
Efficient Transformation of Somatic Embryos and Regeneration of Cork Oak Plantlets with a Gene (CsTL1) Encoding a Chestnut Thaumatin-Like Protein

 , ,
, ,
Abstract
1. Introduction
2. Results
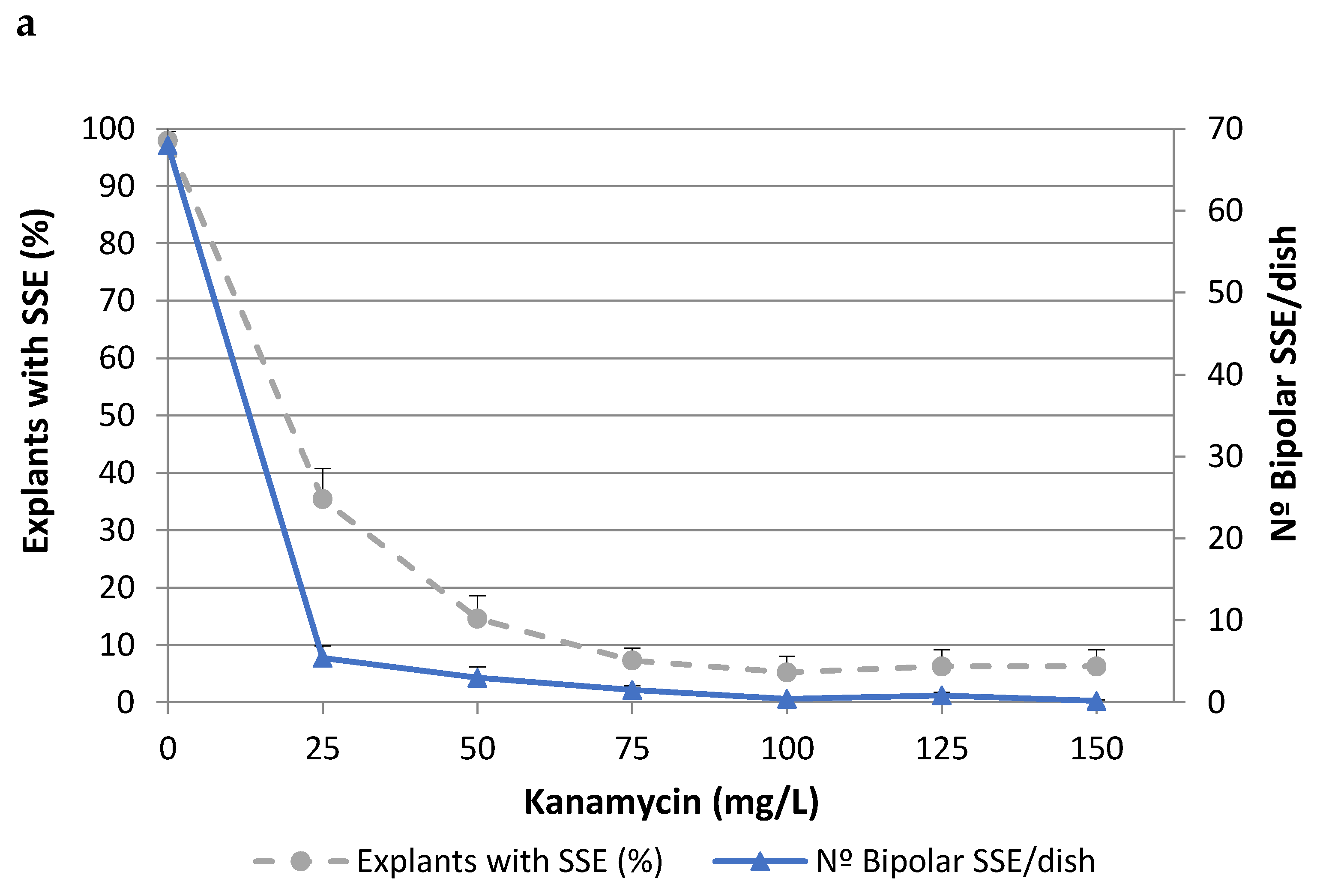
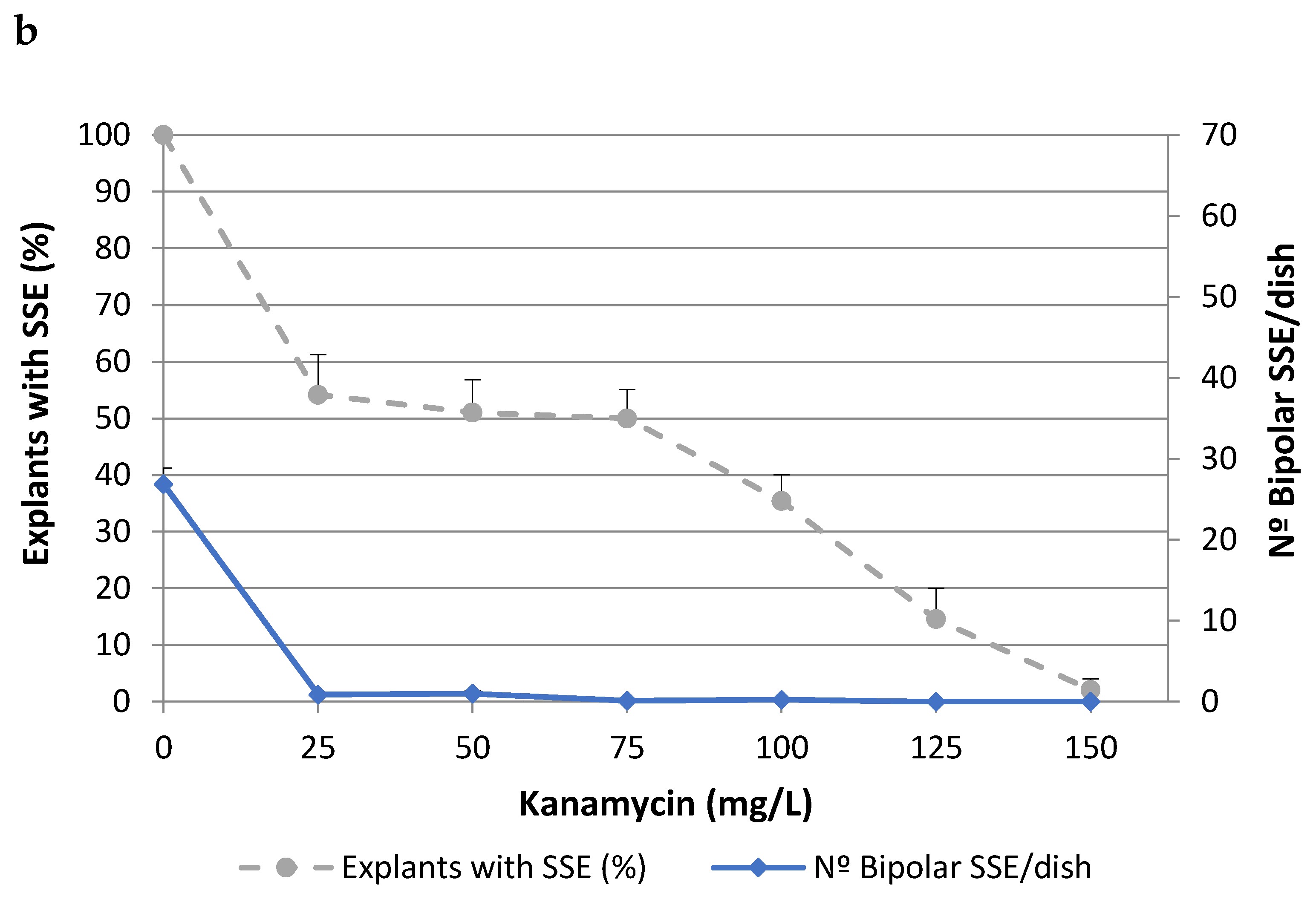
2.1. Effects of Antibiotics on Somatic Embryo Proliferation
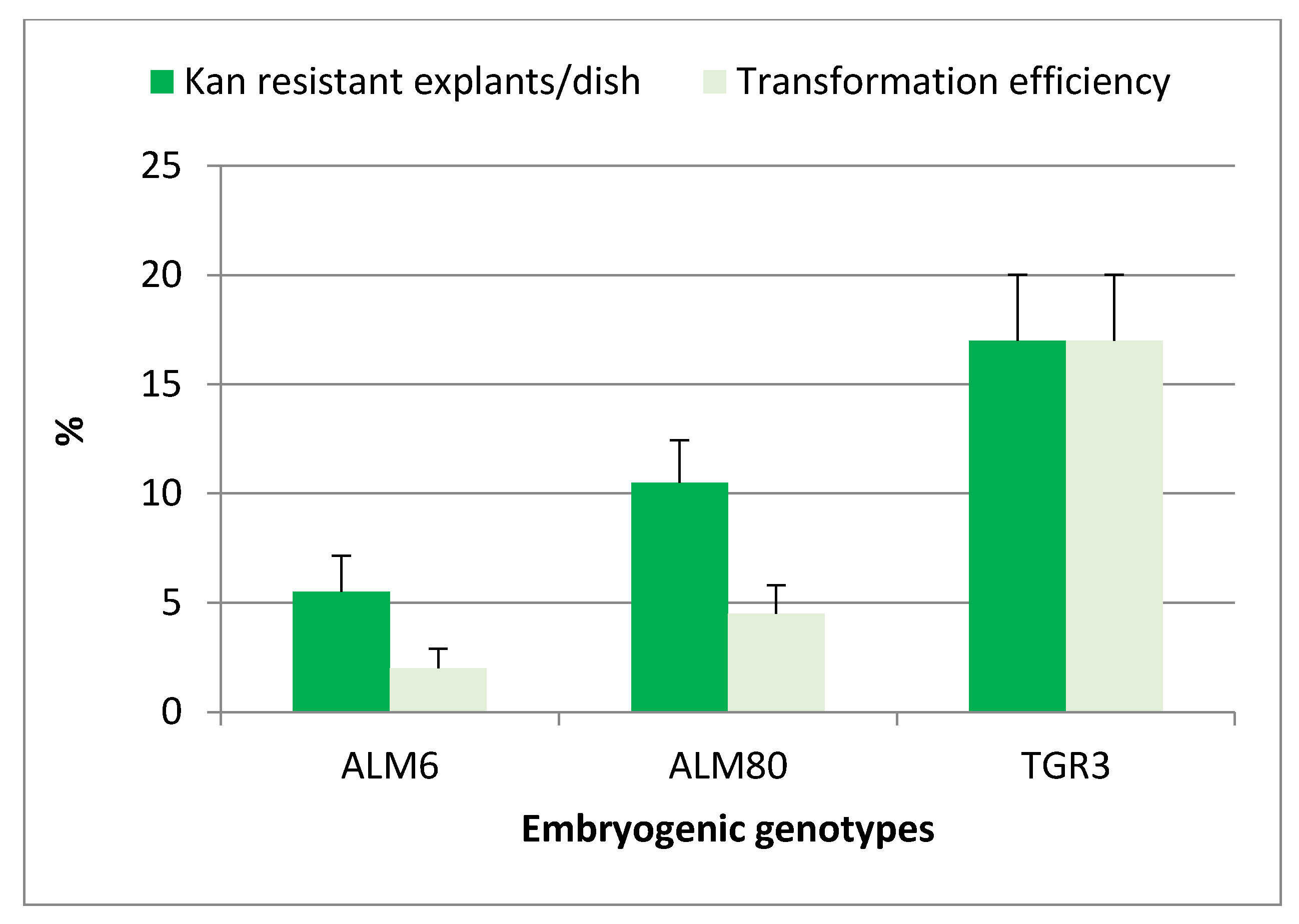
2.2. Genetic Transformation of Somatic Embryos
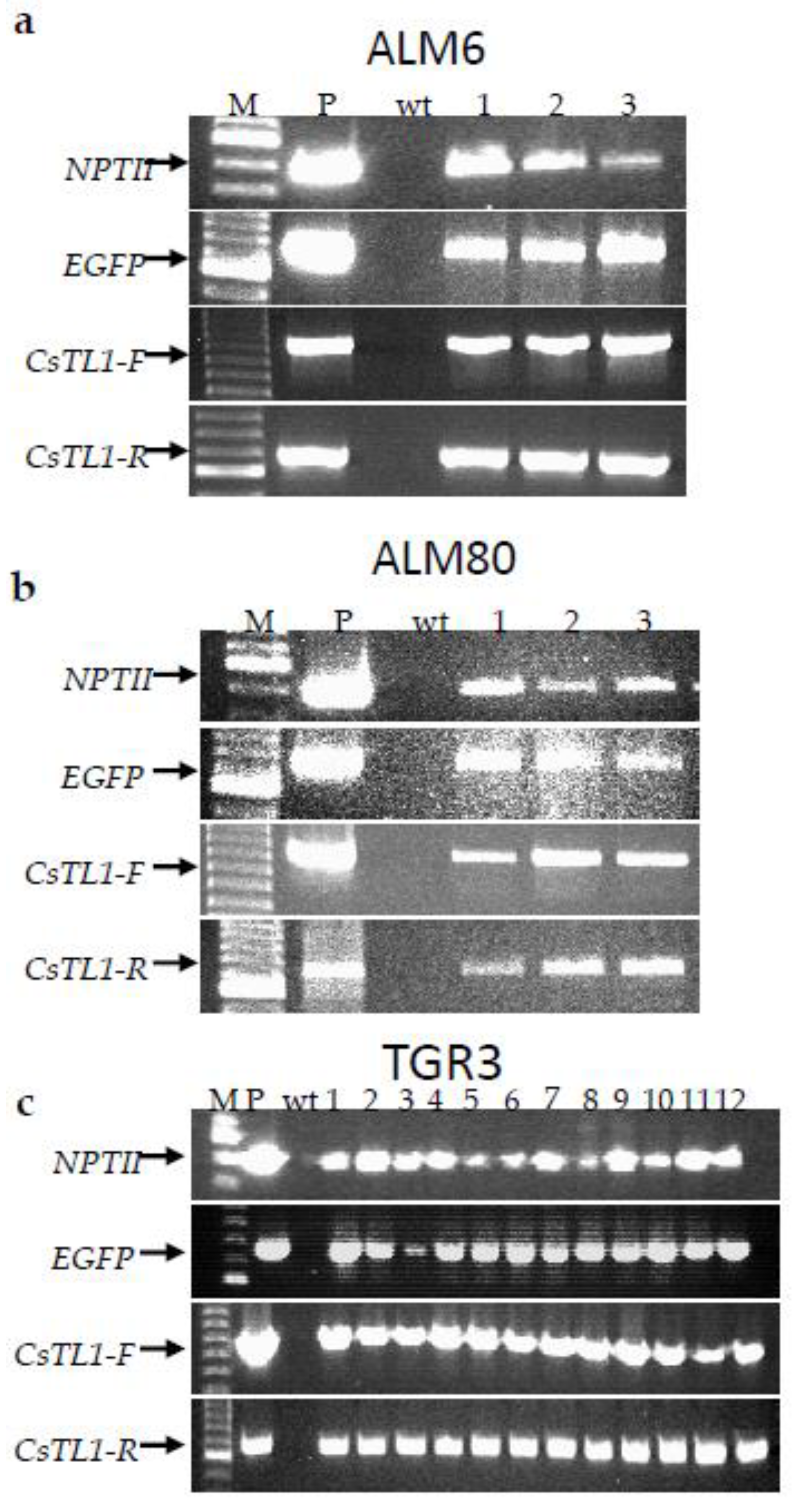
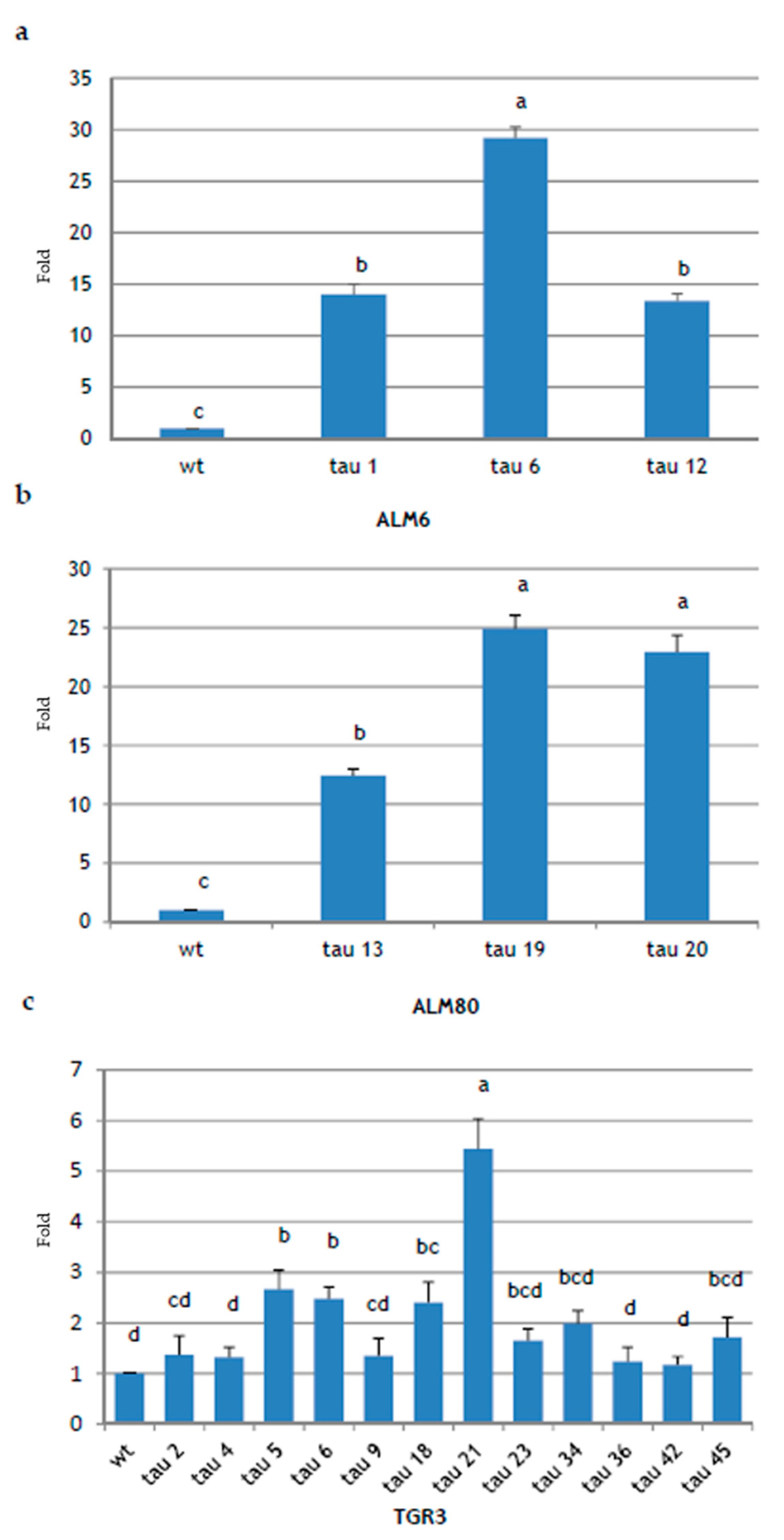
2.3. Molecular Analysis of Transgenic Lines
2.4. Cryopreservation of Somatic Embryos
2.5. Analysis of the Regeneration Ability of Transgenic Embryogenic Lines
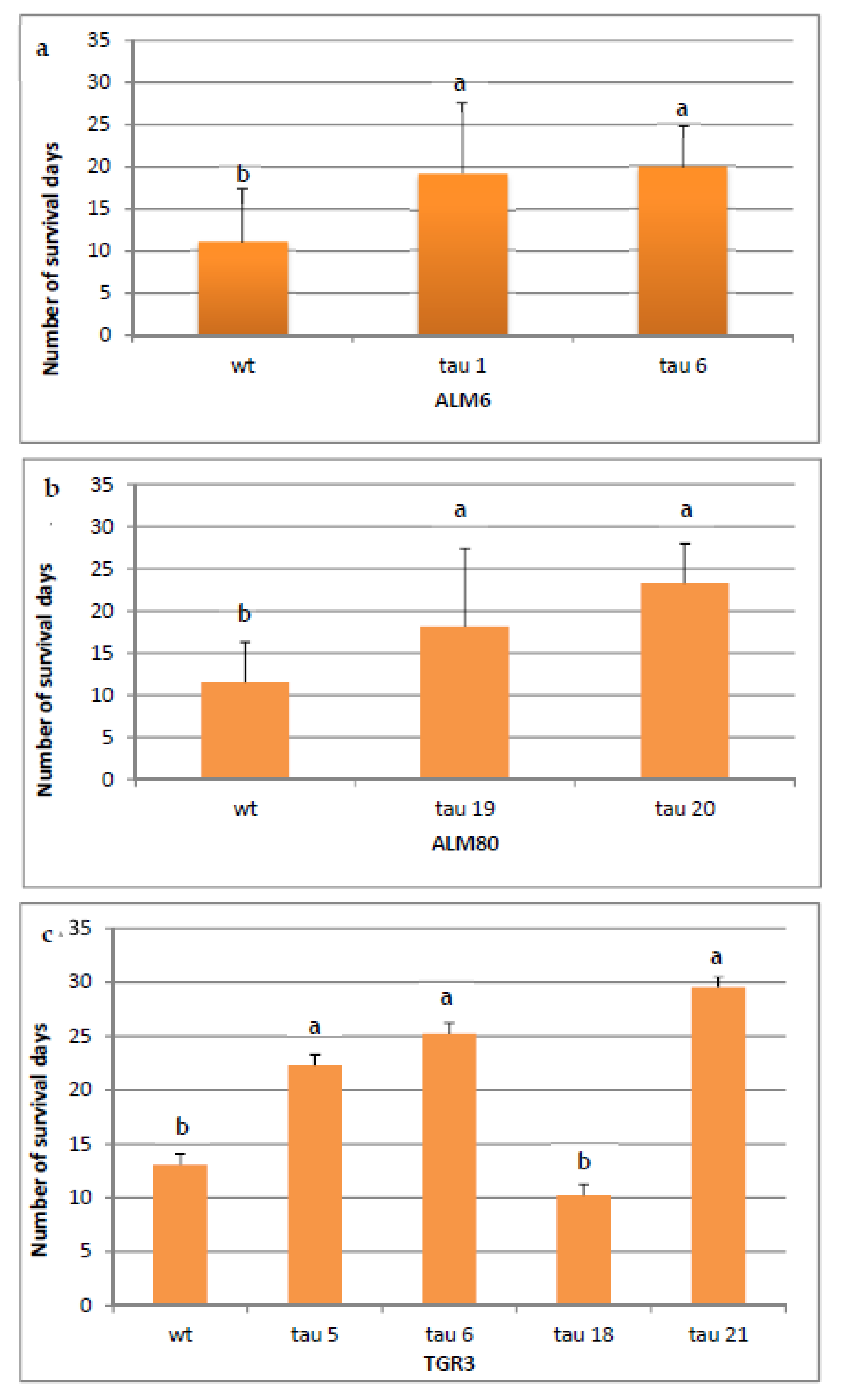
2.6. In Vitro Test of Tolerance to P. cinnamomi
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Binary Vector and Agrobacterium Strain
4.3. Determination of Phytotoxic Levels of Antibiotics
4.4. Agrobacterium Transformation Procedures
4.5. Evaluation of GFP Expresion and Establishment of Transgenic Lines
4.6. Molecular Characterization of Transgenic Embryogenic Lines
4.6.1. PCR Analysis
4.6.2. Quantification of CsTL1 Overexpression by Semi-Quantitative Real-Time PCR
4.7. Cryopreservation of Transgenic Lines
4.8. Analysis of the Regeneration Ability of Transgenic Embryogenic Lines
4.9. P. cinnamomi Infection Bioassay
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gil, L. Cork: A Strategic Material. Front. Chem. 2014, 2, 1–2. [Google Scholar] [CrossRef]
- Sierra-Pérez, J.; Boschmonart-Rives, J.; Gabarrell, X.; Durany, X.G. Production and Trade Analysis in the Iberian Cork Sector: Economic Characterization of a Forest Industry. Resour. Conserv. Recycl. 2015, 98, 55–66. [Google Scholar] [CrossRef]
- Tiberi, R.; Branco, M.; Bracalini, M.; Croci, F.; Panzavolta, T. Cork Oak Pests: A Review of Insect Damage and Management. Ann. For. Sci. 2016, 73, 219–232. [Google Scholar] [CrossRef]
- Agrios, G.N. Control of Plant Diseases. In Plant Pathology, 4th ed.; Academic Press: San Diego, CA, USA, 1997; pp. 200–216. [Google Scholar]
- Brasier, C. Phytophthora cinnamomi and Oak Decline in Southern Europe. Environmental Constraints Including Climate Change. Ann. Sci. For. 1996, 53, 347–358. [Google Scholar] [CrossRef]
- Moreira, A.C.; Medeira, C.; Maia, I.; Quartin, V.; Matos, M.C.; Cravador, A. Studies on the Association of the Quercus suber Decline Disease with Phytophthora cinnamomi in Portugal. Bol. Inf. CIDEU 2006, 1, 31–38. [Google Scholar]
- Corcobado, T.; Cubera, E.; Moreno, G.; Solla, A. Quercus ilex Forests Are Influenced by Annual Variations in Water Table, Soil Water Deficit and Fine Root Loss Caused by Phytophthora cinnamomi. Agric. For. Meteorol. 2013, 169, 92–99. [Google Scholar] [CrossRef]
- Vannini, A.; Valentini, R.; Luisi, N. Impact of Drought and Hypoxylon mediterraneum on Oak Decline in the Mediterranean Region. Annales des Sciences Forestières 1996, 53, 753–760. [Google Scholar] [CrossRef]
- Sánchez, M.E.; Venegas, J.; Romero, M.A.; Phillips, A.J.L.; Trapero, A. El Chancro de Encinas y Alcornoques Causado Por Botryosphaeria spp. en Andalucía. Bol. San. Veg. Plagas 2003, 29, 593–612. [Google Scholar]
- Alves, A.; Correia, A.; Luque, J.; Phillips, A.J. Botryosphaeria corticola, Sp. Nov. on Quercus Species, with Notes and Description of Botryosphaeria stevensii and Its Anamorph, Diplodia mutila. Mycologia 2004, 96, 598. [Google Scholar] [CrossRef]
- Jiménez, A.; Gallardo, A.; Antonietty, C.; Villagran, M.; Ocete, M.; Soria, F. Distribution of Coraebus undatus (Coleoptera: Buprestidae) in Cork Oak Forests of Southern Spain. Int. J. Pest Manag. 2012, 58, 281–288. [Google Scholar] [CrossRef]
- Martínez, M.T.; San-José, M.D.C.; Arrillaga, I.; Cano, V.; Morcillo, M.; Cernadas, M.J.; Corredoira, E. Holm Oak Somatic Embryogenesis: Current Status and Future Perspectives. Front. Plant Sci. 2019, 10, 239. [Google Scholar] [CrossRef] [PubMed]
- Wally, O.; Punja, Z. Genetic Engineering for Increasing Fungal and Bacterial Disease Resistance in Crop Plants. GM Crop. 2010, 1, 199–206. [Google Scholar] [CrossRef]
- Wani, S.H. Inducing Fungus-Resistance into Plants through Biotechnology. Not. Sci. Biol. 2010, 2, 14–21. [Google Scholar] [CrossRef]
- Jain, D.; Khurana, J.P. Role of Pathogenesis-Related (PR) Proteins in Plant Defense Mechanism. In Molecular Aspects of Plant-Pathogen Interaction; Singh, A., Singh, I., Eds.; Springer: Singapore, 2018; pp. 265–281. [Google Scholar]
- Veluthakkal, R.; Dasgupta, M.G. Pathogenesis-Related Genes and Proteins in Forest Tree Species. Trees 2010, 24, 993–1006. [Google Scholar] [CrossRef]
- Liu, J.-J.; Sturrock, R.; Ekramoddoullah, A.K.M. The Superfamily of Thaumatin-Like Proteins: Its Origin, Evolution, and Expression Towards Biological Function. Plant Cell Rep. 2010, 29, 419–436. [Google Scholar] [CrossRef]
- Vanjildorj, E.; Bae, T.-W.; Riu, K.-Z.; Yun, P.-Y.; Park, S.-Y.; Lee, C.-H.; Kim, S.-Y.; Lee, H.-Y. Transgenic Agrostis mongolica Roshev. with Enhanced Tolerance to Drought and Heat Stresses Obtained from Agrobacterium-Mediated Transformation. Plant Cell, Tissue Organ Cult. (PCTOC) 2006, 87, 109–120. [Google Scholar] [CrossRef]
- Schestibratov, K.; Dolgov, S. Transgenic Strawberry Plants Expressing a Thaumatin II Gene Demonstrate Enhanced Resistance to Botrytis cinerea. Sci. Hortic. 2005, 106, 177–189. [Google Scholar] [CrossRef]
- Mackintosh, C.A.; Lewis, J.; Radmer, L.E.; Shin, S.; Heinen, S.J.; Smith, L.A.; Wyckoff, M.N.; Dill-Macky, R.; Evans, C.K.; Kravchenko, S.; et al. Overexpression of Defense Response Genes in Transgenic Wheat Enhances Resistance to Fusarium Head Blight. Plant Cell Rep. 2006, 26, 479–488. [Google Scholar] [CrossRef]
- He, R.; Wu, J.; Zhang, Y.; Agüero, C.B.; Li, X.; Liu, S.; Wang, C.; Walker, M.A.; Lu, J. Overexpression of a Thaumatin-like Protein Gene from Vitis amurensis Improves Downy Mildew Resistance in Vitis vinifera Grapevine. Protoplasma 2016, 254, 1579–1589. [Google Scholar] [CrossRef] [PubMed]
- Ojola, P.O.; Nyaboga, E.N.; Njiru, P.N.; Orinda, G. Overexpression of Rice Thaumatin-like Protein (Ostlp) Gene in Transgenic Cassava Results in Enhanced Tolerance to Colletotrichum gloeosporioides F. Sp. Manihotis. J. Genet. Eng. Biotechnol. 2018, 16, 125–131. [Google Scholar] [CrossRef]
- Allona, I.; Collada, C.; Casado, R.; Paz-Ares, J.; Aragoncillo, C. Bacterial Expression of an Active Class Ib Chitinase from Castanea sativa Cotyledons. Plant Mol. Biol. 1996, 32, 1171–1176. [Google Scholar] [CrossRef] [PubMed]
- Pernas, M.; López-Solanilla, E.; Sánchez-Monge, R.; Salcedo, G.; Rodríguez-Palenzuela, P. Antifungal Activity of a Plant Cystatin. Mol. Plant-Microbe Interact. 1999, 12, 624–627. [Google Scholar] [CrossRef]
- Garcia-Casado, G.; Collada, C.; Allona, I.; Soto, A.; Casado, R.; Rodriguez-Cerezo, E.; Gomez, L.; Aragoncillo, C. Characterization of an Apoplastic Basic Thaumatin-like Protein from Recalcitrant Chestnut Seeds. Physiol. Plant. 2000, 110, 172–180. [Google Scholar] [CrossRef]
- Alonso, P.; Cortizo, M.; Celestino, C.; Toribio, M. Genetic Transformation of Selected Mature Cork Oak (Quercus suber L.) Trees. Plant Cell Rep. 2004, 23, 218–223. [Google Scholar] [CrossRef]
- Alvarez, R.; Ordás, R. Improved Genetic Transformation Protocol for Cork Oak (Quercus suber L.). Plant Cell Tissue Organ Cult. (PCTOC) 2007, 91, 45–52. [Google Scholar] [CrossRef]
- Álvarez, R.; Ordás, R.J. Sensibilidad a la Kanamicina Y Transformación Genética de Embriones Somáticos de Alcornoque (Quercus suber L.). Cuad. Soc. Esp. Cienc. For. 2008, 24, 101–106. [Google Scholar]
- Sánchez, N.; Manzanera, J.A.; Pintos, B.; Bueno, M.A. Agrobacterium-Mediated Transformation of Cork Oak (Quercus suber L.) Somatic Embryos. New For. 2005, 29, 169–176. [Google Scholar] [CrossRef]
- Álvarez, R.; Álvarez, J.M.; Humara, J.M.; Revilla, Á.; Ordás, R.J. Genetic Transformation of Cork Oak (Quercus suber L.) for Herbicide Resistance. Biotechnol. Lett. 2009, 31, 1477–1483. [Google Scholar] [CrossRef]
- Burgos, L. Transformation of Fruit Trees. Useful Breeding Tool or Continued Future Prospect? Transgenic Res. 2005, 14, 15–26. [Google Scholar] [CrossRef]
- Da Silva, J.; Fukai, S. The Impact of Carbenicillin, Cefotaxime and Vancomycin on Chrysanthemum and Tobacco TCL Morphogenesis and Agrobacterium Growth. J. Appl. Hortic. 2001, 3, 3–12. [Google Scholar] [CrossRef]
- Srinivasan, C.; Scorza, R. Transformation of Somatic Embryos of Fruit Trees and Grapevine. In Endophytes of Forest Trees; Springer Nature: London, UK, 1999; Volume 5, pp. 313–330. [Google Scholar]
- Padilla, I.M.; Burgos, L. Aminoglycoside Antibiotics: Structure, Functions and Effects on in Vitro Plant Culture and Genetic Transformation Protocols. Plant Cell Rep. 2010, 29, 1203–1213. [Google Scholar] [CrossRef] [PubMed]
- Leslie, C.A.; Walawage, S.L.; Uratsu, S.L.; McGranahan, G.; Dandekar, A.M. Walnut (Juglans). In Methods Molecular Biology: Agrobacterium protocols; Wang, K., Ed.; Springer: New York, NY, USA, 2015; Volume 2, pp. 297–312. [Google Scholar]
- Pérez-Barranco, G.; Torreblanca, R.; Padilla, I.M.G.; Sánchez-Romero, C.; Pliego-Alfaro, F.; Mercado, J. Studies on Genetic Transformation of Olive (Olea europaea L.) Somatic Embryos: I. Evaluation of Different Aminoglycoside Antibiotics for nptII Selection; II. Transient Transformation via Particle Bombardment. Plant Cell, Tissue Organ Cult. (PCTOC) 2009, 97, 243–251. [Google Scholar] [CrossRef]
- Palomo-Ríos, E.; Barceló-Muñoz, A.; Mercado, J.; Pliego-Alfaro, F. Evaluation of Key Factors Influencing Agrobacterium-Mediated Transformation of Somatic Embryos of Avocado (Persea americana Mill.). Plant Cell Tissue Organ Cult. (PCTOC) 2011, 109, 201–211. [Google Scholar] [CrossRef]
- Mondal, T.; Bhattacharya, A.; Ahuja, P.; Chand, P. Transgenic Tea [Camellia sinensis (L.) O. Kuntze cv. Kangra Jat] Plants Obtained by Agrobacterium-Mediated Transformation of Somatic Embryos. Plant Cell Rep. 2001, 20, 712–720. [Google Scholar] [CrossRef]
- Vidal, N.; Mallon, R.; Valladares, S.; Meijomín, A.M.; Vieitez, A.M. Regeneration of Transgenic Plants by Agrobacterium-Mediated Transformation of Somatic Embryos of Juvenile and Mature Quercus robur. Plant Cell Rep. 2010, 29, 1411–1422. [Google Scholar] [CrossRef] [PubMed]
- Nauerby, B.; Billing, K.; Wyndaele, R. Influence of the Antibiotic Timentin on Plant Regeneration Compared to Carbenicillin and Cefotaxime in Concentrations Suitable for Elimination of Agrobacterium tumefaciens. Plant Sci. 1997, 123, 169–177. [Google Scholar] [CrossRef]
- Yu, T.A.; Yeh, S.D.; Yang, J.S. Effects of Carbenicillin and Cefotaxime on Callus Growth and Somatic Embryogenesis from Ad-Ventitious Roots of Papaya. Bot. Bull. Acad. Sin. 2001, 42, 281–286. [Google Scholar] [CrossRef]
- Qin, Y.H.; Da Silva, J.A.T.; Bi, J.H.; Zhang, S.L.; Hu, G.B. Response of In Vitro Strawberry to Antibiotics. Plant Growth Regul. 2011, 65, 183–193. [Google Scholar] [CrossRef]
- Chauhan, R.D.; Veale, A.; Cathleen, M.; Strauss, S.H.; Myburg, A.A. Genetic Transformation of Eucalyptus-Challenges and Future Prospects. In Tree Biotechnology; Ramawat, K.G., Mérillon, J.M., Ahuja, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 392–425. [Google Scholar]
- Khan, E.U.; Fu, X.-Z.; Liu, J. Agrobacterium-Mediated Genetic Transformation and Regeneration of Transgenic Plants Using Leaf Segments as Explants in Valencia Sweet Orange. Plant Cell Tissue Organ Cult. (PCTOC) 2012, 109, 383–390. [Google Scholar] [CrossRef]
- Corredoira, E.; José, M.C.S.; Vieitez, A.M.; Allona, I.; Aragoncillo, C.; Ballester, A. Agrobacterium-Mediated Transformation of European Chestnut Somatic Embryos with a Castanea sativa (Mill.) Endochitinase Gene. New For. 2016, 47, 669–684. [Google Scholar] [CrossRef]
- De Oliveira-Cauduro, Y.; Adamuchio, L.G.; Filho, J.C.B.; Gerhardt, I.R.; Degenhardt, J.; Bernardes, M.; Quoirin, M. Optimization of Factors Affecting the Agrobacterium Tumefaciens-Mediated Transformation of Eucalyptus saligna. Rev. Árvore 2018, 41. [Google Scholar] [CrossRef]
- Cano, V.; Martínez, M.T.; José, M.C.S.; Couselo, J.L.; Varas, E.; Bouza-Morcillo, L.; Toribio, M.; Corredoira, E. Regeneration of Transgenic Plants by Agrobacterium-Mediated Transformation of Quercus ilex L. Somatic Embryos with the Gene CsTL1. New For. 2020, 51, 1003–1021. [Google Scholar] [CrossRef]
- Corredoira, E.; Valladares, S.; Allona, I.; Aragoncillo, C.; Vieitez, A.M.; Ballester, A. Genetic Transformation of European Chestnut Somatic Embryos with a Native Thaumatin-Like Protein (CsTL1) Gene Isolated from Castanea sativa Seeds. Tree Physiol. 2012, 32, 1389–1402. [Google Scholar] [CrossRef][Green Version]
- Mallon, R.; Valladares, S.; Corredoira, E.; Vieitez, A.M.; Vidal, N. Overexpression of the Chestnut CsTL1 Gene Coding for a Thaumatin-like Protein in Somatic Embryos of Quercus robur. Plant Cell Tissue Organ Cult. (PCTOC) 2013, 116, 141–151. [Google Scholar] [CrossRef]
- Zhang, B.; Oakes, A.D.; Newhouse, A.E.; Baier, K.M.; Maynard, C.A.; Powell, W. A Threshold Level of Oxalate Oxidase Transgene Expression Reduces Cryphonectria parasitica-Induced Necrosis in a Transgenic American Chestnut (Castanea dentata) Leaf Bioassay. Transgenic Res. 2013, 22, 973–982. [Google Scholar] [CrossRef]
- Bandopadhyay, R.; Haque, I.; Singh, D.; Mukhopadhyay, K. Levels and Stability of Expression of Transgenes. In Transgenic Crop Plants; Kole, C., Michler, C., Abbott, A.G., Hall, T.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 1, pp. 145–186. [Google Scholar]
- Amoah, B.; Wu, H.; Sparks, C.; Jones, H.D. Factors Influencing Agrobacterium-Mediated Transient Expression of UidA in Wheat Inflorescence Tissue. J. Exp. Bot. 2001, 52, 1135–1142. [Google Scholar] [CrossRef]
- Meyer, A.D.; Ichikawa, T.; Meins, F. Horizontal Gene Transfer: Regulated Expression of a Tobacco Homologue of the Agrobacterium rhizogenes RolC Gene. Mol. Genet. Genom. 1995, 249, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Cervera, M.; Pina, J.A.; Juarez, J.; Navarro, L.; Peña, L. A Broad Exploration of a Transgenic Population of Citrus: Stability of Gene Expression and Phenotype. Theor. Appl. Genet. 2000, 100, 670–677. [Google Scholar] [CrossRef]
- Raveendar, S.; Premkumar, A.; Sasikumar, S.; Ignacimuthu, S.; Agastian, P. Transgene and Its Expression in Transgenic Plants: A Review. Plant Cell Biotechnol. Mol. Biol. 2007, 8, 105–112. [Google Scholar]
- Hraška, M.; Rakouský, S.; Čurn, V. Green Fluorescent Protein as a Vital Marker for Non-destructive Detection of Transformation Events in Transgenic Plants. Plant Cell Tissue Organ Cult. (PCTOC) 2006, 86, 303–318. [Google Scholar] [CrossRef]
- Haseloff, J.; Amos, B. GFP in Plants. Trends Genet. 1995, 11, 328–329. [Google Scholar] [CrossRef]
- Dutt, M.; Grosser, J.W. An Embryogenic Suspension Cell Culture System for Agrobacterium-Mediated Transformation of Citrus. Plant Cell Rep. 2010, 29, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Palomo-Ríos, E.; Cerezo, S.; Mercado, J.A.; Pliego-Alfaro, F. Agrobacterium-Mediated Transformation of Avocado (Persea americana Mill.) Somatic Embryos with Fluorescent Marker Genes and Optimization of Transgenic Plant Recovery. Plant Cell Tissue Organ Cult. (PCTOC) 2016, 128, 447–455. [Google Scholar] [CrossRef]
- Sun, Z.-L.; Li, X.-W.; Zhou, W.; Yan, J.-D.; Gao, Y.-R.; Sun, J.-C.; Fang, K.-F.; Zhang, Q.; Xing, Y.; Qin, L.; et al. Agrobacterium-Mediated Genetic Transformation of Chinese Chestnut (Castanea mollissima Blume). Plant Cell Tissue Organ Cult. (PCTOC) 2020, 140, 95–103. [Google Scholar] [CrossRef]
- Corredoira, E.; Martínez, M.T.; Sanjosé, M.C.; Ballester, A. Conservation of Hardwood Forest Species. In Sustainable Development and Biodiversity; Springer Nature: London, UK, 2017; Volume 17, pp. 421–453. [Google Scholar]
- Valladares, S.; Toribio, M.; Celestino, C.; Vieitez, A.M. Cryopreservation of Embryogenic Cultures from Mature Quercus suber Trees Using Vitrification. Cryo Lett. 2004, 25, 177–186. [Google Scholar]
- Ryynänen, L.; Sillanpää, M.; Kontunen-Soppela, S.; Tiimonen, H.; Kangasjärvi, J.; Vapaavuori, E.; Häggman, H. Preservation of Transgenic Silver Birch (Betula pendula Roth) Lines by Means of Cryopreservation. Mol. Breed. 2002, 10, 143–152. [Google Scholar] [CrossRef]
- Jokipii-Lukkari, S.; Ryynänen, L.; Kallio, P.T.; Aronen, T.; Häggman, H. A Cryopreservation Method Maintaining the Genetic Fidelity of a Model Forest Tree, Populus tremula L. × Populus tremuloides Michx. Plant Sci. 2004, 166, 799–806. [Google Scholar] [CrossRef]
- Corredoira, E.; Merkle, S.A.; Martínez, M.T.; Toribio, M.; Canhoto, J.M.; Correia, S.I.; Ballester, A.; Vieitez, A.M. Non-Zygotic Embryogenesis in Hardwood Species. Crit. Rev. Plant Sci. 2019, 38, 29–97. [Google Scholar] [CrossRef]
- Raharjo, S.H.T.; Witjaksono; Gomez-Lim, M.A.; Padilla, G.; Litz, R.E. Recovery of avocado (Persea americana Mill.) Plants Transformed with the Antifungal Plant Defensin Gene PDF1.2. Vitr. Cell. Dev. Biol.—Anim. 2008, 44, 254–262. [Google Scholar] [CrossRef]
- Urtubia, C.; Devia, J.; Castro, Á.; Zamora, P.; Aguirre, C.; Tapia, E.; Barba, P.; Dell‘Orto, P.; Moynihan, M.R.; Petri, C.; et al. Agrobacterium-Mediated Genetic Transformation of Prunus salicina. Plant Cell Rep. 2008, 27, 1333–1340. [Google Scholar] [CrossRef]
- Chetty, V.J.; Ceballos, N.; Garcia, D.; Narváez-Vásquez, J.; Lopez, W.; Orozco-Cárdenas, M.L. Evaluation of Four Agrobacterium Tumefaciens Strains for the Genetic Transformation of Tomato (Solanum lycopersicum L.) Cultivar Micro-Tom. Plant Cell Rep. 2012, 32, 239–247. [Google Scholar] [CrossRef]
- Maynard, C.A.; Powell, W.A.; Polin-McGuigan, L.D.; Viéitez, A.M.; Ballester, A.; Corredoira, E.; Merkle, S.A.; Andrade, G.M. Chestnut. In Compendium of Transgenic Crop Plants; Wiley: Hoboken, NJ, USA, 2008; pp. 169–192. [Google Scholar]
- Hernandez, I.M.T.; Celestino, C.; Alegre, J.; Toribio, M.L. Vegetative Propagation of Quercus suber L. by Somatic Embryogenesis. II. Plant regeneration from selected cork oak trees. Plant Cell Rep. 2003, 21, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Andrade, G.M.; Merkle, S.A. Enhancement of American Chestnut Somatic Seedling Production. Plant Cell Rep. 2005, 24, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Corredoira, E.; Valladares, S.; Vieitez, A.M.; Ballester, A. Improved Germination of Somatic Embryos and Plant Recovery of European Chestnut. Vitr. Cell. Dev. Biol.—Anim. 2008, 44, 307–315. [Google Scholar] [CrossRef]
- Martínez, M.T.; Vieitez, A.M.; Corredoira, E. Improved Secondary Embryo Production in Quercus alba and Q. Rubra by Activated Charcoal, Silver Thiosulphate and Sucrose: Influence of Embryogenic Explant Used for Subculture. Plant Cell, Tissue Organ Cult. (PCTOC) 2015, 121, 531–546. [Google Scholar] [CrossRef]
- Martínez, M.T.; José, M.C.S.; Vieitez, A.M.; Cernadas, M.J.; Allester, A.B.; Corredoira, E. Propagation of Mature Quercus ilex L. (Holm Oak) Trees by Somatic Embryogenesis. Plant Cell, Tissue Organ Cult. (PCTOC) 2017, 131, 321–333. [Google Scholar] [CrossRef]
- De Buck, S.; De Wilde, C.; Van Montagu, M.; Depicker, A. T-DNA Vector Backbone Sequences Are Frequently Integrated into the Genome of Transgenic Plants Obtained by Agrobacterium-Mediated Transformation. Mol. Breed. 2000, 6, 459–468. [Google Scholar] [CrossRef]
- Ballester, A.; Corredoira, E.; Vieitez, A.M. Limitations of Somatic Embryogenesis in Hardwood Trees. In Vegetative Propagation of Forest Trees; Park, Y.S., Bonga, J.M., Moon, H.K., Eds.; National Institute of Forest Science (NiFos): Seoul, Korea, 2016; pp. 56–74. [Google Scholar]
- Fagoaga, C.; Rodrigo, I.; Conejero, V.; Hinarejos, C.; Tuset, J.J.; Arnau, J.; Pina, J.A.; Navarro, L.; Peña, L. Increased Tolerance to Phytophthora citrophthora in Transgenic Orange Plants Constitutively Expressing a Tomato Pathogenesis Related Protein PR-5. Mol. Breed. 2001, 7, 175–185. [Google Scholar] [CrossRef]
- Liu, D.; He, X.; Li, W.; Chen, C.; Ge, F. Molecular Cloning of a Thaumatin-like Protein Gene from Pyrus pyrifolia and Overexpression of This Gene in Tobacco Increased Resistance to Pathogenic Fungi. Plant Cell Tissue Organ Cult. (PCTOC) 2012, 111, 29–39. [Google Scholar] [CrossRef]
- Mahdavi-Darvari, F.; Sariah, M.; Maziah, M. Expression of Rice Thaumatin-Like Protein Gene in Transgenic Banana Plants Enhances Resistance to Fusarium Wilt. Appl. Biochem. Biotechnol. 2011, 166, 1008–1019. [Google Scholar] [CrossRef]
- López-Gómez, R.; Cortés-Rodríguez, M.A.; Herbert-Moreno, P.C.; Sánchez-Pérez, J.L.; Vidales-Fernández, I.; Fernández-Pavía, S.; García-Chávez, A.; Salgado-Garciglia, R. Micropropagación Y Pruebas de Resistencia in Vitro a Phytophthora cinnamomi de Materiales de Aguacate Raza Mexicana. In Proceedings of the VI World Avocado Congress. Actas VI Congreso Mundial del Aguacate, Viña Del Mar, Chile, 12–16 November 2007; pp. 12–16. [Google Scholar]
- Cuenca, B.; Ocana, L.; Salinero, C.; Pintos, C.; Mansilla, J.; Rial, C. Selection of Castanea sativa Mill. for Resistance to Phytophthora cinnamomi: Micropropagation and Testing of Selected Clones. Acta Hortic. 2010, 866, 111–119. [Google Scholar] [CrossRef]
- Hernández, I.; Celestino, C.; Toribio, M. Vegetative Propagation of Quercus suber L. by Somatic Embryogenesis. I. Factors affecting the induction in leaves from mature cork oak trees. Plant Cell Rep. 2003, 21, 759–764. [Google Scholar] [CrossRef]
- Schenk, R.U.; Hildebrandt, A.C. Medium and Techniques for Induction and Growth of Monocotyledonous and Dicotyledonous Plant Cell Cultures. Can. J. Bot. 1972, 50, 199–204. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bioassays with Tobacco Tissue Cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Hood, E.E.; Gelvin, S.B.; Melchers, L.S.; Hoekema, A. New Agrobacterium Helper Plasmids for Gene Transfer to Plants. Transgenic Res. 1993, 2, 208–218. [Google Scholar] [CrossRef]
- Xu, R.; Li, Q.Q. Protocol: Streamline Cloning of Genes into Binary Vectors in Agrobacterium via the Gateway® TOPO Vector System. Plant Methods 2008, 4, 4–7. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning. A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989; p. 1546. [Google Scholar]
- Porth, I.; Koch, M.; Berenyi, M.; Burg, A.; Burg, K. Identification of Adaptation-Specify Differences in mRNA Expression of Sessile and Pedunculate Oak Based on Osmotic-Stress-Induced Genes. Tree Physiol. 2005, 25, 1317–1329. [Google Scholar] [CrossRef]
- Soler, M.; Serra, O.; Molinas, M.; García-Berthou, E.; Caritat, A.; Figueras, M. Seasonal Variation in Transcript Abundance in Cork Tissue Analyzed by Real Time RT-PCR. Tree Physiol. 2008, 28, 743–751. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, 3. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative CT Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Sakai, A.; Kobayashi, S.; Oiyama, I. Cryopreservation of Nucellar Cells of Navel Orange (Citrus sinensis Osb. var. brasiliensis Tanaka) by Vitrification. Plant Cell Rep. 1990, 9, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Toribio, M.; Celestino, C.; Molinas, M. Cork Oak, Quercus suber L. In Protocol for Somatic Embryogenesis in Woody Plants; Jain, S.M., Gupta, P.K., Eds.; Springer: Amsterdam, The Netherlands, 2005; pp. 445–457. [Google Scholar]
- Gresshoff, P.M.; Doy, C.H. Development and Differentiation of Haploid Lycopersicon esculentum (Tomato). Planta 1972, 107, 161–170. [Google Scholar] [CrossRef]
- Corcobado, T.; Cubera, E.; Juárez, E.; Moreno, G.; Solla, A. Drought Events Determine Performance of Quercus ilex Seedlings and Increase their Susceptibility to Phytophthora cinnamomi. Agric. For. Meteorol. 2014, 192–193, 1–8. [Google Scholar] [CrossRef]
- Chen, D.-W.; Zentmyer, G.A. Production of Sporangia by Phytophthora cinnamomi in Axenic Culture. Mycology 1970, 62, 397. [Google Scholar] [CrossRef]
- Wheller, T.; Erwin, D.C.; Ribeiro, O.K. Phytophthora Diseases Worldwide. Mycology 1998, 90, 1092. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotics | Explants with SSE (%) | Number of Bipolar SSE/Dish | ||
|---|---|---|---|---|
| (mg/L) | TGR3 | ALM80 | TGR3 | ALM80 |
| 0 | 100.0 ± 0.0 | 98.4 ± 1.5 | 20.5 ± 0.7 | 71.3 ± 3.3 |
| CF 200 | 100.0 ± 0.0 | 98.4 ± 1.5 | 19.6 ± 0.5 | 55.5 ± 3.7 |
| CF 300 | 100.0 ± 0.0 | 100.0 ± 0.0 | 18.5 ± 1.0 | 48.5 ± 2.6 |
| CB 300 | 100.0 ± 0.0 | 98.4 ± 1.5 | 20.5 ± 0.4 | 85.0 ± 9.7 |
| CF 200 + CB 200 | 100.0 ± 0.0 | 100.0 ± 0.0 | 18.0 ± 0.8 | 68.3 ± 6.4 |
| CF 200 + CB 300 | 100.0 ± 0.0 | 90.6 ± 2.9 | 18.3 ± 0.8 | 46.4 ± 3.2 |
| F-test | ns | ns | ns | p ≤ 0.05 |
| Line | Root Only | Shoot Only | Conversion (Shoot + Root) | |||
|---|---|---|---|---|---|---|
| (%) | (%) | (%) | RL (mm) | SL (mm) | NL | |
| ALM6-wt | 29.2 ± 10.9 | 0.0 ± 0.0 | 41.7 ± 7.6 | 85.2 ± 14.7 | 21.9 ± 4.7 | 4.1 ± 0.9 |
| ALM6-tau 1 | 25.0 ± 8.6 | 0.0 ± 0.0 | 19.4 ± 2.5 | 59.9 ± 18.0 | 18.5 ± 2.5 | 4.9 ± 1.6 |
| ALM6-tau 6 | 41.7 ± 12.9 | 0.0 ± 0.0 | 33.3 ± 11.2 | 57.9 ± 12.6 | 19.2 ± 3.0 | 5.7 ± 1.0 |
| ALM6-tau 12 | 55.6 ± 9.4 | 8.3 ± 3.4 | 13.9 ± 4.7 | 51.5 ± 8.0 | 30.6 ± 2.9 | 5.5 ± 0.2 |
| H test | ns | p ≤ 0.05 | p ≤ 0.05 | ns | ns | ns |
| ALM80-wt | 47.2 ± 7.3 | 2.8 ± 2.6 | 16.7 ± 3.9 | 93.2 ± 22.9 | 27.0 ± 14.0 | 6.2 ± 2.5 |
| ALM80-tau 13 | 47.2 ± 14.4 | 0.0 ± 0.0 | 2.8 ± 2.5 | 58.0 ± 0.0 | 7.0 ± 0.0 | 3.0 ± 0.0 |
| ALM80-tau 19 | 47.2 ± 7.3 | 0.0 ± 0.0 | 5.6 ± 5.1 | 105.0 ± 0.0 | 15.0 ± 0.0 | 4.0 ± 0.0 |
| ALM80-tau 20 | 44.4 ± 10.2 | 0.0 ± 0.0 | 8.3 ± 5.2 | 149.3 ± 33.1 | 7.5 ± 1.0 | 6.0 ± 0.4 |
| H test | ns | ns | ns | ns | ns | ns |
| TGR3-wt | 22.2 ± 9.4 | 5.6 ± 3.2 | 18.1 ± 9.7 | 34.8 ± 9.3 | 21.5 ± 2.5 | 6.7 ± 1.1 |
| TGR3-tau 2 | 27.8 ± 6.4 | 2.8 ± 2.5 | 13.9 ± 8.3 | 75.0 ± 12.3 | 28.2 ± 4.2 | 5.2 ± 0.5 |
| TGR3-tau 4 | 56.9 ± 10.9 | 0.0 ± 0.0 | 16.7 ± 7.9 | 60.6 ± 10.5 | 22.8 ± 5.1 | 9.2 ± 1.1 |
| TGR3-tau 5 | 38.9 ± 9.3 | 2.8 ± 0.0 | 16.7 ± 7.5 | 47.0 ± 24.4 | 8.7 ± 3.1 | 2.6 ± 1.0 |
| TGR3-tau 6 | 47.2 ± 10.4 | 2.8 ± 2.6 | 11.1 ± 7.6 | 54.3 ± 3.4 | 16.3 ± 1.5 | 5.5 ± 0.2 |
| TGR3-tau 9 | 43.1 ± 11.6 | 0.0 ± 0.0 | 5.6 ± 3.2 | 55.0 ± 8.2 | 5.0 ± 0.0 | 6.5 ± 1.0 |
| TGR3-tau 18 | 37.5 ± 8.1 | 2.8 ± 2.6 | 5.6 ± 5.1 | 110.0 ± 0.0 | 22.0 ± 0.0 | 5.0 ± 0.0 |
| TGR3-tau 21 | 23.6 ± 6.4 | 5.6 ± 3.2 | 34.7 ± 10.1 | 58.7 ± 23.2 | 12.2 ± 2.8 | 5.7 ± 0.8 |
| TGR3-tau 23 | 8.3 ± 5.2 | 4.2 ± 3.8 | 15.3 ± 6.4 | 22.3 ± 3.0 | 19.2 ± 1.8 | 3.3 ± 0.5 |
| TGR3-tau 34 | 41.7 ± 5.2 | 2.8 ± 2.5 | 16.7 ± 5.6 | 57.5 ± 13.0 | 11.9 ± 2.2 | 5.8 ± 0.5 |
| TGR3-tau 36 | 38.9 ± 14.7 | 0.0 ± 0.0 | 36.1 ± 6.7 | 83.6 ± 15.1 | 8.6 ± 0.4 | 3.5 ± 0.4 |
| TGR3-tau 42 | 36.1 ± 14.4 | 5.6 ± 3.2 | 11.1 ± 5.1 | 77.0 ± 10.6 | 13.3 ± 2.6 | 3.5 ± 0.2 |
| TGR3-tau 45 | 34.7 ± 9.3 | 0.0 ± 0.0 | 11.1 ± 7.6 | 27.8 ± 1.1 | 19.3 ± 2.6 | 6.5 ± 1.0 |
| H test | ns | ns | ns | ns | ns | ns |
| Line | Rooting (%) | Number of Roots |
|---|---|---|
| ALM6-wt | 86.7 ± 5.2 | 2.8 ± 0.3 |
| ALM6-tau 1 | 65.0 ± 6.9 | 2.3 ± 0.3 |
| ALM6-tau 6 | 93.3 ± 3.5 | 4.6 ± 0.6 |
| F-test | p ≤ 0.05 | p ≤ 0.05 |
| ALM80-wt | 75.0 ± 4.3 | 1.9 ± 0.1 |
| ALM80-tau 19 | 51.7 ± 3.7 | 1.6 ± 0.2 |
| ALM80-tau 20 | 86.7 ± 4.6 | 2.4 ± 0.2 |
| F-test | p ≤ 0.05 | p ≤ 0.05 |
| TGR3-wt | 76.7 ± 5.9 | 5.1 ± 0.5 |
| TGR3-tau 5 | 81.7 ± 5.0 | 5.6 ± 0.6 |
| TGR3-tau 6 | 91.7 ± 3.5 | 5.2 ± 0.5 |
| TGR3-tau 18 | 80.0 ± 5.7 | 4.7 ± 0.5 |
| TGR3-tau 21 | 98.3 ± 1.6 | 5.0 ± 0.3 |
| F-test | p ≤ 0.05 | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cano, V.; Martínez, M.T.; Couselo, J.L.; Varas, E.; Vieitez, F.J.; Corredoira, E. Efficient Transformation of Somatic Embryos and Regeneration of Cork Oak Plantlets with a Gene (CsTL1) Encoding a Chestnut Thaumatin-Like Protein. Int. J. Mol. Sci. 2021, 22, 1757. https://doi.org/10.3390/ijms22041757
Cano V, Martínez MT, Couselo JL, Varas E, Vieitez FJ, Corredoira E. Efficient Transformation of Somatic Embryos and Regeneration of Cork Oak Plantlets with a Gene (CsTL1) Encoding a Chestnut Thaumatin-Like Protein. International Journal of Molecular Sciences. 2021; 22(4):1757. https://doi.org/10.3390/ijms22041757
Chicago/Turabian StyleCano, Vanesa, Mᵃ Teresa Martínez, José Luis Couselo, Elena Varas, Francisco Javier Vieitez, and Elena Corredoira. 2021. "Efficient Transformation of Somatic Embryos and Regeneration of Cork Oak Plantlets with a Gene (CsTL1) Encoding a Chestnut Thaumatin-Like Protein" International Journal of Molecular Sciences 22, no. 4: 1757. https://doi.org/10.3390/ijms22041757
APA StyleCano, V., Martínez, M. T., Couselo, J. L., Varas, E., Vieitez, F. J., & Corredoira, E. (2021). Efficient Transformation of Somatic Embryos and Regeneration of Cork Oak Plantlets with a Gene (CsTL1) Encoding a Chestnut Thaumatin-Like Protein. International Journal of Molecular Sciences, 22(4), 1757. https://doi.org/10.3390/ijms22041757