
Drosophila Homeodomain-Interacting Protein Kinase (Hipk) Phosphorylates the Hippo/Warts Signalling Effector Yorkie


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
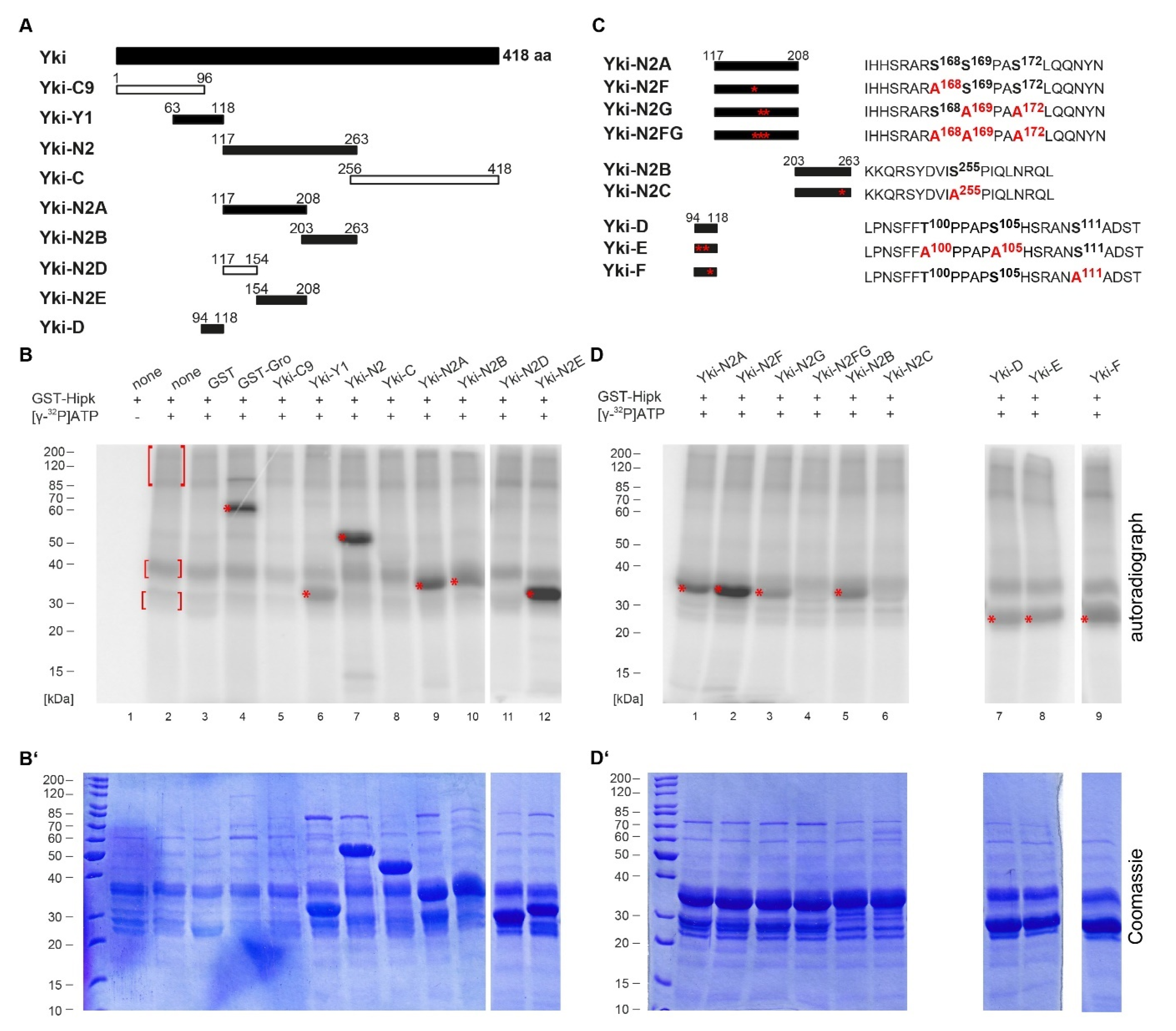
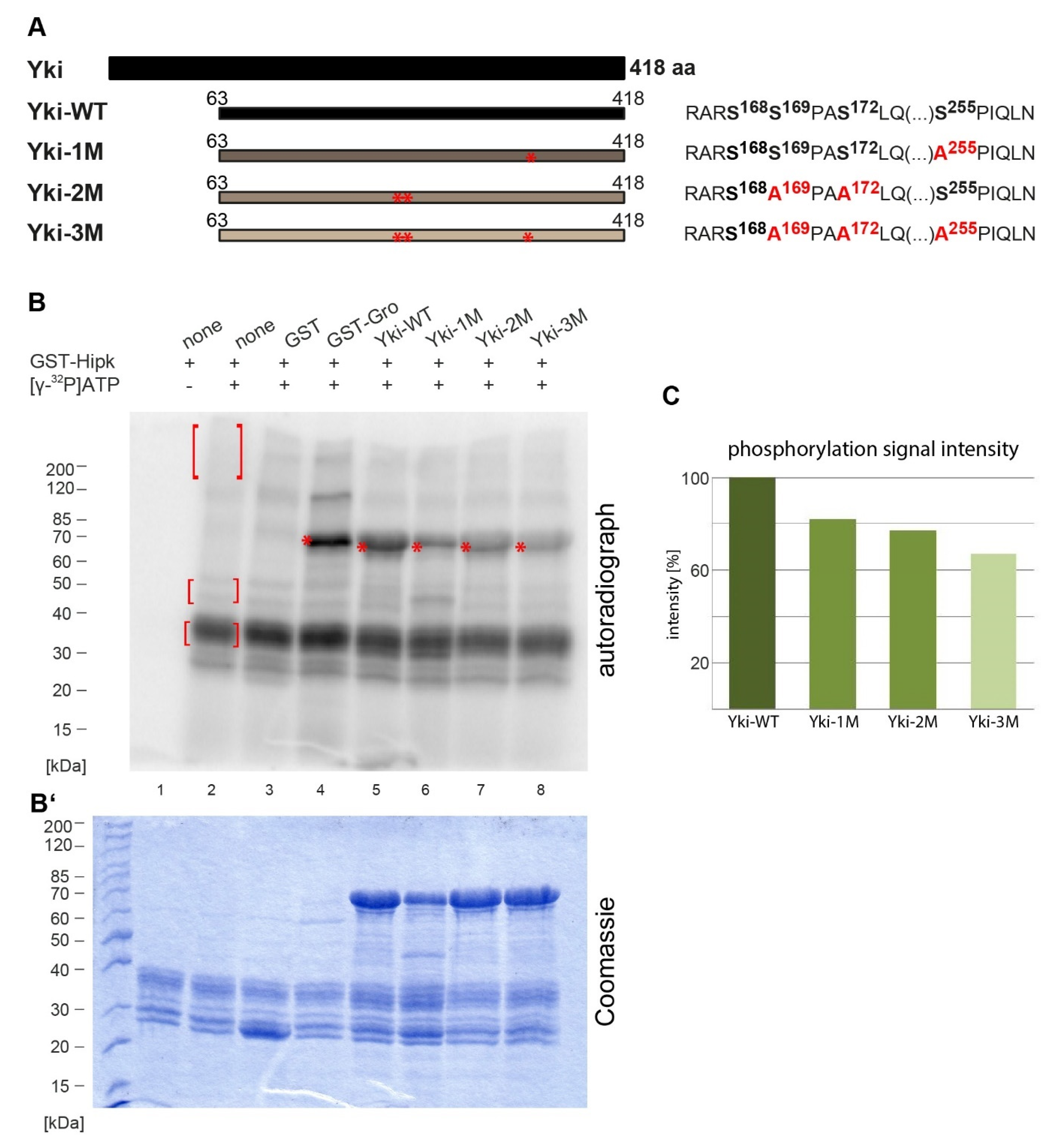
2.1. Hipk Phosphorylates Yorkie at Several Sites
2.2. Hipk Phosphorylation Regulates Yorkie In Vivo
3. Discussion
4. Materials and Methods
4.1. Purification of Recombinant Proteins
4.2. In Vitro Mutagenesis
4.3. In Vitro Phosphorylation Assay
4.4. Fly Stocks
4.5. Phenotypical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| HIPK/Hipk | Homeodomain-interacting protein kinase |
| Yki | Yorkie, aa amino acids |
References
- Badouel, C.; Garg, A.; McNeill, H. Herding hippos: Regulating growth in flies and man. Curr. Opin. Cell Biol. 2009, 21, 837–843. [Google Scholar] [CrossRef]
- Zheng, Y.; Pan, D. The hippo signaling pathway in development and disease. Dev. Cell. 2019, 50, 264–282. [Google Scholar] [CrossRef] [PubMed]
- Snigdha, K.; Gangwani, K.S.; Lapalikar, G.V.; Singh, A.; Kango-Singh, M. Hippo signaling in cancer: Lessons from Drosophila models. Front. Cell Dev. Biol. 2019, 7, 85. [Google Scholar] [CrossRef] [PubMed]
- Koontz, L.M.; Liu-Chittenden, Y.; Yin, F.; Zheng, Y.; Yu, J.; Huang, B.; Chen, Q.; Wu, S.; Pan, D. The Hippo effector Yorkie controls normal tissue growth by antagonizing scalloped-mediated default repression. Dev. Cell. 2013, 25, 388–401. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Pan, D. Validating upstream regulators of Yorkie activity in Hippo signaling through scalloped-based genetic epistasis. Development 2018, 145, dev157545. [Google Scholar] [CrossRef] [PubMed]
- Mackintosh, C. Dynamic interactions between 14-3-3 proteins and phosphoproteins regulate diverse cellular processes. Biochem. J. 2004, 381, 329–342. [Google Scholar] [CrossRef]
- Huang, J.; Wu, S.; Barrera, J.; Matthews, K.; Pan, D. The Hippo signaling pathway coordinately regulates cell proliferation and apoptosis by inactivating Yorkie, the Drosophila Homolog of YAP. Cell 2005, 122, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Feldmann, G.; Huang, J.; Wu, S.; Zhang, N.; Comerford, S.A.; Gayyed, M.F.; Anders, R.A.; Maitra, A.; Pan, D. Elucidation of a universal size-control mechanism in Drosophila and mammals. Cell 2007, 130, 1120–1133. [Google Scholar] [CrossRef]
- Hao, Y.; Chun, A.; Cheung, K.; Rashidi, B.; Yang, X. Tumor suppressor LATS1 is a negative regulator of oncogene YAP. J. Biol. Chem. 2008, 283, 5496–5509. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Smolen, G.A.; Haber, D.A. Negative regulation of YAP by LATS1 underscores evolutionary conservation of the Drosophila Hippo pathway. Cancer Res. 2008, 68, 2789–2794. [Google Scholar] [CrossRef]
- Zhao, B.; Ye, X.; Yu, J.; Li, L.; Li, W.; Li, S.; Yu, J.; Lin, J.D.; Wang, C.Y.; Chinnaiyan, A.M.; et al. TEAD mediates YAP-dependent gene induction and growth control. Genes Dev. 2008, 22, 1962–1971. [Google Scholar] [CrossRef]
- Oh, H.; Irvine, K.D. In vivo regulation of Yorkie phosphorylation and localization. Development 2008, 135, 1081–1088. [Google Scholar] [CrossRef]
- Oh, H.; Irvine, K.D. In vivo analysis of Yorkie phosphorylation sites. Oncogene 2009, 28, 1916–1927. [Google Scholar] [CrossRef] [PubMed]
- Ren, F.; Zhang, L.; Jiang, J. Hippo signaling regulates Yorkie nuclear localization and activity through 14-3-3 dependent and independent mechanisms. Dev. Biol. 2010, 337, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Wei, X.; Li, W.; Udan, R.S.; Yang, Q.; Kim, J.; Xie, J.; Ikenoue, T.; Yu, J.; Li, L.; et al. Inactivation of YAP oncoprotein by the Hippo pathway is involved in cell contact inhibition and tissue growth control. Genes Dev. 2007, 21, 2747–2761. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Li, L.; Tumaneng, K.; Wang, C.Y.; Guan, K.L. A coordinated phosphorylation by Lats and CK1 regulates YAP stability through SCF (beta-TRCP). Genes Dev. 2010, 24, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Verheyen, E.M. Homeodomain-interacting protein kinase regulates Yorkie activity to promote tissue growth. Curr. Biol. 2012, 22, 1582–1586. [Google Scholar] [CrossRef] [PubMed]
- Poon, C.L.; Zhang, X.; Lin, J.I.; Manning, S.A.; Harvey, K.F. Homeodomain-interacting protein kinase regulates Hippo pathway-dependent tissue growth. Curr. Biol. 2012, 22, 1587–1594. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Choi, C.Y.; Lee, S.-J.; Conti, M.A.; Kim, Y. Homeodomain-interacting protein kinases, a novel family of co- repressors for homeodomain transcription factors. J. Biol. Chem. 1998, 273, 25875–25879. [Google Scholar] [CrossRef]
- Manning, G.; Whyte, D.B.; Martinez, R.; Hunter, T.; Sudarsanam, S. The protein kinase complement of the human genome. Science 2002, 298, 1912–1934. [Google Scholar] [CrossRef]
- Link, N.; Chen, P.; Lu, W.J.; Pogue, K.; Chuong, A.; Mata, M.; Checketts, J.; Abrams, J.M. A collective form of cell death requires homeodomain interacting protein kinase. J. Cell Biol. 2007, 178, 567–574. [Google Scholar] [CrossRef]
- Choi, C.Y.; Kim, Y.H.; Kim, Y.O.; Park, S.J.; Kim, E.A.; Riemenschneider, W.; Gajewski, K.; Schulz, R.A.; Kim, Y. Phosphorylation by the DHIPK2 protein kinase modulates the corepressor activity of Groucho. J. Biol. Chem. 2005, 280, 21427–21436. [Google Scholar] [CrossRef]
- Steinmetz, E.L. Analyse von Wechselwirkungen der Homeodomänen Interagierenden Proteinkinase (Hipk) mit Genen der Augenentwicklung von Drosophila melanogaster. Ph.D. Thesis, Saarland University, Saarbrücken, Germany, 2018. [Google Scholar] [CrossRef]
- Rinaldo, C.; Prodosmo, A.; Siepi, F.; Soddu, S. HIPK2: A multitalented partner for transcription factors in DNA damage response and development. Biochem. Cell Biol. 2007, 85, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Rinaldo, C.; Siepi, F.; Prodosmo, A.; Soddu, S. HIPKs: Jack of all trades in basic nuclear activities. Biochim. Biophys. Acta Mol. Cell Res. 2008, 1783, 2124–2129. [Google Scholar] [CrossRef] [PubMed]
- Kuwano, Y.; Nishida, K.; Akaike, Y.; Kurokawa, K.; Nishikawa, T.; Masuda, K.; Rokutan, K. Homeodomain-interacting protein kinase-2: A critical regulator of the DNA damage response and the epigenome. Int. J. Mol. Sci. 2016, 17, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Swarup, S.; Chen, J.; Ishitani, T.; Verheyen, E.M. Homeodomain-interacting protein kinases (Hipks) promote Wnt/Wg signaling through stabilization of β -catenin / Arm and stimulation of target gene expression. Development 2009, 136, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Andrews, B.C.; Faust, M.; Walldorf, U.; Verheyen, E.M. Hipk is an essential protein that promotes Notch signal transduction in the Drosophila eye by inhibition of the global co-repressor Groucho. Dev. Biol. 2009, 325, 263–272. [Google Scholar] [CrossRef]
- Sudol, M. Newcomers to the WW domain-mediated network of the Hippo tumor suppressor pathway. Genes Cancer. 2010, 1, 1115–1118. [Google Scholar] [CrossRef]
- Kim, E.A.; Noh, Y.T.; Ryu, M.-J.; Kim, H.-T.; Lee, S.-E.; Kim, C.-H.; Lee, C.; Kim, Y.H.; Choi, C.Y. Phosphorylation and transactivation of Pax6 by homeodomain-interacting protein kinase 2. J. Biol. Chem. 2006, 281, 7489–7497. [Google Scholar] [CrossRef]
- Dewald, D.N.; Steinmetz, E.L.; Walldorf, U. Homeodomain-interacting protein kinase (Hipk) phosphorylates the small SPOC family protein Spenito. Insect Mol. Biol. 2014, 23, 706–719. [Google Scholar] [CrossRef]
- Steinmetz, E.L.; Dewald, D.N.; Walldorf, U. Homeodomain-interacting protein kinase phosphorylates the Drosophila Paired box protein 6 (Pax6) homologues Twin of eyeless and Eyeless. Insect Mol. Biol. 2018, 27, 198–211. [Google Scholar] [CrossRef]
- Zhai, B.; Villén, J.; Beausoleil, S.A.; Mintseris, J.; Gygi, S.P. Phosphoproteome analysis of Drosophila melanogaster embryos. J. Proteome Res. 2008, 7, 1675–1682. [Google Scholar] [CrossRef] [PubMed]
- Gnad, F.; Gunawardena, J.; Mann, M. PHOSIDA 2011: The posttranslational modification database. Nucleic Acids Res. 2011, 39, 253–260. [Google Scholar] [CrossRef]
- Seger, R.; Krebs, E.G. The MAPK signaling cascade. FASEB J. 1995, 9, 726–735. [Google Scholar] [CrossRef] [PubMed]
- An, R.; Da Silva, X.G.; Semplici, F.; Vakhshouri, S.; Hao, H.X.; Rutter, J.; Pagano, M.A.; Meggio, F.; Pinna, L.A.; Rutter, G.A. Pancreatic and duodenal homeobox 1 (PDX1) phosphorylation at serine-269 is HIPK2-dependent and affects PDX1 subnuclear localization. Biochem. Biophys. Res. Commun. 2010, 399, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Wang, H.; Liu, S.H.; Shahi, K.M.; Lin, X.; Wu, J.; Feng, X.H.; Qin, J.; Tan, T.H.; Brunicardi, F.C. p38 MAP kinase interacts with and stabilizes pancreatic and duodenal homeobox-1. Curr. Mol. 2013, 13, 377–386. [Google Scholar] [CrossRef]
- D’Orazi, G.; Cecchinelli, B.; Bruno, T.; Manni, I.; Higashimoto, Y.; Saito, S.; Gostissa, M.; Coen, S.; Marchetti, A.; Del Sal, G.; et al. Homeodomain-interacting protein kinase-2 phosphorylates p53 at Ser 46 and mediates apoptosis. Nat. Cell Biol. 2002, 4, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, T.G.; Möller, A.; Sirma, H.; Zentgraf, H.; Taya, Y.; Dröge, W.; Will, H.; Schmitz, M.L. Regulation of p53 activity by its interaction with homeodomain- interacting protein kinase-2. Nat. Cell Biol. 2002, 4, 1–10. [Google Scholar] [CrossRef]
- Oh, H.; Slattery, M.; Ma, L.; White, K.P.; Mann, R.S.; Irvine, K.D. Yorkie promotes transcription by recruiting a histone methyltransferase complex. Cell Rep. 2014, 8, 449–459. [Google Scholar] [CrossRef]
- Bischof, J.; Maeda, R.K.; Hediger, M.; Karch, F.; Basler, K. An optimized transgenesis system for Drosophila using germ-line-specific phiC31 integrases. Proc. Natl. Acad. Sci. USA 2007, 104, 3312–3317. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steinmetz, E.L.; Dewald, D.N.; Walldorf, U. Drosophila Homeodomain-Interacting Protein Kinase (Hipk) Phosphorylates the Hippo/Warts Signalling Effector Yorkie. Int. J. Mol. Sci. 2021, 22, 1862. https://doi.org/10.3390/ijms22041862
Steinmetz EL, Dewald DN, Walldorf U. Drosophila Homeodomain-Interacting Protein Kinase (Hipk) Phosphorylates the Hippo/Warts Signalling Effector Yorkie. International Journal of Molecular Sciences. 2021; 22(4):1862. https://doi.org/10.3390/ijms22041862
Chicago/Turabian StyleSteinmetz, Eva Louise, Denise Nicole Dewald, and Uwe Walldorf. 2021. "Drosophila Homeodomain-Interacting Protein Kinase (Hipk) Phosphorylates the Hippo/Warts Signalling Effector Yorkie" International Journal of Molecular Sciences 22, no. 4: 1862. https://doi.org/10.3390/ijms22041862
APA StyleSteinmetz, E. L., Dewald, D. N., & Walldorf, U. (2021). Drosophila Homeodomain-Interacting Protein Kinase (Hipk) Phosphorylates the Hippo/Warts Signalling Effector Yorkie. International Journal of Molecular Sciences, 22(4), 1862. https://doi.org/10.3390/ijms22041862