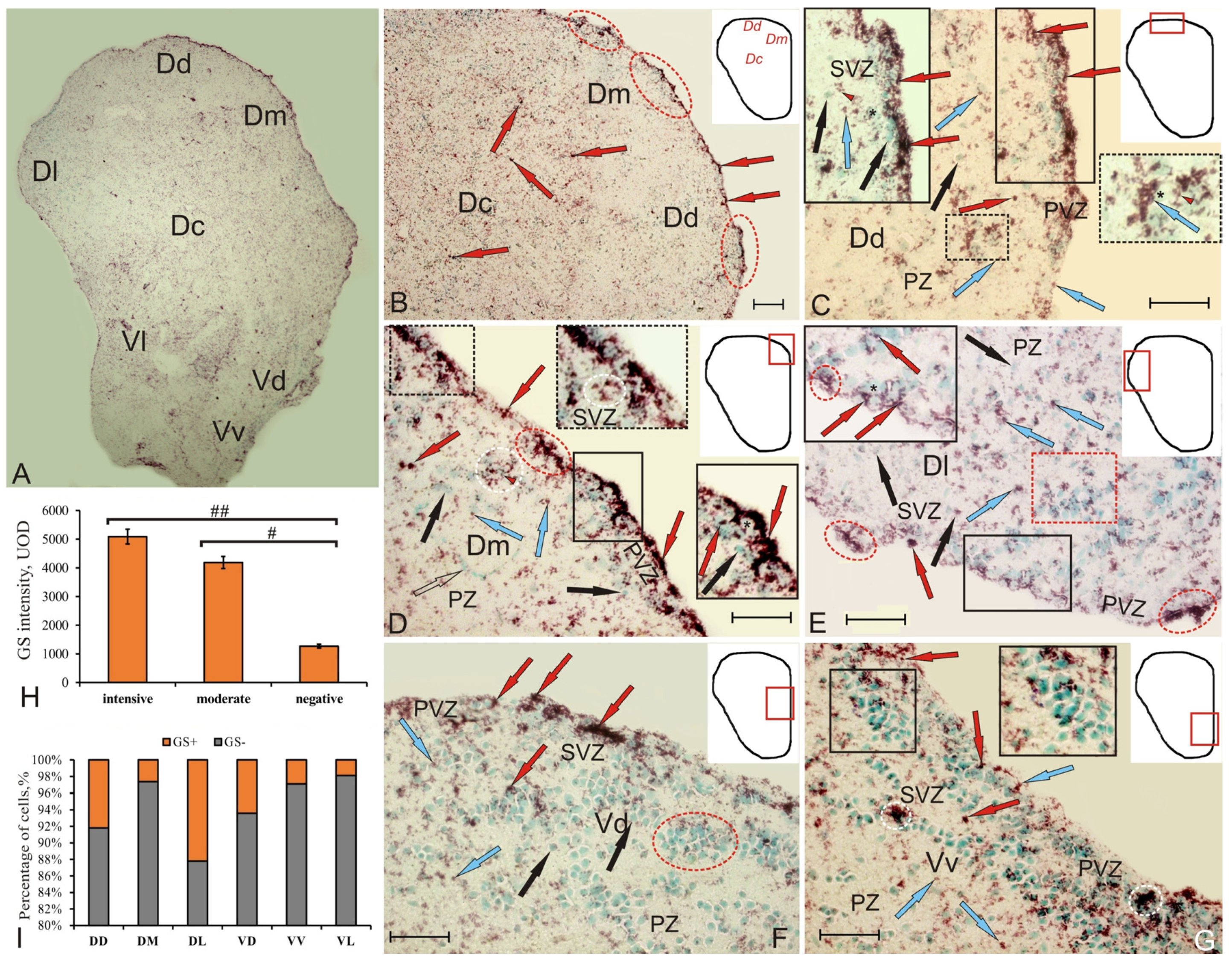
Figure 1.
Glutamine synthetase (GS) in the pallial and subpallial regions of intact telencephalon in juvenile chum salmon, Oncorhynchus keta. (A) General view of the GS immunolocalization pattern in the telencephalon of intact juvenile chum salmon, Dd—dorsal, Dm—medial, Dc—central zones of pallium, Vd—dorsal, Vv—ventral, Vl—lateral zones of subpallium; (B) in the pallial area, the pictogram shows the zones of the dorsal telencephalon (pallium), ovals outline periventricular aggregations of GS+ cells, intensely labeled cells (red arrows); (C) dorsal pallial zone (Dd) at higher magnification, inset (in red rectangle) shows a fragment including periventricular zone (PVZ) and subventricular zone (SVZ); inset (in black rectangle) shows intensely labeled GS+ neuroepithelial cells (red arrows) located in the PVZ above the layer of immunonegative cells (black asterisk), in the SVZ there were (dotted inset) clusters of small moderately labeled cells (red triangular arrow), a moderately GS+ cell (blue arrow), GS–cell (black arrow), PZ—parenchymal zone; (D) the medial pallial zone (Dm) contains small GS+ cells (outlined with a white dotted line) in SVZ (black dotted inset), dense groups of small intensively labeled cells in PVZ of irregular shape (inset in a black rectangle), endothelial cells are shown with a transparent arrow; (E) lateral pallial zone (Dl) including small moderately labeled GS cells (in the red oval) and GS+ cytoplasm of neuroepithelial cells surrounding the GS nuclei in PVZ (inset in the black rectangle), the neurogenic niche of parenchymal localization is in the red rectangle; (F) the dorsal subpallial zone (Vd) contains intensely labeled groups of neuroepithelial cells (red arrows) and parenchymal neurogenic niches (in red oval); (G) the ventral subpallial zone (Vv) contains single (red arrows) or paired (blue arrows) moderately labeled cells in the apical part of PVZ, aggregations of intensely labeled granule-like subcellular elements (in white ovals), pseudo-unipolar GS+ cells occurred in SVZ and penetrate into the deeper layers of PZ (inset). Immunohistochemical labeling of glutamine synthetase. Scale bar: 100 µm. (H) Comparative densitometric analysis of labeling of GS+ cells; significant intergroup differences # (p < 0.05), ## (p < 0.01) between the groups of intensely, moderately labeled cells, and immunonegative cells (n = 5 in each group). One-way ANOVA. Intensity of GS labeling, units of optical density (UOD). (I) Percentage ratio between GS+ and GS–cells in all intact pallial and subpallial areas of juvenile chum salmon, Oncorhynchus keta.
Figure 1.
Glutamine synthetase (GS) in the pallial and subpallial regions of intact telencephalon in juvenile chum salmon, Oncorhynchus keta. (A) General view of the GS immunolocalization pattern in the telencephalon of intact juvenile chum salmon, Dd—dorsal, Dm—medial, Dc—central zones of pallium, Vd—dorsal, Vv—ventral, Vl—lateral zones of subpallium; (B) in the pallial area, the pictogram shows the zones of the dorsal telencephalon (pallium), ovals outline periventricular aggregations of GS+ cells, intensely labeled cells (red arrows); (C) dorsal pallial zone (Dd) at higher magnification, inset (in red rectangle) shows a fragment including periventricular zone (PVZ) and subventricular zone (SVZ); inset (in black rectangle) shows intensely labeled GS+ neuroepithelial cells (red arrows) located in the PVZ above the layer of immunonegative cells (black asterisk), in the SVZ there were (dotted inset) clusters of small moderately labeled cells (red triangular arrow), a moderately GS+ cell (blue arrow), GS–cell (black arrow), PZ—parenchymal zone; (D) the medial pallial zone (Dm) contains small GS+ cells (outlined with a white dotted line) in SVZ (black dotted inset), dense groups of small intensively labeled cells in PVZ of irregular shape (inset in a black rectangle), endothelial cells are shown with a transparent arrow; (E) lateral pallial zone (Dl) including small moderately labeled GS cells (in the red oval) and GS+ cytoplasm of neuroepithelial cells surrounding the GS nuclei in PVZ (inset in the black rectangle), the neurogenic niche of parenchymal localization is in the red rectangle; (F) the dorsal subpallial zone (Vd) contains intensely labeled groups of neuroepithelial cells (red arrows) and parenchymal neurogenic niches (in red oval); (G) the ventral subpallial zone (Vv) contains single (red arrows) or paired (blue arrows) moderately labeled cells in the apical part of PVZ, aggregations of intensely labeled granule-like subcellular elements (in white ovals), pseudo-unipolar GS+ cells occurred in SVZ and penetrate into the deeper layers of PZ (inset). Immunohistochemical labeling of glutamine synthetase. Scale bar: 100 µm. (H) Comparative densitometric analysis of labeling of GS+ cells; significant intergroup differences # (p < 0.05), ## (p < 0.01) between the groups of intensely, moderately labeled cells, and immunonegative cells (n = 5 in each group). One-way ANOVA. Intensity of GS labeling, units of optical density (UOD). (I) Percentage ratio between GS+ and GS–cells in all intact pallial and subpallial areas of juvenile chum salmon, Oncorhynchus keta.

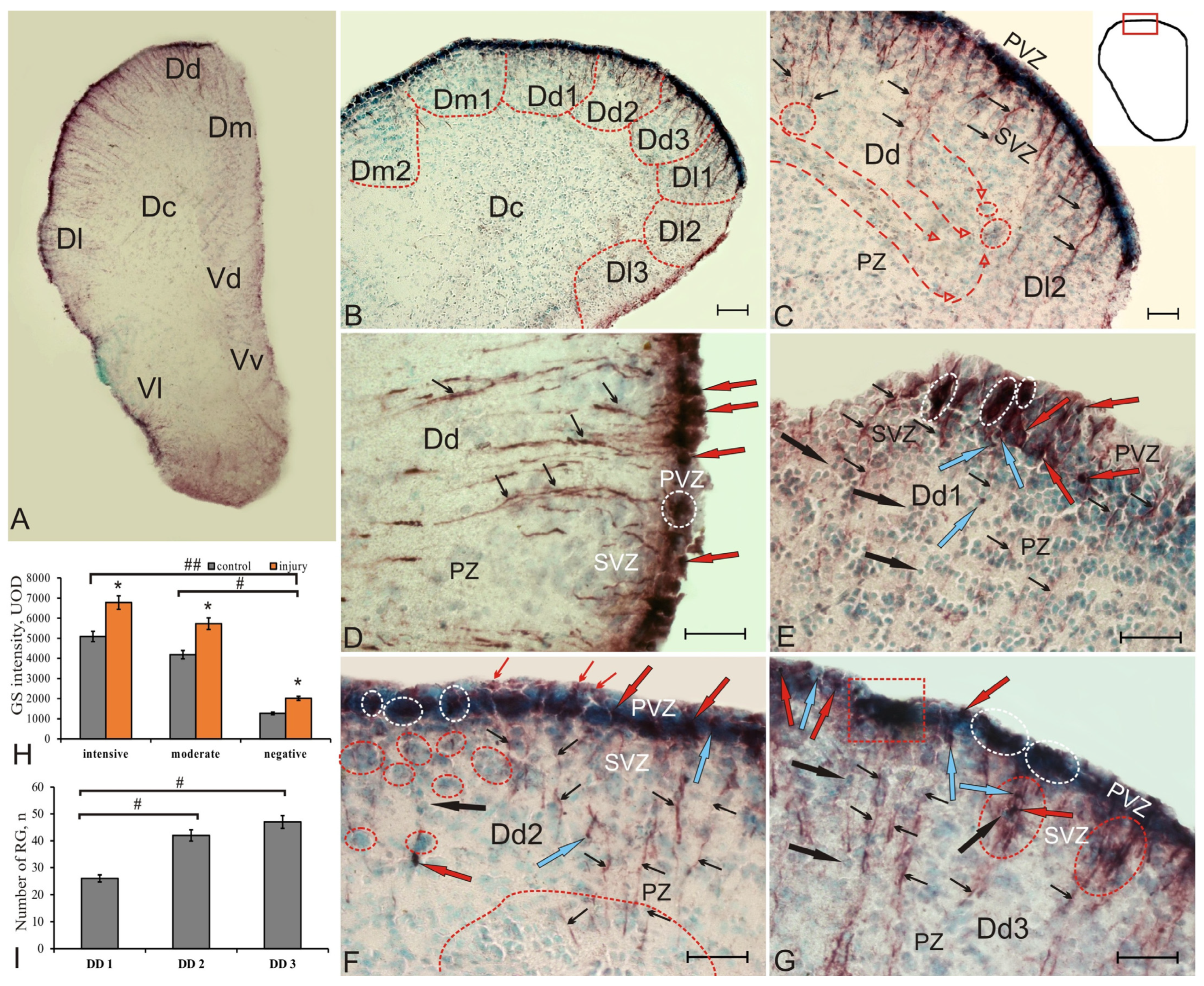
Figure 2.
Glutamine synthetase in the pallial region of the telencephalon of juvenile chum salmon, Oncorhynchus keta, at 3 days after traumatic injury. (A) General view of the GS immunolocalization pattern in the damaged telencephalon of juvenile chum salmon, (B) pallial zone, the red dotted line indicates the boundaries of the subzones; (C) patterns of cell migration (directions are indicated by red dashed arrows) and distribution of radial glia (black arrows) in DD, reactive neurogenic niches of parenchymal localization are outlined by red ovals; (D) patterns of distribution of radial glia (RG) and reactive neurogenic niches (in white oval) in DD; (E) cellular composition of reactive neurogenic niches in DD1, small reactive intensely labeled GS clusters of cells (in white ovals), GS–cells are indicated by black arrows; (F) dense clusters of GS + RG in DD2, thin red arrows indicate the end-feets of the immunopositive RG in PVZ, the red dotted line outlines the reactive aggregations of neuroblasts in SVZ and the largest in PVZ; (G) patterns of RG distribution in DD3, single radial fibers (thin black arrows), along which immunonegative neuroblasts migrate (black arrows), combined with bundles of radial fibers (red ovals); large reactive neurogenic niches (RNN) (in a red rectangle). Immunohistochemical labeling of glutamine synthetase. Scale bars: (B) 100 µm, (C–G) 50 µm. (H) Comparative densitometric analysis of GS+ cell activity in intact animals and on day 3 post-injury; significant intergroup differences # (p < 0.05), ## (p < 0.01) between the groups of intensely, moderately labeled cells, and immunonegative cells (n = 5 in each group). Data are expressed as the mean ± SD (n = 5 in each group); one-way ANOVA followed by the Student–Newman–Keuls post hoc test was used to determine significant difference between control animals and those on day 3 post-injury, p < 0.05 vs. control group. Intensity of GS labeling, UOD; * p ≤ 0.05. (I) Comparative distribution of RG in DD1, DD2 and DD3, on day 3 post-injury. One-way ANOVA (n = 5 in each group, #—significant intergroup differences (p < 0.05).
Figure 2.
Glutamine synthetase in the pallial region of the telencephalon of juvenile chum salmon, Oncorhynchus keta, at 3 days after traumatic injury. (A) General view of the GS immunolocalization pattern in the damaged telencephalon of juvenile chum salmon, (B) pallial zone, the red dotted line indicates the boundaries of the subzones; (C) patterns of cell migration (directions are indicated by red dashed arrows) and distribution of radial glia (black arrows) in DD, reactive neurogenic niches of parenchymal localization are outlined by red ovals; (D) patterns of distribution of radial glia (RG) and reactive neurogenic niches (in white oval) in DD; (E) cellular composition of reactive neurogenic niches in DD1, small reactive intensely labeled GS clusters of cells (in white ovals), GS–cells are indicated by black arrows; (F) dense clusters of GS + RG in DD2, thin red arrows indicate the end-feets of the immunopositive RG in PVZ, the red dotted line outlines the reactive aggregations of neuroblasts in SVZ and the largest in PVZ; (G) patterns of RG distribution in DD3, single radial fibers (thin black arrows), along which immunonegative neuroblasts migrate (black arrows), combined with bundles of radial fibers (red ovals); large reactive neurogenic niches (RNN) (in a red rectangle). Immunohistochemical labeling of glutamine synthetase. Scale bars: (B) 100 µm, (C–G) 50 µm. (H) Comparative densitometric analysis of GS+ cell activity in intact animals and on day 3 post-injury; significant intergroup differences # (p < 0.05), ## (p < 0.01) between the groups of intensely, moderately labeled cells, and immunonegative cells (n = 5 in each group). Data are expressed as the mean ± SD (n = 5 in each group); one-way ANOVA followed by the Student–Newman–Keuls post hoc test was used to determine significant difference between control animals and those on day 3 post-injury, p < 0.05 vs. control group. Intensity of GS labeling, UOD; * p ≤ 0.05. (I) Comparative distribution of RG in DD1, DD2 and DD3, on day 3 post-injury. One-way ANOVA (n = 5 in each group, #—significant intergroup differences (p < 0.05).

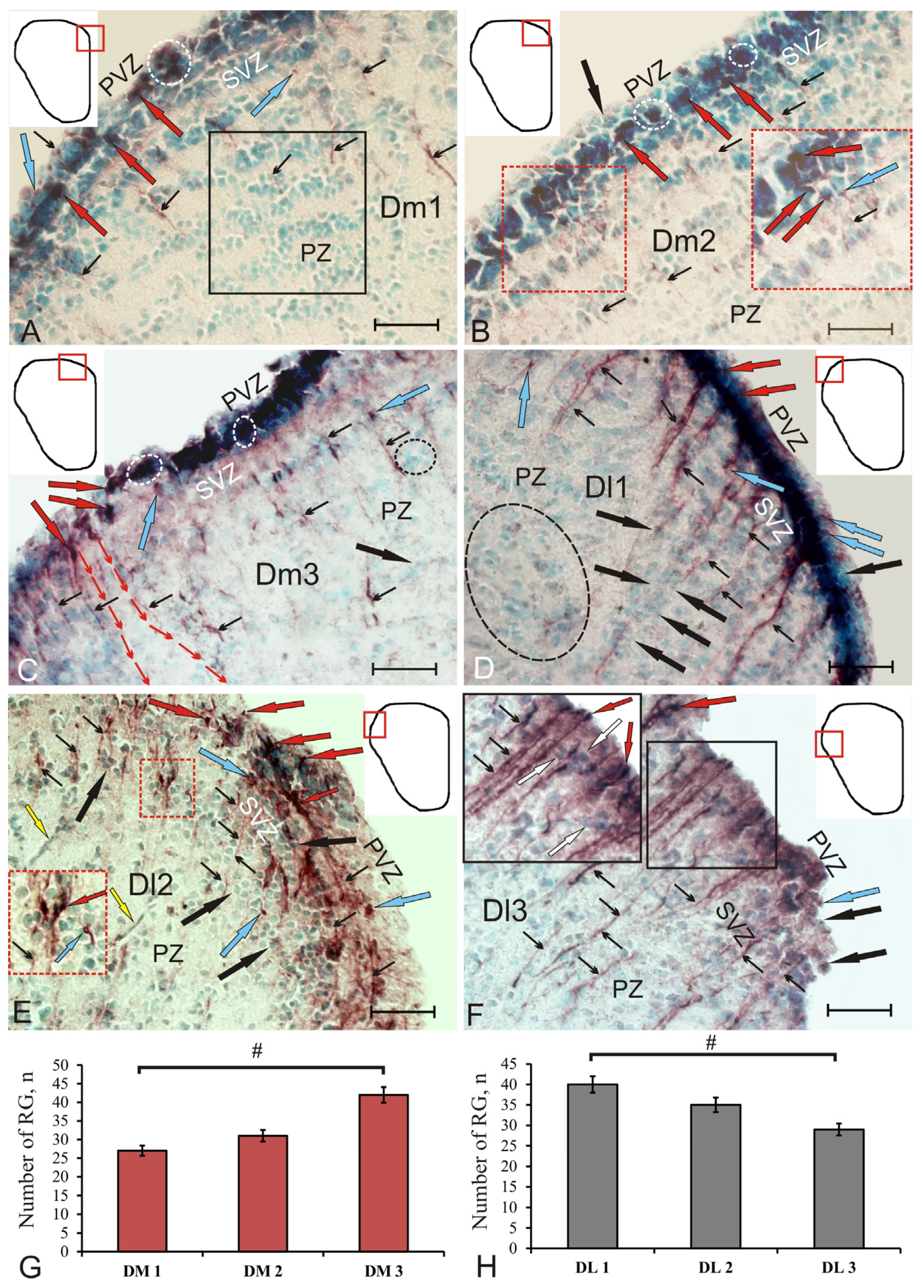
Figure 3.
Glutamine synthetase in the medial (A–C) and lateral (D–F) pallial areas of the telencephalon of juvenile chum salmon, Oncorhynchus keta, at 3 days after the traumatic injury to the telencephalon. (A) Diffuse pattern of GS+ radial glia distribution in DM1, densely or moderately labeled GS+ neuroepithelial cells (blue arrows), a fragment of the pseudo-stratified structure of PZ with migrating neuroblasts and RG fibers is in the black rectangle; (B) reactive neurogenic niches containing GS+ neuroepithelial cells (dotted inset) in DM2; (C) patterns of neuroblasts’ migration in DM3 (the direction of migration is indicated by red thin arrows), RNN of the neuroepithelial type in PVZ (in white ovals), RNN of parenchymal localization (in the black oval); (D) the largest and longest RG fibers and bundles in DL1, with GS–neuroblasts migrating along RG (black arrows), aggregation of GS neuroblasts (in the black oval); (E) patterns of tangentially oriented GS+ fibers in DL2; along the RG fibers, extensive migration of a heterogeneous population of cells was observed (inset), yellow arrows indicate endothelial cells; (F) dense population of GS + RG in DL3, long outgrowths deeply spreading in PZ (inset). Immunohistochemical labeling of glutamine synthetase. Scale bar: 50 µm. (G) Comparative distribution of RG in DM1, DM2 and DM3, and in DL1, DL2 and DL3 on (H) day 3 post-injury. One-way ANOVA (n = 5 in each group, #—significant intergroup differences (p < 0.05).
Figure 3.
Glutamine synthetase in the medial (A–C) and lateral (D–F) pallial areas of the telencephalon of juvenile chum salmon, Oncorhynchus keta, at 3 days after the traumatic injury to the telencephalon. (A) Diffuse pattern of GS+ radial glia distribution in DM1, densely or moderately labeled GS+ neuroepithelial cells (blue arrows), a fragment of the pseudo-stratified structure of PZ with migrating neuroblasts and RG fibers is in the black rectangle; (B) reactive neurogenic niches containing GS+ neuroepithelial cells (dotted inset) in DM2; (C) patterns of neuroblasts’ migration in DM3 (the direction of migration is indicated by red thin arrows), RNN of the neuroepithelial type in PVZ (in white ovals), RNN of parenchymal localization (in the black oval); (D) the largest and longest RG fibers and bundles in DL1, with GS–neuroblasts migrating along RG (black arrows), aggregation of GS neuroblasts (in the black oval); (E) patterns of tangentially oriented GS+ fibers in DL2; along the RG fibers, extensive migration of a heterogeneous population of cells was observed (inset), yellow arrows indicate endothelial cells; (F) dense population of GS + RG in DL3, long outgrowths deeply spreading in PZ (inset). Immunohistochemical labeling of glutamine synthetase. Scale bar: 50 µm. (G) Comparative distribution of RG in DM1, DM2 and DM3, and in DL1, DL2 and DL3 on (H) day 3 post-injury. One-way ANOVA (n = 5 in each group, #—significant intergroup differences (p < 0.05).

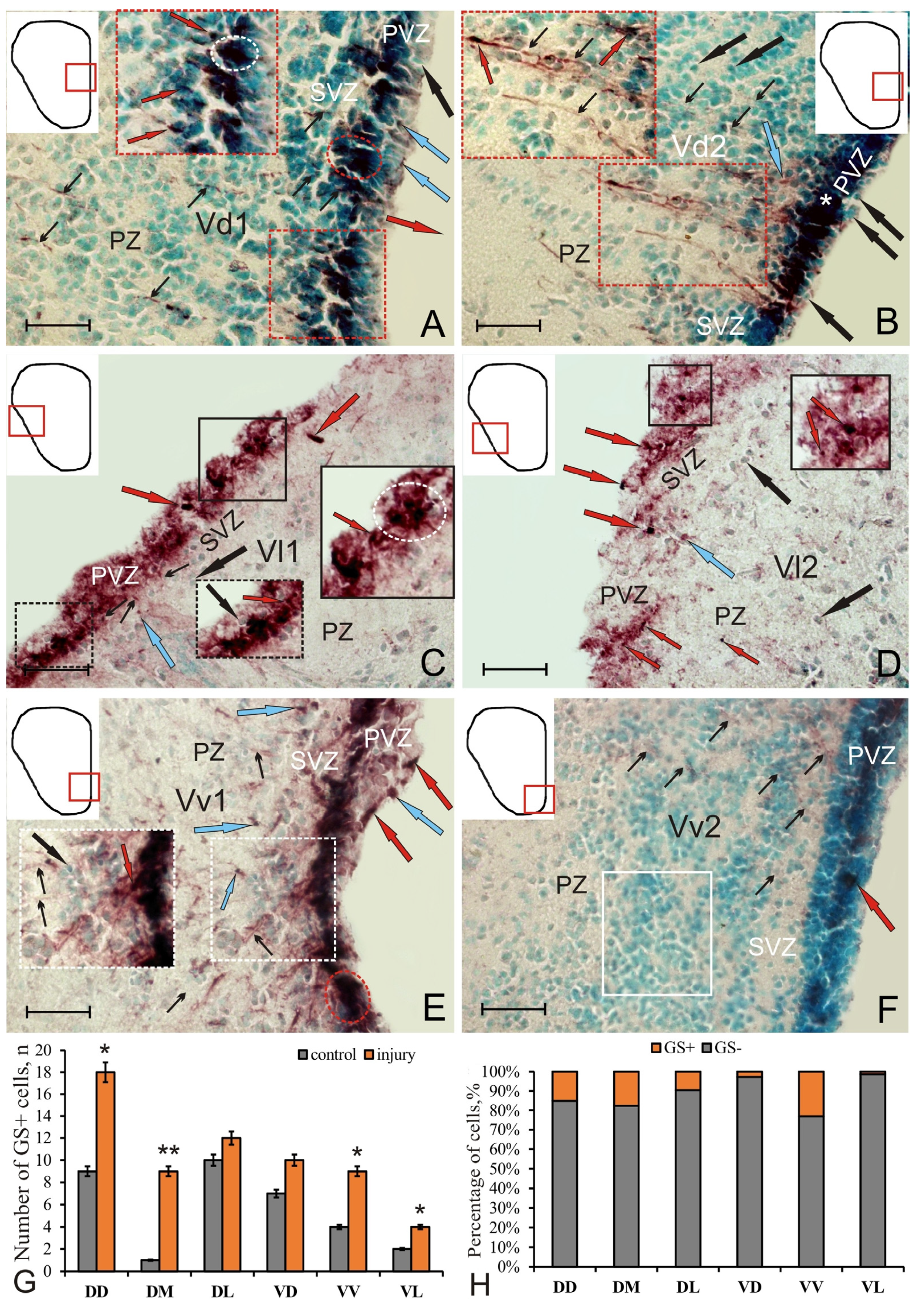
Figure 4.
Glutamine synthetase in the subpallial areas of juvenile chum salmon, Oncorhynchus keta, at 3 days after the traumatic injury to the telencephalon. (A) In VD1, numerous neuroepithelial cell clusters were detected in PVZ and SVZ (inset), producing the RNN (in the red oval), and the number of RG fibers (black thin arrows) was reduced; (B) migrating GS+ cells were found in VD2 along GS+ RG fibers (red arrows, inset); in the PVZ, elongated GS+ cells (white asterisk) were revealed, above which a layer of GS–neuroepithelial cells was found (black arrows); (C) VL1 was dominated by aggregations of GS+ neuroepithelial cells (in the white oval) in the dorsal part of PVZ (black inset) and in the ventral part (dotted inset), and the number of RG (thin arrows) was limited; (D) single GS+ neuroepithelial cells (red arrow) were detected in VL2 in PVZ (inset), and RG fibers were not revealed; (E) in VV1, mixed RNNs were found (inset) containing large densely stained GS+ groups of neuroepithelial cells (red arrows), RG fibers (thin black arrows), and individual moderately labeled cells (blue arrow); (F) in VV2, a few immunolabeled RG fibers, single GS+ accumulations of neuroepithelial cells in PVZ (red arrow) and a high concentration of GS–cells in PVZ (in the white square) were revealed; (G) quantitative ratio of GS+ cells in intact animals (control group) and at 3 days after traumatic injury to the telencephalon (n = 5 in each group, * p ≤ 0.05; ** p ≤ 0.01; significant difference from control groups). One-way ANOVA followed by the Student–Newman–Keuls post hoc test. (H) Percentage proportion between GS+/GS–cells in all areas of the pallium and subpallium of juvenile chum salmon on day 3 posy-injury.
Figure 4.
Glutamine synthetase in the subpallial areas of juvenile chum salmon, Oncorhynchus keta, at 3 days after the traumatic injury to the telencephalon. (A) In VD1, numerous neuroepithelial cell clusters were detected in PVZ and SVZ (inset), producing the RNN (in the red oval), and the number of RG fibers (black thin arrows) was reduced; (B) migrating GS+ cells were found in VD2 along GS+ RG fibers (red arrows, inset); in the PVZ, elongated GS+ cells (white asterisk) were revealed, above which a layer of GS–neuroepithelial cells was found (black arrows); (C) VL1 was dominated by aggregations of GS+ neuroepithelial cells (in the white oval) in the dorsal part of PVZ (black inset) and in the ventral part (dotted inset), and the number of RG (thin arrows) was limited; (D) single GS+ neuroepithelial cells (red arrow) were detected in VL2 in PVZ (inset), and RG fibers were not revealed; (E) in VV1, mixed RNNs were found (inset) containing large densely stained GS+ groups of neuroepithelial cells (red arrows), RG fibers (thin black arrows), and individual moderately labeled cells (blue arrow); (F) in VV2, a few immunolabeled RG fibers, single GS+ accumulations of neuroepithelial cells in PVZ (red arrow) and a high concentration of GS–cells in PVZ (in the white square) were revealed; (G) quantitative ratio of GS+ cells in intact animals (control group) and at 3 days after traumatic injury to the telencephalon (n = 5 in each group, * p ≤ 0.05; ** p ≤ 0.01; significant difference from control groups). One-way ANOVA followed by the Student–Newman–Keuls post hoc test. (H) Percentage proportion between GS+/GS–cells in all areas of the pallium and subpallium of juvenile chum salmon on day 3 posy-injury.

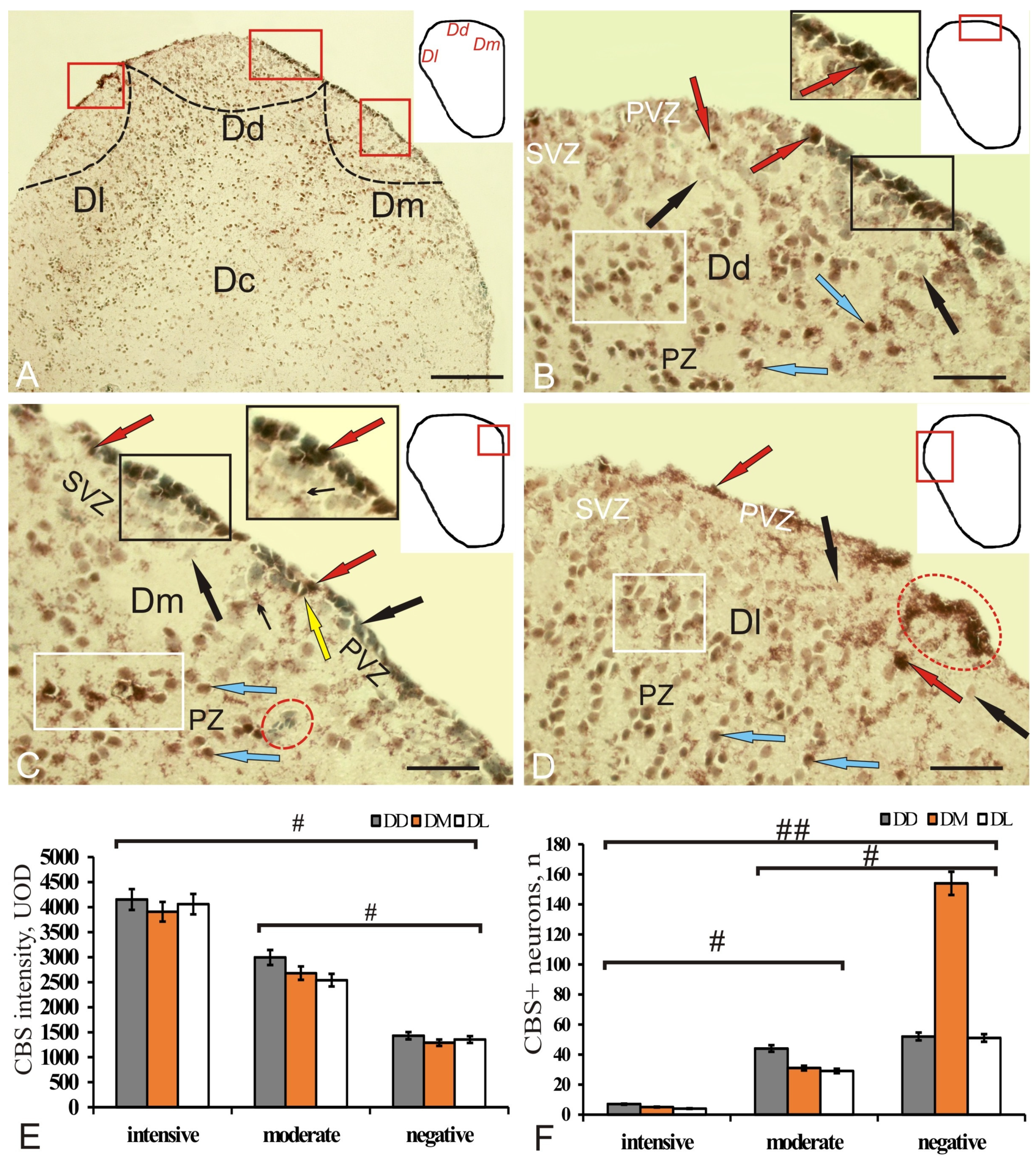
Figure 5.
Cystathionine β-synthase (CBS) in the pallium of intact juvenile chum salmon, Oncorhynchus keta. (A) General view of the pallial region of the telencephalon, the pictogram shows the zones of the dorsal telencephalon (pallium), Dd—dorsal, Dm—medial, Dl—lateral, (outlined with red rectangles) black dotted lines indicate the boundaries of the pallial zones; (B) dorsal pallial zone (Dd) at higher magnification, inset (in black rectangle) shows a fragment including PVZ and SVZ, intensely labeled CBS+ neuroepithelial cells (red arrows), moderately labeled CBS cells (blue arrow), and negative cells (black arrow) in all areas; (C) medial pallial zone (Dm), some CBS+ cells showed proximal areas of radially directed processes (yellow arrow), clusters of small moderately CBS–labeled cells were detected in the SVZ (inset), and small clusters of microglia were revealed in PZ (in the red oval) and groups of intensely and moderately labeled cells (in a white rectangle); (D) lateral pallial zone (Dl) with a diffuse pattern of CBS-immunolocalization, aggregation of CBS+ cells in PVZ (in the red oval). Immunohistochemical labeling of cystathionine β-synthase. Scale bars: (A) 100 µm, (B–D) 50 µm. (E) Comparative densitometric analysis of CBS+ cells’ activity; #—significant intergroup differences (p < 0.05), in groups of intensely, moderately labeled cells and immunonegative cells (n = 5 in each group). One-way ANOVA. Intensity of CBS labeling, UOD. (F) Comparative distribution of CBS+ cells in DD, DM and DL; significant intergroup differences # (p < 0.05), ## (p < 0.01) between the groups of intensively, moderately labeled cells, and immunonegative cells (n = 5 in each group). One-way ANOVA.
Figure 5.
Cystathionine β-synthase (CBS) in the pallium of intact juvenile chum salmon, Oncorhynchus keta. (A) General view of the pallial region of the telencephalon, the pictogram shows the zones of the dorsal telencephalon (pallium), Dd—dorsal, Dm—medial, Dl—lateral, (outlined with red rectangles) black dotted lines indicate the boundaries of the pallial zones; (B) dorsal pallial zone (Dd) at higher magnification, inset (in black rectangle) shows a fragment including PVZ and SVZ, intensely labeled CBS+ neuroepithelial cells (red arrows), moderately labeled CBS cells (blue arrow), and negative cells (black arrow) in all areas; (C) medial pallial zone (Dm), some CBS+ cells showed proximal areas of radially directed processes (yellow arrow), clusters of small moderately CBS–labeled cells were detected in the SVZ (inset), and small clusters of microglia were revealed in PZ (in the red oval) and groups of intensely and moderately labeled cells (in a white rectangle); (D) lateral pallial zone (Dl) with a diffuse pattern of CBS-immunolocalization, aggregation of CBS+ cells in PVZ (in the red oval). Immunohistochemical labeling of cystathionine β-synthase. Scale bars: (A) 100 µm, (B–D) 50 µm. (E) Comparative densitometric analysis of CBS+ cells’ activity; #—significant intergroup differences (p < 0.05), in groups of intensely, moderately labeled cells and immunonegative cells (n = 5 in each group). One-way ANOVA. Intensity of CBS labeling, UOD. (F) Comparative distribution of CBS+ cells in DD, DM and DL; significant intergroup differences # (p < 0.05), ## (p < 0.01) between the groups of intensively, moderately labeled cells, and immunonegative cells (n = 5 in each group). One-way ANOVA.

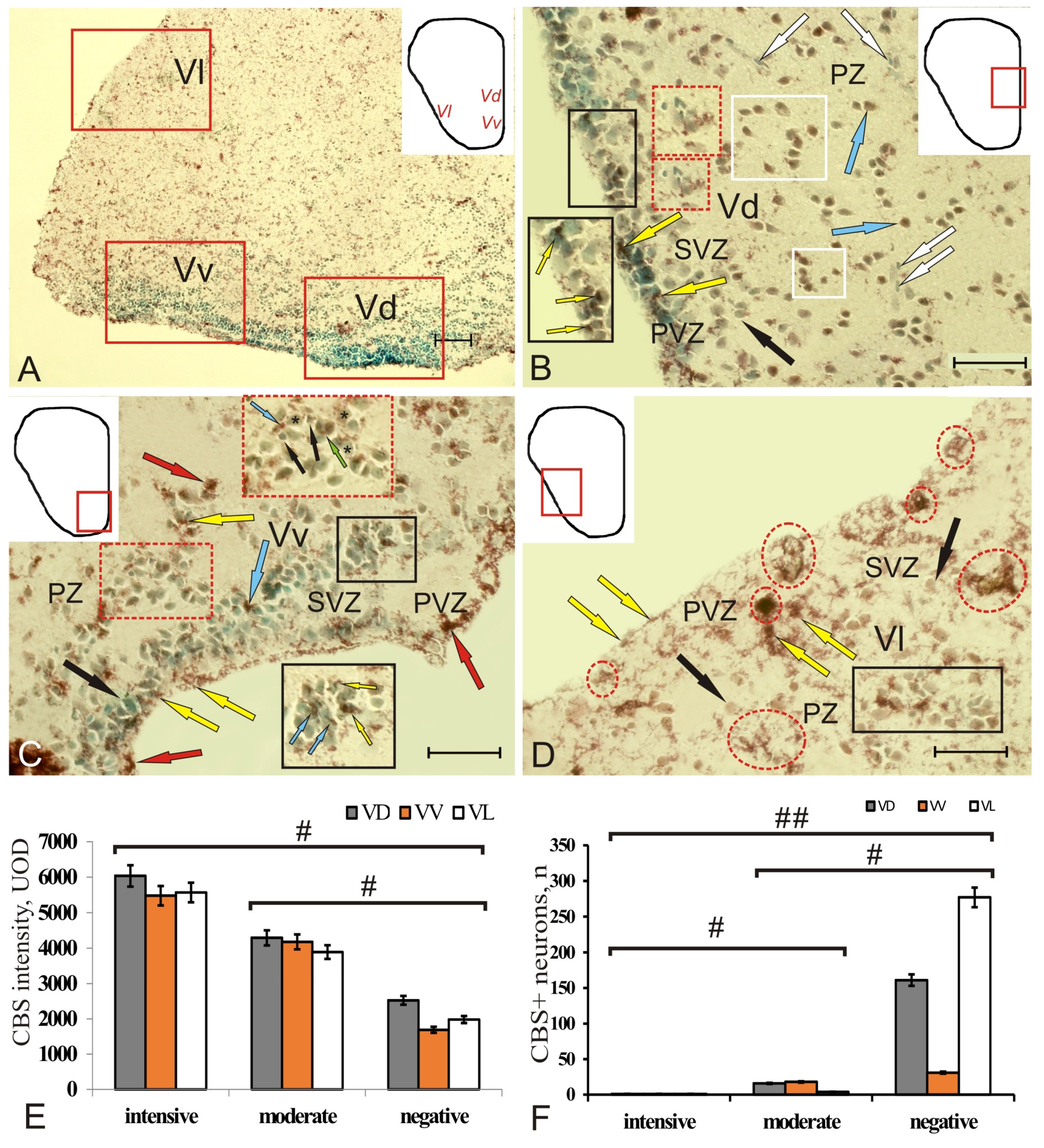
Figure 6.
Cystathionine β-synthase (CBS) in the subpallium of intact juvenile chum salmon, Oncorhynchus keta. (A) General view of the subpallial region of the telencephalon, the pictogram shows the zones of the ventral telencephalon (subpallium), Vd—dorsal, Vv—ventral, Vl—lateral, (outlined with red rectangles) areas; (B) dorsal subpallial zone (Vd) at higher magnification, the inset (in the black rectangle) shows CBS+ granules in PVZ and SVZ (yellow arrows), a fragment (red inset) including CBS–cells surrounded by an aggregation of small elongated basophilic cells (white arrows), clusters of CBS+ cells in PZ (in a white rectangle); (C) ventral subpallial region (Vv), CBS+ granules (yellow arrows) on the apical surface of PVZ neuroepithelial cells and clusters in SVZ (black inset), elongated moderately labeled CBS cells and granules inside and on the surface of CBS cells (red inset), nuclei stained with methyl green are indicated by an asterisk, a green arrow indicates the figure of mitosis; (D) the lateral subpallial region (Vl) of the accumulation of small, intensely labeled cells (in the red oval); multiple CBS+ granules (in the black rectangle) were located in PZ on the surface of CBS cells. Immunohistochemical labeling of cystathionine β-synthase. Scale bars: (A) 100 µm, (B–D) 50 µm. (E) Comparative densitometric analysis of CBS+ cells’ activity; #—significant intergroup differences (p < 0.05) between groups of intensely, moderately labeled cells, and immunonegative cells (n = 5 in each group). One-way ANOVA. Intensity of CBS labeling, UOD. (F) Comparative distribution of CBS+ cells in VD, VV, and VL; significant intergroup differences # (p < 0.05), ## (p < 0.01) between the groups of intensively, moderately labeled cells, and immunonegative cells (n = 5 in each group). One-way ANOVA.
Figure 6.
Cystathionine β-synthase (CBS) in the subpallium of intact juvenile chum salmon, Oncorhynchus keta. (A) General view of the subpallial region of the telencephalon, the pictogram shows the zones of the ventral telencephalon (subpallium), Vd—dorsal, Vv—ventral, Vl—lateral, (outlined with red rectangles) areas; (B) dorsal subpallial zone (Vd) at higher magnification, the inset (in the black rectangle) shows CBS+ granules in PVZ and SVZ (yellow arrows), a fragment (red inset) including CBS–cells surrounded by an aggregation of small elongated basophilic cells (white arrows), clusters of CBS+ cells in PZ (in a white rectangle); (C) ventral subpallial region (Vv), CBS+ granules (yellow arrows) on the apical surface of PVZ neuroepithelial cells and clusters in SVZ (black inset), elongated moderately labeled CBS cells and granules inside and on the surface of CBS cells (red inset), nuclei stained with methyl green are indicated by an asterisk, a green arrow indicates the figure of mitosis; (D) the lateral subpallial region (Vl) of the accumulation of small, intensely labeled cells (in the red oval); multiple CBS+ granules (in the black rectangle) were located in PZ on the surface of CBS cells. Immunohistochemical labeling of cystathionine β-synthase. Scale bars: (A) 100 µm, (B–D) 50 µm. (E) Comparative densitometric analysis of CBS+ cells’ activity; #—significant intergroup differences (p < 0.05) between groups of intensely, moderately labeled cells, and immunonegative cells (n = 5 in each group). One-way ANOVA. Intensity of CBS labeling, UOD. (F) Comparative distribution of CBS+ cells in VD, VV, and VL; significant intergroup differences # (p < 0.05), ## (p < 0.01) between the groups of intensively, moderately labeled cells, and immunonegative cells (n = 5 in each group). One-way ANOVA.

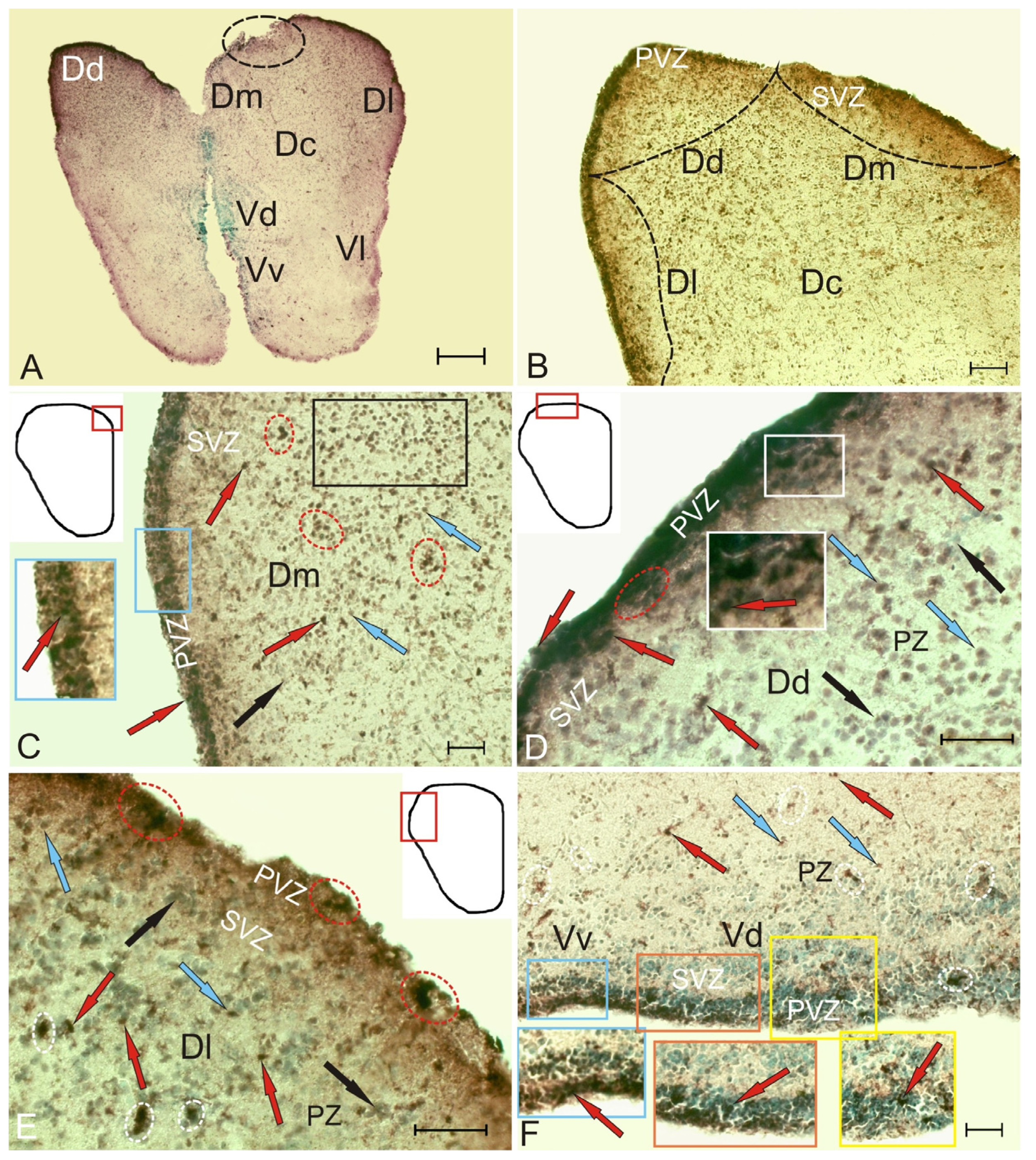
Figure 7.
Cystathionine β-synthase in the pallium and subpallium of juvenile chum salmon, Oncorhynchus keta, at 3 days after the traumatic injury to the telencephalon. (A) General view of CBS immunolocalization in the telencephalon, where the black dotted line indicates the area of injury; (B) general view of the pallial zone, where black dotted lines indicate the boundaries of the pallial subzones; (C) small clusters of CBS+ cells in PVZ and SVZ (inset), CBS+ cells in PZ located discretely (red arrows) or forming small clusters (in red ovals), a large accumulation of CBS+ cells in PVZ (in the black rectangle); (D) the number of heterogeneous CBS+ cells and their aggregations (in the red oval) in SVZ increased in DD (inset); (E) clusters of CBS cells (black arrows), including moderately CBS+ cells (blue arrows), appeared in DL clusters containing intensely labeled cells in PVZ (in the red oval), in SVZ, clusters with intensely CBS-labeled cells appeared (in white ovals); (F) a layer of CBS+ cells occurred in the subpallium in VD in PVZ (yellow inset), SVZ (red inset), and VV (blue inset); single and paired CBS+ cells and their small clusters (in white ovals) in PZ. Immunohistochemical labeling of cystathionine β-synthase. Scale bars: (A) 500 µm; (B,C,F) 100 µm; (D,E) 50 µm.
Figure 7.
Cystathionine β-synthase in the pallium and subpallium of juvenile chum salmon, Oncorhynchus keta, at 3 days after the traumatic injury to the telencephalon. (A) General view of CBS immunolocalization in the telencephalon, where the black dotted line indicates the area of injury; (B) general view of the pallial zone, where black dotted lines indicate the boundaries of the pallial subzones; (C) small clusters of CBS+ cells in PVZ and SVZ (inset), CBS+ cells in PZ located discretely (red arrows) or forming small clusters (in red ovals), a large accumulation of CBS+ cells in PVZ (in the black rectangle); (D) the number of heterogeneous CBS+ cells and their aggregations (in the red oval) in SVZ increased in DD (inset); (E) clusters of CBS cells (black arrows), including moderately CBS+ cells (blue arrows), appeared in DL clusters containing intensely labeled cells in PVZ (in the red oval), in SVZ, clusters with intensely CBS-labeled cells appeared (in white ovals); (F) a layer of CBS+ cells occurred in the subpallium in VD in PVZ (yellow inset), SVZ (red inset), and VV (blue inset); single and paired CBS+ cells and their small clusters (in white ovals) in PZ. Immunohistochemical labeling of cystathionine β-synthase. Scale bars: (A) 500 µm; (B,C,F) 100 µm; (D,E) 50 µm.

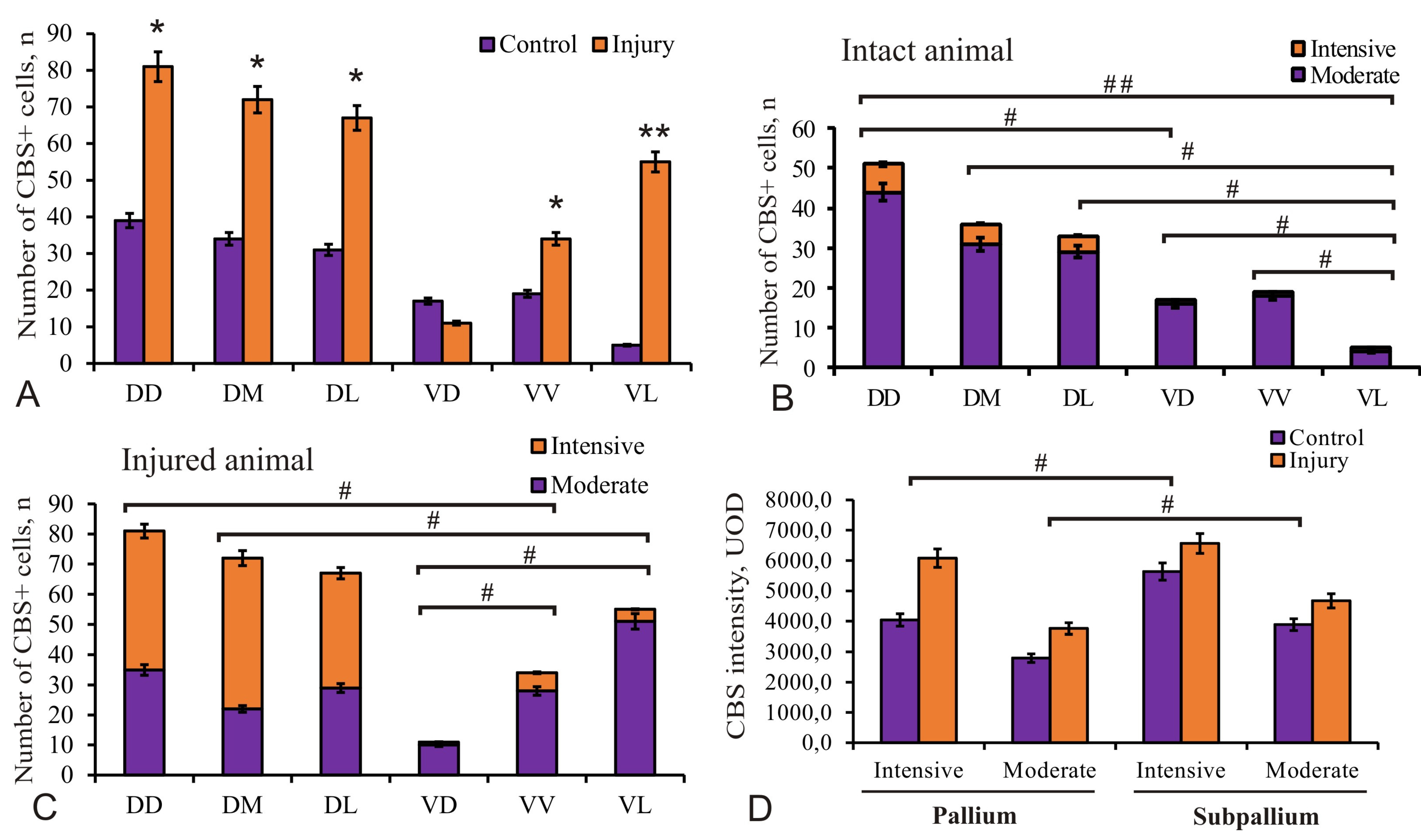
Figure 8.
Quantitative ratio of CBS+/CBS–cells in the pallium and subpallium of intact juvenile chum salmon Oncorhynchus keta and at 3 days after the telencephalon injury. (A) Quantitative proportion of CBS+ cells in intact animals (control group) and in those at 3 days after the traumatic injury to the telencephalon (n = 5 in each group, * p ≤ 0.05; ** p ≤ 0.01; significant difference vs. control groups). One-way ANOVA followed by the Student–Newman–Keuls post hoc test. (B) Ratio of CBS+ intensely/moderately labeled cells in intact animals; (C) on day 3 post-injury, significant intergroup differences # (p < 0.05), ## (p < 0.01) between the groups of intensely, moderately labeled cells, and immunonegative cells (n = 5 in each group). One-way ANOVA. (D) Comparative densitometric analysis of the activity of CBS+ cells in the pallium and subpallium in intact animals and on day 3 days post-injury; significant intergroup differences # (p < 0.05), ## (p < 0.01) between the groups of intensely, moderately labeled cells, and immunonegative cells (n = 5 in each group). One-way ANOVA. Intensity of CBS labeling, UOD.
Figure 8.
Quantitative ratio of CBS+/CBS–cells in the pallium and subpallium of intact juvenile chum salmon Oncorhynchus keta and at 3 days after the telencephalon injury. (A) Quantitative proportion of CBS+ cells in intact animals (control group) and in those at 3 days after the traumatic injury to the telencephalon (n = 5 in each group, * p ≤ 0.05; ** p ≤ 0.01; significant difference vs. control groups). One-way ANOVA followed by the Student–Newman–Keuls post hoc test. (B) Ratio of CBS+ intensely/moderately labeled cells in intact animals; (C) on day 3 post-injury, significant intergroup differences # (p < 0.05), ## (p < 0.01) between the groups of intensely, moderately labeled cells, and immunonegative cells (n = 5 in each group). One-way ANOVA. (D) Comparative densitometric analysis of the activity of CBS+ cells in the pallium and subpallium in intact animals and on day 3 days post-injury; significant intergroup differences # (p < 0.05), ## (p < 0.01) between the groups of intensely, moderately labeled cells, and immunonegative cells (n = 5 in each group). One-way ANOVA. Intensity of CBS labeling, UOD.

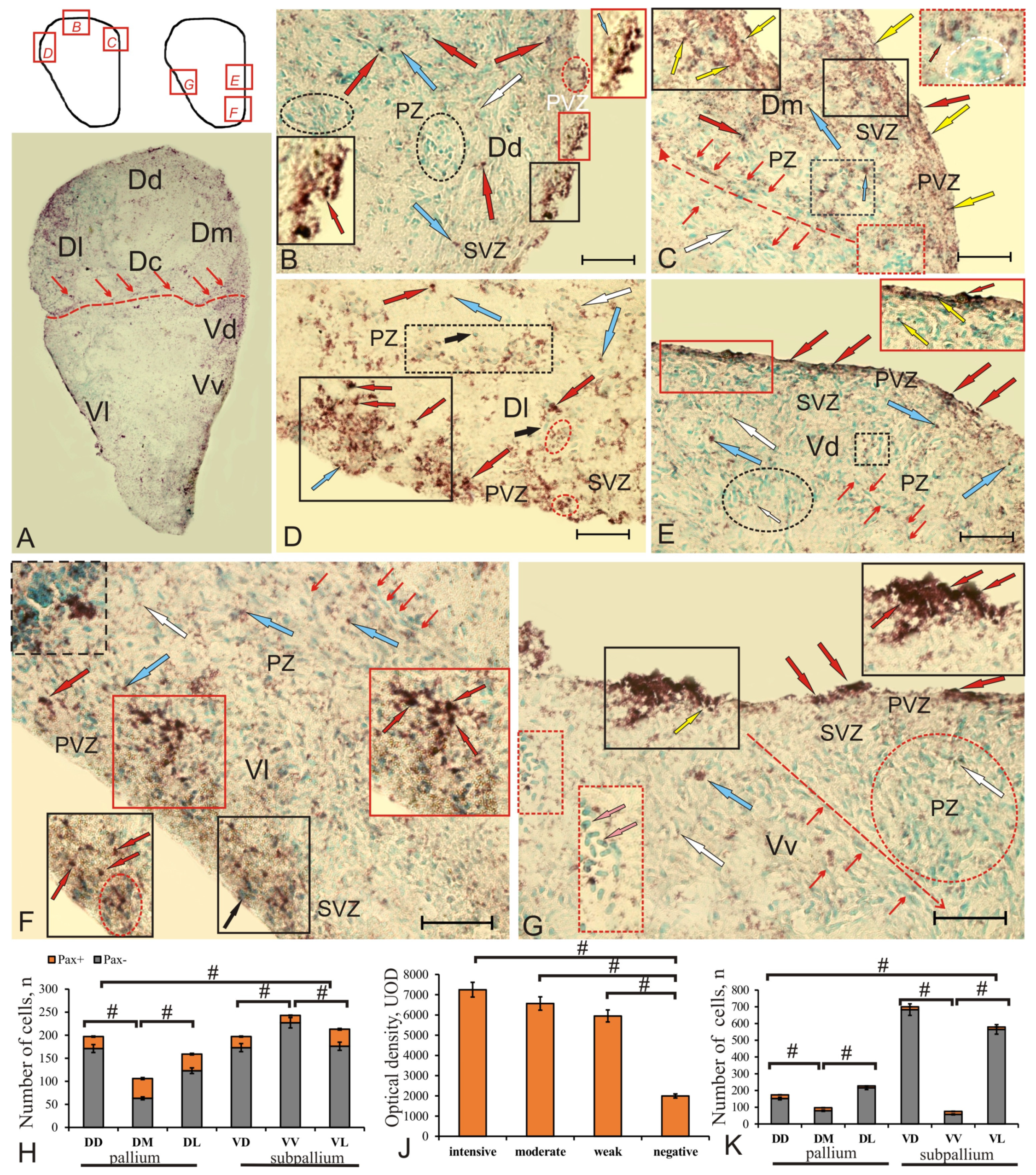
Figure 9.
Pax2 expression in the pallium and subpallium of intact juvenile chum salmon, Oncorhynchus keta. (A) General view of the Pax2 expression pattern in the telencephalon of intact juvenile chum salmon where the boundaries between the pallial and subpallial regions are indicated by the red dashed line, Pax2+ cells along the border are indicated by thin red arrows; the pictograms indicate the areas of the brain shown in Figures (B–D) and (E–G). (C) Pax2 expression in DD, clusters of Pax2-expressing cells (red arrow) in PVZ (black inset) and nuclei (blue arrow, shown in red inset); the aggregation of nuclei is outlined by the red oval, and in black ovals there are aggregations of immunonegative cells in PZ, white arrows indicate weakly labeled cells; (C) patterns of Pax2 expression in DM; immunolabeling of Pax2 was detected in nuclei (shown by yellow arrows), which form extended regions in PVZ and SVZ (black inset). the granular pattern of Pax2 expression predominated in PZ, and the red dotted line indicates the radial direction of neuroblasts’ migration from constitutive neurogenic niches (CNN) (red inset) containing clusters of Pax2– (encircled by the white dotted line) and Pax2+ cells; (D) morphological heterogeneity of Pax2+ cells in DL, where the black rectangle outlines nuclei labeled with Pax2; Pax2+ cells were found in the basal part of PVZ and SVZ, both cellular (in the dashed rectangle) and nuclear expression of Pax2 (in the red ovals); (E) in PVZ of the dorsal subpallial region (VD), Pax2+ cells (red arrows) had a surface tangential orientation (red inset) and in SVZ and PZ there were moderately (in the dotted square) and weakly labeled Pax2 cells (in the oval), migrating cells (thin red arrows) and nuclei (yellow arrows) that form morphogenetic fields; (F) In VL, individual Pax2+ nuclei were encountered in PVZ (black inset); in SVZ, individual intensely labeled clusters of cells (red inset) were identified. In VL, local Pax2– cell clusters were identified in combination with dense clusters of Pax2+ cells (in dotted rectangle ); (G) intensely labeled clusters of Pax2+ cells (black inset) were detected in PVZ and the aggregation of immunonegative basophilic cells was found in PZ (indicated by pink arrows, red inset); the red dotted line indicates the direction of neuroblast migration and in the red oval there is a morphogenetic field with weak expression of Pax2 and neuroblasts. Immunohistochemical labeling of the protein product of the Pax2 transcription factor. Scale bars: (A) 500 µm; (B,C,F) 100 µm; (D,E) 50 µm. (H) Ratio of Pax2+/Pax2–cells in the pallium and subpallium of intact chum salmon (n = 5 in each group, #—significant intergroup differences (p < 0.05)). One-way ANOVA. (J) Comparative densitometric analysis of Pax2+ cells; significant intergroup differences # (p < 0.05) between groups of intensively, moderately, weakly labeled, and immunonegative cells (n = 5 in each group). One-way ANOVA. Intensity of Pax2 labeling, UOD. (K) Ratio of the number of Pax2+/Pax2—cells in the pallium and subpallium zones at 3 days after the injury to the telencephalon of juvenile chum salmon; #—significant intergroup differences (n = 5 in each group, #—significant intergroup differences (p < 0.05). One-way ANOVA.
Figure 9.
Pax2 expression in the pallium and subpallium of intact juvenile chum salmon, Oncorhynchus keta. (A) General view of the Pax2 expression pattern in the telencephalon of intact juvenile chum salmon where the boundaries between the pallial and subpallial regions are indicated by the red dashed line, Pax2+ cells along the border are indicated by thin red arrows; the pictograms indicate the areas of the brain shown in Figures (B–D) and (E–G). (C) Pax2 expression in DD, clusters of Pax2-expressing cells (red arrow) in PVZ (black inset) and nuclei (blue arrow, shown in red inset); the aggregation of nuclei is outlined by the red oval, and in black ovals there are aggregations of immunonegative cells in PZ, white arrows indicate weakly labeled cells; (C) patterns of Pax2 expression in DM; immunolabeling of Pax2 was detected in nuclei (shown by yellow arrows), which form extended regions in PVZ and SVZ (black inset). the granular pattern of Pax2 expression predominated in PZ, and the red dotted line indicates the radial direction of neuroblasts’ migration from constitutive neurogenic niches (CNN) (red inset) containing clusters of Pax2– (encircled by the white dotted line) and Pax2+ cells; (D) morphological heterogeneity of Pax2+ cells in DL, where the black rectangle outlines nuclei labeled with Pax2; Pax2+ cells were found in the basal part of PVZ and SVZ, both cellular (in the dashed rectangle) and nuclear expression of Pax2 (in the red ovals); (E) in PVZ of the dorsal subpallial region (VD), Pax2+ cells (red arrows) had a surface tangential orientation (red inset) and in SVZ and PZ there were moderately (in the dotted square) and weakly labeled Pax2 cells (in the oval), migrating cells (thin red arrows) and nuclei (yellow arrows) that form morphogenetic fields; (F) In VL, individual Pax2+ nuclei were encountered in PVZ (black inset); in SVZ, individual intensely labeled clusters of cells (red inset) were identified. In VL, local Pax2– cell clusters were identified in combination with dense clusters of Pax2+ cells (in dotted rectangle ); (G) intensely labeled clusters of Pax2+ cells (black inset) were detected in PVZ and the aggregation of immunonegative basophilic cells was found in PZ (indicated by pink arrows, red inset); the red dotted line indicates the direction of neuroblast migration and in the red oval there is a morphogenetic field with weak expression of Pax2 and neuroblasts. Immunohistochemical labeling of the protein product of the Pax2 transcription factor. Scale bars: (A) 500 µm; (B,C,F) 100 µm; (D,E) 50 µm. (H) Ratio of Pax2+/Pax2–cells in the pallium and subpallium of intact chum salmon (n = 5 in each group, #—significant intergroup differences (p < 0.05)). One-way ANOVA. (J) Comparative densitometric analysis of Pax2+ cells; significant intergroup differences # (p < 0.05) between groups of intensively, moderately, weakly labeled, and immunonegative cells (n = 5 in each group). One-way ANOVA. Intensity of Pax2 labeling, UOD. (K) Ratio of the number of Pax2+/Pax2—cells in the pallium and subpallium zones at 3 days after the injury to the telencephalon of juvenile chum salmon; #—significant intergroup differences (n = 5 in each group, #—significant intergroup differences (p < 0.05). One-way ANOVA.

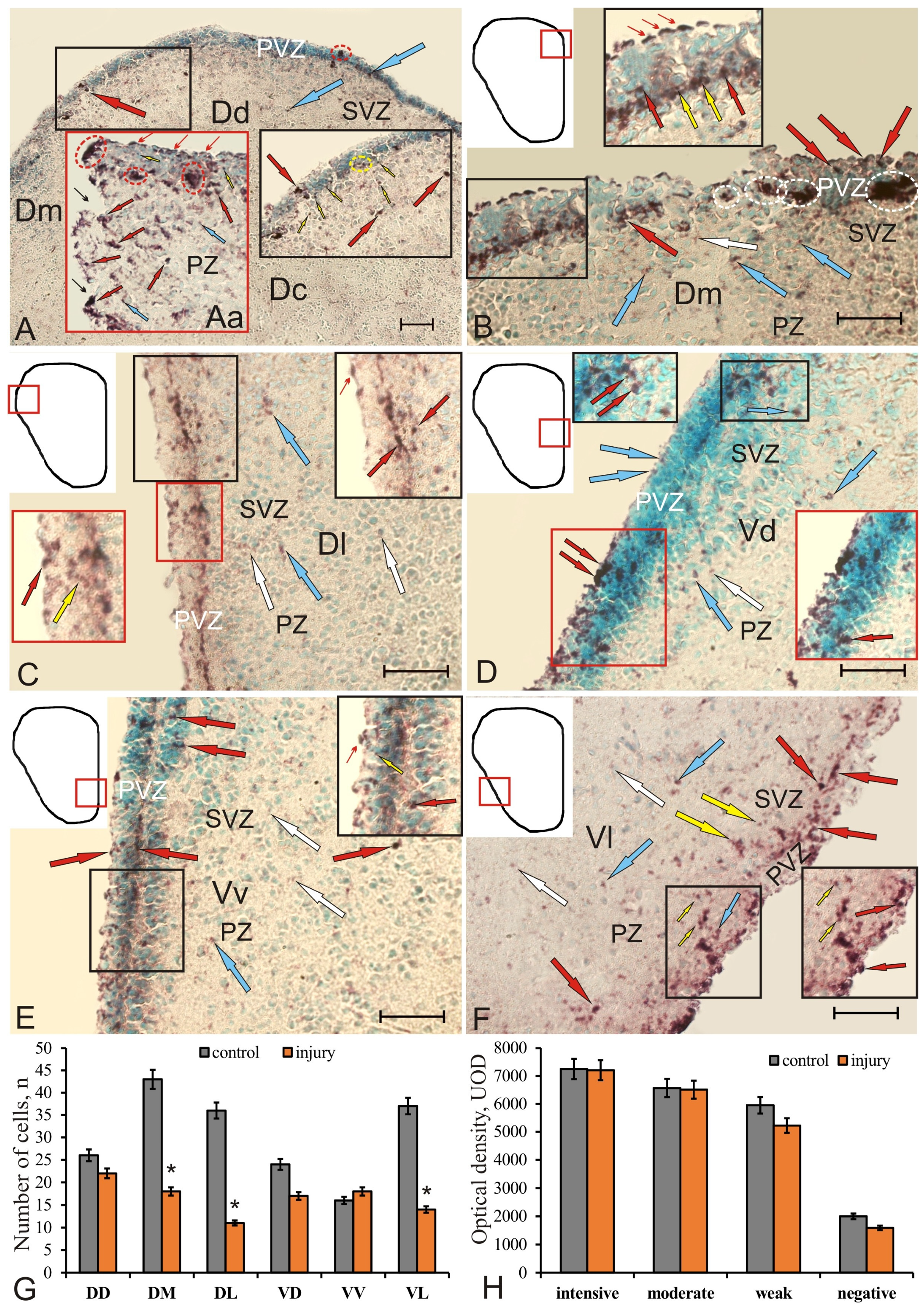
Figure 10.
Pax2 expression in the pallium and subpallium of juvenile chum salmon, Oncorhynchus keta, at 3 days after the traumatic injury. (A) Pattern of Pax2 expression in DD, immunopositive cells (red arrows) and nuclei (yellow arrows) localized in PVZ and SVZ (in the black inset), where the aggregation of cells in PVZ is outlined by the red oval, nuclei (in the yellow oval); (Aa, in a red rectangle) in the area of injury, black narrow arrows indicate the traumatic lumen, RNN in red ovals; cells migrating to the area of injury are indicated by red narrow arrows, blue arrows indicate moderately labeled nuclei; (B) Pax2 expression in DM, migrating to the injury area, reactive cells in PVZ (red thin arrows, inset), local dense clusters of Pax2+ cells in the basal part of PVZ and SVZ (in white ovals), nuclei (yellow arrows); (C) Pax2 expression in DL, tangentially migrating cells in the apical and basal parts of PVZ (black inset), neuroepithelial cells (red arrow) and nuclei (yellow arrow in the red inset); (D) Pax2 expression in VD; Pax2+ nuclei and neuroepithelial cells were labeled with surface localization in PVZ (red inset) and SVZ (black inset), moderately labeled cells (blue arrows), weakly labeled cells (white arrows); (E) Pax2 expression in VV, numerous Pax2+ cells and nuclei were identified in the apical and basal parts of PVZ (inset); in SVZ and PZ, individual moderately (blue arrows) and weakly labeled Pax2 (white arrows) cells and nuclei prevailed; (F) in VL, an increased distribution of Pax2+ cells and their accumulations in the surface and basal parts of PVZ (inset), designations as in E. Immunohistochemical labeling of the protein product of the transcription factor Pax2. Scale bars: (A) 100 µm; (B–F) 50 µm. (G) Quantitative ratio of Pax2+ cells in intact animals (control group) and at 3 days after the traumatic injury to the telencephalon (n = 5 in each group, * p ≤ 0.05; significant difference vs. control groups). One-way ANOVA followed by the Student–Newman–Keuls post hoc test. (H) Comparative densitometric analysis of Pax2+ and Pax2– cells’ activity in intact juvenile chum salmon and at 3 days after the telencephalon injury (M ± SD). Intensity of Pax2 labeling, UOD.
Figure 10.
Pax2 expression in the pallium and subpallium of juvenile chum salmon, Oncorhynchus keta, at 3 days after the traumatic injury. (A) Pattern of Pax2 expression in DD, immunopositive cells (red arrows) and nuclei (yellow arrows) localized in PVZ and SVZ (in the black inset), where the aggregation of cells in PVZ is outlined by the red oval, nuclei (in the yellow oval); (Aa, in a red rectangle) in the area of injury, black narrow arrows indicate the traumatic lumen, RNN in red ovals; cells migrating to the area of injury are indicated by red narrow arrows, blue arrows indicate moderately labeled nuclei; (B) Pax2 expression in DM, migrating to the injury area, reactive cells in PVZ (red thin arrows, inset), local dense clusters of Pax2+ cells in the basal part of PVZ and SVZ (in white ovals), nuclei (yellow arrows); (C) Pax2 expression in DL, tangentially migrating cells in the apical and basal parts of PVZ (black inset), neuroepithelial cells (red arrow) and nuclei (yellow arrow in the red inset); (D) Pax2 expression in VD; Pax2+ nuclei and neuroepithelial cells were labeled with surface localization in PVZ (red inset) and SVZ (black inset), moderately labeled cells (blue arrows), weakly labeled cells (white arrows); (E) Pax2 expression in VV, numerous Pax2+ cells and nuclei were identified in the apical and basal parts of PVZ (inset); in SVZ and PZ, individual moderately (blue arrows) and weakly labeled Pax2 (white arrows) cells and nuclei prevailed; (F) in VL, an increased distribution of Pax2+ cells and their accumulations in the surface and basal parts of PVZ (inset), designations as in E. Immunohistochemical labeling of the protein product of the transcription factor Pax2. Scale bars: (A) 100 µm; (B–F) 50 µm. (G) Quantitative ratio of Pax2+ cells in intact animals (control group) and at 3 days after the traumatic injury to the telencephalon (n = 5 in each group, * p ≤ 0.05; significant difference vs. control groups). One-way ANOVA followed by the Student–Newman–Keuls post hoc test. (H) Comparative densitometric analysis of Pax2+ and Pax2– cells’ activity in intact juvenile chum salmon and at 3 days after the telencephalon injury (M ± SD). Intensity of Pax2 labeling, UOD.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









