
Role of the NUDT Enzymes in Breast Cancer


Abstract
1. Introduction
1.1. Metastatic Breast Cancer
1.2. Use of 3D Cell Culture as a Model for Metastatic Breast Cancer
2. Role of NUDT Enzymes in Cancer
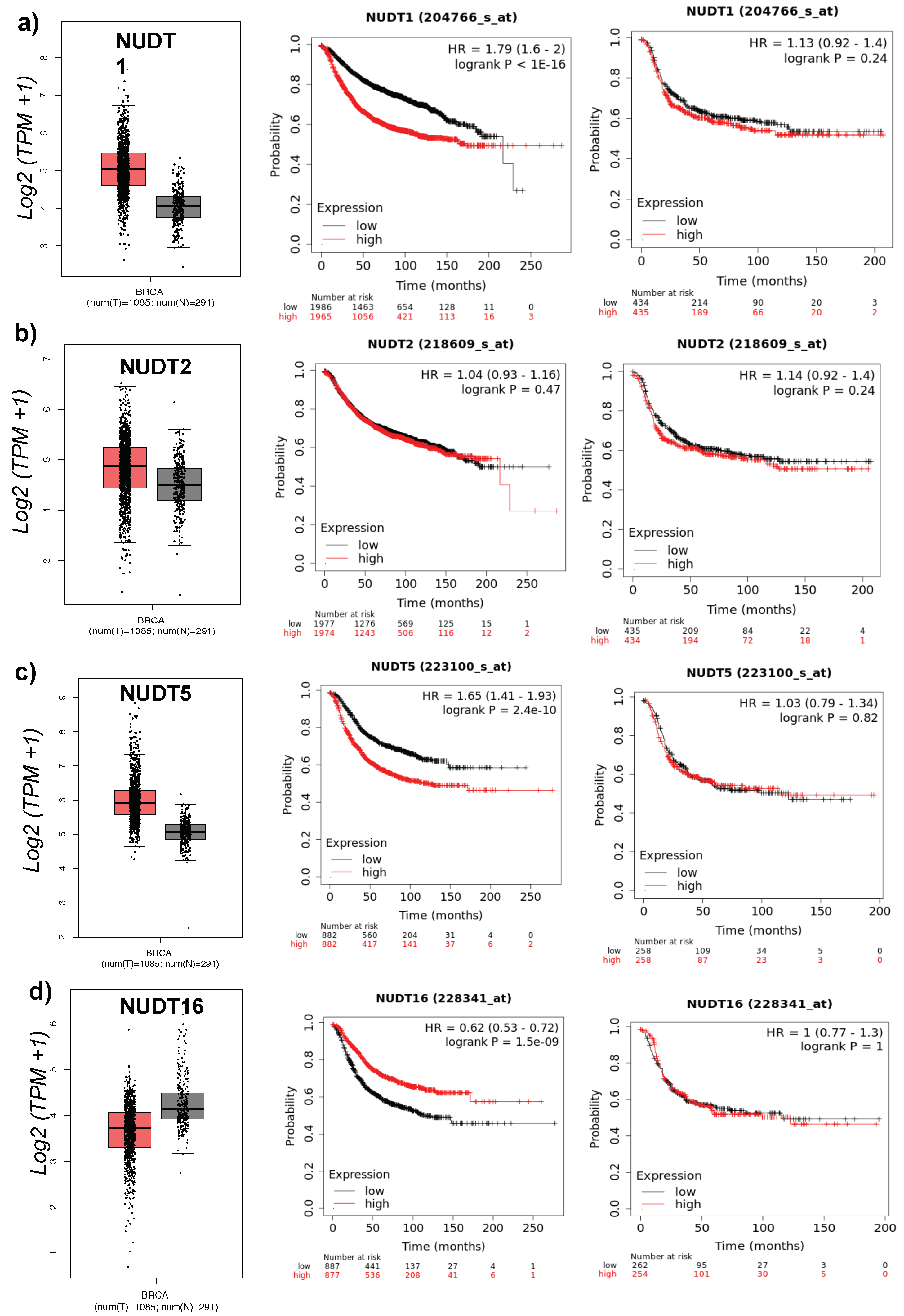
2.1. Role of NUDT Enzymes in Breast Cancer
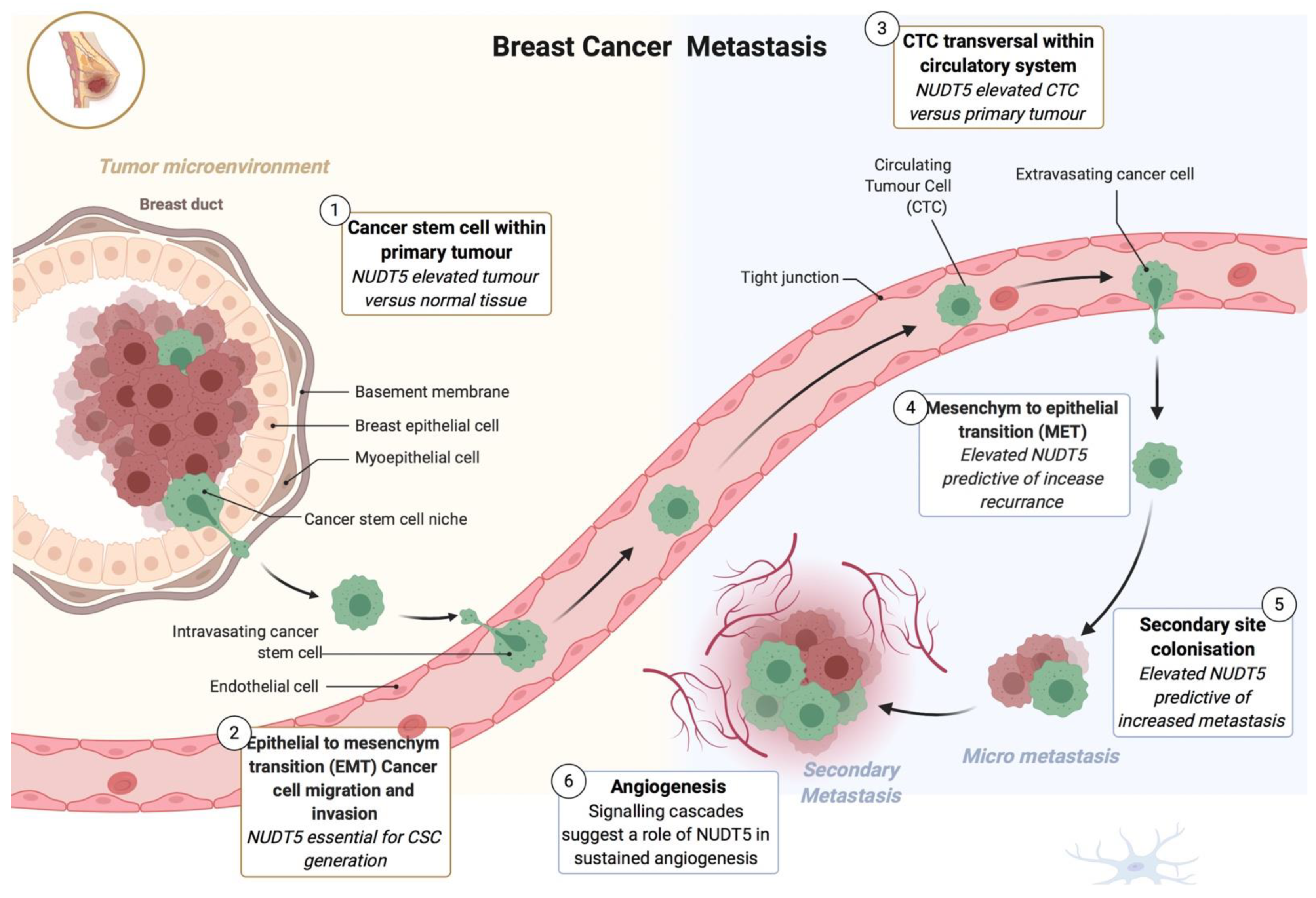
2.2. Role of NUDT5 in Breast Cancer Stem Cells (BCSC)
2.3. A Specific Antibody to the Hexameric form of NUDT5 as a Tool for Breast Cancer Stratification
2.4. Role of NUDT Enzymes in Other Cancer Types
3. Exploiting Global Signaling and Omic Data to Discover Novel Therapeutic Strategies
Progesterone Signaling in Breast Cancer Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ARDT1 | ADP-ribosyltransferase diphtheria toxin-like 1 |
| BCSC | Breast Cancer Stem Cell |
| DCIS | Ductal carcinoma in situ |
| DFS | Disease-free survival |
| ER | Estrogen Receptor |
| H1 | Histone H1 |
| H2A/B | Histone H2A/B |
| HER2 | Human epidermal growth factor receptor 2 |
| HR+ | Hormone Receptor-positive |
| HR- | Hormone Receptor-negative |
| IDC | Invasive ductal carcinoma |
| ILC | Invasive lobular carcinoma |
| NUDT | Nucleoside diphosphate linked moiety X |
| OS | Overall survival |
| PAR | Poly-ADP-ribose |
| PARG | Poly(ADP-ribose) glycohydrolase |
| PARP1 | Poly-ADP-ribose polymerase 1 |
| PR | Progesterone Receptor |
| RFS | Recurrence-free survival |
| SERM | Selective estrogen receptor modulators |
| TGCA | The Cancer Genome Altas |
References
- National Cancer Institute. SEER Stat Fact Sheets: Breast Cancer. In National Cancer Institute Surveillance, Epidemiology, and End Results Program; 2017. Available online: https://seer.cancer.gov/ (accessed on 1 January 2021).
- Hortobagyi, G.N.; de la Garza Salazar, J.; Pritchard, K.; Amadori, D.; Haidinger, R.; Hudis, C.A.; Khaled, H.; Liu, M.C.; Martin, M.; Namer, M.; et al. The global breast cancer burden: Variations in epidemiology and survival. Clin. Breast Cancer 2005, 6, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Society, A.C. Breast Cancer Facts & Figures 2019–2020; American Cancer Society, Inc.: Atlanta, GA, USA, 2019. [Google Scholar]
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment: A Review. JAMA 2019, 321, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Moo, T.A.; Sanford, R.; Dang, C.; Morrow, M. Overview of Breast Cancer Therapy. PET Clin. 2018, 13, 339–354. [Google Scholar] [CrossRef] [PubMed]
- McGale, P.; Taylor, C.; Correa, C.; Cutter, D.; Duane, F.; Ewertz, M.; Gray, R.; Peto, R.; Whelan, T.; Darby, S.; et al. Effect of radiotherapy after mastectomy and axillary surgery on 10-year recurrence and 20-year breast cancer mortality: Meta-analysis of individual patient data for 8135 women in 22 randomised trials. Lancet 2014, 383, 2127–2135. [Google Scholar]
- Zhang, X.H.; Giuliano, M.; Trivedi, M.V.; Schiff, R.; Osborne, C.K. Metastasis dormancy in estrogen receptor-positive breast cancer. Clin. Cancer Res. 2013, 19, 6389–6397. [Google Scholar] [CrossRef]
- Lee, S.B.; Sohn, G.; Kim, J.; Chung, I.Y.; Lee, J.W.; Kim, H.J.; Ko, B.S.; Son, B.H.; Ahn, S.-H. Survival analysis according to period and analysis of the factors influencing changes in survival in patients with recurrent breast cancer: A large-scale, single-center study. Breast Cancer 2018, 25, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Ignatiadis, M.L.; Sotiriou, C. Luminal breast cancer: From biology to treatment. Nat. Rev. Clin. Oncol. 2013, 10, 494–506. [Google Scholar] [CrossRef] [PubMed]
- Perez, E.A. Treatment strategies for advanced hormone receptor-positive and human epidermal growth factor 2-negative breast cancer: The role of treatment order. Drug Resist. Updat. 2016, 24, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.D.; Wang, L.; Kabatas, S.; Ulrich, H.; Zafonte, R.D. Cancer Stem Cells or Tumor Survival Cells? Stem Cells Dev. 2018, 27, 1466–1478. [Google Scholar] [CrossRef]
- Palomeras, S.; Ruiz-Martínez, S.; Puig, T. Targeting Breast Cancer Stem Cells to Overcome Treatment Resistance. Molecules 2018, 23, 2193. [Google Scholar] [CrossRef] [PubMed]
- Schmid, R.; Schmidt, S.K.; Hazur, J.; Detsch, R.; Maurer, E.; Boccaccini, A.R.; Hauptstein, J.; Teßmar, J.; Blunk, T.; Schrüfer, S.; et al. Comparison of Hydrogels for the Development of Well-Defined 3D Cancer Models of Breast Cancer and Melanoma. Cancers 2020, 12, 2320. [Google Scholar] [CrossRef]
- Vinci, M.; Gowan, S.; Boxall, F.; Patterson, L.; Zimmermann, M.; Court, W.; Lomas, C.; Mendiola, M.; Hardisson, D.; Eccles, S.A. Advances in establishment and analysis of three-dimensional tumor spheroid-based functional assays for target validation and drug evaluation. BMC Biol. 2012, 10, 29. [Google Scholar] [CrossRef] [PubMed]
- Kapałczyńska, M.; Kolenda, T.; Przybyła, W.; Zajączkowska, M.; Teresiak, A.; Filas, V.; Ibbs, M.; Bliźniak, R.; Łuczewski, Ł.; Lamperska, K. 2D and 3D cell cultures—A comparison of different types of cancer cell cultures. Arch. Med. Sci. 2018, 14, 910–919. [Google Scholar]
- Wicha, M.S. Targeting breast cancer stem cells. Breast 2009, 18 (Suppl. 3), S56–S58. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, Z.; Dong, D.-L.; Jang, T.-S.; Knowles, J.C.; Kim, H.-W.; Jin, G.-Z.; Xuan, Y. 3D culture technologies of cancer stem cells: Promising ex vivo tumor models. J. Tissue Eng. 2020, 11, 2041731420933407. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, T.; Ohata, H.; Sato, A.; Yamawaki, K.; Enomoto, T.; Okamoto, K. Tumor-derived spheroids: Relevance to cancer stem cells and clinical applications. Cancer Sci. 2017, 108, 283–289. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Xiong, L.; Li, Q.; Lin, L.; Miao, X.; Yan, S.; Hong, Z.; Yang, L.; Wen, Y.; Deng, X. 3D modeling of cancer stem cell niche. Oncotarget 2018, 9, 1326–1345. [Google Scholar] [CrossRef]
- Edmondson, R.; Broglie, J.J.; Adcock, A.F.; Yang, L. Three-dimensional cell culture systems and their applications in drug discovery and cell-based biosensors. Assay Drug Dev. Technol. 2014, 12, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Fontoura, J.C.; Viezzer, C.; Dos Santos, F.G.; Ligabue, R.A.; Weinlich, R.; Puga, R.D.; Antonow, D.; Severino, P.; Bonorino, C. Comparison of 2D and 3D cell culture models for cell growth, gene expression and drug resistance. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 107, 110264. [Google Scholar] [CrossRef]
- Riedl, A.; Schlederer, M.; Pudelko, K.; Stadler, M.; Walter, S.; Unterleuthner, D.; Unger, C.; Kramer, N.; Hengstschläger, M.; Kenner, L.; et al. Comparison of cancer cells in 2D vs 3D culture reveals differences in AKT-mTOR-S6K signaling and drug responses. J. Cell Sci. 2017, 130, 203–218. [Google Scholar] [CrossRef]
- Imamura, Y.; Mukohara, T.; Shimono, Y.; Funakoshi, Y.; Chayahara, N.; Toyoda, M.; Kiyota, N.; Takao, S.; Kono, S.; Nakatsura, T.; et al. Comparison of 2D- and 3D-culture models as drug-testing platforms in breast cancer. Oncol. Rep. 2015, 33, 1837–1843. [Google Scholar] [CrossRef]
- Wright, R.H.; Lioutas, A.; Le Dily, F.; Soronellas, D.; Pohl, A.; Bonet, J.; Nacht, A.S.; Samino, S.; Font-Mateu, J.; Vicent, G.P.; et al. ADP-ribose-derived nuclear ATP synthesis by NUDIX5 is required for chromatin remodeling. Science 2016, 352, 1221–1225. [Google Scholar] [CrossRef] [PubMed]
- Pickup, K.E.; Pardow, F.; Carbonell-Caballero, J.; Lioutas, A.; Villanueva-Cañas, J.L.; Wright, R.H.G.; Beato, M. Expression of Oncogenic Drivers in 3D Cell Culture Depends on Nuclear ATP Synthesis by NUDT5. Cancers 2019, 11, 1337. [Google Scholar] [CrossRef] [PubMed]
- Kulikova, V.A.; Nikiforov, A.A. Role of NUDIX Hydrolases in NAD and ADP-Ribose Metabolism in Mammals. Biochemistry 2020, 85, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Carreras-Puigvert, J.; Zitnik, M.; Jemth, A.S.; Carter, M.; Unterlass, J.E.; Hallström, B.; Loseva, O.; Karem, Z.; Calderón-Montaño, J.M.; Lindskog, C.; et al. A comprehensive structural, biochemical and biological profiling of the human NUDIX hydrolase family. Nat. Commun. 2017, 8, 1541. [Google Scholar] [CrossRef] [PubMed]
- McLennan, A.G.; Cartwright, J.L.; Gasmi, L. The human NUDT family of nucleotide hydrolases. Enzymes of diverse substrate specificity. Adv. Exp. Med. Biol. 2000, 486, 115–118. [Google Scholar] [PubMed]
- Zhang, X.; Song, W.; Zhou, Y.; Mao, F.; Lin, Y.; Guan, J.; Sun, Q. Expression and function of MutT homolog 1 in distinct subtypes of breast cancer. Oncol. Lett. 2017, 13, 2161–2168. [Google Scholar] [CrossRef] [PubMed]
- Oka, K.; Suzuki, T.; Onodera, Y.; Miki, Y.; Takagi, K.; Nagasaki, S.; Akahira, J.-I.; Ishida, T.; Watanabe, M.; Hirakawa, H.; et al. Nudix-type motif 2 in human breast carcinoma: A potent prognostic factor associated with cell proliferation. Int. J. Cancer 2011, 128, 1770–1782. [Google Scholar] [CrossRef]
- Marriott, A.S.; Vasieva, O.; Fang, Y.; Copeland, N.A.; McLennan, A.G.; Jones, N.J. NUDT2 Disruption Elevates Diadenosine Tetraphosphate (Ap4A) and Down-Regulates Immune Response and Cancer Promotion Genes. PLoS ONE 2016, 11, e0154674. [Google Scholar] [CrossRef] [PubMed]
- Harvell, D.M.; Richer, J.K.; Allred, D.C.; Sartorius, C.A.; Horwitz, K.B. Estradiol regulates different genes in human breast tumor xenografts compared with the identical cells in culture. Endocrinology 2006, 147, 700–713. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, B.J.; Liu, D.C.; Guo, Q.Y.; Han, X.W.; Bi, X.M.; Wang, H.; Wu, Z.-S.; Wu, W.-Y. NUDT21 Suppresses Breast Cancer Tumorigenesis Through Regulating CPSF6 Expression. Cancer Manag. Res. 2020, 12, 3069–3078. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.H.; Castellano, G.; Bonet, J.; Le Dily, F.; Font-Mateu, J.; Ballare, C.; Nacht, A.S.; Soronellas, D.; Oliva, B.; Beato, M. CDK2-dependent activation of PARP-1 is required for hormonal gene regulation in breast cancer cells. Genes Dev. 2012, 26, 1972–1983. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.H.; Beato, M. PARty promoters: Hormone-dependent gene regulation requires CDK2 activation of PARP1. Cell Cycle 2012, 11, 4291–4293. [Google Scholar] [CrossRef][Green Version]
- Poirier, G.G.; Niedergang, C.; Champagne, M.; Mazen, A.; Mandel, P. Adenosine diphosphate ribosylation of chicken-erythrocyte histones H1, H5 and high-mobility-group proteins by purified calf-thymus poly(adenosinediphosphate-ribose) polymerase. Eur. J. Biochem. 1982, 127, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Poirier, G.G.; De Murcia, G.; Jongstra-Bilen, J.; Niedergang, C.; Mandel, P. Poly(ADP-ribosyl)ation of polynucleosomes causes relaxation of chromatin structure. Proc. Natl. Acad. Sci. USA 1982, 79, 3423–3427. [Google Scholar] [CrossRef]
- Aubin, R.J.; Dam, V.T.; Miclette, J.; Brousseau, Y. Poirier GG. Chromosomal protein poly(ADP-ribosyl)ation in pancreatic nucleosomes. Can. J. Biochem. 1982, 60, 295–305. [Google Scholar] [CrossRef]
- Aubin, R.J.; Dam, V.T.; Miclette, J.; Brousseau, Y.; Huletsky, A.; Poirier, G.G. Hyper(ADP-ribosyl)ation of histone H1. Can. J. Biochem. 1982, 60, 1085–1094. [Google Scholar] [CrossRef]
- Mathis, G.; Althaus, F.R. Release of core DNA from nucleosomal core particles following (ADP-ribose)n-modification in vitro. Biochem. Biophys. Res. Commun. 1987, 143, 1049–1054. [Google Scholar] [CrossRef]
- Kim, M.Y.; Mauro, S.; Gévry, N.; Lis, J.T.; Kraus, W. NAD+-dependent modulation of chromatin structure and transcription by nucleosome binding properties of PARP-1. Cell 2004, 119, 803–814. [Google Scholar] [CrossRef]
- Kraus, W.; Lis, J.T. PARP goes transcription. Cell 2003, 113, 677–683. [Google Scholar] [CrossRef]
- Thomas, C.; Tulin, A.V. Poly-ADP-ribose polymerase: Machinery for nuclear processes. Mol. Aspects Med. 2013, 34, 1124–1137. [Google Scholar] [CrossRef]
- Tulin, A.; Stewart, D.; Spradling, A.C. The Drosophila heterochromatic gene encoding poly(ADP-ribose) polymerase (PARP) is required to modulate chromatin structure during development. Genes Dev. 2002, 16, 2108–2119. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.H.; Fernandez-Fuentes, N.; Oliva, B.; Beato, M. Insight into the machinery that oils chromatin dynamics. Nucleus 2016, 7, 532–539. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Maruta, H.; Matsumura, N.; Tanuma, S. Role of (ADP-ribose)n catabolism in DNA repair. Biochem. Biophys. Res. Commun. 1997, 236, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Zha, M.; Guo, Q.; Zhang, Y.; Yu, B.; Ou, Y.; Zhong, C.; Ding, J. Molecular mechanism of ADP-ribose hydrolysis by human NUDT5 from structural and kinetic studies. J. Mol. Biol. 2008, 379, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Zha, M.; Zhong, C.; Peng, Y.; Hu, H.; Ding, J. Crystal structures of human NUDT5 reveal insights into the structural basis of the substrate specificity. J. Mol. Biol. 2006, 364, 1021–1033. [Google Scholar] [CrossRef]
- Jing, X.; Liang, H.; Hao, C.; Yang, X.; Cui, X. Overexpression of MUC1 predicts poor prognosis in patients with breast cancer. Oncol. Rep. 2019, 41, 801–810. [Google Scholar] [CrossRef]
- Ballesta, A.M.; Molina, R.; Filella, X.; Jo, J.; Gimenez, N. Carcinoembryonic antigen in staging and follow-up of patients with solid tumors. Tumour. Biol. 1995, 16, 32–41. [Google Scholar] [CrossRef]
- Powell, E.; Shao, J.; Picon, H.M.; Bristow, C.; Ge, Z.; Peoples, M.; Robinson, F.; Jeter-Jones, S.L.; Schlosberg, C.; Grzeskowiak, C.L.; et al. A functional genomic screen in vivo identifies CEACAM5 as a clinically relevant driver of breast cancer metastasis. NPJ Breast Cancer 2018, 4, 9. [Google Scholar] [CrossRef]
- Pudelko, L.; Rouhi, P.; Sanjiv, K.; Gad, H.; Kalderen, C.; Hoglund, A.; Squatrito, M.; Schuhmacher, A.J.; Edwards, S.; Hägerstrand, D.; et al. Glioblastoma and glioblastoma stem cells are dependent on functional MTH1. Oncotarget 2017, 8, 84671–84684. [Google Scholar] [CrossRef]
- Wang, J.Y.; Liu, G.Z.; Wilmott, J.S.; La, T.; Feng, Y.C.; Yari, H.; Yan, X.G.; Thorne, R.F.; Scolyer, R.A.; Zhang, X.D.; et al. Skp2-Mediated Stabilization of MTH1 Promotes Survival of Melanoma Cells upon Oxidative Stress. Cancer Res. 2017, 77, 6226–6239. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Ma, L.; Yang, J.; Qiao, H.; Li, L.; Guo, Q.; Ma, J.; Zhao, L.; Wang, J.; Jiang, G.; et al. Potent and specific MTH1 inhibitors targeting gastric cancer. Cell Death Dis. 2019, 10, 434. [Google Scholar] [CrossRef]
- Anadón, C.; Van Tetering, G.; Ferreira, H.J.; Moutinho, C.; Martínez-Cardús, A.; Villanueva, A.; Soler, M.; Heyn, H.; Moran, S.; De Moura, M.C.; et al. Epigenetic loss of the RNA decapping enzyme NUDT16 mediates C-MYC activation in T-cell acute lymphoblastic leukemia. Leukemia 2017, 31, 1622–1625. [Google Scholar] [CrossRef][Green Version]
- Iyama, T.; Abolhassani, N.; Tsuchimoto, D.; Nonaka, M.; Nakabeppu, Y. NUDT16 is a (deoxy)inosine diphosphatase, and its deficiency induces accumulation of single-strand breaks in nuclear DNA and growth arrest. Nucleic Acids Res. 2010, 38, 4834–4843. [Google Scholar] [CrossRef]
- Wang, Y.; Wan, F.; Chang, K.; Lu, X.; Dai, B.; Ye, D. NUDT expression is predictive of prognosis in patients with clear cell renal cell carcinoma. Oncol. Lett. 2017, 14, 6121–6128. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Liu, T.H.; Li, J.; Li, D.N.; Tian, X.Y.; Ouyang, Q.G.; Cai, J.-P. The high expression of MTH1 and NUDT5 predict a poor survival and are associated with malignancy of esophageal squamous cell carcinoma. PeerJ 2020, 8, e9195. [Google Scholar] [CrossRef] [PubMed]
- Kwon, O.; Kwak, D.; Ha, S.H.; Jeon, H.; Park, M.; Chang, Y.; Suh, P.-G.; Ryu, S.H. Nudix-type motif 2 contributes to cancer proliferation through the regulation of Rag GTPase-mediated mammalian target of rapamycin complex 1 localization. Cell Signal. 2017, 32, 24–35. [Google Scholar] [CrossRef]
- Zhang, F.; Lou, L.; Peng, B.; Song, X.; Reizes, O.; Almasan, A.; Gong, Z. Nudix Hydrolase NUDT16 Regulates 53BP1 Protein by Reversing 53BP1 ADP-Ribosylation. Cancer Res. 2020, 80, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- Gyorffy, B.; Serra, V.; Jurchott, K.; Abdul-Ghani, R.; Garber, M.; Stein, U.; Petersen, I.; Lage, H.; Dietel, M.; Schäfer, R. Prediction of doxorubicin sensitivity in breast tumors based on gene expression profiles of drug-resistant cell lines correlates with patient survival. Oncogene 2005, 24, 7542–7551. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Park, S.; Oh, J.; Kim, D.; Ryu, J.H.; Park, W.C.; Baek, I.-J.; Cheng, X.; Lu, X.; Jin, E.-J. NUDT7 Loss Promotes Kras(G12D) CRC Development. Cancers 2020, 12, 576. [Google Scholar] [CrossRef]
- Barupal, D.K.; Gao, B.; Budczies, J.; Phinney, B.S.; Perroud, B.; Denkert, C.; Fiehn, O. Prioritization of metabolic genes as novel therapeutic targets in estrogen-receptor negative breast tumors using multi-omics data and text mining. Oncotarget 2019, 10, 3894–3909. [Google Scholar] [CrossRef]
- Wright, R.H.G.; Vastolo, V.; Oliete, J.Q.; Carbonell-Caballero, J.; Beato, M. Signalling Network of Breast Cancer Cells in Response to Progesterone. BioRxiv 2020. [Google Scholar] [CrossRef]
- Reyes, D.; Ballare, C.; Castellano, G.; Soronellas, D.; Bagó, J.R.; Blanco, J.; Beato, M. Activation of mitogen- and stress-activated kinase 1 is required for proliferation of breast cancer cells in response to estrogens or progestins. Oncogene 2014, 33, 1570–1580. [Google Scholar] [CrossRef] [PubMed]
- Ballare, C.; Uhrig, M.; Bechtold, T.; Sancho, E.; Di Domenico, M.; Migliaccio, A.; Auricchio, F.; Beato, M. Two domains of the progesterone receptor interact with the estrogen receptor and are required for progesterone activation of the c-Src/Erk pathway in mammalian cells. Mol. Cell Biol. 2003, 23, 1994–2008. [Google Scholar] [CrossRef] [PubMed]
- Moukengue, B.; Brown, H.K.; Charrier, C.; Battaglia, S.; Baud’Huin, M.; Quillard, T.; Pham, T.M.; Pateras, I.; Gorgoulis, V.G.; Helleday, T.; et al. TH1579, MTH1 inhibitor, delays tumour growth and inhibits metastases development in osteosarcoma model. EBioMedicine 2020, 53, 102704. [Google Scholar] [CrossRef]
- Gad, H.; Koolmeister, T.; Jemth, A.S.; Eshtad, S.; Jacques, S.A.; Ström, C.E.; Svensson, L.M.; Schultz, N.; Lundbäck, T.; Einarsdottir, B.O.; et al. MTH1 inhibition eradicates cancer by preventing sanitation of the dNTP pool. Nature 2014, 508, 215–221. [Google Scholar] [CrossRef]
- Van der Waals, L.M.; Laoukili, J.; Jongen, J.M.J.; Raats, D.A.; Borel Rinkes, I.H.M.; Kranenburg, O. Differential anti-tumour effects of MTH1 inhibitors in patient-derived 3D colorectal cancer cultures. Sci. Rep. 2019, 9, 819. [Google Scholar] [CrossRef]
- Farand, J.; Kropf, J.E.; Blomgren, P.; Xu, J.; Schmitt, A.C.; Newby, Z.E.; Wang, T.; Murakami, E.; Barauskas, O.; Sudhamsu, J.; et al. Discovery of Potent and Selective MTH1 Inhibitors for Oncology: Enabling Rapid Target (In)Validation. ACS Med. Chem. Lett. 2020, 11, 358–364. [Google Scholar] [CrossRef]
- Page, B.D.G.; Valerie, N.C.K.; Wright, R.H.G.; Wallner, O.; Isaksson, R.; Carter, M.; Rudd, S.G.; Loseva, O.; Jemth, A.-S.; Almlöf, I.; et al. Targeted NUDT5 inhibitors block hormone signaling in breast cancer cells. Nat. Commun. 2018, 9, 250. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NUDT (Alternative Name) | Substrate Hydrolase Activity | Product Hydrolase Activity | Role | Enzyme Entry (Expasy) |
|---|---|---|---|---|
| NUDT1 (MTH1) | 8-oxo-dGTP+ H2O 8-oxo-dATP 8-OH-dATP | 8-oxo-dGMP +diphosphate+H+ | Sanitizing oxidized nucleotides | EC 3.6.1.55 EC 3.6.1.56 |
| NUDT2 (APAH1) | Ap4A | AMP + ATP | Homeostasis | EC 3.6.17 |
| NUDT3 (DIPP-1) | Diphospho-myo-inositol polyphosphate + H2O | myo-inositol polyphosphate + phosphate. | EC 3.6.1.52 | |
| NUDT4 (DIPP-2) | Diphospho-myo-inositol polyphosphate + H2O | myo-inositol polyphosphate + phosphate. | Signal transduction | EC:3.6.1.52 |
| NUDT5 (NUDIX5) | ATP + D-ribose 5-phosphate + H+ ADP-D-ribose + H2O 8-oxo-dGDP + H2O ADP-D-ribose + PPi + H2O | ADP-D-ribose + diphosphate AMP + D-ribose 5-phosphate + 2 H+ 8-oxo-dGMP + H+ + phosphate ATP+ D-ribose-5-phosphate | Sanitizing oxidized nucleotides | EC 2.7.7.96 EC 3.6.1.13 EC 3.6.1.58 |
| NUDT6 (FGF2AS) | ADP-ribose + H2O NADH + H2O | AMP + D-ribose 5-phosphate. AMP + NMNH + 2 H+ | Cell proliferation | EC 2.7.7.96 |
| NUDT7 | acetyl-CoA + H2O | acetate + CoA + H(+). | Eliminate oxidized coenzyme A (CoA) | EC 3.1.2.20 |
| NUDT8 | unknown | unknown | unknown | |
| NUDT9 | ADP-D-ribose + H2O | AMP + D-ribose 5-phosphate + 2 H+ | EC 3.6.1.13 | |
| NUDT10 (DIPP3A, APS2) | H2O + P1,P6-bis(5′-adenosyl) hexaphosphate H2O + P1,P5-bis(5′-adenosyl) pentaphosphate Diphospho-myo-inositol polyphosphate + H2O | adenosine 5′-pentaphosphate + AMP + 2 H+ adenosine 5′-tetraphosphate + AMP + 2 H+ myo-inositol polyphosphate + phosphate | Signal transduction | EC 3.6.1.60 EC 3.6.1.60 EC 3.6.1.52 |
| NUDT (Alternative Name) | Substrate Hydrolase Activity | Product Hydrolase Activity | Role | Enzyme Entry (Expasy) |
| NUDT11 (DIPP3B, APS1) | H2O + P1,P6-bis(5′-adenosyl) hexaphosphate H2O + P1,P5-bis(5′-adenosyl) pentaphosphate Diphospho-myo-inositol polyphosphate + H2O | adenosine 5′-pentaphosphate + AMP + 2 H+ adenosine 5′-tetraphosphate + AMP + 2 H+ myo-inositol polyphosphate + phosphate | Signal transduction | EC 3.6.1.60 EC 3.6.1.60 EC 3.6.1.52 |
| NUDT12 | H2O + NAD+ H2O + NADH | AMP + β-nicotinamide D-ribonucleotide + 2 H+ AMP + 2 H+ + reduced β-nicotinamide D-ribonucleotide | Regulate nicotinamide | EC 3.6.1.22 EC 3.6.1.22 |
| NUDT13 | unknown | unknown | unknown | |
| NUDT14 (UGPP) | UDP-sugar + H2O | UMP + alpha-D-aldose 1-phosphate | unknown | EC:3.6.1.45 |
| NUDT15 (MTH2) | a ribonucleoside 5′-triphosphate + H2O a 2′-deoxyribonucleoside 5′-triphosphate + H2O | a ribonucleoside 5′-phosphate + diphosphate + H+ a 2′-deoxyribonucleoside 5′-phosphate + diphosphate + H+ | Sanitizing oxidized nucleotides | EC:3.6.1.9 |
| NUDT16 | a 5′-end (N7-methyl 5′-triphosphoguanosine)-adenosine in mRNA + H2O | a 5′-end phospho-adenosine in mRNA + 2 H+ + N7-methylguanosine 5′-diphosphate | RNA decapping enzyme | EC 3.6.1.62 |
| NUDT17 | unknown | unknown | unknown | unknown |
| NUDT18 (MTH3) | 8-oxo-dGDP + H2O | 8-oxo-dGMP + H+ + phosphate | Removes oxidized guanine from DNA and RNA | EC 3.6.1.58 |
| NUDT19 | acyl-CoA + H2O | CoA + a carboxylate. | Hydrolysis CoA esters | EC 3.1.2.20 |
| NUDT20 (DCP2) | a 5′-end (N7-methyl 5′-triphosphoguanosine)-adenosine in mRNA+ H2O | a 5′-end phospho-adenosine in mRNA + 2 H+ + N7-methylguanosine 5′-diphosphate | RNA decapping enzyme | EC 3.6.1.62 |
| NUDT21 (CFIm25) | No hydrolase activity | No hydrolase activity | Pre-mRNA processing |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wright, R.H.G.; Beato, M. Role of the NUDT Enzymes in Breast Cancer. Int. J. Mol. Sci. 2021, 22, 2267. https://doi.org/10.3390/ijms22052267
Wright RHG, Beato M. Role of the NUDT Enzymes in Breast Cancer. International Journal of Molecular Sciences. 2021; 22(5):2267. https://doi.org/10.3390/ijms22052267
Chicago/Turabian StyleWright, Roni H. G., and Miguel Beato. 2021. "Role of the NUDT Enzymes in Breast Cancer" International Journal of Molecular Sciences 22, no. 5: 2267. https://doi.org/10.3390/ijms22052267
APA StyleWright, R. H. G., & Beato, M. (2021). Role of the NUDT Enzymes in Breast Cancer. International Journal of Molecular Sciences, 22(5), 2267. https://doi.org/10.3390/ijms22052267