
Differential Responses to Bioink-Induced Oxidative Stress in Endothelial Cells and Fibroblasts

 , ,
, ,  ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
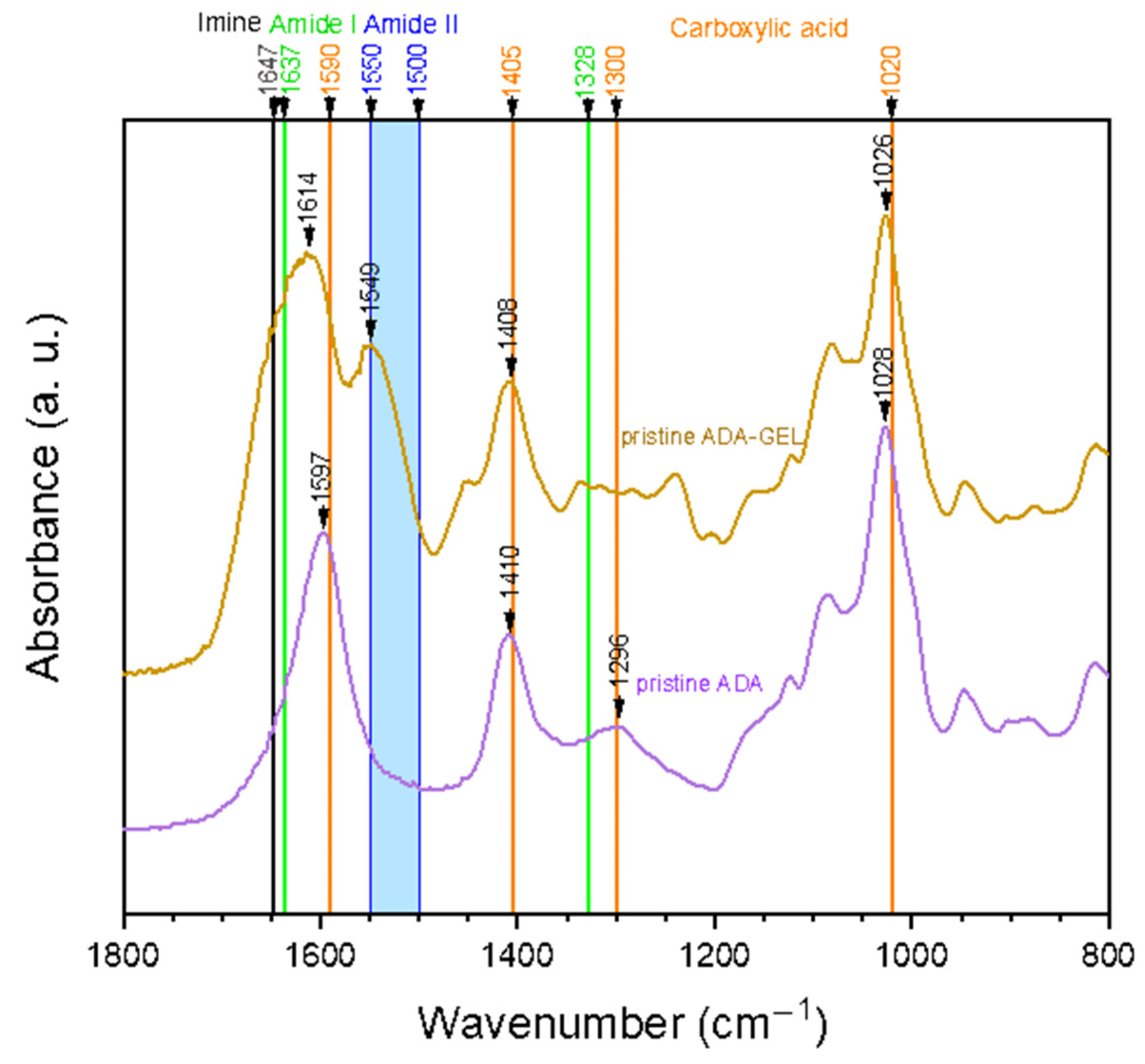
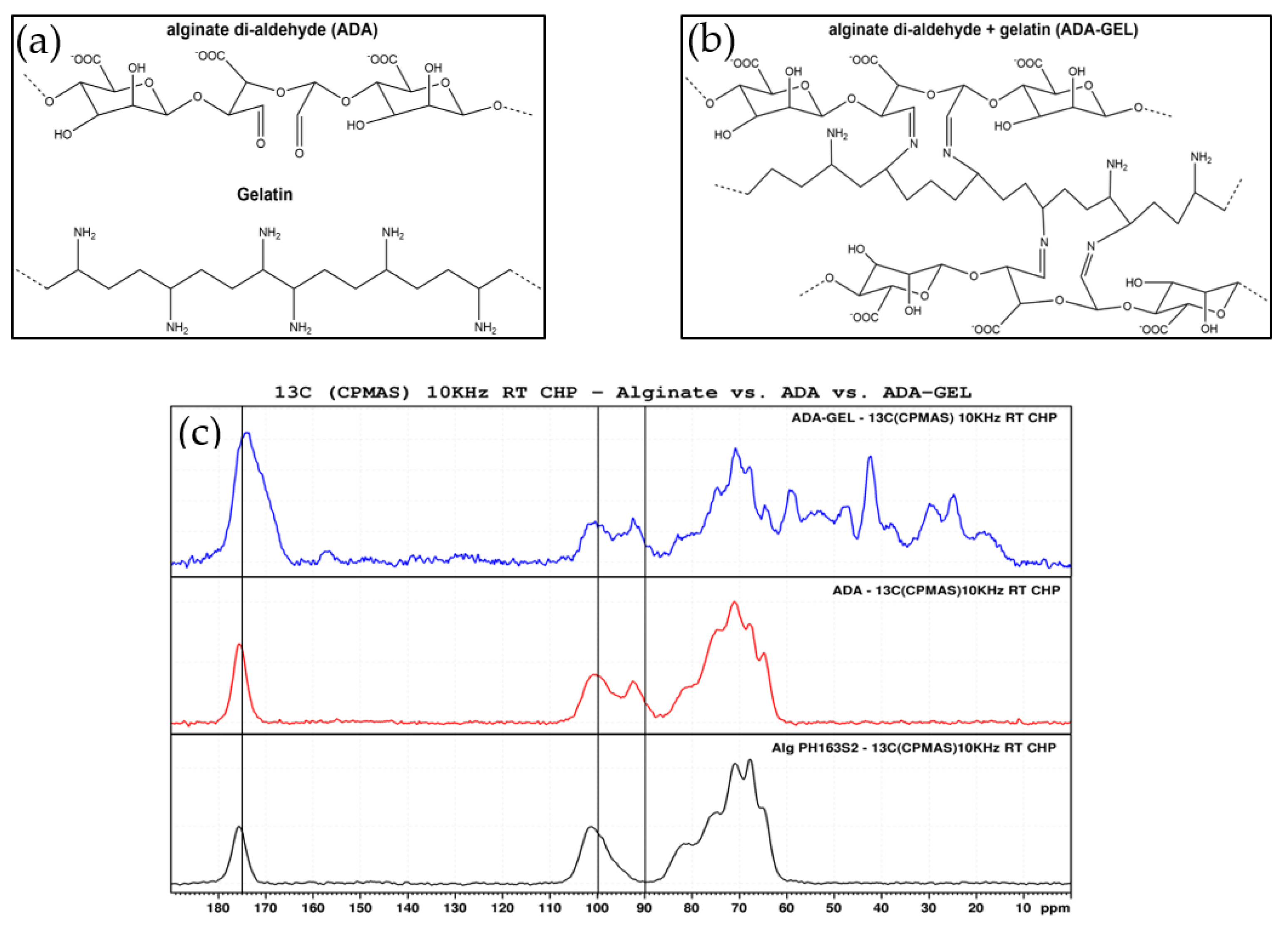
2.1. Hydrogel Characterization
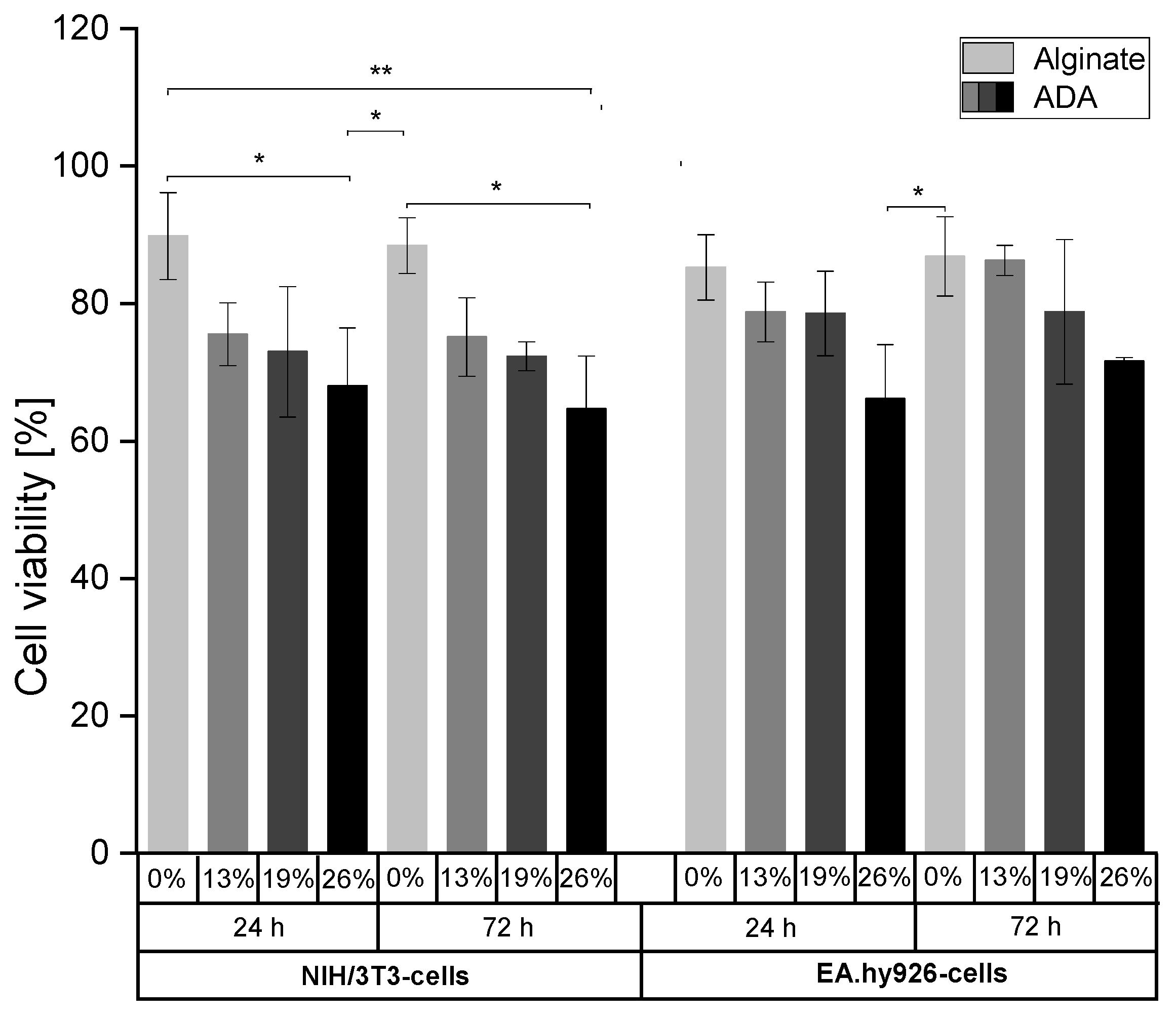
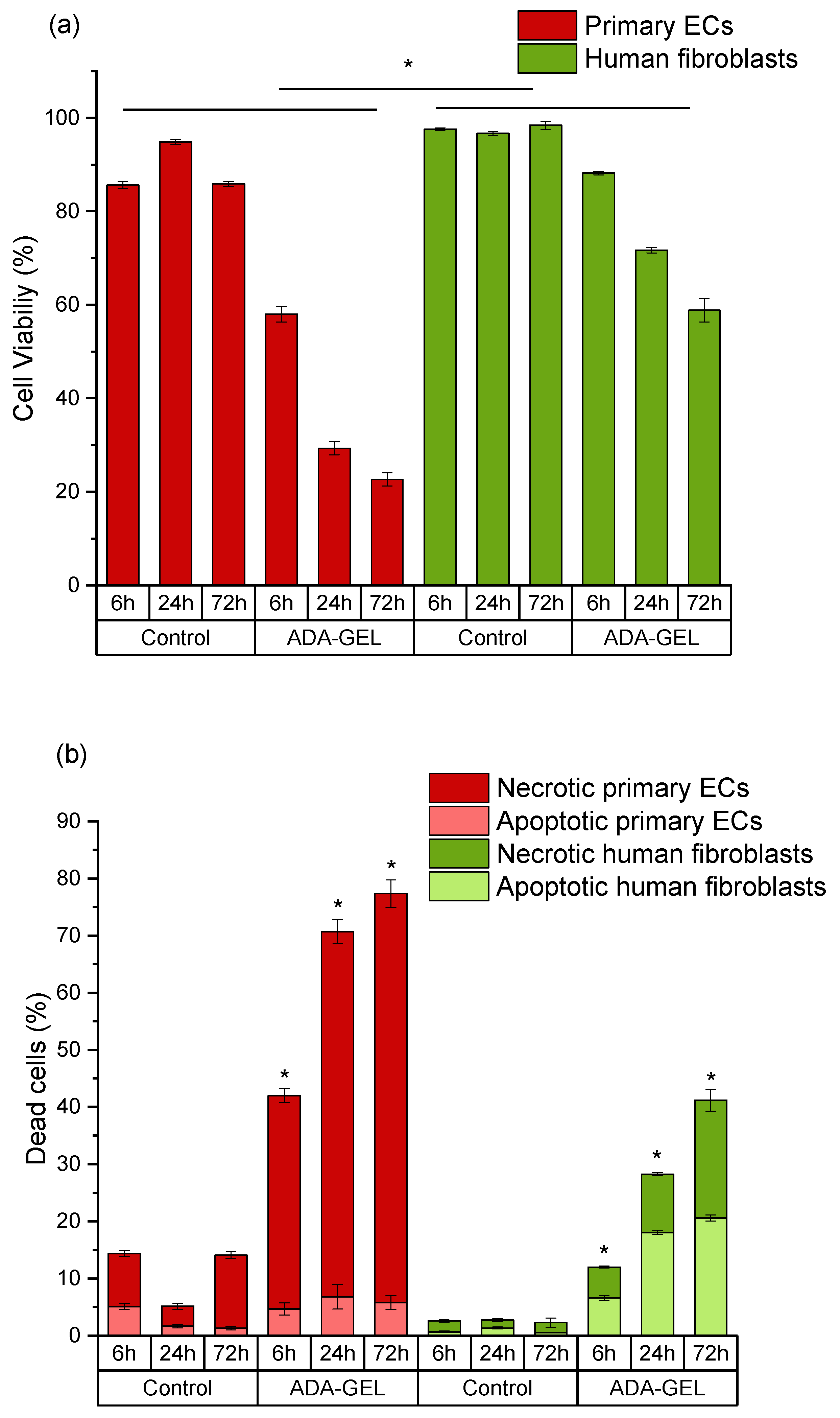
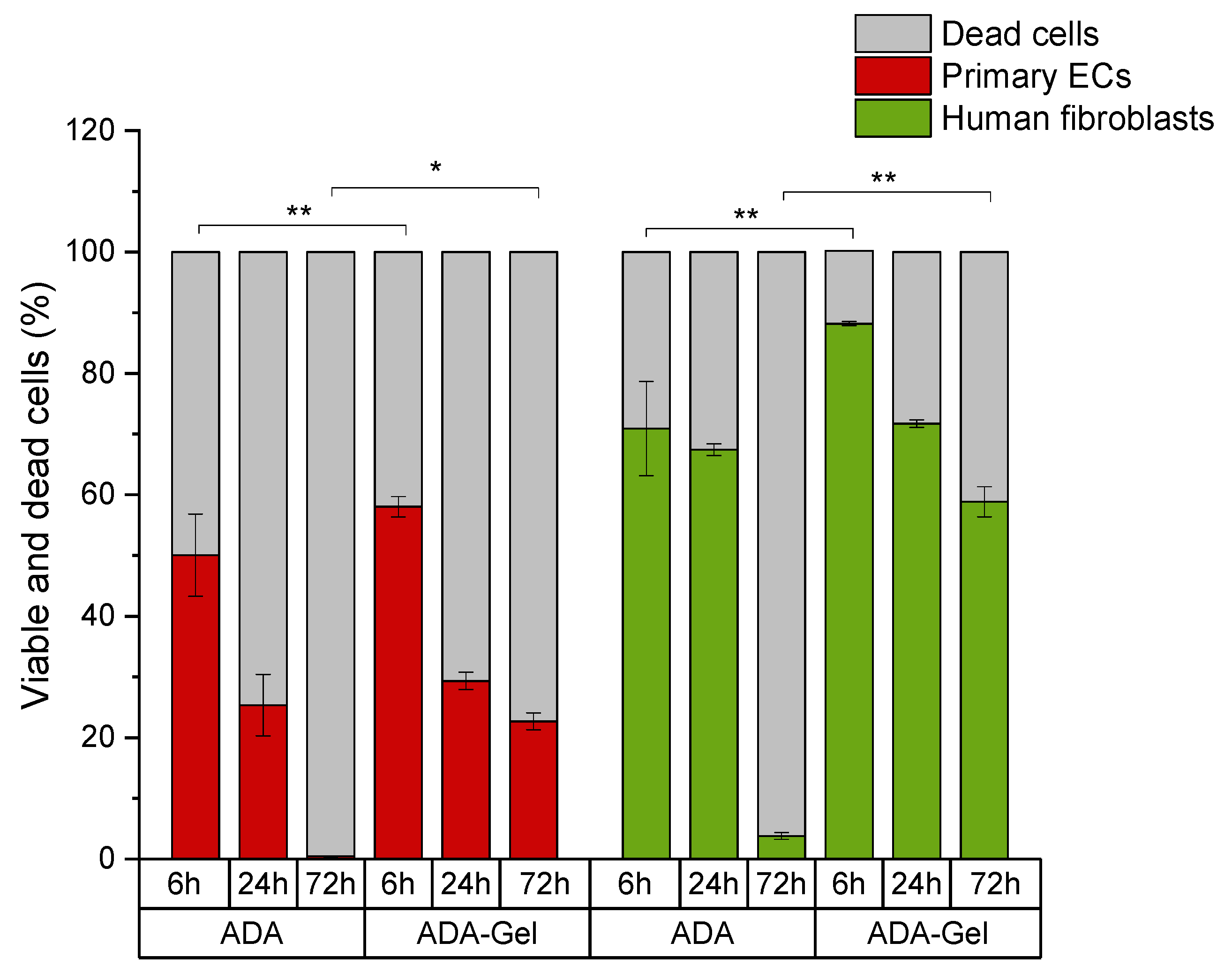
2.2. Evaluation of Cell Viability as a Function of Oxidation Degree
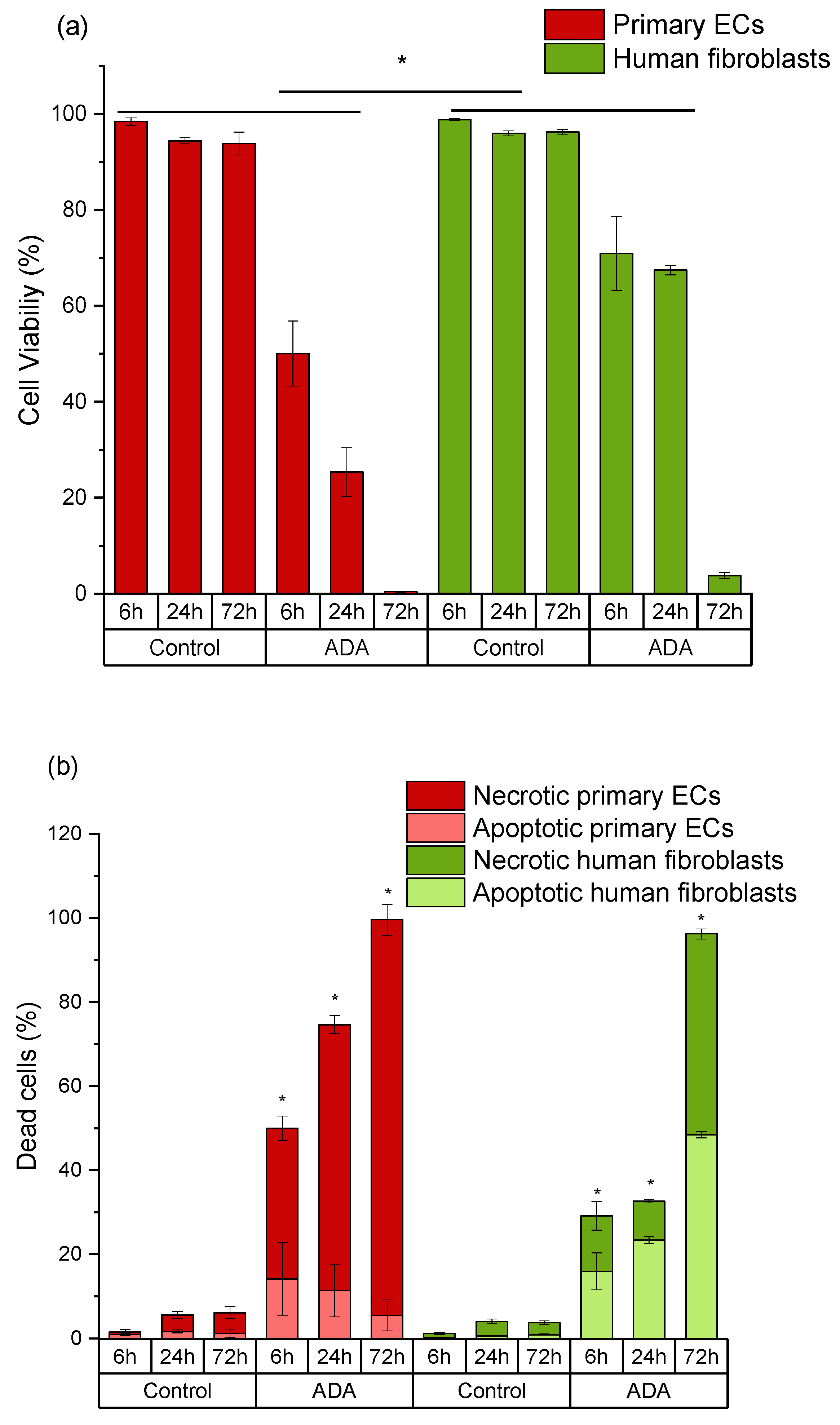
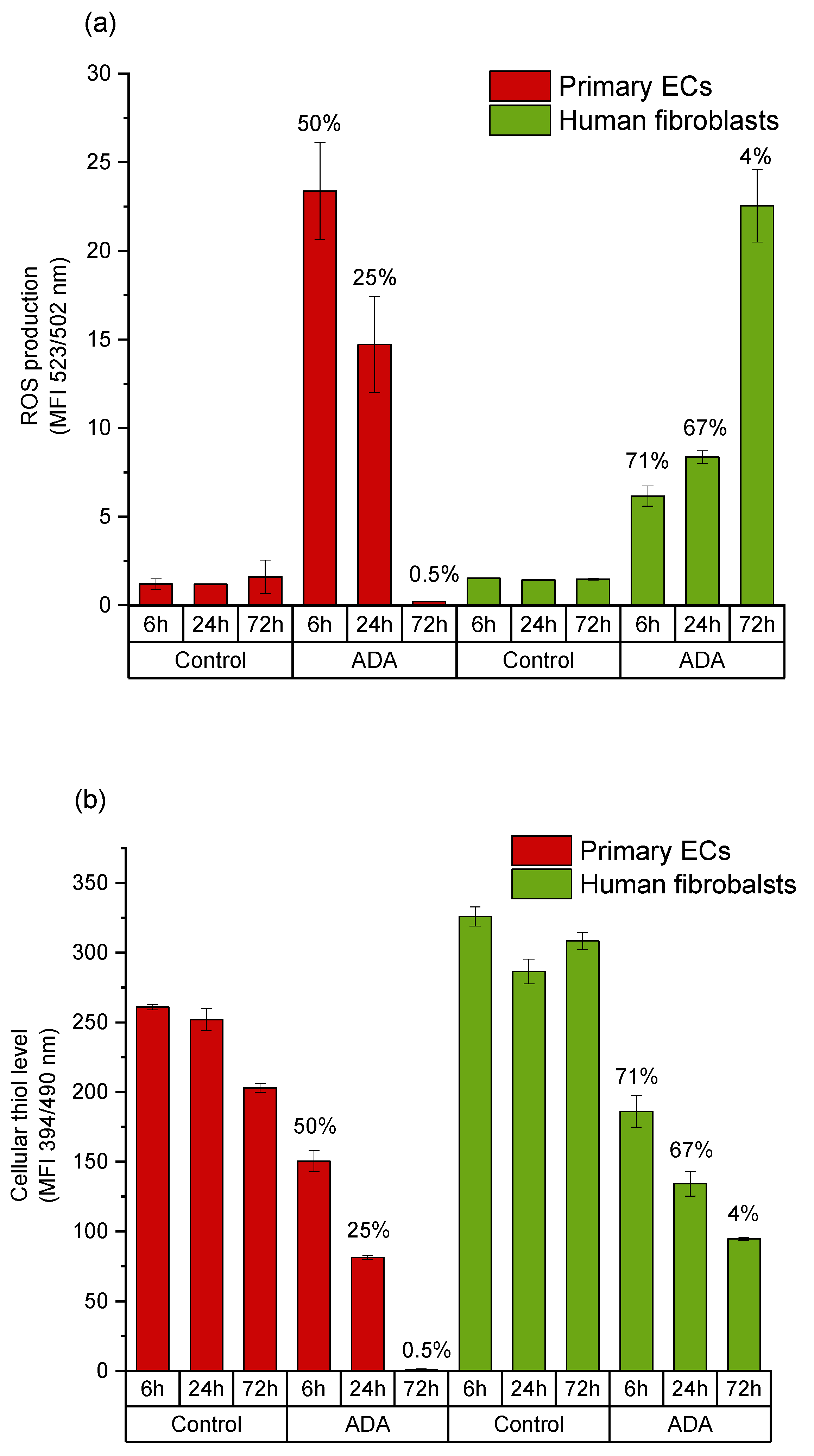
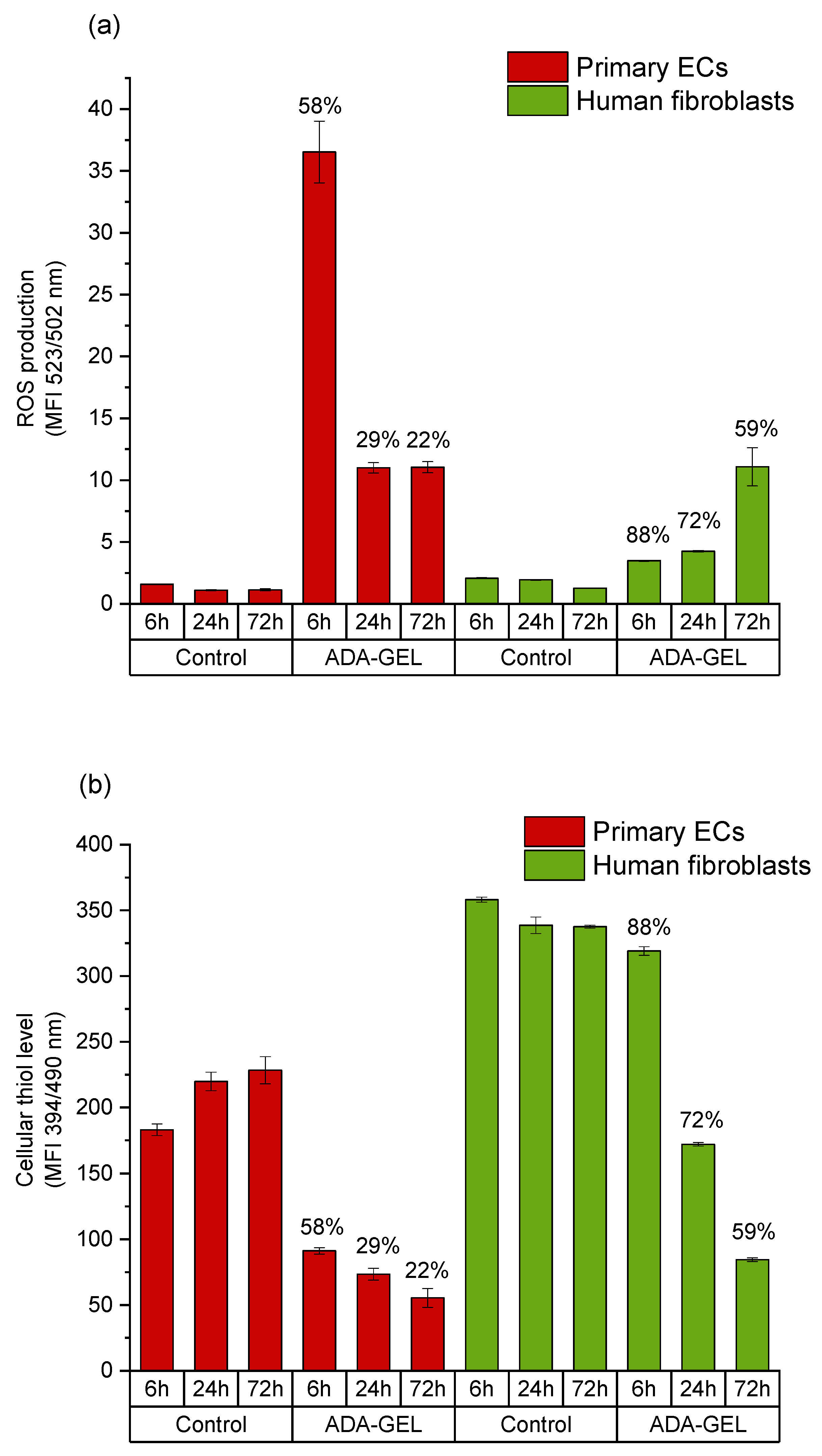
2.3. Mechanisms of Oxidized Material-Induced Cytotoxicity in Primary Human Cells
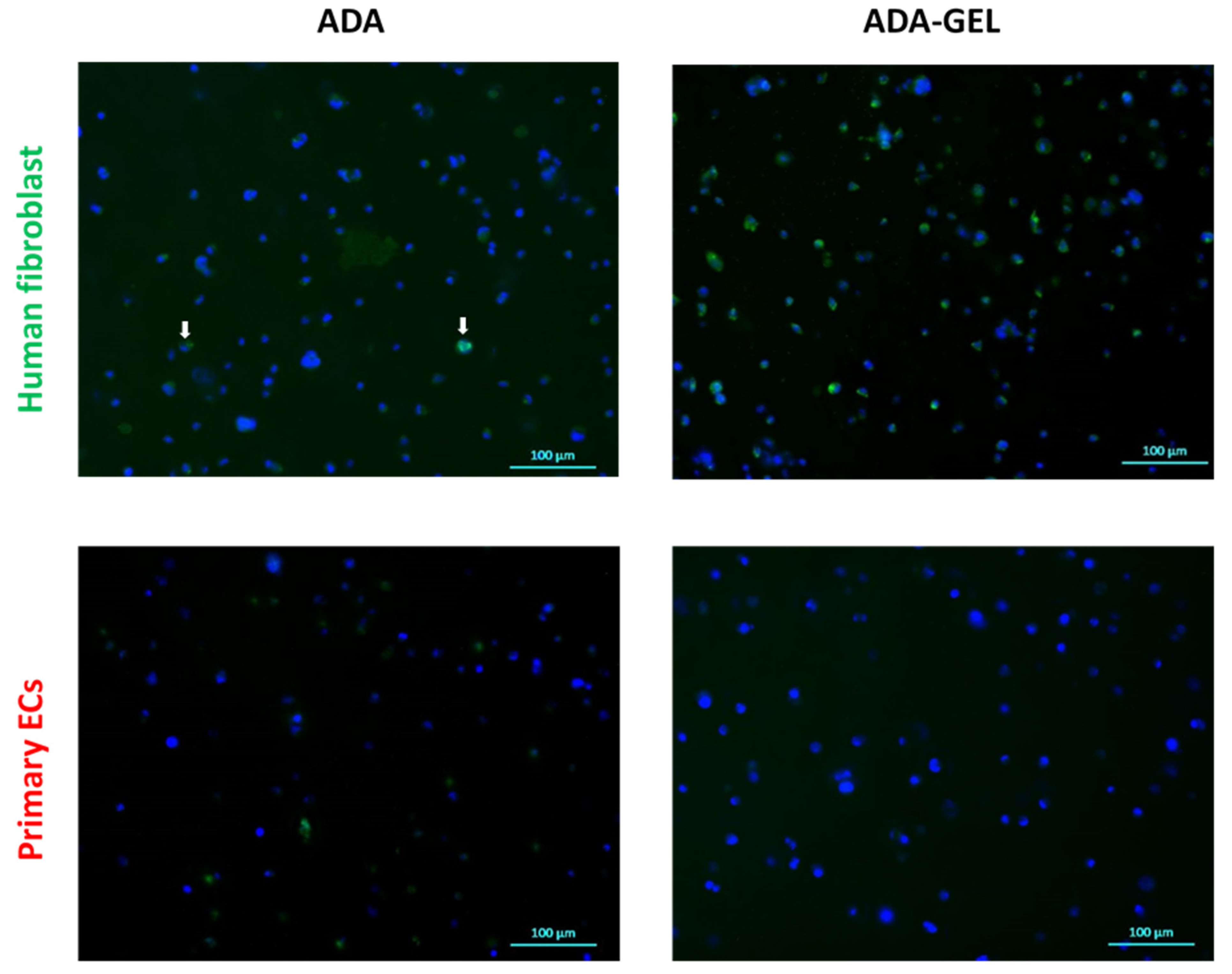
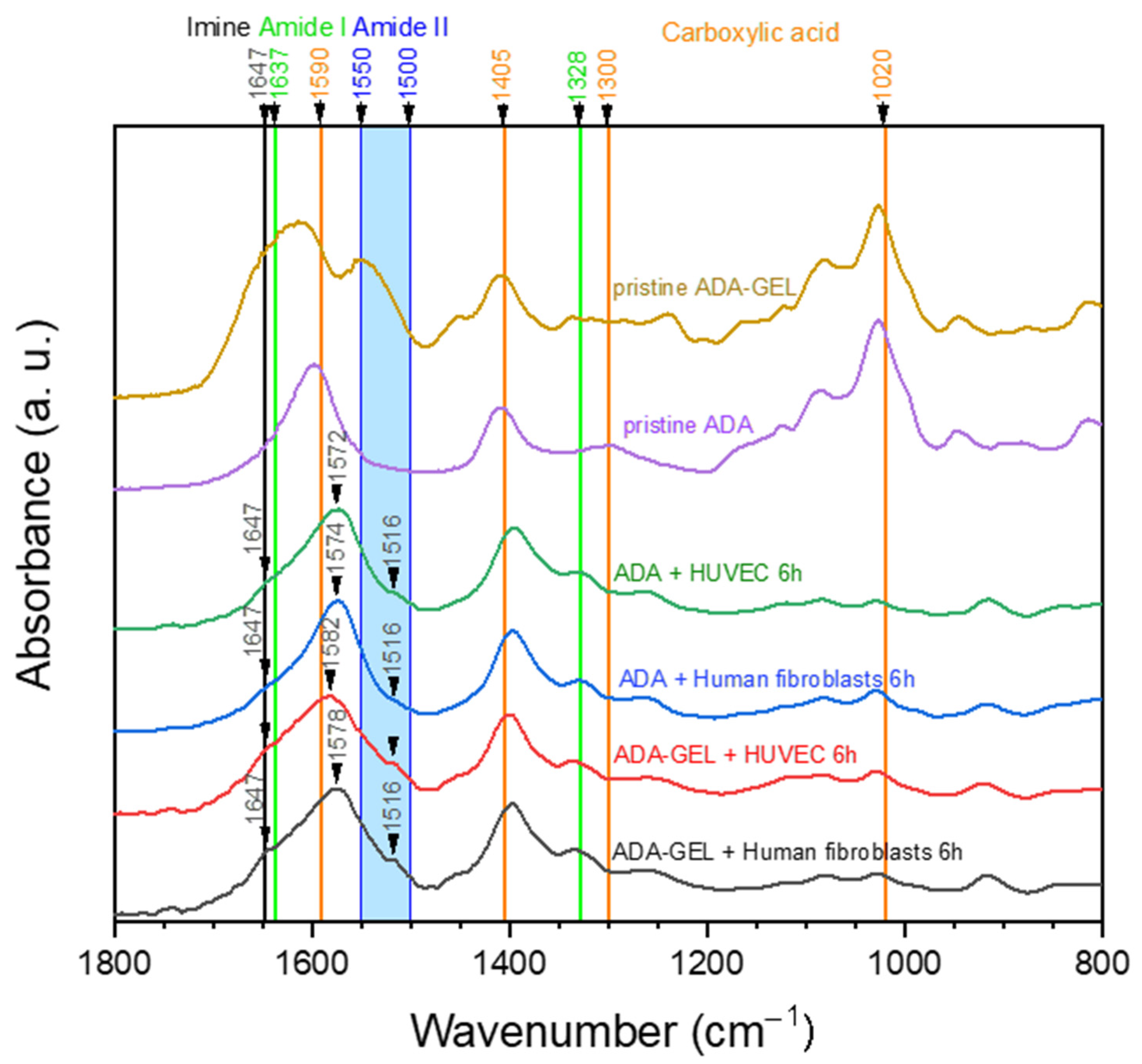
2.4. ATR-FTIR of Cell-Containing Hydrogels
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Synthesis of Alginate Di-Aldehyde (ADA)
4.3. Hydrogel Formulations
4.4. Chemical Characterization of ADA and ADA-GEL
4.4.1. Fourier-Transform Infrared Spectroscopy
4.4.2. Solid State 13 C NMR Spectroscopy
4.5. Cell Culture
4.5.1. Endothelial Cells
4.5.2. Fibroblasts
4.6. Cell Seeding in the Hydrogels
4.7. Analysis of Cell Viability
4.8. Flow-Cytometry Analyses of Cells
4.8.1. Preparation of 2′,7′-Dichlorofluorescin Diacetate (DCFH-DA) Probe for Intracellular ROS Detection
4.8.2. Preparation of Staining Solutions and Analysis of Flow Data
4.9. Immunofluorescence Staining of Samples
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ADA | alginate di-aldehyde |
| ATR-FTIR | attenuated total reflectance-Fourier transform infrared spectroscopy |
| DAPI | 2-[4-(aminoiminomethyl)phenyl]-1H-indole-6-carboximidamide hydrochloride |
| DCF | 2′,7′-dichlorofluorescein |
| DCHF-DA | 2′,7′-dichlorofluorescein diacetate |
| DiI | 1,1′,3,3,3′,3′-hexamethylindodicarbocyanine iodide |
| DMEM | Dulbecco’s modified Eagle medium |
| DNA | deoxyribonucleic acid |
| DO | degree of oxidation in percent |
| DPBS | Dulbecco’s phosphate buffered saline |
| EA.hy926 | human endothelial cell line |
| ECM | extracellular matrix |
| ECs | endothelial cells |
| EDTA | ethylenediaminetetraacetic acid |
| FCS | fetal calf serum |
| GEL | gelatin |
| GSH | glutathione |
| HBSS | Hank’s balanced salt solution |
| HUVEC | human umbilical vein endothelial cells |
| MBB | monobromobimane |
| NIH/3T3 | mouse fibroblast cell line |
| NMR | nuclear magnetic resonance spectroscopy |
| PI | propidium iodide |
| ROS | reactive oxygen species |
| SEM | standard error of mean |
References
- Schwab, A.; Levato, R.; D’Este, M.; Piluso, S.; Eglin, D.; Malda, J. Printability and Shape Fidelity of Bioinks in 3D Bioprinting. Chem. Rev. 2020, 120, 10850–10877. [Google Scholar] [CrossRef] [PubMed]
- Li, H.J.; Tan, C.; Li, L. Review of 3D printable hydrogels and constructs. Mater. Des. 2018, 159, 20–38. [Google Scholar] [CrossRef]
- Holzl, K.; Lin, S.M.; Tytgat, L.; Van Vlierberghe, S.; Gu, L.X.; Ovsianikov, A. Bioink properties before, during and after 3D bioprinting. Biofabrication 2016, 8, 032002. [Google Scholar] [CrossRef]
- Chung, J.H.Y.; Naficy, S.; Yue, Z.L.; Kapsa, R.; Quigley, A.; Moulton, S.E.; Wallace, G.G. Bio-ink properties and printability for extrusion printing living cells. Biomater. Sci. 2013, 1, 763–773. [Google Scholar] [CrossRef]
- Levato, R.; Jungst, T.; Scheuring, R.G.; Blunk, T.; Groll, J.; Malda, J. From Shape to Function: The Next Step in Bioprinting. Adv. Mater. 2020, 32, 1906423. [Google Scholar] [CrossRef]
- Ashton, R.S.; Banerjee, A.; Punyani, S.; Schaffer, D.V.; Kane, R.S. Scaffolds based on degradable alginate hydrogels and poly(lactide-co-glycolide) microspheres for stem cell culture. Biomaterials 2007, 28, 5518–5525. [Google Scholar] [CrossRef]
- Ruvinov, E.; Cohen, S. Alginate biomaterial for the treatment of myocardial infarction: Progress, translational strategies, and clinical outlook: From ocean algae to patient bedside. Adv. Drug Deliv. Rev. 2016, 96, 54–76. [Google Scholar] [CrossRef]
- Mann, D.L.; Lee, R.J.; Coats, A.J.; Neagoe, G.; Dragomir, D.; Pusineri, E.; Piredda, M.; Bettari, L.; Kirwan, B.A.; Dowling, R.; et al. One-year follow-up results from AUGMENT-HF: A multicentre randomized controlled clinical trial of the efficacy of left ventricular augmentation with Algisyl in the treatment of heart failure. Eur. J. Heart Fail. 2016, 18, 314–325. [Google Scholar] [CrossRef]
- Jia, J.; Richards, D.J.; Pollard, S.; Tan, Y.; Rodriguez, J.; Visconti, R.P.; Trusk, T.C.; Yost, M.J.; Yao, H.; Markwald, R.R.; et al. Engineering alginate as bioink for bioprinting. Acta Biomater. 2014, 10, 4323–4331. [Google Scholar] [CrossRef] [PubMed]
- Sarker, B.; Papageorgiou, D.G.; Silva, R.; Zehnder, T.; Gul-E.-Noor, F.; Bertmer, M.; Kaschta, J.; Chrissafis, K.; Detsch, R.; Boccaccini, A.R. Fabrication of alginate-gelatin crosslinked hydrogel microcapsules and evaluation of the microstructure and physico-chemical properties. J. Mater. Chem. B 2014, 2, 1470–1482. [Google Scholar] [CrossRef] [PubMed]
- Rottensteiner, U.; Sarker, B.; Heusinger, D.; Dafinova, D.; Rath, S.N.; Beier, J.P.; Kneser, U.; Horch, R.E.; Detsch, R.; Boccaccini, A.R.; et al. In vitro and in vivo Biocompatibility of Alginate Dialdehyde/Gelatin Hydrogels with and without Nanoscaled Bioactive Glass for Bone Tissue Engineering Applications. Materials 2014, 7, 1957–1974. [Google Scholar] [CrossRef]
- Reakasame, S.; Boccaccini, A.R. Oxidized Alginate-Based Hydrogels for Tissue Engineering Applications: A Review. Biomacromolecules 2018, 19, 3–21. [Google Scholar] [CrossRef]
- Weis, M.; Shan, J.W.; Kuhlmann, M.; Jungst, T.; Tessmar, J.; Groll, J. Evaluation of Hydrogels Based on Oxidized Hyaluronic Acid for Bioprinting. Gels 2018, 4, 82. [Google Scholar] [CrossRef]
- Amirian, J.; Zeng, Y.; Shekh, M.I.; Sharma, G.; Stadler, F.J.; Song, J.; Du, B.; Zhu, Y.X. In-situ crosslinked hydrogel based on amidated pectin/oxidized chitosan as potential wound dressing for skin repairing. Carbohyd. Polym 2021, 251, 117005. [Google Scholar] [CrossRef]
- Balakrishnan, B.; Mohanty, M.; Umashankar, P.R.; Jayakrishnan, A. Evaluation of an in situ forming hydrogel wound dressing based on oxidized alginate and gelatin. Biomaterials 2005, 26, 6335–6342. [Google Scholar] [CrossRef]
- Bouhadir, K.H.; Lee, K.Y.; Alsberg, E.; Damm, K.L.; Anderson, K.W.; Mooney, D.J. Degradation of partially oxidized alginate and its potential application for tissue engineering. Biotechnol. Progr. 2001, 17, 945–950. [Google Scholar] [CrossRef]
- Sakai, S.; Yamaguchi, S.; Takei, T.; Kawakami, K. Oxidized alginate-cross-linked alginate/gelatin hydrogel fibers for fabricating tubular constructs with layered smooth muscle cells and endothelial cells in collagen gels. Biomacromolecules 2008, 9, 2036–2041. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.S.; Mooney, D.J.; Arany, P.R.; Lee, K.; Huebsch, N.; Kim, J. Adipose Tissue Engineering Using Injectable, Oxidized Alginate Hydrogels. Tissue Eng. Pt. A 2012, 18, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Sarker, B.; Singh, R.; Silva, R.; Roether, J.A.; Kaschta, J.; Detsch, R.; Schubert, D.W.; Cicha, I.; Boccaccini, A.R. Evaluation of Fibroblasts Adhesion and Proliferation on Alginate-Gelatin Crosslinked Hydrogel. PLoS ONE 2014, 9, e107952. [Google Scholar] [CrossRef] [PubMed]
- Zehnder, T.; Sarker, B.; Boccaccini, A.R.; Detsch, R. Evaluation of an alginate-gelatine crosslinked hydrogel for bioplotting. Biofabrication 2015, 7, 025001. [Google Scholar] [CrossRef]
- Silva, R.; Singh, R.; Sarker, B.; Papageorgiou, D.G.; Juhasz-Bortuzzo, J.A.; Roether, J.A.; Cicha, I.; Kaschta, J.; Schubert, D.W.; Chrissafis, K.; et al. Hydrogel matrices based on elastin and alginate for tissue engineering applications. Int. J. Biol. Macromol. 2018, 114, 614–625. [Google Scholar] [CrossRef]
- Distler, T.; McDonald, K.; Heid, S.; Karakaya, E.; Detsch, R.; Boccaccini, A.R. Ionically and Enzymatically Dual Cross-Linked Oxidized Alginate Gelatin Hydrogels with Tunable Stiffness and Degradation Behavior for Tissue Engineering. ACS Biomater. Sci. Eng. 2020, 6, 3899–3914. [Google Scholar] [CrossRef]
- Ivanovska, J.; Zehnder, T.; Lennert, P.; Sarker, B.; Boccaccini, A.R.; Hartmann, A.; Schneider-Stock, R.; Detsch, R. Biofabrication of 3D Alginate-Based Hydrogel for Cancer Research: Comparison of Cell Spreading, Viability, and Adhesion Characteristics of Colorectal HCT116 Tumor Cells. Tissue Eng. Part. C 2016, 22, 708–715. [Google Scholar] [CrossRef]
- Leite, A.J.; Sarker, B.; Zehnder, T.; Silva, R.; Mano, J.F.; Boccaccini, A.R. Bioplotting of a bioactive alginate dialdehyde-gelatin composite hydrogel containing bioactive glass nanoparticles. Biofabrication 2016, 8, 035005. [Google Scholar] [CrossRef] [PubMed]
- Dranseikiene, D.; Schrufer, S.; Schubert, D.W.; Reakasame, S.; Boccaccini, A.R. Cell-laden alginate dialdehyde-gelatin hydrogels formed in 3D printed sacrificial gel. J. Mater. Sci.-Mater. 2020, 31, 1–5. [Google Scholar] [CrossRef]
- Feng, Q.; Wei, K.C.; Zhang, K.Y.; Yang, B.G.; Tian, F.; Wang, G.X.; Bian, L.M. One-pot solvent exchange preparation of non-swellable, thermoplastic, stretchable and adhesive supramolecular hydrogels based on dual synergistic physical crosslinking. NPG Asia Mater. 2018, 10, e455. [Google Scholar] [CrossRef]
- You, F.; Wu, X.; Kelly, M.; Chen, X.B. Bioprinting and in vitro characterization of alginate dialdehyde-gelatin hydrogel bio-ink. Bio-Des. Manuf. 2020, 3, 48–59. [Google Scholar] [CrossRef]
- Soltan, N.; Ning, L.Q.; Mohabatpour, F.; Papagerakis, P.; Chen, X.B. Printability and Cell Viability in Bioprinting Alginate Dialdehyde-Gelatin Scaffolds. ACS Biomater. Sci. Eng. 2019, 5, 2976–2987. [Google Scholar] [CrossRef] [PubMed]
- Baniasadi, H.; Mashayekhan, S.; Fadaoddini, S.; Haghirsharifzamini, Y. Design, fabrication and characterization of oxidized alginate-gelatin hydrogels for muscle tissue engineering applications. J. Biomater. Appl. 2016, 31, 152–161. [Google Scholar] [CrossRef]
- Rosellini, E.; Cristallini, C.; Barbani, N.; Vozzi, G.; Giusti, P. Preparation and characterization of alginate/gelatin blend films for cardiac tissue engineering. J. Biomed. Mater. Res. A 2009, 91, 447–453. [Google Scholar] [CrossRef]
- Medeiros, M.H.G. DNA Damage by Endogenous and Exogenous Aldehydes. J. Braz. Chem. Soc. 2019, 30, 2000–2009. [Google Scholar] [CrossRef]
- Stone, R.C.; Kim, S.; Barnes, B.J.; Aviv, A. Diverging Antioxidative Responses to IGF-1 in Cultured Human Skin Fibroblasts Versus Vascular Endothelial Cells. J. Gerontol. A-Biol. 2012, 67, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.A.; Ackerman, S.L. Oxidative stress, cell cycle, and neurodegeneration. J. Clin. Investig. 2003, 111, 785–793. [Google Scholar] [CrossRef]
- Martindale, J.L.; Holbrook, N.J. Cellular response to oxidative stress: Signaling for suicide and survival. J. Cell Physiol. 2002, 192, 1–15. [Google Scholar] [CrossRef]
- Qiao, L.Y.; Liu, C.D.; Liu, C.; Yang, L.Q.; Zhang, M.X.; Liu, W.T.; Wang, J.Y.; Jian, X.G. Self-healing alginate hydrogel based on dynamic acylhydrazone and multiple hydrogen bonds. J. Mater. Sci. 2019, 54, 8814–8828. [Google Scholar] [CrossRef]
- Chao, S.C.; Wang, M.J.; Pai, N.S.; Yen, S.K. Preparation and characterization of gelatin-hydroxyapatite composite microspheres for hard tissue repair. Mat. Sci. Eng. C-Mater. 2015, 57, 113–122. [Google Scholar] [CrossRef]
- Yuan, L.; Geng, X.; Li, J.; Sun, B.; Mo, X. Aldehyde-sodium alginate and amino-gelatin preparation as soft tissue adhesive. J. Donghua Univ. 2014, 31, 503–506. [Google Scholar]
- Wu, Y.; Yuan, L.; Sheng, N.A.; Gu, Z.Q.; Feng, W.H.; Yin, H.Y.; Morsi, Y.; Mo, X.M. A soft tissue adhesive based on aldehyde-sodium alginate and amino-carboxymethyl chitosan preparation through the Schiff reaction. Front. Mater. Sci. 2017, 11, 215–222. [Google Scholar] [CrossRef]
- Chen, F.; Tian, M.; Zhang, D.M.; Wang, J.Y.; Wang, Q.G.; Yu, X.X.; Zhang, X.H.; Wan, C.X. Preparation and characterization of oxidized alginate covalently cross-linked galactosylated chitosan scaffold for liver tissue engineering. Mat. Sci. Eng. C-Mater. 2012, 32, 310–320. [Google Scholar] [CrossRef]
- Ye, J.; Xiong, J.; Sun, R.C. The fluorescence property of Schiff’s bases of carboxymethyl cellulose. Carbohyd. Polym. 2012, 88, 1420–1424. [Google Scholar] [CrossRef]
- Jejurikar, A.; Seow, X.T.; Lawrie, G.; Martin, D.; Jayakrishnan, A.; Grondahl, L. Degradable alginate hydrogels crosslinked by the macromolecular crosslinker alginate dialdehyde. J. Mater. Chem. 2012, 22, 9751–9758. [Google Scholar] [CrossRef]
- Singh, R.; Wieser, A.; Reakasame, S.; Detsch, R.; Dietel, B.; Alexiou, C.; Boccaccini, A.R.; Cicha, I. Cell specificity of magnetic cell seeding approach to hydrogel colonization. J. Biomed. Mater. Res. A 2017, 105, 2948–2956. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, B.; Joshi, N.; Jayakrishnan, A.; Banerjee, R. Self-crosslinked oxidized alginate/gelatin hydrogel as injectable, adhesive biomimetic scaffolds for cartilage regeneration. Acta Biomater. 2014, 10, 3650–3663. [Google Scholar] [CrossRef] [PubMed]
- Grigore, A.; Sarker, B.; Fabry, B.; Boccaccini, A.R.; Detsch, R. Behavior of Encapsulated MG-63 Cells in RGD and Gelatine-Modified Alginate Hydrogels. Tissue Eng. Pt. A 2014, 20, 2140–2150. [Google Scholar] [CrossRef] [PubMed]
- Sokolsky-Papkov, M.; Domb, A.J.; Golenser, J. Impact of aldehyde content on amphotericin B-dextran imine conjugate toxicity. Biomacromolecules 2006, 7, 1529–1535. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, M.; Saretzki, G.; Sitte, N.; Metzkow, S.; von Zglinicki, T. BJ fibroblasts display high antioxidant capacity and slow telomere shortening independent of hTERT transfection. Free Radic. Bio Med. 2001, 31, 824–831. [Google Scholar] [CrossRef]
- Serra, V.; von Zglinicki, T.; Lorenz, M.; Saretzki, G. Extracellular superoxide dismutase is a major antioxidant in human fibroblasts and slows telomere shortening. J. Biol. Chem. 2003, 278, 6824–6830. [Google Scholar] [CrossRef] [PubMed]
- Simon, F.; Fernandez, R. Early lipopolysaccharide-induced reactive oxygen species production evokes necrotic cell death in human umbilical vein endothelial cells. J. Hypertens. 2009, 27, 1202–1216. [Google Scholar] [CrossRef]
- Mammone, T.; Gan, D.; Foyouzi-Youssefi, R. Apoptotic cell death increases with senescence in normal human dermal fibroblast cultures. Cell Biol. Int. 2006, 30, 903–909. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.D.; Joiner, C.S.; Stoddart, J.F. Template-directed synthesis employing reversible imine bond formation. Chem. Soc. Rev. 2007, 36, 1705–1723. [Google Scholar] [CrossRef]
- Erdem, A.; Darabi, M.A.; Nasiri, R.; Sangabathuni, S.; Ertas, Y.N.; Alem, H.; Hosseini, V.; Shamloo, A.; Nasr, A.S.; Ahadian, S.; et al. 3D Bioprinting of Oxygenated Cell-Laden Gelatin Methacryloyl Constructs. Adv. Healthc. Mater. 2020, 9, 1901794. [Google Scholar] [CrossRef] [PubMed]
- Mollica, G.; Ziarelli, F.; Lack, S.; Brunel, F.; Viel, S. Characterization of insoluble calcium alginates by solid-state NMR. Carbohyd. Polym. 2012, 87, 383–391. [Google Scholar] [CrossRef]
- Cicha, I.; Goppelt-Struebe, M.; Muehlich, S.; Yilmaz, A.; Raaz, D.; Daniel, W.G.; Garlichs, C.D. Pharmacological inhibition of RhoA signaling prevents connective tissue growth factor induction in endothelial cells exposed to non-uniform shear stress. Atherosclerosis 2008, 196, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Daum, S.; Reshetnikov, M.S.V.; Sisa, M.; Dumych, T.; Lootsik, M.D.; Bilyy, R.; Bila, E.; Janko, C.; Alexiou, C.; Herrmann, M.; et al. Lysosome-Targeting Amplifiers of Reactive Oxygen Species as Anticancer Prodrugs. Angew. Chem. Int. Ed. 2017, 56, 15545–15549. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Genç, H.; Hazur, J.; Karakaya, E.; Dietel, B.; Bider, F.; Groll, J.; Alexiou, C.; Boccaccini, A.R.; Detsch, R.; Cicha, I. Differential Responses to Bioink-Induced Oxidative Stress in Endothelial Cells and Fibroblasts. Int. J. Mol. Sci. 2021, 22, 2358. https://doi.org/10.3390/ijms22052358
Genç H, Hazur J, Karakaya E, Dietel B, Bider F, Groll J, Alexiou C, Boccaccini AR, Detsch R, Cicha I. Differential Responses to Bioink-Induced Oxidative Stress in Endothelial Cells and Fibroblasts. International Journal of Molecular Sciences. 2021; 22(5):2358. https://doi.org/10.3390/ijms22052358
Chicago/Turabian StyleGenç, Hatice, Jonas Hazur, Emine Karakaya, Barbara Dietel, Faina Bider, Jürgen Groll, Christoph Alexiou, Aldo R. Boccaccini, Rainer Detsch, and Iwona Cicha. 2021. "Differential Responses to Bioink-Induced Oxidative Stress in Endothelial Cells and Fibroblasts" International Journal of Molecular Sciences 22, no. 5: 2358. https://doi.org/10.3390/ijms22052358
APA StyleGenç, H., Hazur, J., Karakaya, E., Dietel, B., Bider, F., Groll, J., Alexiou, C., Boccaccini, A. R., Detsch, R., & Cicha, I. (2021). Differential Responses to Bioink-Induced Oxidative Stress in Endothelial Cells and Fibroblasts. International Journal of Molecular Sciences, 22(5), 2358. https://doi.org/10.3390/ijms22052358