
Treatment-Induced Neuropathy in Diabetes (TIND)—Developing a Disease Model in Type 1 Diabetic Rats

 , and
, and
Abstract
1. Introduction
2. Results
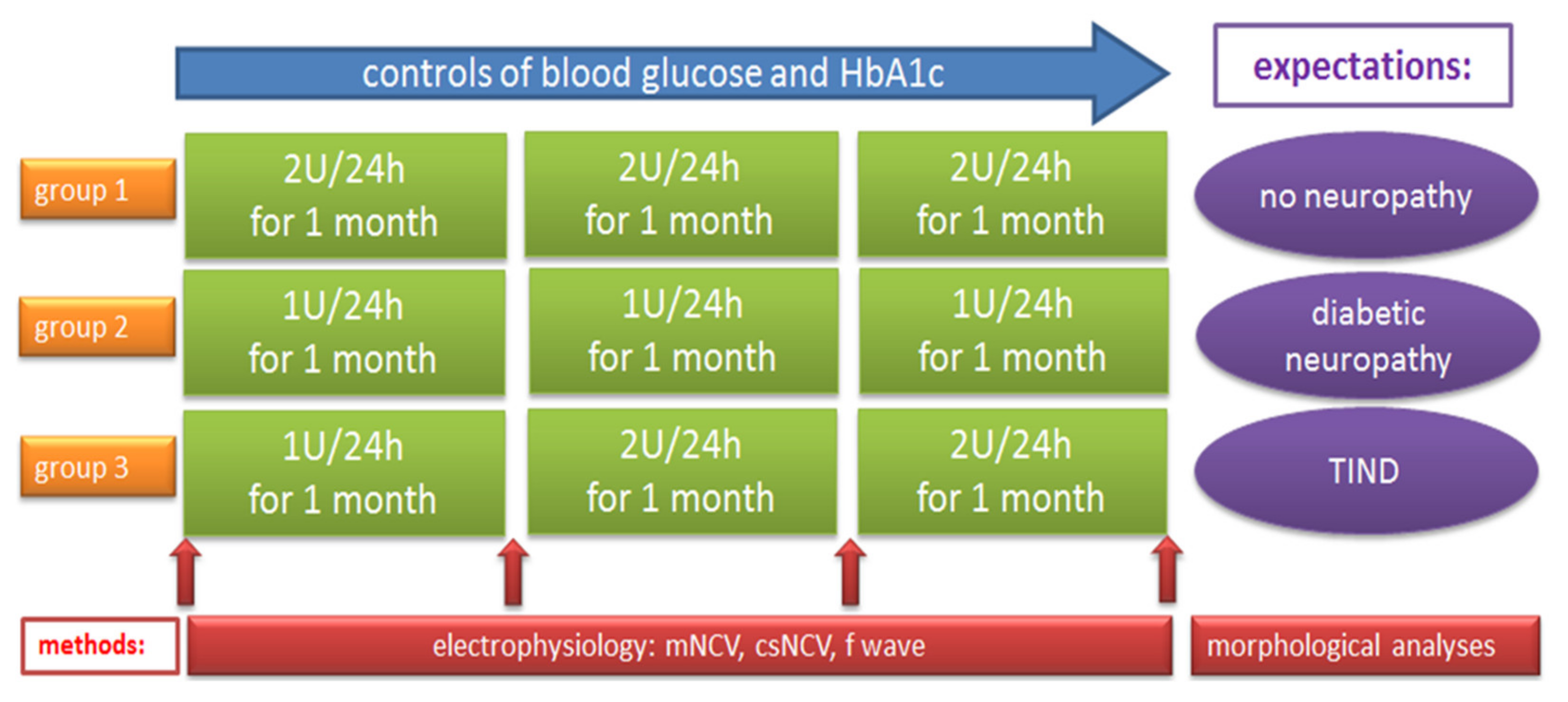
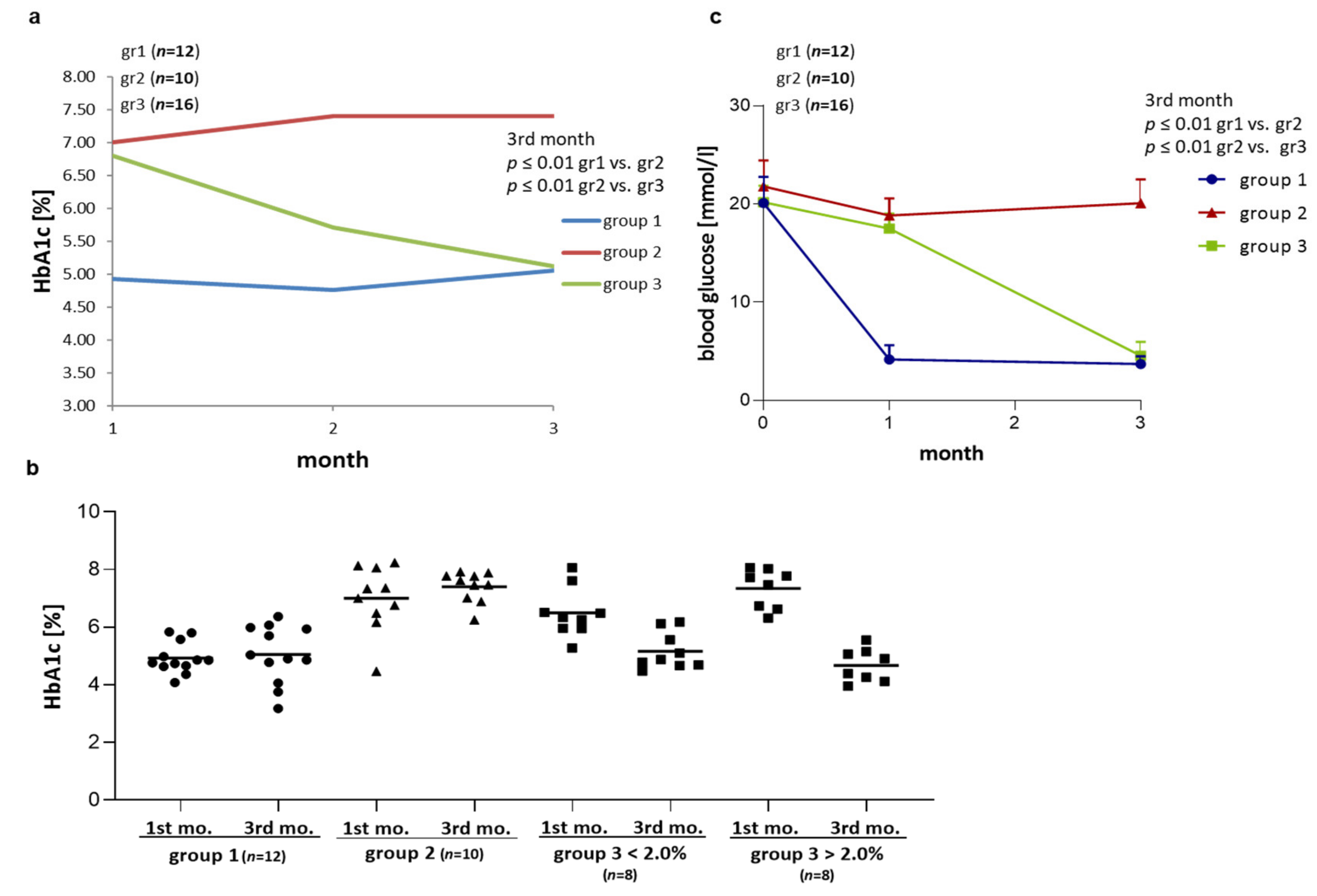
2.1. Effect of Different Modes of Insulin Tretment on HbA1c and Blood Glucose in BB/OKL Diabetic Rats
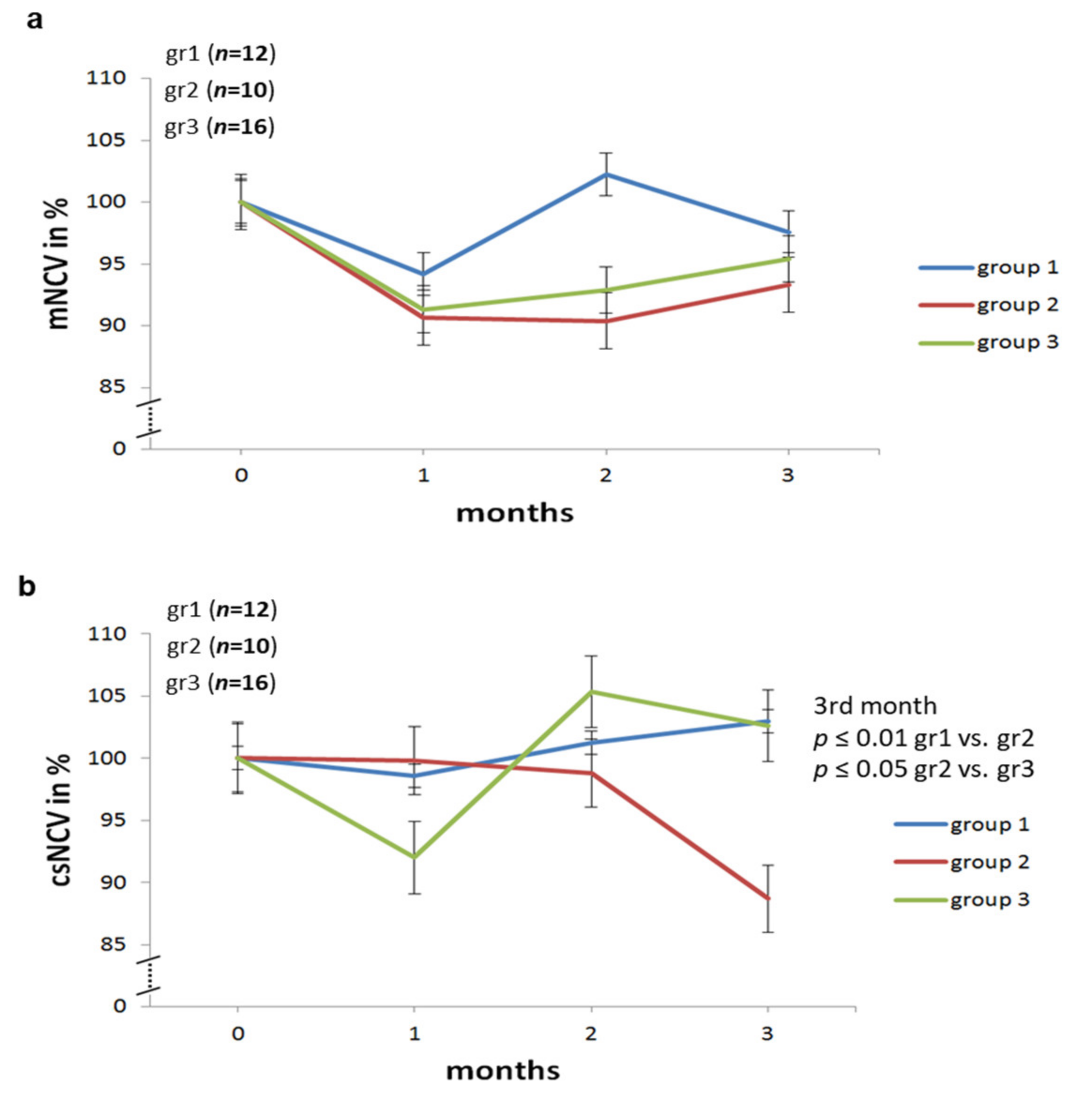
2.2. Effect of Different Modes of Insulin Treatment on Nerve Conduction Velocities, F Waves and Amplitudes of Sciatic Nerve in BB/OKL Diabetic Rats
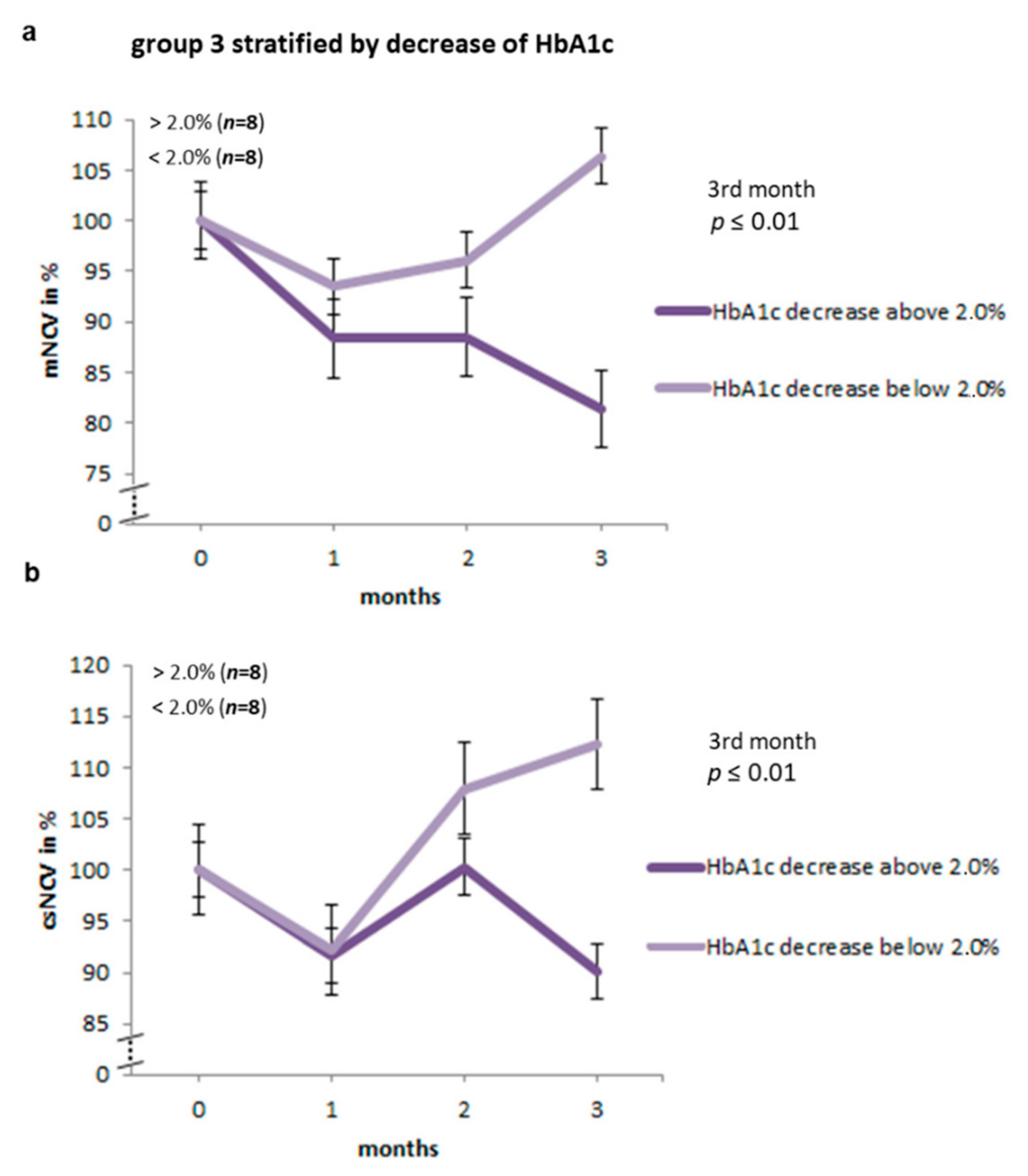
2.3. Nerve Conduction Studies Stratified by the Decrease in HbA1c Levels in Group 3 of BB/OKL Rats
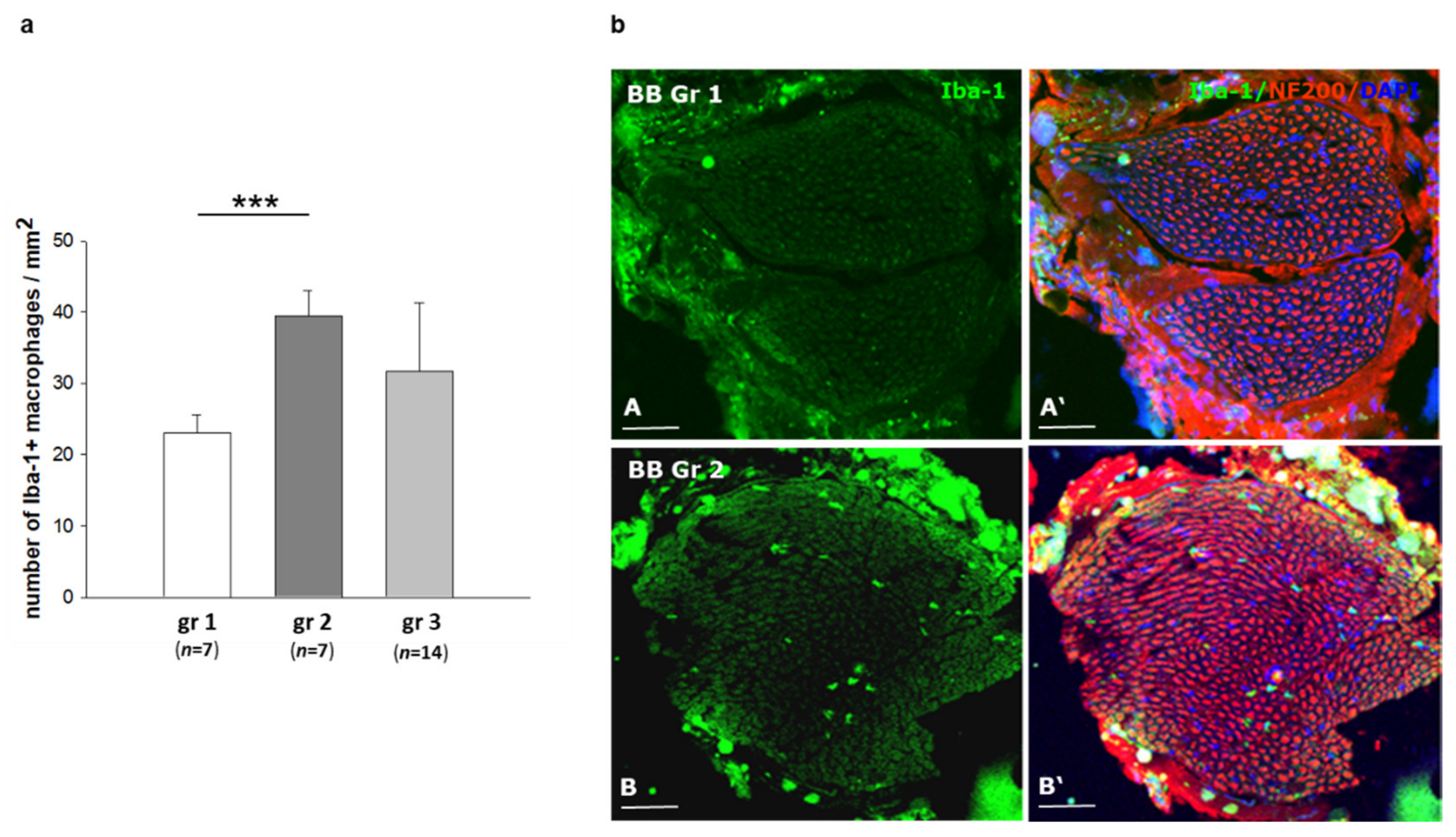
2.4. Infiltration of Inflammatory Cells in Sciatic Nerves of BB/OKL Rats after Different Modes of Insulin Treatment
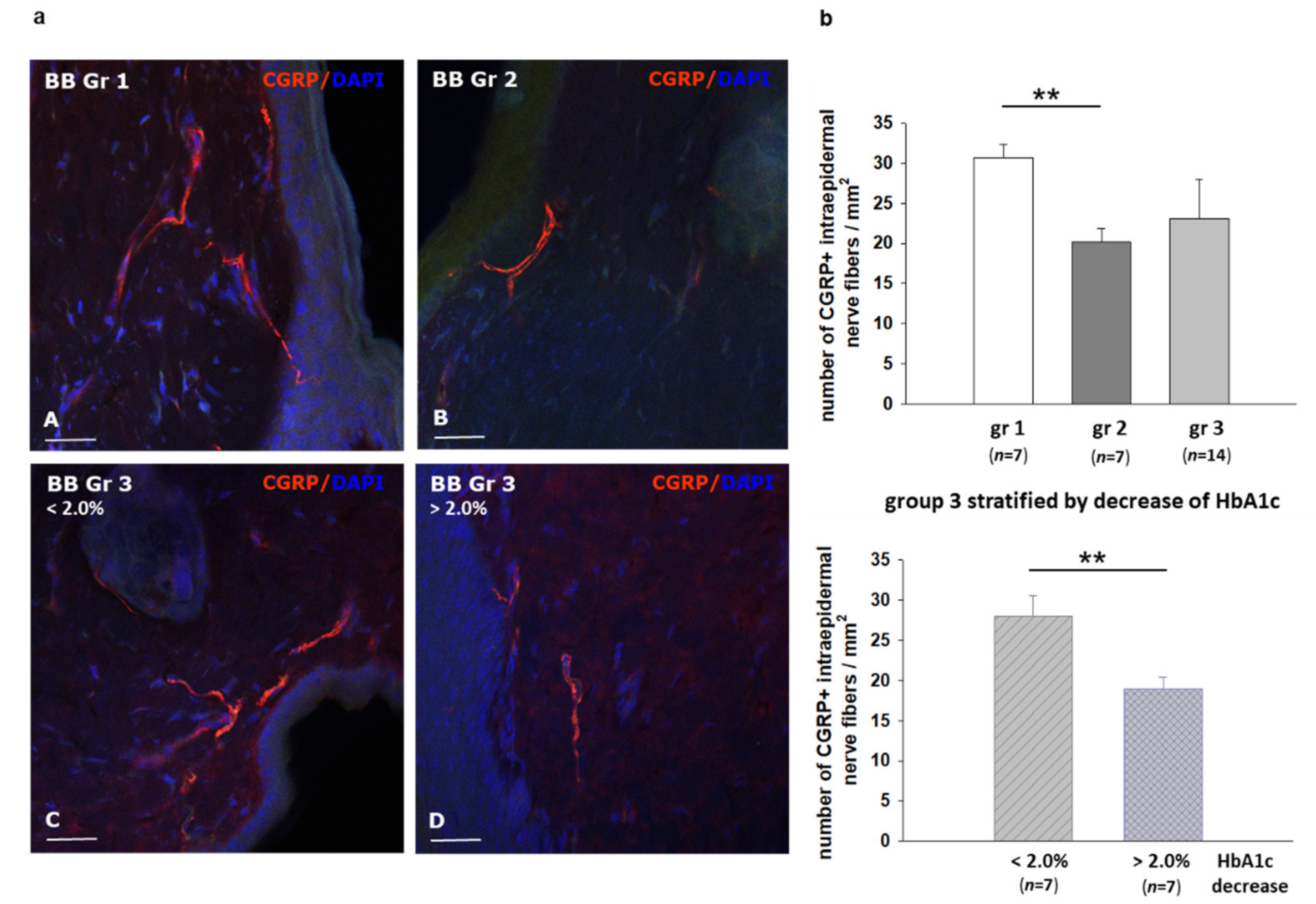
2.5. Effect of Different Modes of Insulin Treatment on Intraepidermal Nerve Fiber Density (IENFD) and Calcitonin Gene Related Peptide CGRP Expression in Skin Speciments of BB/OKL Rats
3. Discussion
4. Material and Methods
4.1. Animals and Treatments
4.2. Electroneurography
4.3. Immunostaining
4.4. Determination of Macrophages
4.5. Quantification of the Intraepidermal Nerve Fiber Density (IENFD) and CGRP-Positive Nerve Endings
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gibbons, C.H.; Freeman, R. Treatment-induced diabetic neuropathy: A reversible painful autonomic neuropathy. Ann. Neurol. 2010, 67, 534–541. [Google Scholar] [CrossRef]
- Gibbons, C.H.; Freeman, R. Treatment-induced neuropathy of diabetes: An acute, iatrogenic complication of diabetes. Brain 2015, 138, 43–52. [Google Scholar] [CrossRef]
- Storz, M.A. Treatment-induced Neuropathy of Diabetes: A Call for Acknowledgement. Diabet. Med. 2020, 37, 369–370. [Google Scholar] [CrossRef]
- Tesfaye, S.; Stevens, L.K.; Stephenson, J.M.; Fuller, J.H.; Plater, M.; Ionescu-Tirgoviste, C.; Nuber, A.; Pozza, G.; Ward, J.D. Prevalence of diabetic peripheral neuropathy and its relation to glycaemic control and potential risk factors: The EURODIAB IDDM Complications Study. Diabetologia 1996, 39, 1377–1384. [Google Scholar] [CrossRef]
- Fullerton, B.; Jeitler, K.; Seitz, M.; Horvath, K.; Berghold, A.; Siebenhofer, A. Intensive glucose control versus conventional glucose control for type 1 diabetes mellitus. Cochrane Database Syst. Rev. 2014, 14, CD009122. [Google Scholar] [CrossRef]
- Low, P.A.; Singer, W. Treatment-induced neuropathy of diabetes: An energy crisis? Brain 2015, 138, 2–3. [Google Scholar] [CrossRef][Green Version]
- Nicodemus, J.M.; Enriquez, C.; Marquez, A.; Anaya, C.J.; Jolivalt, C.G. Murine model and mechanisms of treatment-induced painful diabetic neuropathy. Neuroscience 2017, 354, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Marliss, E.B.; Nakhooda, A.F.; Poussier, P.; Sima, A.A. The diabetic syndrome of the ‘BB’ Wistar rat: Possible relevance to type 1 (insulin-dependent) diabetes in man. Diabetologia 1982, 22, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Sood, A.; Cunningham, C.; Lin, S. The BB Wistar Rat as a Diabetic Model for Fracture Healing. ISRN Endocrinol. 2013, 2013, 349604. [Google Scholar] [CrossRef]
- Heininger, K.; Stoll, G.; Linington, C.; Toyka, K.V.; Wekerle, H. Conduction failure and nerve conduction slowing in experimental allergic neuritis induced by P2-specific T-cell lines. Ann. Neurol. 1986, 19, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Kosacka, J.; Nowicki, M.; Blüher, M.; Baum, P.; Stockinger, M.; Toyka, K.V.; Klöting, I.; Stumvoll, M.; Serke, H.; Bechmann, I.; et al. Increased autophagy in peripheral nerves may protect Wistar Ottawa Karlsburg W rats against neuropathy. Exp. Neurol. 2013, 250, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Nowicki, M.; Baum, P.; Kosacka, J.; Stockinger, M.; Klöting, N.; Blüher, M.; Bechmann, I.; Toyka, K.V. Effects of isoflurane anesthesia on F-waves in the sciatic nerve of the adult rat. Muscle Nerve 2014, 50, 257–261. [Google Scholar] [CrossRef]
- Carenini, S.; Mäurer, M.; Werner, A.; Blazyca, H.; Toyka, K.V.; Schmid, C.D.; Raivich, G.; Martini, R. The Role of Macrophages in Demyelinating Peripheral Nervous System of Mice Heterozygously Deficient in p0. J. Cell Biol. 2001, 152, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Jann, S.; Bramerio, M.A.; Beretta, S.; Koch, S.; Defanti, C.A.; Toyka, K.V.; Sommer, C. Diagnostic Value of Sural Nerve Matrix metalloproteinase-9 in Diabetic Patients With CIDP. Neurology 2003, 61, 1607–1610. [Google Scholar] [CrossRef]
- Klöting, I.; Stark, O.; Brdicka, R. Incidence of the insulin-dependent Diabetes mellitus in BB rats: Their genetic heterogeneity and susceptibility to infection. Folia Biol (Praha) 1984, 30, 33–42. [Google Scholar] [PubMed]
- Thulé, P.M.; Campbell, A.G.; Jia, D.; Lin, Y.; You, S.; Paveglio, S.; Olson, D.E.; Kozlowski, M. Long-term glycemic control with hepatic insulin gene therapy in streptozotocin-diabetic mice. J. Gene Med. 2015, 17, 141–152. [Google Scholar]
- Hull, R.L.; Willard, J.R.; Struck, M.D.; Barrow, B.M.; Brar, G.S.; Andrikopoulos, S.; Zraika, S. High fat feeding unmasks variable insulin responses in male C57BL/6 mouse substrains. J. Endocrinol. 2017, 233, 53–64. [Google Scholar] [CrossRef]
- Tuck, R.R.; Schmelzer, J.D.; Low, P.A. Endoneurial blood flow and oxygen tension in the sciatic nerves of rats with experimental diabetic neuropathy. Brain 1984, 107, 935–950. [Google Scholar] [CrossRef]
- Chiarelli, F.; Verrotti, A.; Catino, M.; Sabatino, G.; Pinelli, L. Hypoglycaemia in Children with Type 1 Diabetes Mellitus. Acta Paediatr. 1999, Suppl. 88, 31–34. [Google Scholar]
- Nordfeldt, S.; Ludvigsson, J. Adverse Events in Intensively Treated Children and Adolescents with Type 1 Diabetes. Acta Paediatr. 1999, 88, 1184–1193. [Google Scholar] [CrossRef]
- Mohseni, S. Hypoglycemic Neuropathy. Acta Neuropathol. 2001, 102, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Mohseni, S.; Lillesaar, C.; Theodorsson, E.; Hildebrand, C. Hypoglycaemic Neuropathy: Occurrence of Axon Terminals in Plantar Skin and Plantar Muscle of Diabetic BB/Wor Rats Treated With Insulin Implants. Acta Neuropathol. 2000, 99, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Boulton, A.J.; Vinik, A.I.; Arezzo, J.C.; Bril, V.; Feldman, E.L.; Freeman, R.; Malik, R.A.; Maser, R.E.; Sosenko, J.M.; Ziegler, D. Diabetic neuropathies: A statement by the American Diabetes Association. Diabetes Care. 2005, 28, 956–962. [Google Scholar] [CrossRef]
- Barrett, A.M.; Lucero, M.A.; Le, T.; Robinson, R.L.; Dworkin, R.H.; Chappell, A.S. Epidemiology, public health burden, and treatment of diabetic peripheral neuropathic pain: A review. Pain Med. 2007, 8, S50–S62. [Google Scholar] [CrossRef] [PubMed]
- Hatch, M.N.; Cushing, T.R.; Carlson, G.D.; Chang, E.Y. Neuropathic pain and SCI: Identification and treatment strategies in the 21st century. J. Neurol. Sci. 2018, 384, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Scholz, J.; Woolf, C.J. The Neuropathic Pain Triad: Neurons, Immune Cells and Glia. Nat. Neurosci. 2007, 10, 1361–1368. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Kiguchi, N.; Fukazawa, Y.; Saika, F.; Maeda, T.; Kishioka, S. Macrophage-T cell interactions mediate neuropathic pain through the glucocorticoid-induced tumor necrosis factor ligand system. J. Biol. Chem. 2015, 290, 12603–12613. [Google Scholar] [CrossRef]
- Saika, F.; Kiguchi, N.; Matsuzaki, S.; Kobayashi, D.; Kishioka, S. Inflammatory Macrophages in the Sciatic Nerves Facilitate Neuropathic Pain Associated with Type 2 Diabetes Mellitus. J. Pharmacol. Exp. Ther. 2019, 368, 535–544. [Google Scholar] [CrossRef]
- Sommer, C.; Kress, M. Recent Findings on How Proinflammatory Cytokines Cause Pain: Peripheral Mechanisms in Inflammatory and Neuropathic Hyperalgesia. Neurosci. Lett. 2004, 361, 184–187. [Google Scholar] [CrossRef]
- Saika, F.; Kiguchi, N.; Kobayashi, Y.; Fukazawa, Y.; Kishioka, S. CC-chemokine ligand 4/macrophage inflammatory protein-1β participates in the induction of neuropathic pain after peripheral nerve injury. Eur. J. Pain. 2012, 16, 1271–1280. [Google Scholar] [CrossRef]
- Zhang, D.; Mou, J.Y.; Wang, F.; Liu, J.; Hu, X. CRNDE enhances neuropathic pain via modulating miR-136/IL6R axis in CCI rat models. J. Cell. Physiol. 2019, 234, 22234–22241. [Google Scholar] [CrossRef] [PubMed]
- Davies, A.J.; Rinaldi, S.; Costigan, M.; Bae Oh, S. Cytotoxic Immunity in Peripheral Nerve Injury and Pain. Front. Neurosci. 2020, 14, 142. [Google Scholar] [CrossRef] [PubMed]
- Echeverry, S.; Wu, Y.; Zhang, J. Selectively reducing cytokine/chemokine expressing macrophages in injured nerves impairs the development of neuropathic pain. Exp. Neurol. 2013, 240, 205–218. [Google Scholar] [CrossRef]
- Kiguchi, N.; Kobayashi, D.; Saika, F.; Matsuzaki, S.; Kishioka, S. Inhibition of peripheral macrophages by nicotinic acetylcholine receptor agonists suppresses spinal microglial activation and neuropathic pain in mice with peripheral nerve injury. J. Neuroinflammation. 2018, 15, 96. [Google Scholar] [CrossRef]
- Gibbons, C.H. Treatment-Induced Neuropathy of Diabetes. Curr. Diab. Rep. 2017, 17, 127. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Pascual, C.; Lieu, C.; Oh, S.; Wang, J. Analgesic Microneedle Patch for Neuropathic Pain Therapy. ACS Nano. 2017, 11, 395–406. [Google Scholar] [CrossRef]
- Dyck, P.J.; Lambert, E.H.; O’Brien, P.C. Pain in peripheral neuropathy related to rate and kind of fiber degeneration. Neurology 1976, 26, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Sommer, C.; Leinders, M.; Üçeyler, N. Inflammation in the Pathophysiology of Neuropathic Pain. Pain 2018, 159, 595–602. [Google Scholar] [CrossRef]
- Gerstein, H.C.; Miller, M.E.; Byington, R.P.; Goff, D.C., Jr.; Bigger, J.T.; Buse, J.B.; Cushman, W.C.; Genuth, S.; Ismail-Beigi, F.; Grimm, R.H., Jr.; et al. Action to Control Cardiovascular Risk in Diabetes Study Group. Effects of Intensive Glucose Lowering in Type 2 Diabetes. N. Engl. J. Med. 2008, 358, 2545–2559. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blood Glucose (mmol/L) ±SD | 0 Month | After 1st Month | After 3rd Month |
|---|---|---|---|
| group 1 | 20.8 ± 2.5 | 3.9 ± 1.0 | 4.1 ± 0.7 |
| group 2 | 21.4 ± 2.4 | 18.8 ± 1.7 | 20.2 ± 2.4 |
| group 3 | 20.3 ± 2.1 | 19.1 ± 1.9 | 4.3 ± 1.2 |
| HbA1C (%) ±SD | After 1st Month | After 2nd Month | After 3rd Month |
|---|---|---|---|
| group 1 | 4.9 ± 0.9 | 4.8 ± 1.5 | 5.1 ± 0.8 |
| group 2 | 7.0 ± 1.1 | 7.4 ± 0.8 | 7.4 ± 0.6 |
| group 3 | 6.8 ± 1.2 | 5.7 ± 1.3 | 5.1 ± 1.0 |
| Start | After 1st Month | After 2nd Month | After 3rd Month | ||
|---|---|---|---|---|---|
| group 1 | mNCV | 100 | 94.2 | 102.2 | 97.6 |
| csNCV | 100 | 98.6 | 101.2 | 103 | |
| F wave | 100 | 95 | 93 | 92 | |
| group 2 | mNCV | 100 | 90.7 | 90.4 | 93.3 |
| csNCV | 100 | 99.8 | 98.8 | 88.7 | |
| F wave | 100 | 101 | 94.3 | 99 | |
| group 3 | mNCV | 100 | 91.3 | 93 | 95.4 |
| csNCV | 100 | 92 | 105.4 | 102.6 | |
| F wave | 100 | 98.5 | 95 | 91.4 | |
| Start | After 1st Month | After 2nd Month | After 3rd Month | ||
|---|---|---|---|---|---|
| decrease in HbA1c >2% | mNCV | 100 | 88.3 | 88.5 | 81.3 |
| csNCV | 100 | 91.7 | 100.3 | 90 | |
| F wave | 100 | 95.6 | 92 | 88 | |
| decrease in HbA1c <2% | mNCV | 100 | 93.5 | 96 | 106.6 |
| csNCV | 100 | 92.2 | 108 | 112.3 | |
| F wave | 100 | 100.5 | 97.2 | 94 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baum, P.; Koj, S.; Klöting, N.; Blüher, M.; Classen, J.; Paeschke, S.; Gericke, M.; Toyka, K.V.; Nowicki, M.; Kosacka, J. Treatment-Induced Neuropathy in Diabetes (TIND)—Developing a Disease Model in Type 1 Diabetic Rats. Int. J. Mol. Sci. 2021, 22, 1571. https://doi.org/10.3390/ijms22041571
Baum P, Koj S, Klöting N, Blüher M, Classen J, Paeschke S, Gericke M, Toyka KV, Nowicki M, Kosacka J. Treatment-Induced Neuropathy in Diabetes (TIND)—Developing a Disease Model in Type 1 Diabetic Rats. International Journal of Molecular Sciences. 2021; 22(4):1571. https://doi.org/10.3390/ijms22041571
Chicago/Turabian StyleBaum, Petra, Severin Koj, Nora Klöting, Matthias Blüher, Joseph Classen, Sabine Paeschke, Martin Gericke, Klaus V. Toyka, Marcin Nowicki, and Joanna Kosacka. 2021. "Treatment-Induced Neuropathy in Diabetes (TIND)—Developing a Disease Model in Type 1 Diabetic Rats" International Journal of Molecular Sciences 22, no. 4: 1571. https://doi.org/10.3390/ijms22041571
APA StyleBaum, P., Koj, S., Klöting, N., Blüher, M., Classen, J., Paeschke, S., Gericke, M., Toyka, K. V., Nowicki, M., & Kosacka, J. (2021). Treatment-Induced Neuropathy in Diabetes (TIND)—Developing a Disease Model in Type 1 Diabetic Rats. International Journal of Molecular Sciences, 22(4), 1571. https://doi.org/10.3390/ijms22041571