
Growth Factors VEGF-A165 and FGF-2 as Multifunctional Biomolecules Governing Cell Adhesion and Proliferation

 , ,
, ,  , ,
, ,  ,
,  ,
,  and
and 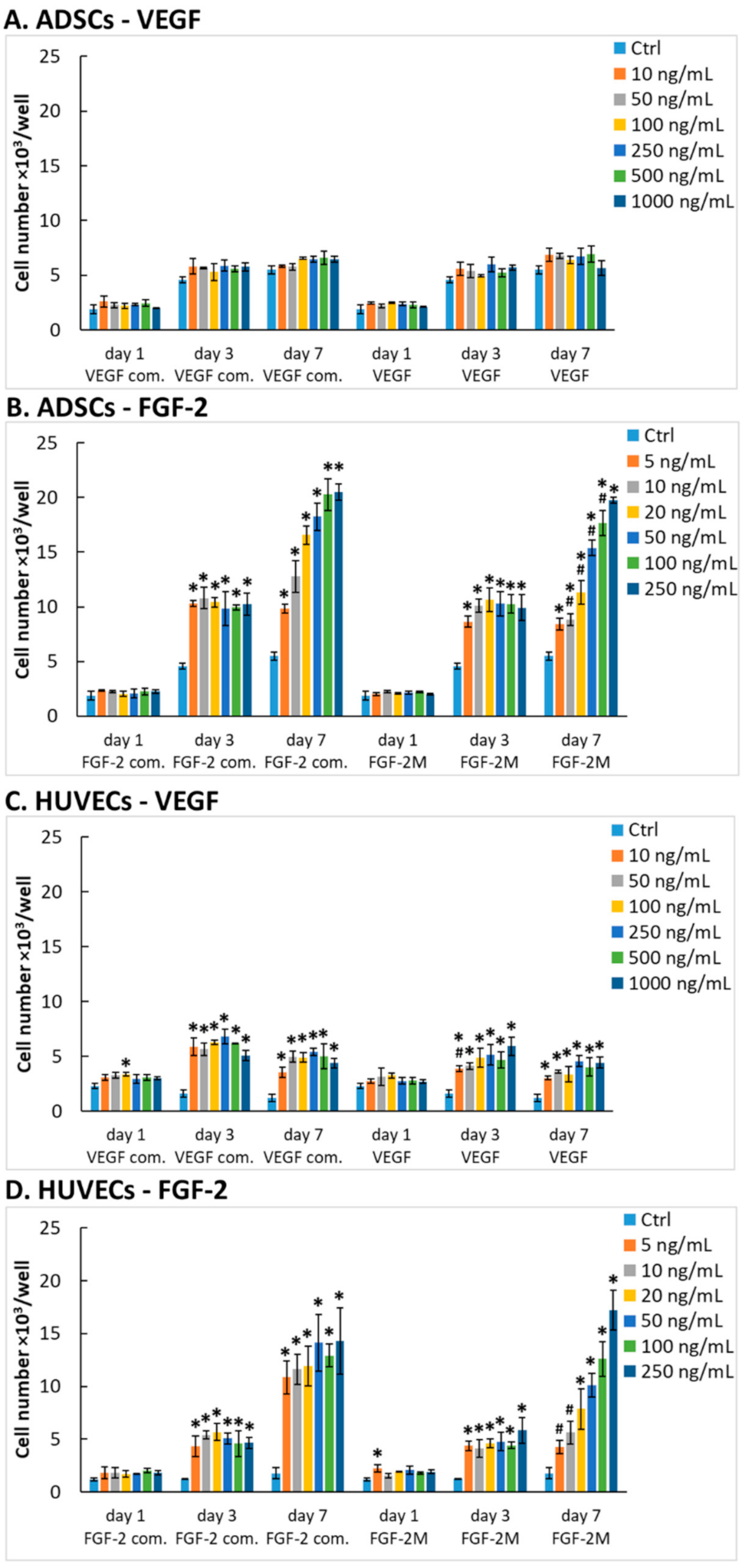{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
2.1. Expression and Purification of Vascular Endothelial Growth Factor (VEGF)-A165 and Fibroblast Growth Factor (FGF)-2M Growth Factors
2.2. Mitogenic Activity of Soluble VEGF-A165 and FGF-2M
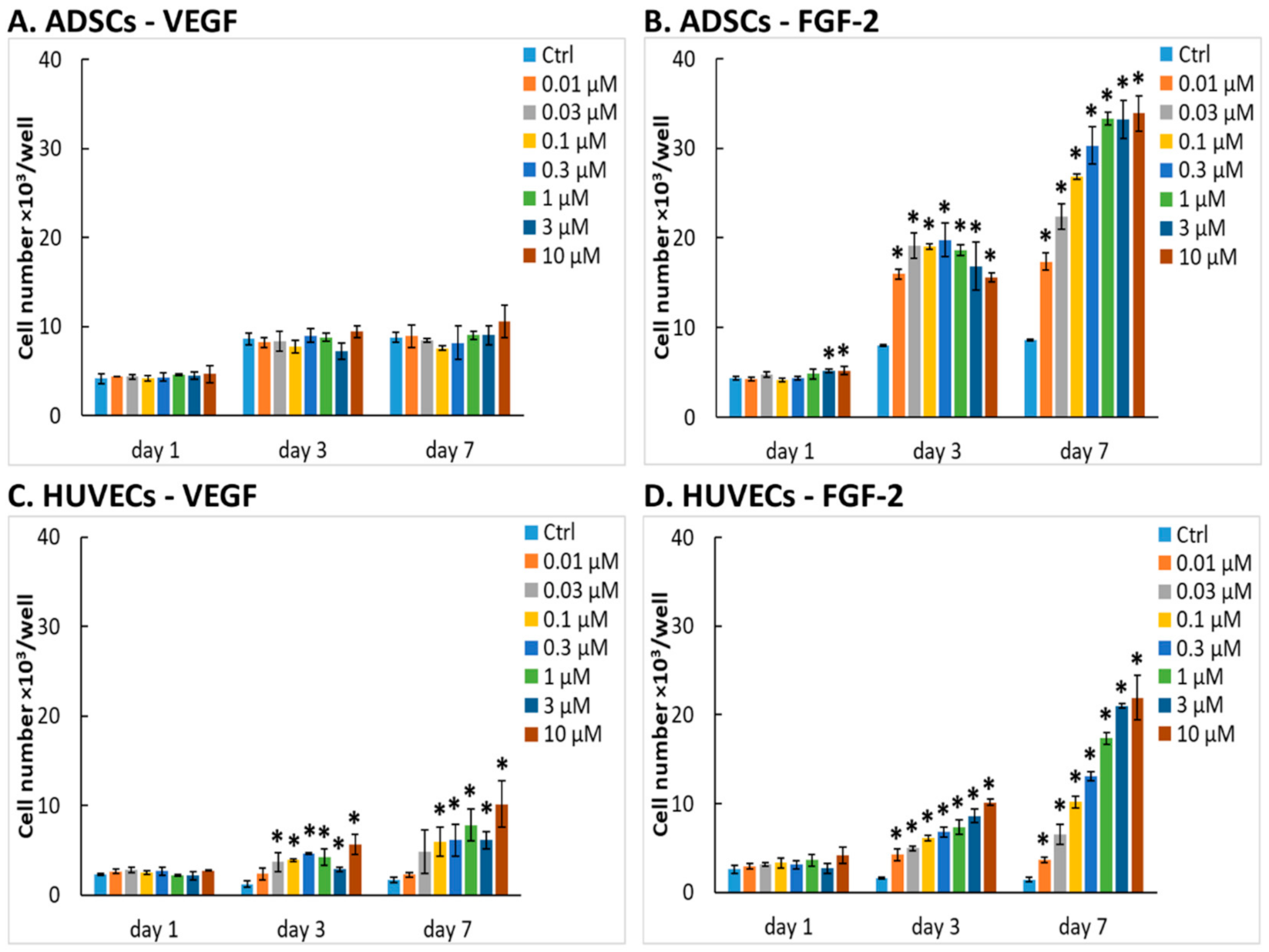
2.2.1. Number of Cells in Media with Growth Factors
2.2.2. Comparison of Our Recombinant and Commercial Growth Factors
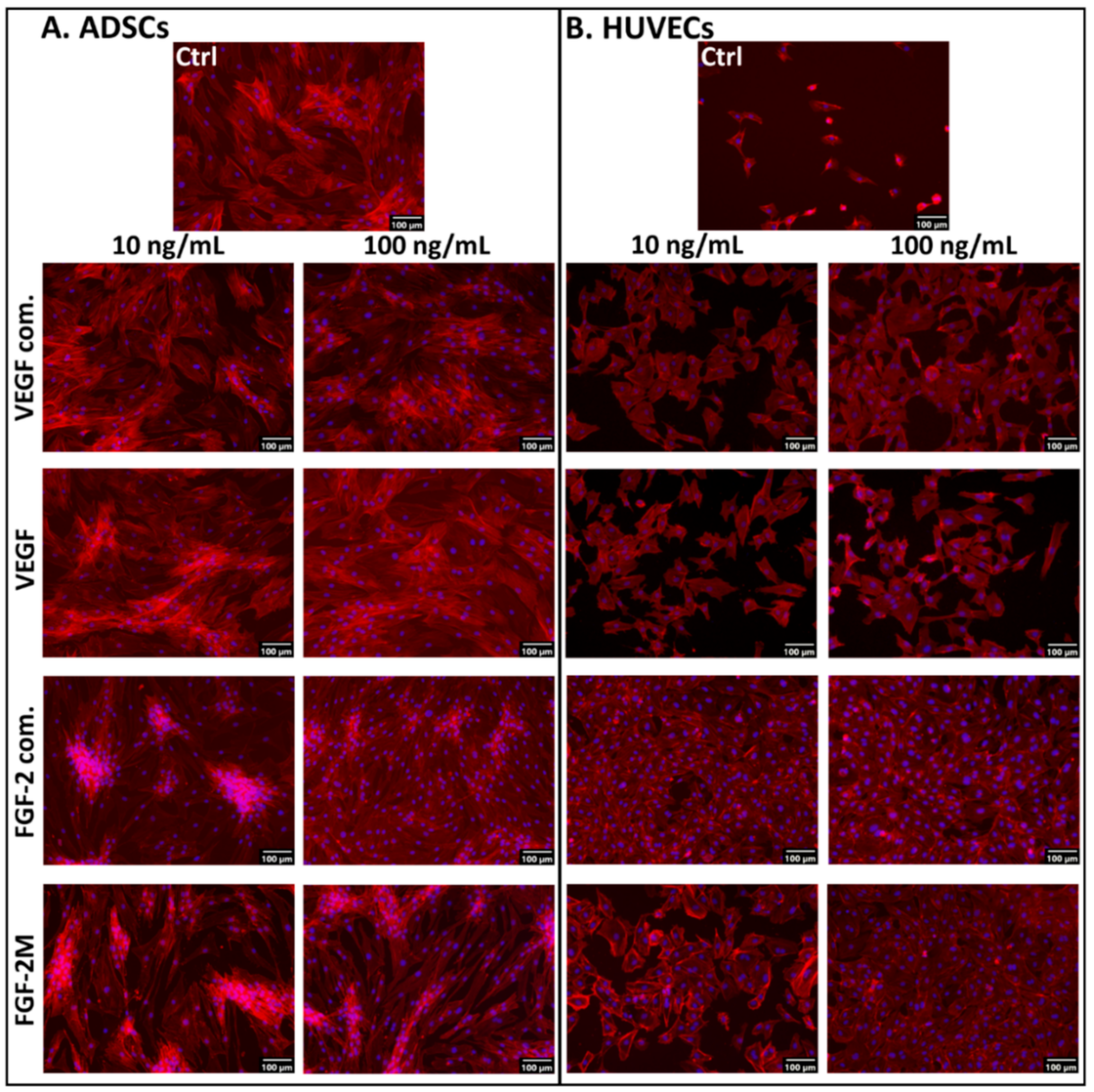
2.2.3. Morphology of Cells in Media with Soluble Growth Factors
2.2.4. Metabolic Activity of Cells in Media with Soluble VEGF-A165 and FGF-2M
2.3. Mitogenic Activity of Adsorbed VEGF-A165 and FGF-2M
2.3.1. Number and Metabolic Activity of Cells on Cultivation Substrates Pre-Adsorbed with Growth Factors
2.3.2. Morphology of Cells on Cultivation Substrates Pre-Adsorbed with Growth Factors
2.4. VEGF-A165 and FGF-2M as Adhesion Ligands for Cells
2.4.1. Adhesion of ADSCs on Cultivation Substrates Pre-Adsorbed with Growth Factors
2.4.2. Adhesion of Human Umbilical Vein Endothelial Cells (HUVECs) on Cultivation Substrates Pre-Adsorbed with Growth Factors
2.5. Limitation of an In Vitro Study
3. Materials and Methods
3.1. Expression and Purification of VEGF-A165
3.2. Expression and Purification of FGF-2M
3.3. Expression of VEGF-A165-FXIIIa and FGF-2M-FXIIIa
3.4. Cell Models
3.5. Cell Cultivation with Recombinant Growth Factors Diluted in Culture Medium
3.6. Cell Cultivation with Recombinant Growth Factors Adsorbed on Culture Wells
3.7. Monitoring the Initial Cell Adhesion to Adsorbed VEGF-A165 and FGF-2M Using xCELLigence System
3.8. Fluorescence Staining and Counting of Cells
3.9. Metabolic Activity Assay
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABAM | Antibiotic antimycotic solution |
| ADSC | Adipose tissue-derived stem cell |
| BMGH | Buffered minimal glycerol medium |
| BMMH | Buffered minimal methanol medium |
| BMGY | Buffered glycerol complex medium |
| BMMY | Buffered methanol complex medium |
| BSA | Bovine serum albumin |
| DAPI | 2-(4-Amidinophenyl)-1H-indole-6-carboxamidine |
| DMEM | Dulbecco’s modified Eagle’s medium |
| EBM2 | Endothelial cell basal medium 2 |
| EGF | Epidermal growth factor |
| EGM2 | Endothelial cell growth medium 2 |
| FBS | Fetal bovine serum |
| FGF-2 | Fibroblast growth factor 2 |
| FGF-2M | Fibroblast growth factor 2 double mutant |
| HEK | Human embryonic kidney |
| HUVEC | Human umbilical vein endothelial cell |
| IGF-1 | Insulin-like growth factor 1 |
| MTS | 3-(4,5-Dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium |
| MTT | 3-[4,5-Dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide |
| PBS | Phosphate-buffered saline |
| PrADSC | Porcine adipose tissue-derived stem cell |
| SDS-PAGE | Sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| TRITC | Tetramethylrhodamine |
| VEGF | Vascular endothelial growth factor |
| VEGF-A | Vascular endothelial growth factor A |
| VEGFR2 | Vascular endothelial growth factor receptor 2 |
| WST | Water-soluble tetrazolium salt |
| XTT | 2,3-bis-(2-Methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide |
| YPD | Yeast extract peptone dextrose |
References
- Bhisitkul, R.B. Vascular endothelial growth factor biology: Clinical implications for ocular treatments. Br. J. Ophthalmol. 2006, 90, 1542–1547. [Google Scholar] [CrossRef]
- Luzuriaga, J.; Irurzun, J.; Irastorza, I.; Unda, F.; Ibarretxe, G.; Pineda, J.R. Vasculogenesis from human dental pulp stem cells grown in matrigel with fully defined serum-free culture media. Biomedicines 2020, 8, 483. [Google Scholar] [CrossRef]
- Kukula, K.; Urbanowicz, A.; Klopotowski, M.; Dabrowski, M.; Pregowski, J.; Kadziela, J.; Chmielak, Z.; Witkowski, A.; Ruzyllo, W. Long-term follow-up and safety assessment of angiogenic gene therapy trial VIF-CAD: Transcatheter intramyocardial administration of a bicistronic plasmid expressing VEGF-A165/bFGF cDNA for the treatment of refractory coronary artery disease. Am. Heart J. 2019, 215, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Anttila, V.; Saraste, A.; Knuuti, J.; Jaakkola, P.; Hedman, M.; Svedlund, S.; Lagerstrom-Fermer, M.; Kjaer, M.; Jeppsson, A.; Gan, L.M. Synthetic mRNA encoding VEGF-A in patients undergoing coronary artery bypass grafting: Design of a phase 2a clinical trial. Mol. Ther. Methods Clin. Dev. 2020, 18, 464–472. [Google Scholar] [CrossRef]
- Kastrup, J. Therapeutic angiogenesis in ischemic heart disease: Gene or recombinant vascular growth factor protein therapy? Curr. Gene Ther. 2003, 3, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Qiu, J.L.; Sun, Q.F.; Yan, S.G.; Wang, W.X.; Yang, P.S.; Song, A.M. One year results evaluating the effects of concentrated growth factors on the healing of intrabony defects treated with or without bone substitute in chronic periodontitis. Med. Sci. Monit. Int. Med J. Exp. Clin. Res. 2019, 25, 4384–4389. [Google Scholar] [CrossRef]
- Fadeev, F.O.; Bashirov, F.V.; Markosyan, V.A.; Izmailov, A.A.; Povysheva, T.V.; Sokolov, M.E.; Kuznetsov, M.S.; Eremeev, A.A.; Salafutdinov, I.I.; Rizvanov, A.A.; et al. Combination of epidural electrical stimulation with ex vivo triple gene therapy for spinal cord injury: A proof of principle study. Neural Regen. Res. 2021, 16, 550–560. [Google Scholar] [CrossRef]
- Masgutov, R.; Zeinalova, A.; Bogov, A.; Masgutova, G.; Salafutdinov, I.; Garanina, E.; Syromiatnikova, V.; Idrisova, K.; Mullakhmetova, A.; Andreeva, D.; et al. Angiogenesis and nerve regeneration induced by local administration of plasmid pBud-coVEGF165-coFGF2 into the intact rat sciatic nerve. Neural Regen. Res. 2021, 16, 1882–1889. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.J.; Nasiri, B.; Kann, J.; Yergeau, D.; Bard, J.E.; Swartz, D.D.; Andreadis, S.T. Endothelialization of arterial vascular grafts by circulating monocytes. Nat. Commun. 2020, 11, 1622. [Google Scholar] [CrossRef] [PubMed]
- Rossi, C.; Lees, M.; Mehta, V.; Heikura, T.; Martin, J.; Zachary, I.; Spencer, R.; Peebles, D.M.; Shaw, R.; Karhinen, M.; et al. Comparison of efficiency and function of vascular endothelial growth factor adenovirus vectors in endothelial cells for gene therapy of placental insufficiency. Hum. Gene Ther. 2020, 31, 1190–1202. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, J.; Ishihara, A.; Starke, R.D.; Peghaire, C.R.; Smith, K.E.; McKinnon, T.A.J.; Tabata, Y.; Sasaki, K.; White, M.J.V.; Fukunaga, K.; et al. The heparin binding domain of von Willebrand factor binds to growth factors and promotes angiogenesis in wound healing. Blood 2019, 133, 2559–2569. [Google Scholar] [CrossRef] [PubMed]
- Knaack, S.; Lode, A.; Hoyer, B.; Rosen-Wolff, A.; Gabrielyan, A.; Roeder, I.; Gelinsky, M. Heparin modification of a biomimetic bone matrix for controlled release of VEGF. J. Biomed. Mater. Res. Part A 2014, 102, 3500–3511. [Google Scholar] [CrossRef] [PubMed]
- Filova, E.; Steinerova, M.; Travnickova, M.; Knitlova, J.; Musilkova, J.; Eckhardt, A.; Hadraba, D.; Matejka, R.; Prazak, S.; Stepanovska, J.; et al. Accelerated in in vitro recellularization of decellularized porcine pericardium for cardiovascular grafts. Biomed. Mater. 2020. [Google Scholar] [CrossRef]
- Hutchings, H.; Ortega, N.; Plouet, J. Extracellular matrix-bound vascular endothelial growth factor promotes endothelial cell adhesion, migration, and survival through integrin ligation. FASEB J. 2003, 17, 1520–1522. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Gonzalez, A.; Aguilera-Montilla, N.; Ugarte-Berzal, E.; Bailon, E.; Cerro-Pardo, I.; Sanchez-Maroto, C.; Garcia-Campillo, L.; Garcia-Marco, J.A.; Garcia-Pardo, A. α4β1 integrin associates with VEGFR2 in CLL cells and contributes to VEGF binding and intracellular signaling. Blood Adv. 2019, 3, 2144–2148. [Google Scholar] [CrossRef]
- Bikfalvi, A.; Klein, S.; Pintucci, G.; Rifkin, D.B. Biological roles of fibroblast growth factor-2. Endocr. Rev. 1997, 18, 26–45. [Google Scholar] [CrossRef]
- Gharibi, B.; Hughes, F.J. Effects of medium supplements on proliferation, differentiation potential, and in vitro expansion of mesenchymal stem cells. Stem Cells Transl. Med. 2012, 1, 771–782. [Google Scholar] [CrossRef]
- Al-Masawa, M.E.; Zaman, W.S.W.K.; Chua, K.H. Biosafety evaluation of culture-expanded human chondrocytes with growth factor cocktail: A preclinical study. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Vahdat, S.; Pahlavan, S.; Mahmoudi, E.; Barekat, M.; Ansari, H.; Bakhshandeh, B.; Aghdami, N.; Baharvand, H. Expansion of human pluripotent stem cell-derived early cardiovascular progenitor cells by a cocktail of signaling factors. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Ahn, H.N.; Kang, H.S.; Park, S.J.; Park, M.H.; Chun, W.; Cho, E. Safety and efficacy of basic fibroblast growth factors for deep second-degree burn patients. Burns 2020, 46, 1857–1866. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.B.; Shen, Z.Y.; Chen, Y.L.; Xie, J.H.; Guo, Z.R.; Zhang, M.L.; Sheng, Z.Y. Recombinant bovine basic fibroblast growth factor accelerates wound healing in patients with burns, donor sites and chronic dermal ulcers. Chin. Med. J. 2000, 113, 367–371. [Google Scholar]
- Kuroda, Y.; Kawai, T.; Goto, K.; Matsuda, S. Clinical application of injectable growth factor for bone regeneration: A systematic review. Inflamm. Regen. 2019, 39. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, W.; Takeuchi, T.; Imamura, K.; Seshima, F.; Saito, A.; Tomita, S. Treatment of chronic periodontitis with recombinant human fibroblast growth factor-2 and deproteinized bovine bone mineral in wide intrabony defects:12-month follow-up case series. Bull. Tokyo Dent. Coll. 2020, 61, 231–241. [Google Scholar] [CrossRef]
- Fiorillo, L.; Cervino, G.; Galindo-Moreno, P.; Herford, A.S.; Spagnuolo, G.; Cicciu, M. Growth factors in oral tissue engineering: New perspectives and current therapeutic options. BioMed Res. Int. 2021, 2021, 8840598. [Google Scholar] [CrossRef]
- Wang, H.; Zhou, W.X.; Huang, J.F.; Zheng, X.Q.; Tian, H.J.; Wang, B.; Fu, W.L.; Wu, A.M. Endocrine therapy for the functional recovery of spinal cord injury. Front. Neurosci. 2020, 14, 590570. [Google Scholar] [CrossRef] [PubMed]
- Salem, S.A.M.; Fezeaa, T.A.; El Khazragy, N.; Soltan, M.Y. Effect of platelet-rich plasma on the outcome of mini-punch grafting procedure in localized stable vitiligo: Clinical evaluation and relation to lesional basic fibroblast growth factor. Dermatol. Ther. 2021, e14738. [Google Scholar] [CrossRef]
- Edamura, K.; Takahashi, Y.; Fujii, A.; Masuhiro, Y.; Narita, T.; Seki, M.; Asano, K. Recombinant canine basic fibroblast growth factor-induced differentiation of canine bone marrow mesenchymal stem cells into voltage- and glutamate-responsive neuron-like cells. Regen. Ther. 2020, 15, 121–128. [Google Scholar] [CrossRef]
- Kurniawan, D.W.; Booijink, R.; Pater, L.; Wols, I.; Vrynas, A.; Storm, G.; Prakash, J.; Bansal, R. Fibroblast growth factor 2 conjugated superparamagnetic iron oxide nanoparticles (FGF2-SPIONs) ameliorate hepatic stellate cells activation in vitro and acute liver injury in vivo. J. Control. Release 2020, 328, 640–652. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, R.; Shome, D.; Vadera, S.; Kumar, V.; Ram, M.S. QR678 & QR678 neo hair growth formulations: A cellular toxicity & animal efficacy study. Plast. Reconstr. Surg. Glob. Open 2020, 8. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Z.; Tang, J.; Li, Y.; You, Q.; Yang, J.; Jin, Y.; Zou, G.; Ge, Z.; Zhu, X.; et al. Fibroblast growth factor 2-induced human amniotic mesenchymal stem cells combined with autologous platelet rich plasma augmented tendon-to-bone healing. J. Orthop. Transl. 2020, 24, 155–165. [Google Scholar] [CrossRef]
- Rusnati, M.; Tanghetti, E.; Dell’Era, P.; Gualandris, A.; Presta, M. αvβ3 integrin mediates the cell-adhesive capacity and biological activity of basic fibroblast growth factor (FGF-2) in cultured endothelial cells. Mol. Biol. Cell 1997, 8, 2449–2461. [Google Scholar] [CrossRef]
- Kang, J.M.; Han, M.; Park, I.S.; Jung, Y.; Kim, S.H.; Kim, S.H. Adhesion and differentiation of adipose-derived stem cells on a substrate with immobilized fibroblast growth factor. Acta Biomater. 2012, 8, 1759–1767. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.M.; Lee, Y.B.; Kim, S.J.; Kang, J.K.; Park, J.C.; Jang, W.; Shin, H. Mussel-inspired immobilization of vascular endothelial growth factor (VEGF) for enhanced endothelialization of vascular grafts. Biomacromolecules 2012, 13, 2020–2028. [Google Scholar] [CrossRef]
- Shen, Y.H.; Shoichet, M.S.; Radisic, M. Vascular endothelial growth factor immobilized in collagen scaffold promotes penetration and proliferation of endothelial cells. Acta Biomater. 2008, 4, 477–489. [Google Scholar] [CrossRef]
- Robinson, D.E.; Smith, L.E.; Steele, D.A.; Short, R.D.; Whittle, J.D. Development of a surface to enhance the effectiveness of fibroblast growth factor 2 (FGF-2). Biomater. Sci. 2014, 2, 875–882. [Google Scholar] [CrossRef]
- Firoozi, N.; Kang, Y. Immobilization of FGF on poly(xylitol dodecanedioic acid) polymer for tissue regeneration. Sci. Rep. 2020, 10, 10419. [Google Scholar] [CrossRef]
- Taborska, J.; Riedelova, Z.; Brynda, E.; Majek, P.; Riedel, T. Endothelialisation of ePTFE vessel prosthesis modified with an antithrombogenic fibrin/heparin coating enriched with bound growth factors. RSC Adv. 2021, 11, 5903–5913. [Google Scholar] [CrossRef]
- Kaplan, O.; Zarubova, J.; Mikulova, B.; Filova, E.; Bartova, J.; Bacakova, L.; Brynda, E. Enhanced mitogenic activity of recombinant human vascular endothelial growth factor VEGF121 expressed in E. coli Origami B (DE3) with molecular chaperones. PLoS ONE 2016, 11, e0163697. [Google Scholar] [CrossRef]
- Taktak-BenAmar, A.; Morjen, M.; Ben Mabrouk, H.; Abdelmaksoud-Dammak, R.; Guerfali, M.; Fourati-Masmoudi, N.; Marrakchi, N.; Gargouri, A. Expression, purification and functionality of bioactive recombinant human vascular endothelial growth factor VEGF165 in E. coli. AMB Express 2017, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Lee, J.E.; Kang, K.J.; Jang, Y.J. Functional efficacy of human recombinant FGF-2s tagged with (His)6 and (His-Asn)6 at the N- and C-termini in human gingival fibroblast and periodontal ligament-derived cells. Protein Expr. Purif. 2017, 135, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Sauer, D.G.; Mosor, M.; Frank, A.C.; Weiss, F.; Christler, A.; Walch, N.; Jungbauer, A.; Durauer, A. A two-step process for capture and purification of human basic fibroblast growth factor from E. coli homogenate: Yield versus endotoxin clearance. Protein Expr. Purif. 2019, 153, 70–82. [Google Scholar] [CrossRef]
- Slamova, K.; Bojarova, P.; Gerstorferova, D.; Fliedrova, B.; Hofmeisterova, J.; Fiala, M.; Pompach, P.; Kren, V. Sequencing, cloning and high-yield expression of a fungal β-N-acetylhexosaminidase in Pichia pastoris. Protein Expr. Purif. 2012, 82, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Krejzova, J.; Kulik, N.; Slamova, K.; Kren, V. Expression of human β-N-acetylhexosaminidase B in yeast eases the search for selective inhibitors. Enzym. Microb. Technol. 2016, 89, 1–6. [Google Scholar] [CrossRef]
- Arjmand, S.; Tavasoli, Z.; Siadat, S.O.R.; Saeidi, B.; Tavana, H. Enhancing chimeric hydrophobin II-vascular endothelial growth factor A165 expression in Pichia pastoris and its efficient purification using hydrophobin counterpart. Int. J. Biol. Macromol. 2019, 139, 1028–1034. [Google Scholar] [CrossRef]
- Zisch, A.H.; Schenk, U.; Schense, J.C.; Sakiyama-Elbert, S.E.; Hubbell, J.A. Covalently conjugated VEGF-fibrin matrices for endothelialization. J. Control. Release 2001, 72, 101–113. [Google Scholar] [CrossRef]
- Rockwell, N.C.; Krysan, D.J.; Komiyama, T.; Fuller, R.S. Precursor processing by Kex2/furin proteases. Chem. Rev. 2002, 102, 4525–4548. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Villalobos, M.A.; Choron, R.L.; Chang, S.; Brown, S.A.; Carpenter, J.P.; Tulenko, T.N.; Zhang, P. Fibroblast growth factor and vascular endothelial growth factor play a critical role in endotheliogenesis from human adipose-derived stem cells. J. Vasc. Surg. 2017, 65, 1483–1492. [Google Scholar] [CrossRef] [PubMed]
- Bassaneze, V.; Barauna, V.G.; Lavini-Ramos, C.; Kalil, J.; Schettert, I.T.; Miyakawa, A.A.; Krieger, J.E. Shear stress induces nitric oxide-mediated vascular endothelial growth factor production in human adipose tissue mesenchymal stem cells. Stem Cells Dev. 2010, 19, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Estes, B.T.; Diekman, B.O.; Gimble, J.M.; Guilak, F. Isolation of adipose-derived stem cells and their induction to a chondrogenic phenotype. Nat. Protoc. 2010, 5, 1294–1311. [Google Scholar] [CrossRef] [PubMed]
- Ori, A.; Free, P.; Courty, J.; Wilkinson, M.C.; Fernig, D.G. Identification of heparin-binding sites in proteins by selective labeling. Mol. Cell. Proteom. 2009, 8, 2256–2265. [Google Scholar] [CrossRef]
- Thompson, L.D.; Pantoliano, M.W.; Springer, B.A. Energetic characterization of the basic fibroblast growth factor-heparin interaction: Identification of the heparin binding domain. Biochemistry 1994, 33, 3831–3840. [Google Scholar] [CrossRef]
- Presta, M.; Statuto, M.; Isacchi, A.; Caccia, P.; Pozzi, A.; Gualandris, A.; Rusnati, M.; Bergonzoni, L.; Sarmientos, P. Structure-function relationship of basic fibroblast growth factor: Site-directed mutagenesis of a putative heparin-binding and receptor-binding region. Biochem. Biophys. Res. Commun. 1992, 185, 1098–1107. [Google Scholar] [CrossRef]
- Isacchi, A.; Bergonzoni, L.; Statuto, M.; Rusnati, M.; Chiesa, R.; Caccia, P.; Sarmientos, P.; Presta, M.; Ragnoiti, G. A mutant of basic fibroblast growth factor that has lost the ability to stimulate plasminogen activator synthesis in endothelial cells. Ann. N. Y. Acad. Sci. 1991, 638, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Tahara, H.; Matsuda, S.; Yamamoto, Y.; Yoshizawa, H.; Fujita, M.; Katsuoka, Y.; Kasahara, T. High-content image analysis (HCIA) assay has the highest correlation with direct counting cell suspension compared to the ATP, WST-8 and Alamar blue assays for measurement of cytotoxicity. J. Pharmacol. Toxicol. Methods 2017, 88, 92–99. [Google Scholar] [CrossRef]
- Walzl, A.; Unger, C.; Kramer, N.; Unterleuthner, D.; Scherzer, M.; Hengstschlager, M.; Schwanzer-Pfeiffer, D.; Dolznig, H. The resazurin reduction assay can distinguish cytotoxic from cytostatic compounds in spheroid screening assays. J. Biomol. Screen 2014, 19, 1047–1059. [Google Scholar] [CrossRef]
- Uzarski, J.S.; DiVito, M.D.; Wertheim, J.A.; Miller, W.M. Essential design considerations for the resazurin reduction assay to noninvasively quantify cell expansion within perfused extracellular matrix scaffolds. Biomaterials 2017, 129, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Chiu, L.L.; Radisic, M. Scaffolds with covalently immobilized VEGF and angiopoietin-1 for vascularization of engineered tissues. Biomaterials 2010, 31, 226–241. [Google Scholar] [CrossRef] [PubMed]
- Miyagi, Y.; Chiu, L.L.; Cimini, M.; Weisel, R.D.; Radisic, M.; Li, R.K. Biodegradable collagen patch with covalently immobilized VEGF for myocardial repair. Biomaterials 2011, 32, 1280–1290. [Google Scholar] [CrossRef]
- Underwood, P.A.; Whitelock, J.M.; Bean, P.A.; Steele, J.G. Effects of base material, plasma proteins and FGF2 on endothelial cell adhesion and growth. J. Biomater. Sci. Polym. Ed. 2002, 13, 845–862. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Park, H.M.; Kim, Y.W.; Kim, Y.H.; Varghese, S.; Seok, H.K.; Kim, Y.G.; Kim, S.H. Control of mesenchymal stem cell phenotype and differentiation depending on cell adhesion mechanism. Eur. Cell Mater. 2014, 28, 387–403. [Google Scholar] [CrossRef] [PubMed]
- Tanghetti, E.; Ria, R.; Dell’Era, P.; Urbinati, C.; Rusnati, M.; Ennas, M.G.; Presta, M. Biological activity of substrate-bound basic fibroblast growth factor (FGF2): Recruitment of FGF receptor-1 in endothelial cell adhesion contacts. Oncogene 2002, 21, 3889–3897. [Google Scholar] [CrossRef] [PubMed]
- Kumorek, M.; Kubies, D.; Filova, E.; Houska, M.; Kasoju, N.; Mazl Chanova, E.; Matejka, R.; Kryslova, M.; Bacakova, L.; Rypacek, F. Cellular responses modulated by FGF-2 adsorbed on albumin/heparin layer-by-layer assemblies. PLoS ONE 2015, 10, e0125484. [Google Scholar] [CrossRef]
- Spitaleri, A.; Mari, S.; Curnis, F.; Traversari, C.; Longhi, R.; Bordignon, C.; Corti, A.; Rizzardi, G.P.; Musco, G. Structural basis for the interaction of isoDGR with the RGD-binding site of αvβ3 integrin. J. Biol. Chem. 2008, 283, 19757–19768. [Google Scholar] [CrossRef]
- Corti, A.; Curnis, F. Isoaspartate-dependent molecular switches for integrin-ligand recognition. J. Cell Sci. 2011, 124, 515–522. [Google Scholar] [CrossRef]
- Traub, S.; Morgner, J.; Martino, M.M.; Honing, S.; Swartz, M.A.; Wickstrom, S.A.; Hubbell, J.A.; Eming, S.A. The promotion of endothelial cell attachment and spreading using FNIII10 fused to VEGF-A165. Biomaterials 2013, 34, 5958–5968. [Google Scholar] [CrossRef]
- Dee, K.C.; Andersen, T.T.; Bizios, R. Design and function of novel osteoblast-adhesive peptides for chemical modification of biomaterials. J. Biomed. Mater. Res. 1998, 40, 371–377. [Google Scholar] [CrossRef]
- Baird, A.; Schubert, D.; Ling, N.; Guillemin, R. Receptor-binding and heparin-binding domains of basic fibroblast growth factor. Proc. Natl. Acad. Sci. USA 1988, 85, 2324–2328. [Google Scholar] [CrossRef]
- Mitchell, A.C.; Briquez, P.S.; Hubbell, J.A.; Cochran, J.R. Engineering growth factors for regenerative medicine applications. Acta Biomater. 2016, 30, 1–12. [Google Scholar] [CrossRef]
- Bacakova, L.; Zarubova, J.; Travnickova, M.; Musilkova, J.; Pajorova, J.; Slepicka, P.; Kasalkova, N.S.; Svorcik, V.; Kolska, Z.; Motarjemi, H.; et al. Stem cells: Their source, potency and use in regenerative therapies with focus on adipose-derived stem cells—A review. Biotechnol. Adv. 2018, 36, 1111–1126. [Google Scholar] [CrossRef] [PubMed]
- Matejka, R.; Konarik, M.; Stepanovska, J.; Lipensky, J.; Chlupac, J.; Turek, D.; Prazak, I.; Broz, A.; Simunkova, Z.; Mrazova, I.; et al. Bioreactor processed stromal cell seeding and cultivation on decellularized pericardium patches for cardiovascular use. Appl. Sci. 2020, 10, 5473. [Google Scholar] [CrossRef]
- Bradford, M.M.; Williams, W.L. New, Rapid, sensitive method for protein determination. Fed. Proc. 1976, 35, 274. [Google Scholar]
- Przekora, A.; Vandrovcova, M.; Travnickova, M.; Pajorova, J.; Molitor, M.; Ginalska, G.; Bacakova, L. Evaluation of the potential of chitosan/β-1,3-glucan/hydroxyapatite material as a scaffold for living bone graft production in vitro by comparison of ADSC and BMDSC behaviour on its surface. Biomed. Mater. 2017, 12, 015030. [Google Scholar] [CrossRef] [PubMed]
- Travnickova, M.; Pajorova, J.; Zarubova, J.; Krocilova, N.; Molitor, M.; Bacakova, L. The influence of negative pressure and of the harvesting site on the characteristics of human adipose tissue-derived stromal cells from lipoaspirates. Stem Cells Int. 2020, 2020, 1016231. [Google Scholar] [CrossRef] [PubMed]
- Bunnell, B.A.; Flaat, M.; Gagliardi, C.; Patel, B.; Ripoll, C. Adipose-derived stem cells: Isolation, expansion and differentiation. Methods 2008, 45, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Megaloikonomos, P.D.; Panagopoulos, G.N.; Bami, M.; Igoumenou, V.G.; Dimopoulos, L.; Milonaki, A.; Kyriakidou, M.; Mitsiokapa, E.; Anastassopoulou, J.; Mavrogenis, A.F. Harvesting, isolation and differentiation of rat adipose-derived stem cells. Curr. Pharm. Biotechnol. 2018, 19, 19–29. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sedlář, A.; Trávníčková, M.; Matějka, R.; Pražák, Š.; Mészáros, Z.; Bojarová, P.; Bačáková, L.; Křen, V.; Slámová, K. Growth Factors VEGF-A165 and FGF-2 as Multifunctional Biomolecules Governing Cell Adhesion and Proliferation. Int. J. Mol. Sci. 2021, 22, 1843. https://doi.org/10.3390/ijms22041843
Sedlář A, Trávníčková M, Matějka R, Pražák Š, Mészáros Z, Bojarová P, Bačáková L, Křen V, Slámová K. Growth Factors VEGF-A165 and FGF-2 as Multifunctional Biomolecules Governing Cell Adhesion and Proliferation. International Journal of Molecular Sciences. 2021; 22(4):1843. https://doi.org/10.3390/ijms22041843
Chicago/Turabian StyleSedlář, Antonín, Martina Trávníčková, Roman Matějka, Šimon Pražák, Zuzana Mészáros, Pavla Bojarová, Lucie Bačáková, Vladimír Křen, and Kristýna Slámová. 2021. "Growth Factors VEGF-A165 and FGF-2 as Multifunctional Biomolecules Governing Cell Adhesion and Proliferation" International Journal of Molecular Sciences 22, no. 4: 1843. https://doi.org/10.3390/ijms22041843
APA StyleSedlář, A., Trávníčková, M., Matějka, R., Pražák, Š., Mészáros, Z., Bojarová, P., Bačáková, L., Křen, V., & Slámová, K. (2021). Growth Factors VEGF-A165 and FGF-2 as Multifunctional Biomolecules Governing Cell Adhesion and Proliferation. International Journal of Molecular Sciences, 22(4), 1843. https://doi.org/10.3390/ijms22041843