
Feed Restriction Modulates Growth, Gut Morphology and Gene Expression in Zebrafish


 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Fish Husbandry and Feeding Strategy
2.3. Feed Restriction Experiment and Sample Collection
2.4. Histology and Microscopy
2.5. RNA Extraction, Library Construction and Sequencing
2.6. Mapping of Reads and Differential Gene Expression Analysis
2.7. Gene Ontology (GO), Eukaryotic Orthologous Groups (KOG) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Analysis
2.8. Statistics
3. Results
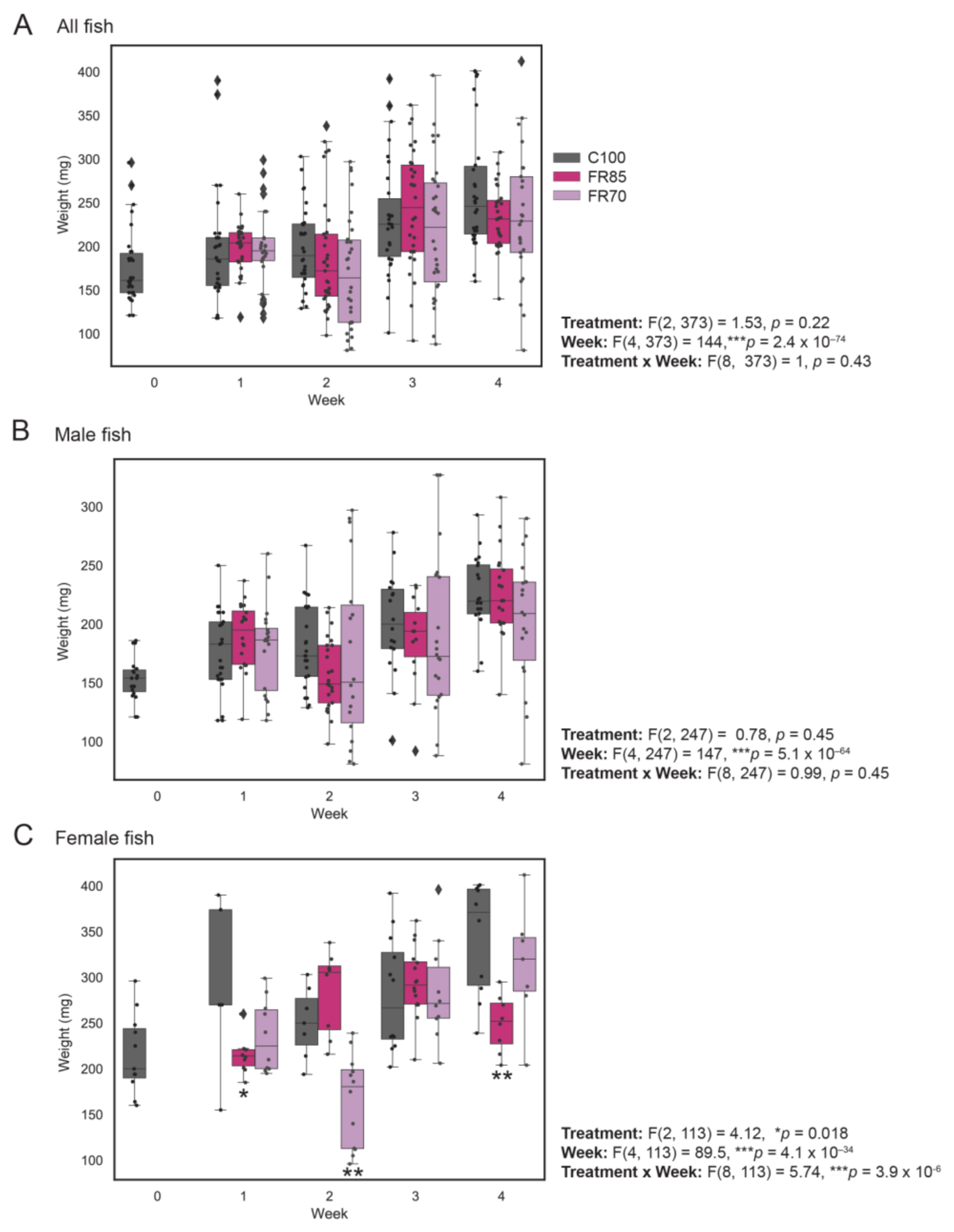
3.1. Weight Profiles of Fully Fed and Feed-Restricted Fish Revealed Minor Differences
3.2. Histological Analyses Reveal Differences in Some Morphological Characteristics
3.3. RNA Sequencing-Based Transcriptomic Profiling
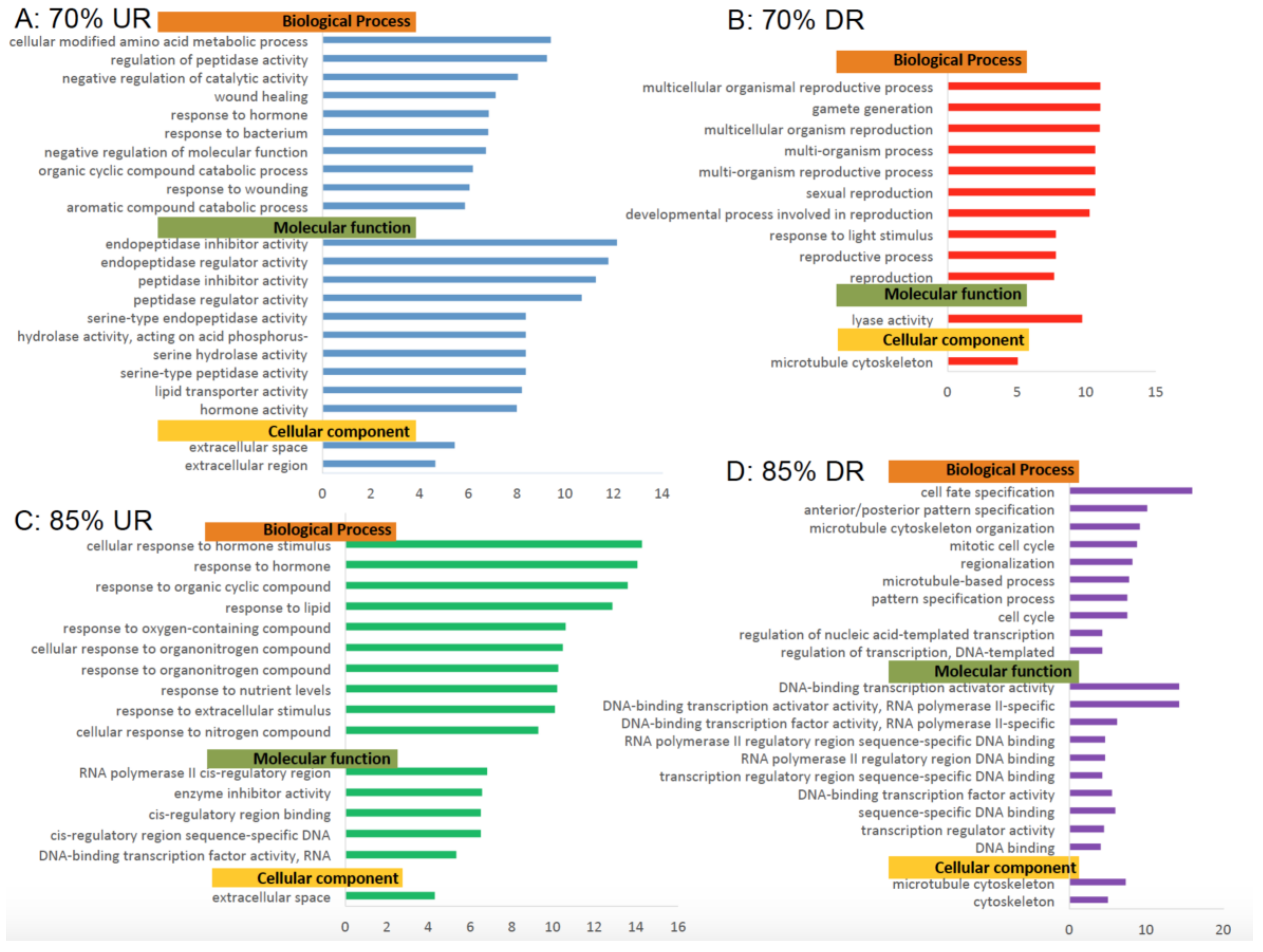
3.4. Bioinformatics Analysis of RNA-Sequencing Data: GO Pathways-Results (GO)
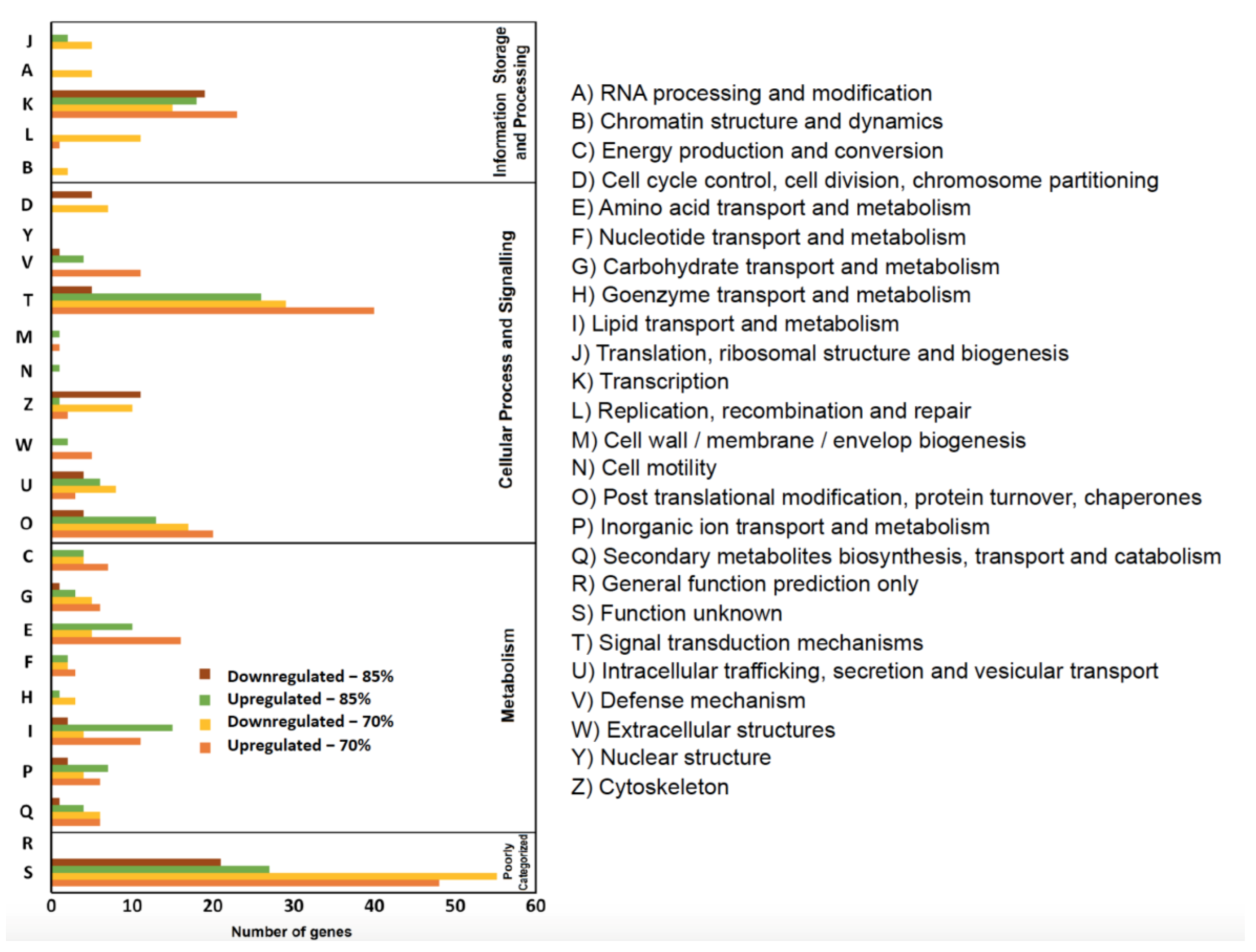
3.5. Bioinformatics Analysis of RNA Sequencing Data: KOG and KEGG Pathways
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sahraei, M. Effects of feed restriction on metabolic disorders in broiler chickens: A review. Biotechnol. Anim. Husb. 2014, 30, 1–13. [Google Scholar] [CrossRef]
- Weber, T.E.; Bosworth, B.G. Effects of 28 day exposure to cold temperature or feed restriction on growth, body composition, and expression of genes related to muscle growth and metabolism in channel catfish. Aquaculture 2005, 246, 483–492. [Google Scholar] [CrossRef]
- Tian, X.; Qin, J.G. Effects of previous ration restriction on compensatory growth in barramundi Lates calcarifer. Aquaculture 2004, 235, 273–283. [Google Scholar] [CrossRef]
- Kim, M.K.; Lovell, R.T. Effect of overwinter feeding regimen on body weight, body composition and resistance to Edwardsiella ictaluri in channel catfish, Ictarulus punctatus. Aquaculture 1995, 134, 237–246. [Google Scholar] [CrossRef]
- Kim, M.K.; Lovell, R.T. Effect of restricted feeding regimens on compensatory weight gain and body tissue changes in channel catfish Ictalurus punctatus in ponds. Aquaculture 1995, 135, 285–293. [Google Scholar] [CrossRef]
- Lovell, T. Fish, feed & nutrition: Feed deprivation increases resistance of channel catfish to bacterial infection. Aquac. Mag.-Ark. 1996, 22, 65–67. [Google Scholar]
- Okwoche, V.O.; Lovell, R.T. Cool Weather Feeding Influences Responses of Channel Catfish to Edwardsiella ictaluri Challenge. J. Aquat. Anim. Health 1997, 9, 163–171. [Google Scholar] [CrossRef]
- Maciel, J.C.; Francisco, C.J.; Miranda-Filho, K.C. Compensatory growth and feed restriction in marine shrimp production, with emphasis on biofloc technology. Aquac. Int. 2017, 26, 203–212. [Google Scholar] [CrossRef]
- Koubova, J.; Guarente, L. How does calorie restriction work? Genes. Dev. 2003, 17, 313–321. [Google Scholar] [CrossRef]
- Tsuchiya, T.; Dhahbi, J.M.; Cui, X.; Mote, P.L.; Bartke, A.; Spindler, S.R. Additive regulation of hepatic gene expression by dwarfism and caloric restriction. Physiol. Genom. 2004, 17, 307–315. [Google Scholar] [CrossRef]
- Hursting, S.D.; Dunlap, S.M.; Ford, N.A.; Hursting, M.J.; Lashinger, L.M. Calorie restriction and cancer prevention: A mechanistic perspective. Cancer Metab. 2013, 1, 10. [Google Scholar] [CrossRef] [PubMed]
- Ravussin, E.; Redman, L.M.; Rochon, J.; Das, S.K.; Fontana, L.; Kraus, W.E.; Romashkan, S.; Williamson, D.A.; Meydani, S.N.; Villareal, D.T.; et al. A 2-Year Randomized Controlled Trial of Human Caloric Restriction: Feasibility and Effects on Predictors of Health Span and Longevity. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2015, 70, 1097–1104. [Google Scholar] [CrossRef]
- Taormina, G.; Mirisola, M.G. Calorie Restriction in Mammals and Simple Model Organisms. BioMed Res. Int. 2014, 2014, 1–10. [Google Scholar] [CrossRef]
- Varady, K.A.; Hellerstein, M.K. Alternate-day fasting and chronic disease prevention: A review of human and animal trials. Am. J. Clin. Nutr. 2007, 86, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Casirola, D.M.; Lan, Y.; Ferraris, R.P. Effects of changes in calorie intake on intestinal nutrient uptake and transporter mRNA levels in aged mice. J. Gerontol. Ser. A Biol. Sci. Med Sci. 1997, 52, B300–B310. [Google Scholar] [CrossRef]
- Ahmed, T.; Das, S.K.; Golden, J.K.; Saltzman, E.; Roberts, S.B.; Meydani, S.N. Calorie Restriction Enhances T-Cell-Mediated Immune Response in Adult Overweight Men and Women. J. Gerontol. Ser. A Biol. Sci. Med Sci. 2009, 64, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Vanegas, S.M.; Du, X.; Noble, T.; Zingg, J.-M.A.; Meydani, M.; Meydani, S.N.; Wu, D. Caloric restriction favorably impacts metabolic and immune/inflammatory profiles in obese mice but curcumin/piperine consumption adds no further benefit. Nutr. Metab. 2013, 10, 29. [Google Scholar] [CrossRef]
- Tocher, D.R. Metabolism and Functions of Lipids and Fatty Acids in Teleost Fish. Rev. Fish. Sci. 2003, 11, 107–184. [Google Scholar] [CrossRef]
- Hołowko, J.; Michalczyk, M.M.; Zając, A.; Czerwińska-Rogowska, M.; Ryterska, K.; Banaszczak, M.; Jakubczyk, K.; Stachowska, E. Six Weeks of Calorie Restriction Improves Body Composition and Lipid Profile in Obese and Overweight Former Athletes. Nutrients 2019, 11, 1461. [Google Scholar] [CrossRef]
- Mattison, J.A.; Lane, M.A.; Roth, G.S.; Ingram, D.K. Calorie restriction in rhesus monkeys. Exp. Gerontol. 2003, 38, 35–46. [Google Scholar] [CrossRef]
- Ma, S.; Sun, S.; Geng, L.; Song, M.; Wang, W.; Ye, Y.; Ji, Q.; Zou, Z.; Wang, S.; He, X.; et al. Caloric Restriction Reprograms the Single-Cell Transcriptional Landscape of Rattus Norvegicus Aging. Cell 2020, 180, 984–1001.e22. [Google Scholar] [CrossRef]
- Pifferi, F.; Terrien, J.; Marchal, J.; Dal-Pan, A.; Djelti, F.; Hardy, I.; Chahory, S.; Cordonnier, N.; Desquilbet, L.; Hurion, M.; et al. Caloric restriction increases lifespan but affects brain integrity in grey mouse lemur primates. Commun. Biol. 2018, 1, 30. [Google Scholar] [CrossRef]
- Sardeli, A.V.; Komatsu, T.R.; Mori, M.A.; Gáspari, A.F.; Chacon-Mikahil, M.P.T. Resistance Training Prevents Muscle Loss Induced by Caloric Restriction in Obese Elderly Individuals: A Systematic Review and Meta-Analysis. Nutrients 2018, 10, 423. [Google Scholar] [CrossRef]
- Kumar, P.; Sudhagar, S.A.; Harikrishna, V.; Jayant, M. Practical signifi cance of restricted feeding regime in aquaculture. Aquac. Asia Mag. 2017, 21, 18. [Google Scholar]
- Azodi, M.; Nafisi, M.; Morshedi, V.; Modarresi, M.; Faghih-Ahmadani, A. Effects of intermittent feeding on compensatory growth, feed intake and body composition in Asian sea bass (Lates calcarifer). Iran. J. Fish. Sci. 2016, 15, 144–156. [Google Scholar]
- Purushothaman, K.; Lau, D.; Saju, J.M.; Lunny, D.P.; Vij, S.; Orbán, L. Morpho-histological characterisation of the alimentary canal of an important food fish, Asian seabass (Lates calcarifer). PeerJ 2016, 4, e2377. [Google Scholar] [CrossRef] [PubMed]
- Sáez, M.I.; Navarro, G.; García-Mesa, S.; Martínez, T.F.; García-Gallego, M.; Suárez, M.D. Influence of pre-slaughtering feed restriction on muscle characteristics of farmed sea bass (Dicentrarchus labrax L.) during cold storage. J. Sci. Food Agric. 2013, 93, 2323–2330. [Google Scholar] [PubMed]
- Ulloa, P.E.; Medrano, J.F.; Feijóo, C.G. Zebrafish as animal model for aquaculture nutrition research. Front. Genet. 2014, 5, 313. [Google Scholar] [CrossRef]
- Drew, R.E.; Rodnick, K.J.; Settles, M.; Wacyk, J.; Churchill, E.; Powell, M.S.; Hardy, R.W.; Murdoch, G.K.; Hill, R.A.; Robison, B.D. Effect of starvation on transcriptomes of brain and liver in adult female zebrafish (Danio rerio). Physiol. Genom. 2008, 35, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Leibold, S.; Hammerschmidt, M. Long-Term Hyperphagia and Caloric Restriction Caused by Low- or High-Density Husbandry Have Differential Effects on Zebrafish Postembryonic Development, Somatic Growth, Fat Accumulation and Reproduction. PLoS ONE 2015, 10, e0120776. [Google Scholar] [CrossRef]
- Egerton, S.; Culloty, S.; Whooley, J.; Stanton, C.; Ross, R.P. The Gut Microbiota of Marine Fish. Front. Microbiol. 2018, 9, 873. [Google Scholar] [CrossRef] [PubMed]
- Yukgehnaish, K.; Kumar, P.; Sivachandran, P.; Marimuthu, K.; Arshad, A.; Paray, B.A.; Arockiaraj, J. Gut microbiota metagenomics in aquaculture: Factors influencing gut microbiome and its physiological role in fish. Rev. Aquac. 2020, 12, 1903–1927. [Google Scholar] [CrossRef]
- Perry, W.B.; Lindsay, E.; Payne, C.J.; Brodie, C.; Kazlauskaite, R. The role of the gut microbiome in sustainable teleost aquaculture. Proc. R. Soc. B Biol. Sci. 2020, 287, 20200184. [Google Scholar] [CrossRef]
- Le, H.T.M.D.; Shao, X.; Krogdahl, Å.; Kortner, T.M.; Lein, I.; Kousoulaki, K.; Lie, K.K.; Sæle, Ø. Intestinal function of the stomachless fish, Ballan wrasse (Labrus bergylta). Front. Mar. Sci. 2019, 6, 140. [Google Scholar] [CrossRef]
- Wallace, K.N.; Akhter, S.; Smith, E.M.; Lorent, K.; Pack, M. Intestinal growth and differentiation in zebrafish. Mech. Dev. 2005, 122, 157–173. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A.; Koonin, E.V. The COG database: A tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Hiramatsu, N.; Todo, T.; Sullivan, C.V.; Schilling, J.; Reading, B.J.; Matsubara, T.; Ryu, Y.-W.; Mizuta, H.; Luo, W.; Nishimiya, O.; et al. Ovarian yolk formation in fishes: Molecular mechanisms underlying formation of lipid droplets and vitellogenin-derived yolk proteins. Gen. Comp. Endocrinol. 2015, 221, 9–15. [Google Scholar] [CrossRef]
- Yilmaz, O.; Patinote, A.; Nguyen, T.; Bobe, J. Multiple vitellogenins in zebrafish (Danio rerio): Quantitative inventory of genes, transcripts and proteins, and relation to egg quality. Fish Physiol. Biochem. 2018, 44, 1509–1525. [Google Scholar] [CrossRef]
- Hellman, N.E.; Liu, Y.; Merkel, E.; Austin, C.; Le Corre, S.; Beier, D.R.; Sun, Z.; Sharma, N.; Yoder, B.K.; Drummond, I.A. The zebrafish foxj1a transcription factor regulates cilia function in response to injury and epithelial stretch. Proc. Natl. Acad. Sci. USA 2010, 107, 18499–18504. [Google Scholar] [CrossRef]
- Ryan, R.; Failler, M.; Reilly, M.L.; Garfa-Traore, M.; Delous, M.; Filhol, E.; Reboul, T.; Bole-Feysot, C.; Nitschké, P.; Baudouin, V. Functional characterization of tektin-1 in motile cilia and evidence for TEKT1 as a new candidate gene for motile ciliopathies. Hum. Mol. Genet. 2018, 27, 266–282. [Google Scholar] [CrossRef]
- Espenschied, S.T.; Cronan, M.R.; Matty, M.A.; Mueller, O.; Redinbo, M.R.; Tobin, D.M.; Rawls, J.F. Epithelial delamination is protective during pharmaceutical-induced enteropathy. Proc. Natl. Acad. Sci. USA 2019, 116, 16961–16970. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, B.; Kubota, A.; O’Meara, C.M.; Lamb, D.C.; Tanguay, R.L.; Goldstone, J.V.; Stegeman, J.J. Cytochrome P450 20A1 in zebrafish: Cloning, regulation and potential involvement in hyperactivity disorders. Toxicol. Appl. Pharmacol. 2016, 296, 73–84. [Google Scholar] [CrossRef]
- Piccinetti, C.C.; De Leo, A.; Cosoli, G.; Scalise, L.; Randazzo, B.; Cerri, G.; Olivotto, I. Measurement of the 100 MHz EMF radiation in vivo effects on zebrafish D. rerio embryonic development: A multidisciplinary study. Ecotoxicol. Environ. Saf. 2018, 154, 268–279. [Google Scholar] [CrossRef]
- Greiling, T.M.; Houck, S.A.; Clark, J.I. The zebrafish lens proteome during development and aging. Mol. Vis. 2009, 15, 2313. [Google Scholar]
- Yan, Y.; He, F.; Li, Z.; Xu, R.; Li, T.; Su, J.; Liu, X.; Zhao, M.; Wu, W. The important role of apolipoprotein A-II in ezetimibe driven reduction of high cholesterol diet-induced atherosclerosis. Atherosclerosis 2019, 280, 99–108. [Google Scholar] [CrossRef]
- Kwon, O.K.; Kim, S.J.; Lee, S. First profiling of lysine crotonylation of myofilament proteins and ribosomal proteins in zebrafish embryos. Sci. Rep. 2018, 8, 3652. [Google Scholar] [CrossRef]
- Eno, C.; Pelegri, F. Modulation of F-actin dynamics by maternal Mid1ip1L controls germ plasm aggregation and furrow recruitment in the zebrafish embryo. Development 2018, 145. [Google Scholar] [CrossRef] [PubMed]
- Eno, C.; Hansen, C.L.; Pelegri, F. Aggregation, segregation, and dispersal of homotypic germ plasm RNPs in the early zebrafish embryo. Dev. Dyn. 2019, 248, 306–318. [Google Scholar] [CrossRef] [PubMed]
- Eno, C.; Solanki, B.; Pelegri, F. aura(mid1ip1l) regulates the cytoskeleton at the zebrafish egg-to-embryo transition. Development 2016, 143, 1585–1599. [Google Scholar] [CrossRef]
- Jiao, W.; Parker, E.J. Using a Combination of Computational and Experimental Techniques to Understand the Molecular Basis for Protein Allostery. In Insights into Enzyme Mechanisms and Functions from Experimental and Computational Methods; Elsevier: Amsterdam, The Netherlands, 2012; Volume 87, pp. 391–413. [Google Scholar]
- Polonsky, K.; Burant, C. Type 2 diabetes mellitus. In Williams Textbook of Endocrinology; Elsevier: Amsterdam, The Netherlands, 2016; pp. 1385–1450. [Google Scholar]
- Marandel, L.; Panserat, S.; Plagnes-Juan, E.; Arbenoits, E.; Soengas, J.L.; Bobe, J. Evolutionary history of glucose-6-phosphatase encoding genes in vertebrate lineages: Towards a better understanding of the functions of multiple duplicates. BMC Genom. 2017, 18, 342. [Google Scholar] [CrossRef]
- Huss, M.; Byström, P.; Persson, L. Resource heterogeneity, diet shifts and intra-cohort competition: Effects on size divergence in YOY fish. Oecologia 2008, 158, 249–257. [Google Scholar] [CrossRef]
- Wise, D.H. Cannibalism, food limitation, intraspecific competition, and the regulation of spider populations. Annu. Rev. Èntomol. 2006, 51, 441–465. [Google Scholar] [CrossRef]
- Inness, C.L.W.; Metcalfe, N.B. The impact of dietary restriction, intermittent feeding and compensatory growth on reproductive investment and lifespan in a short-lived fish. Proc. R. Soc. B Biol. Sci. 2008, 275, 1703–1708. [Google Scholar] [CrossRef]
- Mattison, J.A.; Roth, G.S.; Beasley, T.M.; Tilmont, E.M.; Handy, A.M.; Herbert, R.L.; Longo, D.L.; Allison, D.B.; Young, J.E.; Bryant, M.; et al. Impact of caloric restriction on health and survival in rhesus monkeys from the NIA study. Nature 2012, 489, 318–321. [Google Scholar] [CrossRef] [PubMed]
- Rana, K.J.; Siriwardena, S.; Hasan, M.R. Impact of Rising Feed Ingredient Prices on Aquafeeds and Aquaculture Production; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2009. [Google Scholar]
- Sankaran-Walters, S.; Macal, M.; Grishina, I.; Nagy, L.; Goulart, L.; Coolidge, K.; Li, J.; Fenton, A.; Williams, T.; Miller, M.K.; et al. Sex differences matter in the gut: Effect on mucosal immune activation and inflammation. Biol. Sex Differ. 2013, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Rao, J.; Wang, J. Luminal nutrients in health and microbes in gut mucosal growth. In Regulation of Gastrointestinal Mucosal Growth; NCBI Bookshelf; Morgan & Claypool Life Sciences: San Rafael, CA, USA, 2010. [Google Scholar]
- Ferraris, R.P.; Carey, H.V. Intestinal transport during fasting and malnutrition. Annu. Rev. Nutr. 2000, 20, 195–219. [Google Scholar] [CrossRef]
- Heller, T.D.; Holt, P.R.; Richardson, A. Food restriction retards age-related histological changes in rat small intestine. Gastroenterology 1990, 98, 387–391. [Google Scholar] [CrossRef]
- Cornick, S.; Tawiah, A.; Chadee, K. Roles and regulation of the mucus barrier in the gut. Tissue Barriers 2015, 3, e982426. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, J.A.; Goodlad, R.A.; Wright, N.A. Maintenance of normal intestinal mucosa: Function, structure, and adaptation. Gut 1994, 35 (Suppl. S1), S1–S4. [Google Scholar] [CrossRef]
- O’Brien, L.E.; Soliman, S.S.; Li, X.; Bilder, D. Altered modes of stem cell division drive adaptive intestinal growth. Cell 2011, 147, 603–614. [Google Scholar] [CrossRef]
- Tanca, A.; Abbondio, M.; Palomba, A.; Fraumene, C.; Marongiu, F.; Serra, M.; Pagnozzi, D.; Laconi, E.; Uzzau, S. Caloric restriction promotes functional changes involving short-chain fatty acid biosynthesis in the rat gut microbiota. Sci. Rep. 2018, 8, 14778. [Google Scholar] [CrossRef] [PubMed]
- Ferraris, R.P.; Cao, Q.-X.; Prabhakaram, S. Chronic but not acute energy restriction increases intestinal nutrient transport in mice. J. Nutr. 2001, 131, 779–786. [Google Scholar] [CrossRef]
- Sreekumar, R.; Unnikrishnan, J.; Fu, A.; Nygren, J.; Short, K.R.; Schimke, J.; Barazzoni, R.; Nair, K.S. Effects of caloric restriction on mitochondrial function and gene transcripts in rat muscle. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E38–E43. [Google Scholar] [CrossRef] [PubMed]
- Cox, L.M.; Schafer, M.J.; Sohn, J.; Vincentini, J.; Weiner, H.L.; Ginsberg, S.D.; Blaser, M.J. Calorie restriction slows age-related microbiota changes in an Alzheimer’s disease model in female mice. Sci. Rep. 2019, 9, 17904. [Google Scholar] [CrossRef]
- Richards, A.L.; Muehlbauer, A.L.; Alazizi, A.; Burns, M.B.; Findley, A.; Messina, F.; Gould, T.J.; Cascardo, C.; Pique-Regi, R.; Blekhman, R.; et al. Gut Microbiota Has a Widespread and Modifiable Effect on Host Gene Regulation. MSystems 2019, 4, e00323-18. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Barrón, E.; Hernández, C.; Llera-Herrera, R.; García-Gasca, A.; Gómez-Gil, B. Overfeeding a High-Fat Diet Promotes Sex-Specific Alterations on the Gut Microbiota of the Zebrafish (Danio rerio). Zebrafish 2019, 16, 268–279. [Google Scholar] [CrossRef]
- Cruzen, C.; Colman, R.J. Effects of Caloric Restriction on Cardiovascular Aging in Non-human Primates and Humans. Clin. Geriatr. Med. 2009, 25, 733–743. [Google Scholar] [CrossRef]
- Longo, V.D.; Fontana, L. Calorie restriction and cancer prevention: Metabolic and molecular mechanisms. Trends Pharmacol. Sci. 2010, 31, 89–98. [Google Scholar] [CrossRef]
- Contreras, N.A.; Fontana, L.; Tosti, V.; Nikolich-Žugich, J. Calorie restriction induces reversible lymphopenia and lymphoid organ atrophy due to cell redistribution. GeroScience 2018, 40, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Peck, M.D.; Babcock, G.F.; Alexander, J.W. The Role of Protein and Calorie Restriction in Outcome From Salmonella Infection in Mice. J. Parenter. Enter. Nutr. 1992, 16, 561–565. [Google Scholar] [CrossRef]
- Reiser, K.; McGee, C.; Rucker, R.; McDonald, R. Effects of Aging and Caloric Restriction on Extracellular Matrix Biosynthesis in a Model of Injury Repair in Rats. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 1995, 50, B40–B47. [Google Scholar] [CrossRef]
- Landgraf, K.; Schuster, S.; Meusel, A.; Garten, A.; Riemer, T.; Schleinitz, D.; Kiess, W.; Körner, A. Short-term overfeeding of zebrafish with normal or high-fat diet as a model for the development of metabolically healthy versus unhealthy obesity. BMC Physiol. 2017, 17, 4. [Google Scholar] [CrossRef] [PubMed]
- Roh, H.; Park, J.; Kim, A.; Kim, N.; Lee, Y.; Kim, B.S.; Vijayan, J.; Lee, M.K.; Park, C.-I.; Kim, D.-H. Overfeeding-Induced Obesity Could Cause Potential Immuno-Physiological Disorders in Rainbow Trout (Oncorhynchus mykiss). Animals 2020, 10, 1499. [Google Scholar] [CrossRef] [PubMed]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef] [PubMed]
- Hussey, G.S.; Keane, T.J.; Badylak, S.F. The extracellular matrix of the gastrointestinal tract: A regenerative medicine platform. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 540–552. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group 3 (100% Feeding) | Group 2 (85% Feeding) | Group 1 (70% Feeding) | ||||
|---|---|---|---|---|---|---|
| Week | Male | Female | Male | Female | Male | Female |
| 0 | 19 | 11 | NA | NA | NA | NA |
| 1 | 25 | 5 | 20 | 10 | 20 | 10 |
| 2 | 23 | 7 | 25 | 8 | 18 | 12 |
| 3 | 18 | 12 | 14 | 16 | 20 | 10 |
| 4 | 20 | 10 | 20 | 8 | 18 | 7 |
| Pathway | Number of High/Low Abundance Genes in FR70 | Number of High/Low Abundance Genes in FR85 |
|---|---|---|
| map01100 Metabolic pathways | 30/28 | 23/3 |
| map04024 cAMP signalling pathway | 6/1 | 5/1 |
| map00230 Purine metabolism | 5/1 | 2/0 |
| map01200 Carbon metabolism | 5/3 | 2/0 |
| map04979 Cholesterol metabolism | 5/5 | 2/1 |
| map04911 Insulin secretion | 4/1 | 3/0 |
| map04978 Mineral absorption | 4/2 | 3/0 |
| map00010 Glycolysis / Gluconeogenesis | 4/2 | 1/0 |
| map00630 Glyoxylate and dicarboxylate metabolism | 4/2 | 2/0 |
| map04218 Cellular senescence | 3/5 | 4/1 |
| map04923 Regulation of lipolysis in adipocytes | 3/0 | 2/0 |
| map00620 Pyruvate metabolism | 3/2 | 0/0 |
| map04810 Regulation of actin cytoskeleton | 3/0 | 4/1 |
| map04510 Focal adhesion | 2/0 | 2/1 |
| map04210 Apoptosis | 2/1 | 3/2 |
| map04973 Carbohydrate digestion and absorption | 2/1 | 2/0 |
| map04975 Fat digestion and absorption | 2/3 | 4/0 |
| map05210 Colorectal cancer | 2/3 | 4/2 |
| map04977 Vitamin digestion and absorption | 2/1 | 3/0 |
| map04217 Necroptosis | 2/1 | 0/3 |
| map01230 Biosynthesis of amino acids | 1/3 | 3/0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Purushothaman, K.; Tan, J.K.H.; Lau, D.; Saju, J.M.; Thevasagayam, N.M.; Wee, C.L.; Vij, S. Feed Restriction Modulates Growth, Gut Morphology and Gene Expression in Zebrafish. Int. J. Mol. Sci. 2021, 22, 1814. https://doi.org/10.3390/ijms22041814
Purushothaman K, Tan JKH, Lau D, Saju JM, Thevasagayam NM, Wee CL, Vij S. Feed Restriction Modulates Growth, Gut Morphology and Gene Expression in Zebrafish. International Journal of Molecular Sciences. 2021; 22(4):1814. https://doi.org/10.3390/ijms22041814
Chicago/Turabian StylePurushothaman, Kathiresan, Jerryl Kim Han Tan, Doreen Lau, Jolly M. Saju, Natascha M. Thevasagayam, Caroline Lei Wee, and Shubha Vij. 2021. "Feed Restriction Modulates Growth, Gut Morphology and Gene Expression in Zebrafish" International Journal of Molecular Sciences 22, no. 4: 1814. https://doi.org/10.3390/ijms22041814
APA StylePurushothaman, K., Tan, J. K. H., Lau, D., Saju, J. M., Thevasagayam, N. M., Wee, C. L., & Vij, S. (2021). Feed Restriction Modulates Growth, Gut Morphology and Gene Expression in Zebrafish. International Journal of Molecular Sciences, 22(4), 1814. https://doi.org/10.3390/ijms22041814