
Centrifugation Removes a Population of Large Vesicles, or “Macroparticles,” Intermediate in Size to RBCs and Microvesicles


Abstract
1. Introduction
2. Results
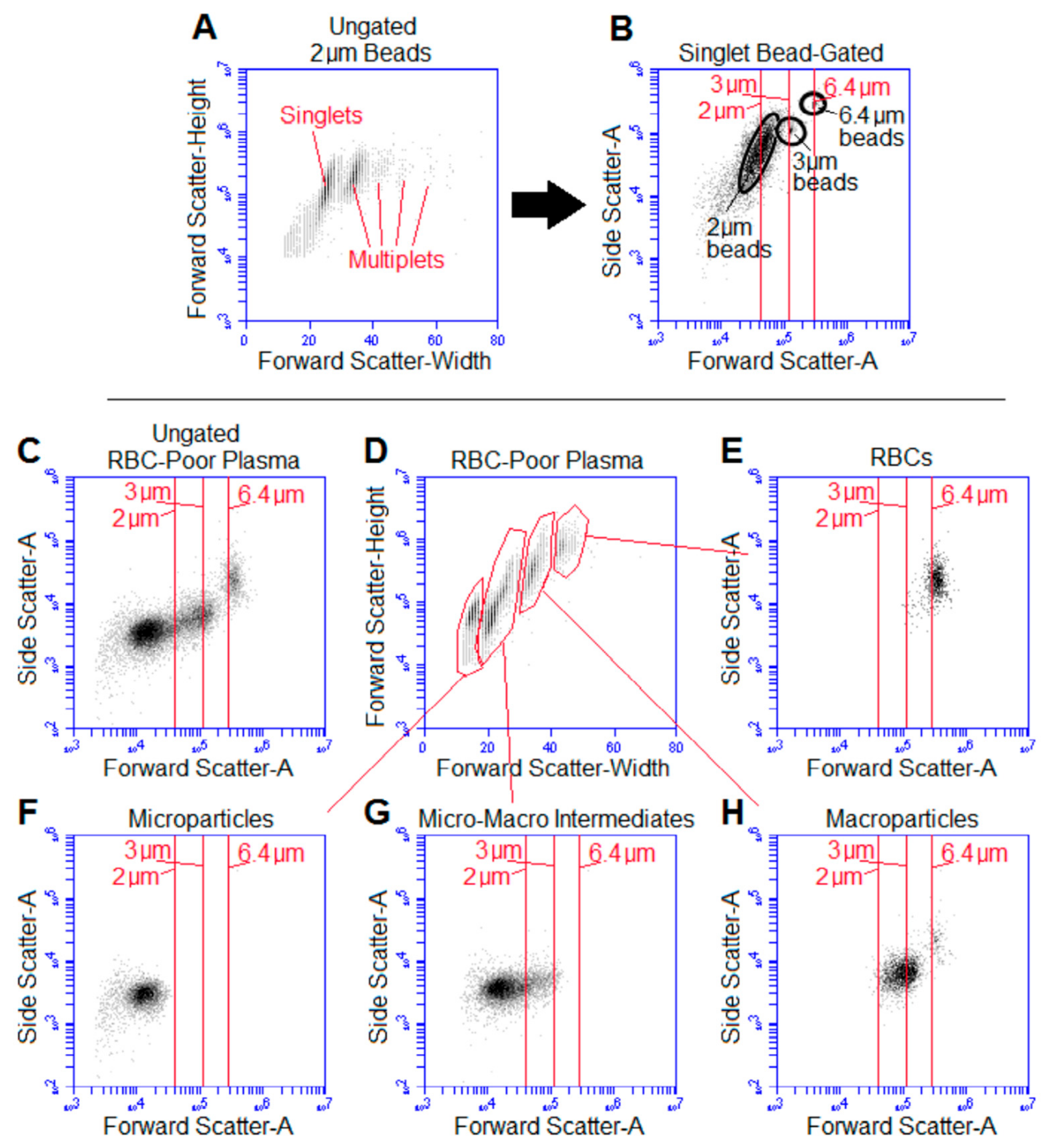
2.1. Gentle Sedimentation Revealed Vesicles Larger than 1 µm in Diameter
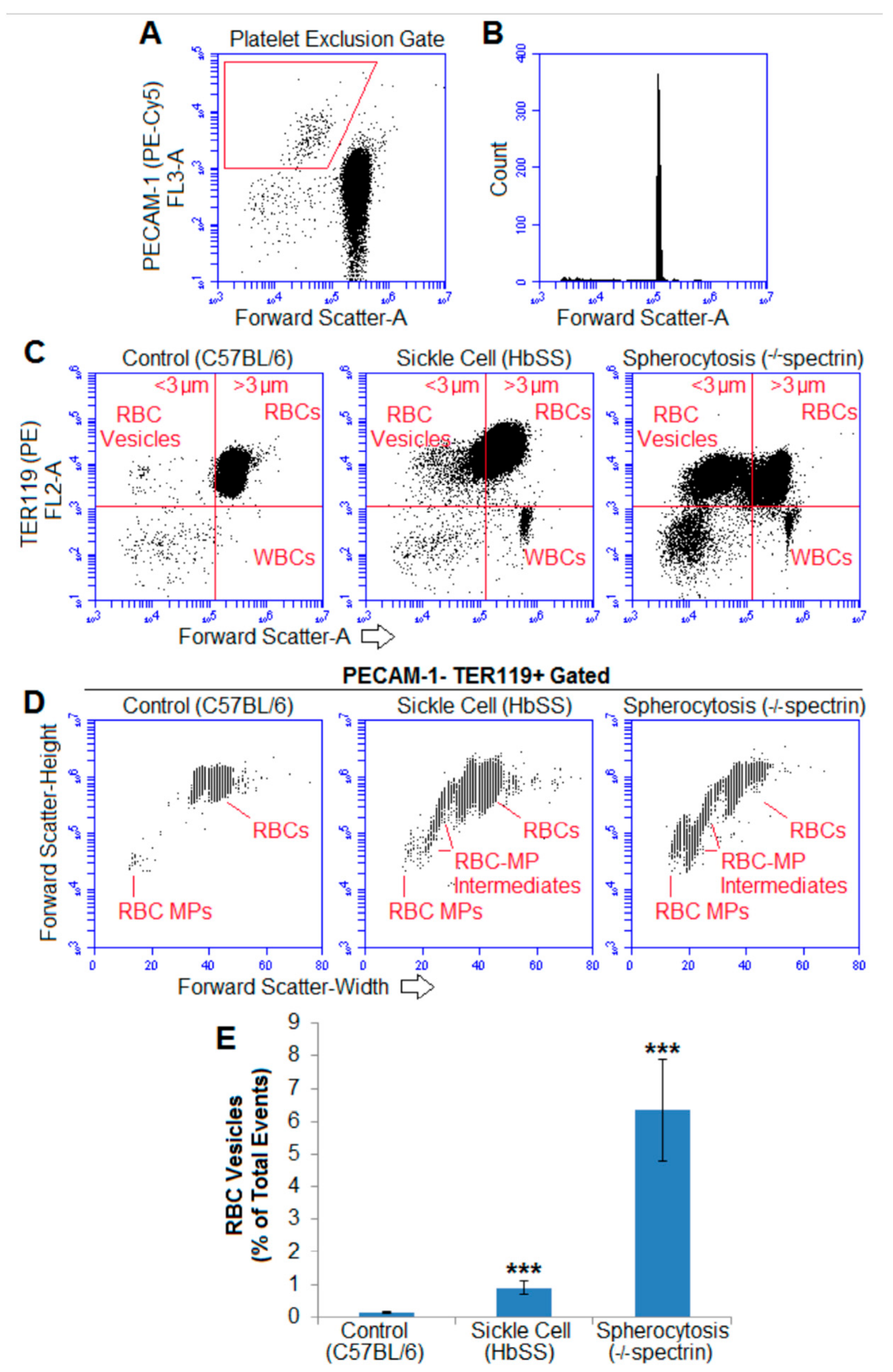
2.2. Size Determination of Cell-to-Microparticle Intermediate “Macroparticles”
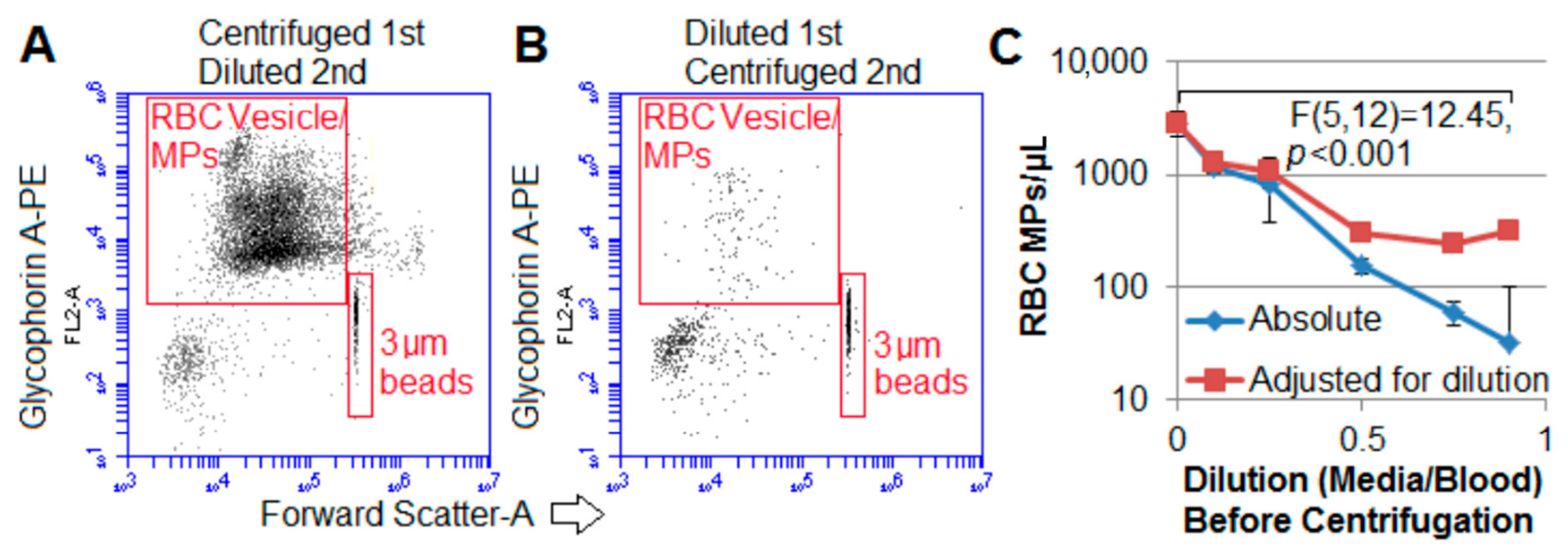
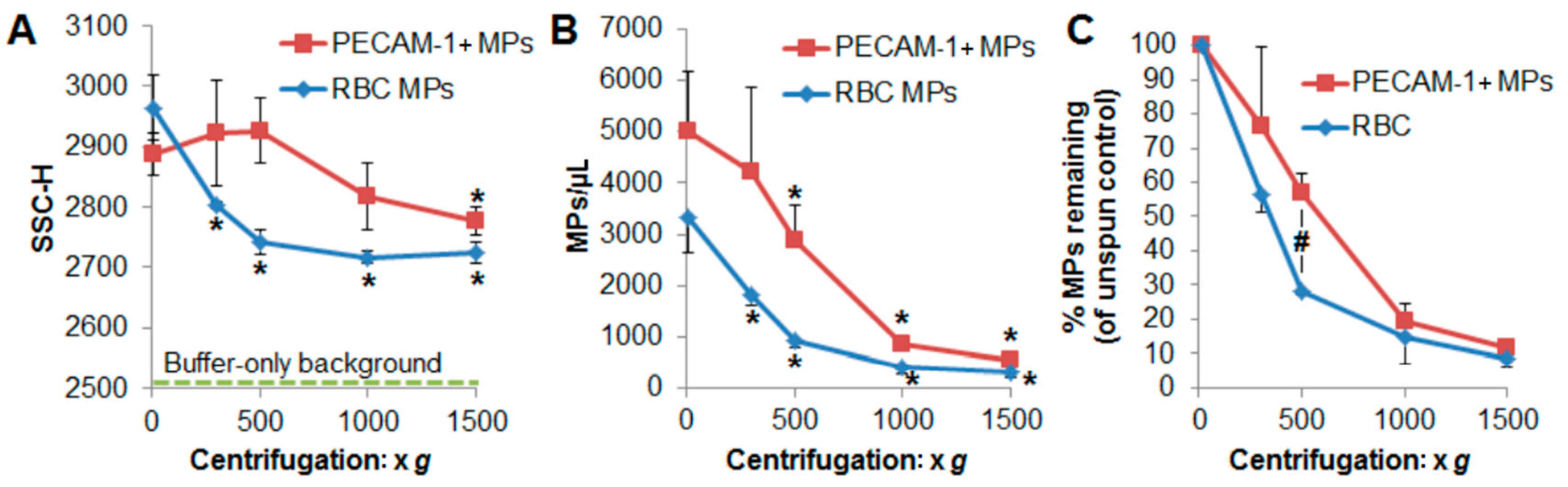
2.3. Centrifugation Removes Macroparticles
2.4. RBC-Derived MVs Sediment Was Relatively Faster than PECAM-1 + MVs
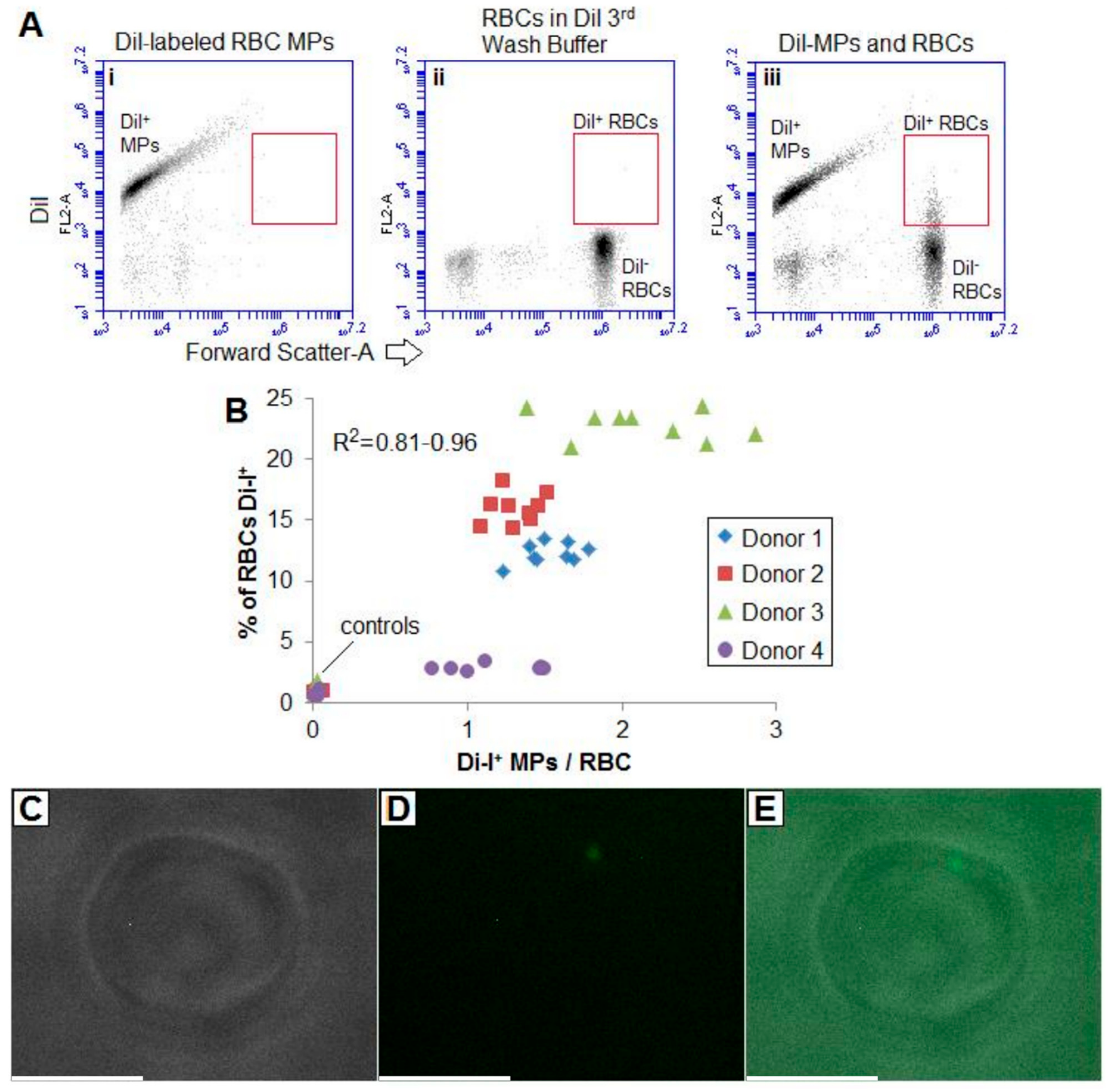
2.5. MVs May Adhere to RBCs
2.6. RBC-Derived Macroparticles Are Also Present in Murine Hemolytic Blood
3. Discussion
4. Materials and Methods
4.1. RBC and RMV Isolation
4.2. Cell and MV Staining
4.3. Flow Cytometry
4.4. Fluorescence Microscopy
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| FSC | Forward Scatter |
| MP/MV | Microparticles or microvesicles |
| PECAM-1 | Platelet Endothelial Cell Adhesion Molecule-1 |
| PMV | Platelet-derived microvesicles |
| RMV | Red blood cell-derived microvesicles |
| SSC | Side Scatter |
| PS | Phosphatidylserine |
| SCD | Sickle Cell Disease |
References
- Dignat-George, F.; Boulanger, C.M. The Many Faces of Endothelial Microparticles. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Rak, J. Microparticles in Cancer. Semin. Thromb. Hemost. 2010, 36, 888–906. [Google Scholar] [CrossRef]
- Wang, B.; Li, T.; Han, X.; Li, Y.; Cheng, W.; Wang, L.; Lu, Z.; Yang, J.; Zhao, M. The Level of Circulating Microparticles in Patients with Coronary Heart Disease: A Systematic Review and Meta-Analysis. J Cardiovasc. Transl. Res. 2020, 13, 702–712. [Google Scholar] [CrossRef]
- Willekens, F.L.A.; Werre, J.M.; Groenen-Döpp, Y.A.M.; Roerdinkholder-Stoelwinder, B.; Pauw, B.D.; Bosman, G.J.C.G.M. Erythrocyte Vesiculation: A Self-Protective Mechanism? Br. J. Haematol. 2008, 141, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Zucker-Franklin, D.; Karpatkin, S. Red-Cell and Platelet Fragmentation in Idiopathic Autoimmune Thrombocytopenic Purpura. N. Engl. J. Med. 1977, 297, 517–523. [Google Scholar] [CrossRef]
- Larson, M.C.; Hillery, C.A.; Hogg, N. Circulating Membrane-Derived Microvesicles in Redox Biology. Free Radic. Biol. Med. 2014, 73, 214–228. [Google Scholar] [CrossRef] [PubMed]
- Kriebardis, A.G.; Antonelou, M.H.; Stamoulis, K.E.; Economou-Petersen, E.; Margaritis, L.H.; Papassideri, I.S. RBC-Derived Vesicles during Storage: Ultrastructure, Protein Composition, Oxidation, and Signaling Components. Transfusion (Paris) 2008, 48, 1943–1953. [Google Scholar] [CrossRef] [PubMed]
- Nantakomol, D.; Palasuwan, A.; Chaowanathikhom, M.; Soogarun, S.; Imwong, M. Red Cell and Platelet-Derived Microparticles Are Increased in G6PD-Deficient Subjects. Eur. J. Haematol. 2012, 89, 423–429. [Google Scholar] [CrossRef]
- Rubin, O.; Crettaz, D.; Canellini, G.; Tissot, J.-D.; Lion, N. Microparticles in Stored Red Blood Cells: An Approach Using Flow Cytometry and Proteomic Tools. Vox Sang. 2008, 95, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Tissot, J.-D.; Rubin, O.; Canellini, G. Analysis and Clinical Relevance of Microparticles from Red Blood Cells. Curr. Opin. Hematol. 2010, 17, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Larson, M.C.; Woodliff, J.E.; Hillery, C.A.; Kearl, T.J.; Zhao, M. Phosphatidylethanolamine Is Externalized at the Surface of Microparticles. Biochim. Biophys. Acta 2012, 1821, 1501–1507. [Google Scholar] [CrossRef] [PubMed]
- Tavoosi, N.; Davis-Harrison, R.L.; Pogorelov, T.V.; Ohkubo, Y.Z.; Arcario, M.J.; Clay, M.C.; Rienstra, C.M.; Tajkhorshid, E.; Morrissey, J.H. Molecular Determinants of Phospholipid Synergy in Blood Clotting. J. Biol. Chem. 2011, 286, 23247–23253. [Google Scholar] [CrossRef] [PubMed]
- Fadok, V.A.; Bratton, D.L.; Rose, D.M.; Pearson, A.; Ezekewitz, R.A.; Henson, P.M. A Receptor for Phosphatidylserine-Specific Clearance of Apoptotic Cells. Nature 2000, 405, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Willekens, F.L.A.; Werre, J.M.; Kruijt, J.K.; Roerdinkholder-Stoelwinder, B.; Groenen-Döpp, Y.A.M.; van den Bos, A.G.; Bosman, G.J.C.G.M.; van Berkel, T.J.C. Liver Kupffer Cells Rapidly Remove Red Blood Cell-Derived Vesicles from the Circulation by Scavenger Receptors. Blood 2005, 105, 2141–2145. [Google Scholar] [CrossRef] [PubMed]
- Simhadri, V.R.; Andersen, J.F.; Calvo, E.; Choi, S.-C.; Coligan, J.E.; Borrego, F. Human CD300a Binds to Phosphatidylethanolamine and Phosphatidylserine, and Modulates the Phagocytosis of Dead Cells. Blood 2012, 119, 2799–2809. [Google Scholar] [CrossRef]
- Frey, B.; Gaipl, U.S. The Immune Functions of Phosphatidylserine in Membranes of Dying Cells and Microvesicles. Semin. Immunopathol. 2011, 33, 497–516. [Google Scholar] [CrossRef]
- Sadallah, S.; Eken, C.; Schifferli, J.A. Ectosomes as Immunomodulators. Semin. Immunopathol. 2010, 33, 487–495. [Google Scholar] [CrossRef]
- Zecher, D.; Cumpelik, A.; Schifferli, J.A. Erythrocyte-Derived Microvesicles Amplify Systemic Inflammation by Thrombin-Dependent Activation of Complement. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 313–320. [Google Scholar] [CrossRef]
- Noubouossie, D.F.; Henderson, M.W.; Mooberry, M.; Ilich, A.; Ellsworth, P.; Piegore, M.; Skinner, S.C.; Pawlinski, R.; Welsby, I.; Renné, T.; et al. Red Blood Cell Microvesicles Activate the Contact System, Leading to Factor IX Activation via 2 Independent Pathways. Blood 2020, 135, 755–765. [Google Scholar] [CrossRef]
- Shet, A.S.; Aras, O.; Gupta, K.; Hass, M.J.; Rausch, D.J.; Saba, N.; Koopmeiners, L.; Key, N.S.; Hebbel, R.P. Sickle Blood Contains Tissue Factor-Positive Microparticles Derived from Endothelial Cells and Monocytes. Blood 2003, 102, 2678–2683. [Google Scholar] [CrossRef]
- Donadee, C.; Raat, N.J.H.; Kanias, T.; Tejero, J.; Lee, J.S.; Kelley, E.E.; Zhao, X.; Liu, C.; Reynolds, H.; Azarov, I.; et al. Nitric Oxide Scavenging by Red Blood Cell Microparticles and Cell-Free Hemoglobin as a Mechanism for the Red Cell Storage Lesion. Circulation 2011, 124, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Vlaar, A.P.J.; Hofstra, J.J.; Levi, M.; Kulik, W.; Nieuwland, R.; Tool, A.T.J.; Schultz, M.J.; de Korte, D.; Juffermans, N.P. Supernatant of Aged Erythrocytes Causes Lung Inflammation and Coagulopathy in a “Two-Hit” in Vivo Syngeneic Transfusion Model. Anesthesiology 2010, 113, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Jy, W.; Ricci, M.; Shariatmadar, S.; Gomez-Marin, O.; Horstman, L.H.; Ahn, Y.S. Microparticles in Stored Red Blood Cells as Potential Mediators of Transfusion Complications. Transfusion (Paris) 2011, 51, 886–893. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, M.K.; Tamagne, M.; Elayeb, R.; Andrieu, M.; Pirenne, F.; Vingert, B. Blood Microparticles Are a Component of Immune Modulation in Red Blood Cell Transfusion. Eur. J. Immunol. 2020, 50, 1237–1240. [Google Scholar] [CrossRef] [PubMed]
- Van Beers, E.J.; Schaap, M.C.L.; Berckmans, R.J.; Nieuwland, R.; Sturk, A.; van Doormaal, F.F.; Meijers, J.C.M.; Biemond, B.J. CURAMA study group Circulating Erythrocyte-Derived Microparticles Are Associated with Coagulation Activation in Sickle Cell Disease. Haematologica 2009, 94, 1513–1519. [Google Scholar] [CrossRef]
- Camus, S.M.; Gausserès, B.; Bonnin, P.; Loufrani, L.; Grimaud, L.; Charue, D.; De Moraes, J.A.; Renard, J.-M.; Tedgui, A.; Boulanger, C.M.; et al. Erythrocyte Microparticles Can Induce Kidney Vaso-Occlusions in a Murine Model of Sickle Cell Disease. Blood 2012, 120, 5050–5058. [Google Scholar] [CrossRef]
- Mullier, F.; Bailly, N.; Chatelain, C.; Chatelain, B.; Dogné, J.-M. Pre-Analytical Issues in the Measurement of Circulating Microparticles: Current Recommendations and Pending Questions. J. Thromb. Haemost. 2013, 11, 693–696. [Google Scholar] [CrossRef]
- Cointe, S.; Judicone, C.; Robert, S.; Mooberry, M.J.; Poncelet, P.; Wauben, M.; Nieuwland, R.; Key, N.S.; Dignat-George, F.; Lacroix, R. Standardization of Microparticle Enumeration across Different Flow Cytometry Platforms: Results of a Multicenter Collaborative Workshop. J. Thromb. Haemost. 2017, 15, 187–193. [Google Scholar] [CrossRef]
- Poncelet, P.; Robert, S.; Bouriche, T.; Bez, J.; Lacroix, R.; Dignat-George, F. Standardized Counting of Circulating Platelet Microparticles Using Currently Available Flow Cytometers and Scatter-Based Triggering: Forward or Side Scatter? Cytom. Part J. Int. Soc. Anal. Cytol. 2016, 89, 148–158. [Google Scholar] [CrossRef]
- Barr, J.D.; Chauhan, A.K.; Schaeffer, G.V.; Hansen, J.K.; Motto, D.G. Red Blood Cells Mediate the Onset of Thrombosis in the Ferric Chloride Murine Model. Blood 2013, 121, 3733–3741. [Google Scholar] [CrossRef]
- Shet, A.S. Characterizing Blood Microparticles: Technical Aspects and Challenges. Vasc. Health Risk Manag. 2008, 4, 769–774. [Google Scholar] [CrossRef][Green Version]
- Dey-Hazra, E.; Hertel, B.; Kirsch, T.; Woywodt, A.; Lovric, S.; Haller, H.; Haubitz, M.; Erdbruegger, U. Detection of Circulating Microparticles by Flow Cytometry: Influence of Centrifugation, Filtration of Buffer, and Freezing. Vasc. Health Risk Manag. 2010, 6, 1125–1133. [Google Scholar] [CrossRef] [PubMed]
- Yuana, Y.; Bertina, R.M.; Osanto, S. Pre-Analytical and Analytical Issues in the Analysis of Blood Microparticles. Thromb. Haemost. 2011, 105, 396–408. [Google Scholar] [CrossRef] [PubMed]
- Van Ierssel, S.H.; Van Craenenbroeck, E.M.; Conraads, V.M.; Van Tendeloo, V.F.; Vrints, C.J.; Jorens, P.G.; Hoymans, V.Y. Flow Cytometric Detection of Endothelial Microparticles (EMP): Effects of Centrifugation and Storage Alter with the Phenotype Studied. Thromb. Res. 2010, 125, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Larson, M.C.; Karafin, M.S.; Hillery, C.A.; Hogg, N. Phosphatidylethanolamine Is Progressively Exposed in RBCs during Storage. Transfus. Med. 2017, 27, 136–141. [Google Scholar] [CrossRef]
- Larson, M.C.; Luthi, M.R.; Hogg, N.; Hillery, C.A. Calcium-Phosphate Microprecipitates Mimic Microparticles When Examined with Flow Cytometry. Cytom. Part J. Int. Soc. Anal. Cytol. 2013, 83, 242–250. [Google Scholar] [CrossRef][Green Version]
- Pankoui Mfonkeu, J.B.; Gouado, I.; Fotso Kuaté, H.; Zambou, O.; Amvam Zollo, P.H.; Grau, G.E.R.; Combes, V. Elevated Cell-Specific Microparticles Are a Biological Marker for Cerebral Dysfunctions in Human Severe Malaria. PLoS ONE 2010, 5, e13415. [Google Scholar] [CrossRef]
- Mrvar-Brecko, A.; Sustar, V.; Jansa, V.; Stukelj, R.; Jansa, R.; Mujagić, E.; Kruljc, P.; Iglic, A.; Hägerstrand, H.; Kralj-Iglic, V. Isolated Microvesicles from Peripheral Blood and Body Fluids as Observed by Scanning Electron Microscope. Blood Cells Mol. Dis. 2010, 44, 307–312. [Google Scholar] [CrossRef]
- Lelkes, G.; Fodor, I. Formation of Large, Membrane Skeleton-Free Erythrocyte Vesicles as a Function of the Intracellular PH and Temperature. Biochim. Biophys. Acta 1991, 1065, 135–144. [Google Scholar] [CrossRef]
- Leonards, K.S.; Ohki, S. Isolation and Characterization of Large (0.5–1.0 Micron) Cytoskeleton-Free Vesicles from Human and Rabbit Erythrocytes. Biochim. Biophys. Acta 1983, 728, 383–393. [Google Scholar] [CrossRef]
- Allan, D.; Thomas, P.; Limbrick, A.R. The Isolation and Characterization of 60 Nm Vesicles (‘nanovesicles’) Produced during Ionophore A23187-Induced Budding of Human Erythrocytes. Biochem. J. 1980, 188, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Allan, D.; Billah, M.M.; Finean, J.B.; Michell, R.H. Release of Diacylglycerol-Enriched Vesicles from Erythrocytes with Increased Intracellular (Ca2+). Nature 1976, 261, 58–60. [Google Scholar] [CrossRef]
- Mullier, F.; Bailly, N.; Chatelain, C.; Dogné, J.M.; Chatelain, B. More on: Calibration for the Measurement of Microparticles: Needs, Interests, and Limitations of Calibrated Polystyrene Beads for Flow Cytometry-Based Quantification of Biological Microparticles. J. Thromb. Haemost. 2011, 9, 1679–1681; author reply 1681–1682. [Google Scholar] [CrossRef] [PubMed]
- Van der Pol, E.; van Gemert, M.J.C.; Sturk, A.; Nieuwland, R.; van Leeuwen, T.G. Single vs. Swarm Detection of Microparticles and Exosomes by Flow Cytometry. J. Thromb. Haemost. 2012, 10, 919–930. [Google Scholar] [CrossRef] [PubMed]
- Momen-Heravi, F.; Balaj, L.; Alian, S.; Trachtenberg, A.J.; Hochberg, F.H.; Skog, J.; Kuo, W.P. Impact of Biofluid Viscosity on Size and Sedimentation Efficiency of the Isolated Microvesicles. Front. Physiol. 2012, 3, 162. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, R.; Robert, S.; Poncelet, P.; Dignat-George, F. Overcoming Limitations of Microparticle Measurement by Flow Cytometry. Semin. Thromb. Hemost. 2010, 36, 807–818. [Google Scholar] [CrossRef]
- Kina, T.; Ikuta, K.; Takayama, E.; Wada, K.; Majumdar, A.S.; Weissman, I.L.; Katsura, Y. The Monoclonal Antibody TER-119 Recognizes a Molecule Associated with Glycophorin A and Specifically Marks the Late Stages of Murine Erythroid Lineage. Br. J. Haematol. 2000, 109, 280–287. [Google Scholar] [CrossRef]
- Ponder, E. The Relation Between Red Blood Cell Density and Corpuscular Hemoglobin Concentration. J. Biol. Chem. 1942, 144, 333–338. [Google Scholar] [CrossRef]
- Corash, L.; Tan, H.; Gralnick, H.R. Heterogeneity of Human Whole Blood Platelet Subpopulations. I. Relationship between Buoyant Density, Cell Volume, and Ultrastructure. Blood 1977, 49, 71–87. [Google Scholar] [CrossRef]
- Lacroix, R.; Robert, S.; Poncelet, P.; Kasthuri, R.S.; Key, N.S.; Dignat-George, F. ISTH SSC Workshop Standardization of Platelet-Derived Microparticle Enumeration by Flow Cytometry with Calibrated Beads: Results of the International Society on Thrombosis and Haemostasis SSC Collaborative Workshop. J. Thromb. Haemost. 2010, 8, 2571–2574. [Google Scholar] [CrossRef] [PubMed]
- Azarov, I.; Liu, C.; Reynolds, H.; Tsekouras, Z.; Lee, J.S.; Gladwin, M.T.; Kim-Shapiro, D.B. Mechanisms of Slower Nitric Oxide Uptake by Red Blood Cells and Other Hemoglobin-Containing Vesicles. J. Biol. Chem. 2011, 286, 33567–33579. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.C.; Hein, T.W.; Vaughn, M.W.; Huang, K.T.; Kuo, L. Intravascular Flow Decreases Erythrocyte Consumption of Nitric Oxide. Proc. Natl. Acad. Sci. USA 1999, 96, 8757–8761. [Google Scholar] [CrossRef] [PubMed]
- Wandersee, N.J.; Punzalan, R.C.; Rettig, M.P.; Kennedy, M.D.; Pajewski, N.M.; Sabina, R.L.; Paul Scott, J.; Low, P.S.; Hillery, C.A. Erythrocyte Adhesion Is Modified by Alterations in Cellular Tonicity and Volume. Br. J. Haematol. 2005, 131, 366–377. [Google Scholar] [CrossRef]
- Toth, B.; Nikolajek, K.; Rank, A.; Nieuwland, R.; Lohse, P.; Pihusch, V.; Friese, K.; Thaler, C.J. Gender-Specific and Menstrual Cycle Dependent Differences in Circulating Microparticles. Platelets 2007, 18, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Tushuizen, M.E.; Nieuwland, R.; Rustemeijer, C.; Hensgens, B.E.; Sturk, A.; Heine, R.J.; Diamant, M. Elevated Endothelial Microparticles Following Consecutive Meals Are Associated with Vascular Endothelial Dysfunction in Type 2 Diabetes. Diabetes Care 2007, 30, 728–730. [Google Scholar] [CrossRef][Green Version]
- Gordon, C.; Gudi, K.; Krause, A.; Sackrowitz, R.; Harvey, B.-G.; Strulovici-Barel, Y.; Mezey, J.G.; Crystal, R.G. Circulating Endothelial Microparticles as a Measure of Early Lung Destruction in Cigarette Smokers. Am. J. Respir. Crit. Care Med. 2011, 184, 224–232. [Google Scholar] [CrossRef]
- Brown, M.D.; Feairheller, D.L.; Thakkar, S.; Veerabhadrappa, P.; Park, J.-Y. Racial Differences in Tumor Necrosis Factor-α-Induced Endothelial Microparticles and Interleukin-6 Production. Vasc. Health Risk Manag. 2011, 7, 541–550. [Google Scholar] [CrossRef]
- Pászty, C.; Brion, C.M.; Manci, E.; Witkowska, H.E.; Stevens, M.E.; Mohandas, N.; Rubin, E.M. Transgenic Knockout Mice with Exclusively Human Sickle Hemoglobin and Sickle Cell Disease. Science 1997, 278, 876–878. [Google Scholar] [CrossRef]
- Frei, A.C.; Guo, Y.; Jones, D.W.; Pritchard, K.A.; Fagan, K.A.; Hogg, N.; Wandersee, N.J. Vascular Dysfunction in a Murine Model of Severe Hemolysis. Blood 2008, 112, 398–405. [Google Scholar] [CrossRef]
- Wandersee, N.J.; Birkenmeier, C.S.; Gifford, E.J.; Mohandas, N.; Barker, J.E. Murine Recessive Hereditary Spherocytosis, Sph/Sph, Is Caused by a Mutation in the Erythroid Alpha-Spectrin Gene. Hematol. J. 2000, 1, 235–242. [Google Scholar] [CrossRef]
- Grant, C.W.M.; McConnell, H.M. Glycophorin in Lipid Bilayers. Proc. Natl. Acad. Sci. USA 1974, 71, 4653–4657. [Google Scholar] [CrossRef] [PubMed]
- Furthmayr, H.; Marchesi, V.T. Subunit Structure of Human Erythrocyte Glycophorin A. Biochemistry 1976, 15, 1137–1144. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of RBC Vesicles | Cell/Vesicle Separation | Vesicle Diameter | Examination Method(s) | Reference |
|---|---|---|---|---|
| (Patho)physiologic RBC Damage or Hemolysis | ||||
| Malaria | 1500× g for 15′ | ≤1 µm * | Flow cytometry | [37] |
| Hemolytic anemia, ±overnight storage | 1500× g for 5’ to remove cells | ~0.25–0.30 µm | Light microscopy | [8] |
| Autoimmune thrombocytopenia | 200× g for 8’ to remove cells | Up to 2 µm | Electron microscopy | [5] |
| Various diseases | 1550× g for 20’ to remove cell | 0.05–1 µm | Electron microscopy | [38] |
| During blood storage | ≥870× g for 20’ to remove cells, direct dilution | 3 µm, 2.78 µm | Light microscopy, flow cytometry | This study |
| Laboratory Techniques | ||||
| Heating RBCs to 45 °C, 42 °C with pH ≤ 6 | Filtered through 3 µm membrane | 1 up to 3.5 µm | Electron microscopy | [39] |
| Heating RBCs to 45 °C with EDTA and Ca2+ | Sucrose gradient centrifugation | 0.5–1 µm | Light and electron microscopy | [40] |
| Ionophore A23187 and Ca2+ | Centrifugation to sediment cells | 60 nm, 100–150 nm | Electron microscopy | [41,42] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Larson, M.C.; Hogg, N.; Hillery, C.A. Centrifugation Removes a Population of Large Vesicles, or “Macroparticles,” Intermediate in Size to RBCs and Microvesicles. Int. J. Mol. Sci. 2021, 22, 1243. https://doi.org/10.3390/ijms22031243
Larson MC, Hogg N, Hillery CA. Centrifugation Removes a Population of Large Vesicles, or “Macroparticles,” Intermediate in Size to RBCs and Microvesicles. International Journal of Molecular Sciences. 2021; 22(3):1243. https://doi.org/10.3390/ijms22031243
Chicago/Turabian StyleLarson, Michael C., Neil Hogg, and Cheryl A. Hillery. 2021. "Centrifugation Removes a Population of Large Vesicles, or “Macroparticles,” Intermediate in Size to RBCs and Microvesicles" International Journal of Molecular Sciences 22, no. 3: 1243. https://doi.org/10.3390/ijms22031243
APA StyleLarson, M. C., Hogg, N., & Hillery, C. A. (2021). Centrifugation Removes a Population of Large Vesicles, or “Macroparticles,” Intermediate in Size to RBCs and Microvesicles. International Journal of Molecular Sciences, 22(3), 1243. https://doi.org/10.3390/ijms22031243