. Dent. J. 2010, 21, 471–481. [Google Scholar] [CrossRef]
- Kunzler, T.P.; Huwiler, C.; Drobek, T.; Voros, J.; Spencer, N.D. Systematic study of osteoblast response to nanotopography by means of nanoparticle-density gradients. Biomaterials 2007, 28, 5000–5006. [Google Scholar] [CrossRef] [PubMed]
- Wennerberg, A.; Albrektsson, T. Effects of titanium surface topography on bone integration: A systematic review. Clin. Oral Implants Res. 2009, 20 (Suppl. 4), 172–184. [Google Scholar] [CrossRef]
- Albrektsson, T.; Wennerberg, A. Oral implant surfaces: Part 1—Review focusing on topographic and chemical properties of different surfaces and in vivo responses to them. Int. J. Prosthodont. 2004, 17, 536–543. [Google Scholar]
- Schneider, G.B.; Perinpanayagam, H.; Clegg, M.; Zaharias, R.; Seabold, D.; Keller, J.; Stanford, C. Implant surface roughness affects osteoblast gene expression. J. Dent. Res. 2003, 82, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Masaki, C.; Schneider, G.B.; Zaharias, R.; Seabold, D.; Stanford, C. Effects of implant surface microtopography on osteoblast gene expression. Clin. Oral Implants Res. 2005, 16, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Roccuzzo, M.; Bonino, F.; Bonino, L.; Dalmasso, P. Surgical therapy of peri-implantitis lesions by means of a bovine-derived xenograft: Comparative results of a prospective study on two different implant surfaces. J. Clin. Periodontol. 2011, 38, 738–745. [Google Scholar] [CrossRef]
- Esposito, M.; Hirsch, J.M.; Lekholm, U.; Thomsen, P. Failure patterns of four osseointegrated oral implant systems. J. Mater. Sci. Mater. Med. 1997, 8, 843–847. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, A.D.; Allis, C.D.; Bernstein, E. Epigenetics: A landscape takes shape. Cell 2007, 128, 635–638. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.D.; Kim, P.J.; Kim, H.G.; Seol, Y.J.; Lee, Y.M.; Ku, Y.; Rhyu, I.C.; Ryoo, H.M. Transcriptomics and methylomics in chronic periodontitis with tobacco use: A pilot study. Clin. Epigenet. 2017, 9, 81. [Google Scholar] [CrossRef]
- Jones, P.A. Functions of DNA methylation: Islands, start sites, gene bodies and beyond. Nat. Rev. Genet. 2012, 13, 484–492. [Google Scholar] [CrossRef]
- Cho, Y.D.; Bae, H.S.; Lee, D.S.; Yoon, W.J.; Woo, K.M.; Baek, J.H.; Lee, G.; Park, J.C.; Ku, Y.; Ryoo, H.M. Epigenetic priming confers direct cell trans-differentiation from adipocyte to osteoblast in a transgene-free state. J. Cell. Physiol. 2016, 231, 1484–1494. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.G.; Putnam, D.; Lavik, E.B.; Mahmood, T.A.; Langer, R. Biomaterial microarrays: Rapid, microscale screening of polymer-cell interaction. Biomaterials 2005, 26, 4892–4897. [Google Scholar] [CrossRef] [PubMed]
- Levenberg, S.; Langer, R. Advances in tissue engineering. Curr. Top. Dev. Biol. 2004, 61, 113–134. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Xiao, Z.; Chen, T.; Wei, J.; Chen, L.; Liu, L.; Chen, B.; Wang, X.; Li, X.; Dai, J. The miR-7 identified from collagen biomaterial-based three-dimensional cultured cells regulates neural stem cell differentiation. Stem Cells Dev. 2014, 23, 393–405. [Google Scholar] [CrossRef]
- Kulangara, K.; Yang, J.; Chellappan, M.; Yang, Y.; Leong, K.W. Nanotopography alters nuclear protein expression, proliferation and differentiation of human mesenchymal stem/stromal cells. PLoS ONE 2014, 9, e114698. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, X.L.; Yang, S.C.; Lin, X.; He, Y.; Yan, C.; Wu, L.; Chen, G.Q.; Wang, Z.Y.; Wu, Q. MicroRNAs in the regulation of interfacial behaviors of MSCs cultured on microgrooved surface pattern. Biomaterials 2011, 32, 9207–9217. [Google Scholar] [CrossRef] [PubMed]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix elasticity directs stem cell lineage specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Tang, Y.; Zhang, P.; Liu, Y.; Bai, X.; Zhou, Y. Biomaterial Cues Regulate Epigenetic State and Cell Functions—A Systematic Review. Tissue Eng. Part B Rev. 2018, 24, 112–132. [Google Scholar] [CrossRef]
- Marsit, C.J. Influence of environmental exposure on human epigenetic regulation. J. Exp. Biol. 2015, 218, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.D.; Yoon, W.J.; Kim, W.J.; Woo, K.M.; Baek, J.H.; Lee, G.; Ku, Y.; van Wijnen, A.J.; Ryoo, H.M. Epigenetic modifications and canonical wingless/int-1 class (WNT) signaling enable trans-differentiation of nonosteogenic cells into osteoblasts. J. Biol. Chem. 2014, 289, 20120–20128. [Google Scholar] [CrossRef]
- Larsson, L.; Pilipchuk, S.P.; Giannobile, W.V.; Castilho, R.M. When epigenetics meets bioengineering-A material characteristics and surface topography perspective. J. Biomed. Mater. Res. B 2018, 106, 2065–2071. [Google Scholar] [CrossRef] [PubMed]
- Ichioka, Y.; Asa’ad, F.; Malekzadeh, B.O.; Westerlund, A.; Larsson, L. Epigenetic changes of osteoblasts in response to titanium surface characteristics. J. Biomed. Mater. Res. A 2020. [Google Scholar] [CrossRef]
- Zheng, Y.; Zheng, Y.F.; Jia, L.F.; Zhang, Y.; Lin, Y. Integrated analysis of lncRNA-mRNA networks associated with an SLA titanium surface reveals the potential role of HIF1A-AS1 in bone remodeling. RSC Adv. 2020, 10, 20972–20990. [Google Scholar] [CrossRef]
- Pacheco, K.A. Epigenetics mediate environment: Gene effects on occupational sensitization. Curr. Opin. Allergy Clin. Immunol. 2012, 12, 111–118. [Google Scholar] [CrossRef]
- Feil, R.; Fraga, M.F. Epigenetics and the environment: Emerging patterns and implications. Nat. Rev. Genet. 2012, 13, 97–109. [Google Scholar] [CrossRef]
- Yu, F.; Shen, H.; Deng, H.W. Systemic analysis of osteoblast-specific DNA methylation marks reveals novel epigenetic basis of osteoblast differentiation. Bone Rep. 2017, 6, 109–119. [Google Scholar] [CrossRef]
- Cho, Y.D.; Kim, B.S.; Kim, W.J.; Kim, H.J.; Baek, J.H.; Woo, K.M.; Seol, Y.J.; Ku, Y.; Ryoo, H.M. Histone acetylation together with DNA demethylation empowers higher plasticity in adipocytes to differentiate into osteoblasts. Gene 2020, 733, 144274. [Google Scholar] [CrossRef]
- Cho, Y.D.; Ryoo, H.M. Trans-differentiation via Epigenetics: A New Paradigm in the Bone Regeneration. J. Bone Metab. 2018, 25, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Kim, B.; Bae, H.; Kim, W.; Baek, J.; Woo, K.; Lee, G.; Seol, Y.; Lee, Y.; Ku, Y.; et al. Direct gingival fibroblast/osteoblast transdifferentiation via epigenetics. J. Dent. Res. 2017, 96, 555–561. [Google Scholar] [CrossRef]
- Ehara, T.; Kamei, Y.; Takahashi, M.; Yuan, X.; Kanai, S.; Tamura, E.; Tanaka, M.; Yamazaki, T.; Miura, S.; Ezaki, O.; et al. Role of DNA methylation in the regulation of lipogenic glycerol-3-phosphate acyltransferase 1 gene expression in the mouse neonatal liver. Diabetes 2012, 61, 2442–2450. [Google Scholar] [CrossRef]
- Michelotti, G.A.; Brinkley, D.M.; Morris, D.P.; Smith, M.P.; Louie, R.J.; Schwinn, D.A. Epigenetic regulation of human alpha1d-adrenergic receptor gene expression: A role for DNA methylation in Sp1-dependent regulation. FASEB J. 2007, 21, 1979–1993. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.P.; Lindroth, A.M.; Cao, X.; Jacobsen, S.E. Control of CpNpG DNA methylation by the KRYPTONITE histone H3 methyltransferase. Nature 2002, 416, 556–560. [Google Scholar] [CrossRef] [PubMed]
- Ghayor, C.; Weber, F.E. Epigenetic Regulation of Bone Remodeling and Its Impacts in Osteoporosis. Int. J. Mol. Sci. 2016, 17, 1446. [Google Scholar] [CrossRef]
- Meyer, M.B.; Benkusky, N.A.; Sen, B.; Rubin, J.; Pike, J.W. Epigenetic Plasticity Drives Adipogenic and Osteogenic Differentiation of Marrow-derived Mesenchymal Stem Cells. J. Biol. Chem. 2016, 291, 17829–17847. [Google Scholar] [CrossRef]
- Zaidi, S.K.; Young, D.W.; Montecino, M.A.; Lian, J.B.; van Wijnen, A.J.; Stein, J.L.; Stein, G.S. Mitotic bookmarking of genes: A novel dimension to epigenetic control. Nat. Rev. Genet. 2010, 11, 583–589. [Google Scholar] [CrossRef]
- Gordon, J.A.R.; Stein, J.L.; Westendorf, J.J.; van Wijnen, A.J. Chromatin modifiers and histone modifications in bone formation, regeneration, and therapeutic intervention for bone-related disease. Bone 2015, 81, 739–745. [Google Scholar] [CrossRef]
- Lv, L.; Liu, Y.; Zhang, P.; Zhang, X.; Liu, J.; Chen, T.; Su, P.; Li, H.; Zhou, Y. The nanoscale geometry of TiO2 nanotubes influences the osteogenic differentiation of human adipose-derived stem cells by modulating H3K4 trimethylation. Biomaterials 2015, 39, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Andrukhov, O.; Huber, R.; Shi, B.; Berner, S.; Rausch-Fan, X.; Moritz, A.; Spencer, N.D.; Schedle, A. Proliferation, behavior, and differentiation of osteoblasts on surfaces of different microroughness. Dent. Mater. 2016, 32, 1374–1384. [Google Scholar] [CrossRef]
- Gittens, R.A.; Scheideler, L.; Rupp, F.; Hyzy, S.L.; Geis-Gerstorfer, J.; Schwartz, Z.; Boyan, B.D. A review on the wettability of dental implant surfaces II: Biological and clinical aspects. Acta Biomater. 2014, 10, 2907–2918. [Google Scholar] [CrossRef]
- Kennedy, S.B.; Washburn, N.R.; Simon, C.G., Jr.; Amis, E.J. Combinatorial screen of the effect of surface energy on fibronectin-mediated osteoblast adhesion, spreading and proliferation. Biomaterials 2006, 27, 3817–3824. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, C.; Nygren, H.; Ohlson, K. Implantation of hydrophilic and hydrophobic titanium discs in rat tibia: Cellular reactions on the surfaces during the first 3 weeks in bone. Biomaterials 2004, 25, 4759–4766. [Google Scholar] [CrossRef] [PubMed]
- Lamolle, S.F.; Monjo, M.; Rubert, M.; Haugen, H.J.; Lyngstadaas, S.P.; Ellingsen, J.E. The effect of hydrofluoric acid treatment of titanium surface on nanostructural and chemical changes and the growth of MC3T3-E1 cells. Biomaterials 2009, 30, 736–742. [Google Scholar] [CrossRef]
- Zareidoost, A.; Yousefpour, M.; Ghaseme, B.; Amanzadeh, A. The relationship of surface roughness and cell response of chemical surface modification of titanium. J. Mater. Sci. Mater. Med. 2012, 23, 1479–1488. [Google Scholar] [CrossRef]
- Silva, T.S.; Machado, D.C.; Viezzer, C.; Silva Junior, A.N.; Oliveira, M.G. Effect of titanium surface roughness on human bone marrow cell proliferation and differentiation: An experimental study. Acta Cir. Bras. 2009, 24, 200–205. [Google Scholar] [CrossRef]
- Cho, Y.D.; Shin, J.C.; Yoon, H.I.; Ku, Y.; Ryoo, H.M.; Kim, D.J.; Kim, D.G.; Han, J.S. Characterization of Human Gingival Fibroblasts on Zirconia Surfaces Containing Niobium Oxide. Materials 2015, 8, 6018–6028. [Google Scholar] [CrossRef]
- Cho, Y.; Hong, J.; Ryoo, H.; Kim, D.; Park, J.; Han, J. Osteogenic responses to zirconia with hydroxyapatite coating by aerosol deposition. J. Dent. Res. 2015, 94, 491–499. [Google Scholar] [CrossRef]
- Cho, Y.D.; Shin, J.C.; Kim, H.L.; Gerelmaa, M.; Yoon, H.I.; Ryoo, H.M.; Kim, D.J.; Han, J.S. Comparison of the osteogenic potential of titanium- and modified zirconia-based bioceramics. Int. J. Mol. Sci. 2014, 15, 4442–4452. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, Y.-D.; Kim, W.-J.; Kim, S.; Ku, Y.; Ryoo, H.-M. Surface Topography of Titanium Affects Their Osteogenic Potential through DNA Methylation. Int. J. Mol. Sci. 2021, 22, 2406. https://doi.org/10.3390/ijms22052406
Cho Y-D, Kim W-J, Kim S, Ku Y, Ryoo H-M. Surface Topography of Titanium Affects Their Osteogenic Potential through DNA Methylation. International Journal of Molecular Sciences. 2021; 22(5):2406. https://doi.org/10.3390/ijms22052406
Chicago/Turabian StyleCho, Young-Dan, Woo-Jin Kim, Sungtae Kim, Young Ku, and Hyun-Mo Ryoo. 2021. "Surface Topography of Titanium Affects Their Osteogenic Potential through DNA Methylation" International Journal of Molecular Sciences 22, no. 5: 2406. https://doi.org/10.3390/ijms22052406
APA StyleCho, Y.-D., Kim, W.-J., Kim, S., Ku, Y., & Ryoo, H.-M. (2021). Surface Topography of Titanium Affects Their Osteogenic Potential through DNA Methylation. International Journal of Molecular Sciences, 22(5), 2406. https://doi.org/10.3390/ijms22052406