
Dual Profile of Environmental Enrichment and Autistic-Like Behaviors in the Maternal Separated Model in Rats

 , and
, and 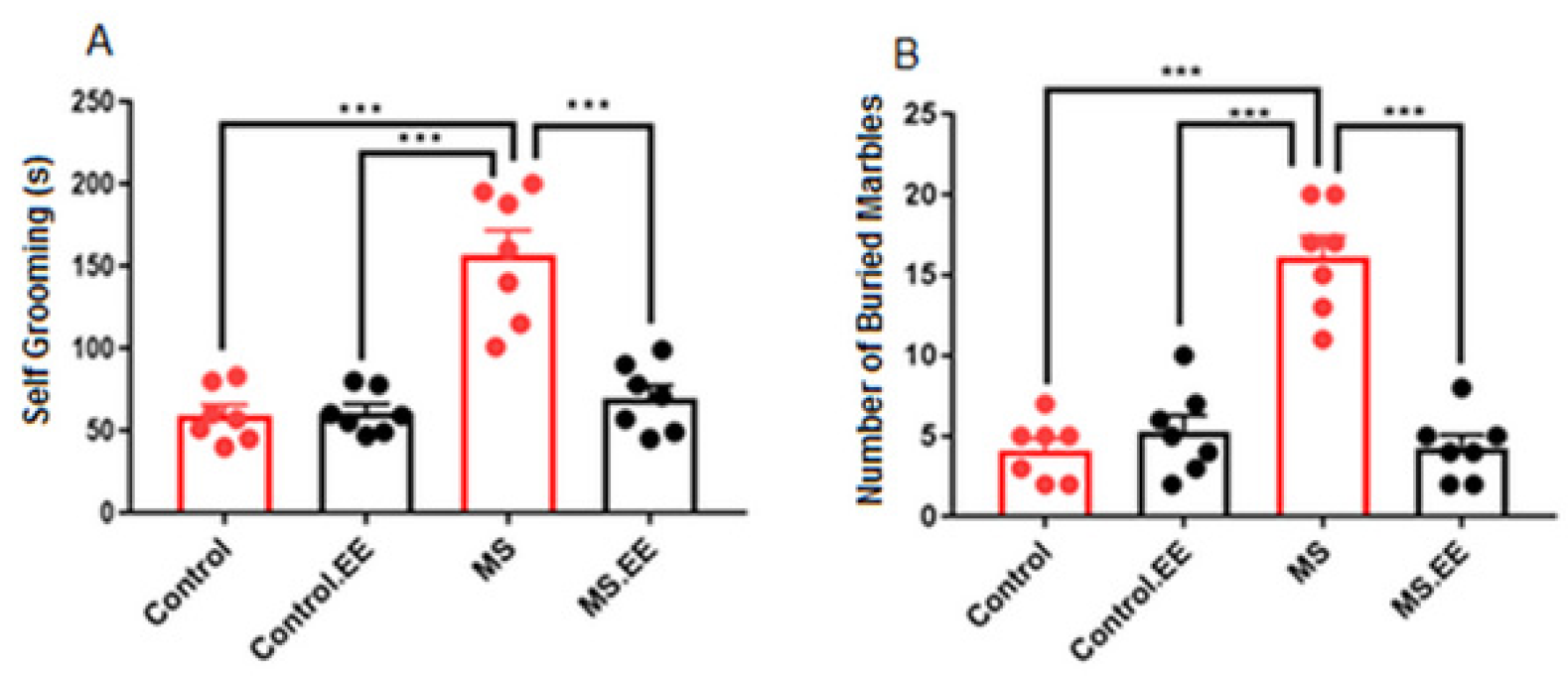{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Repetitive Behaviors
2.1.1. Self-Grooming
2.1.2. Marble Burying
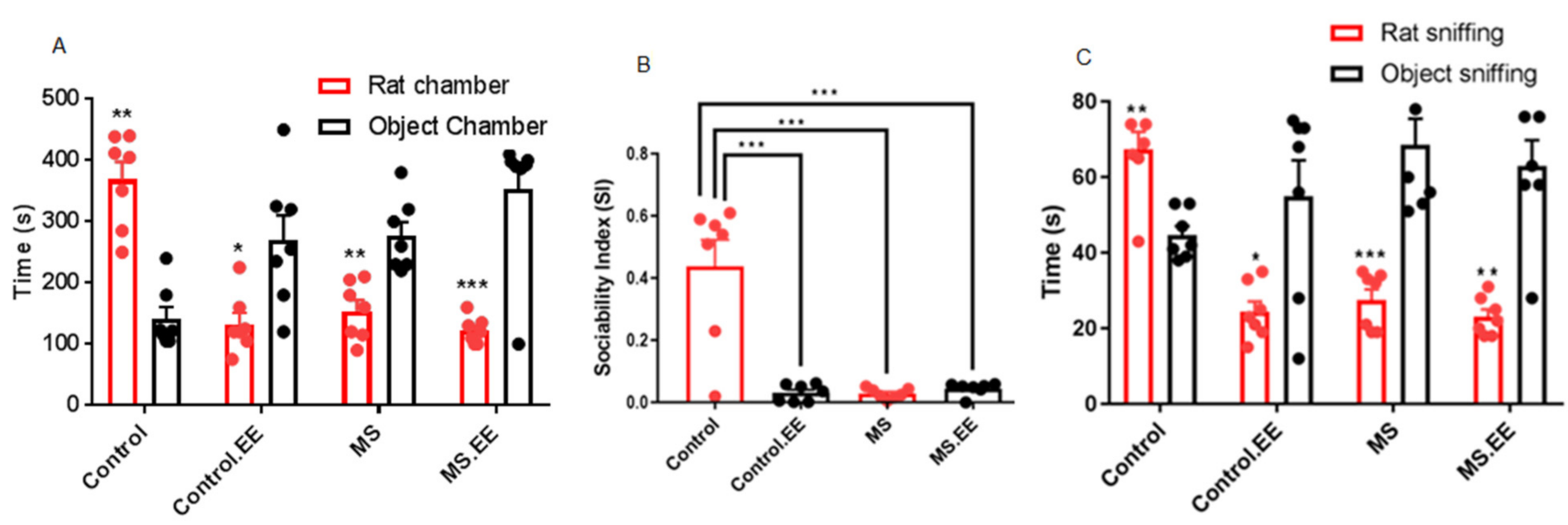
2.2. Social Behavior
2.2.1. Sociability
2.2.2. Direct Social Interaction
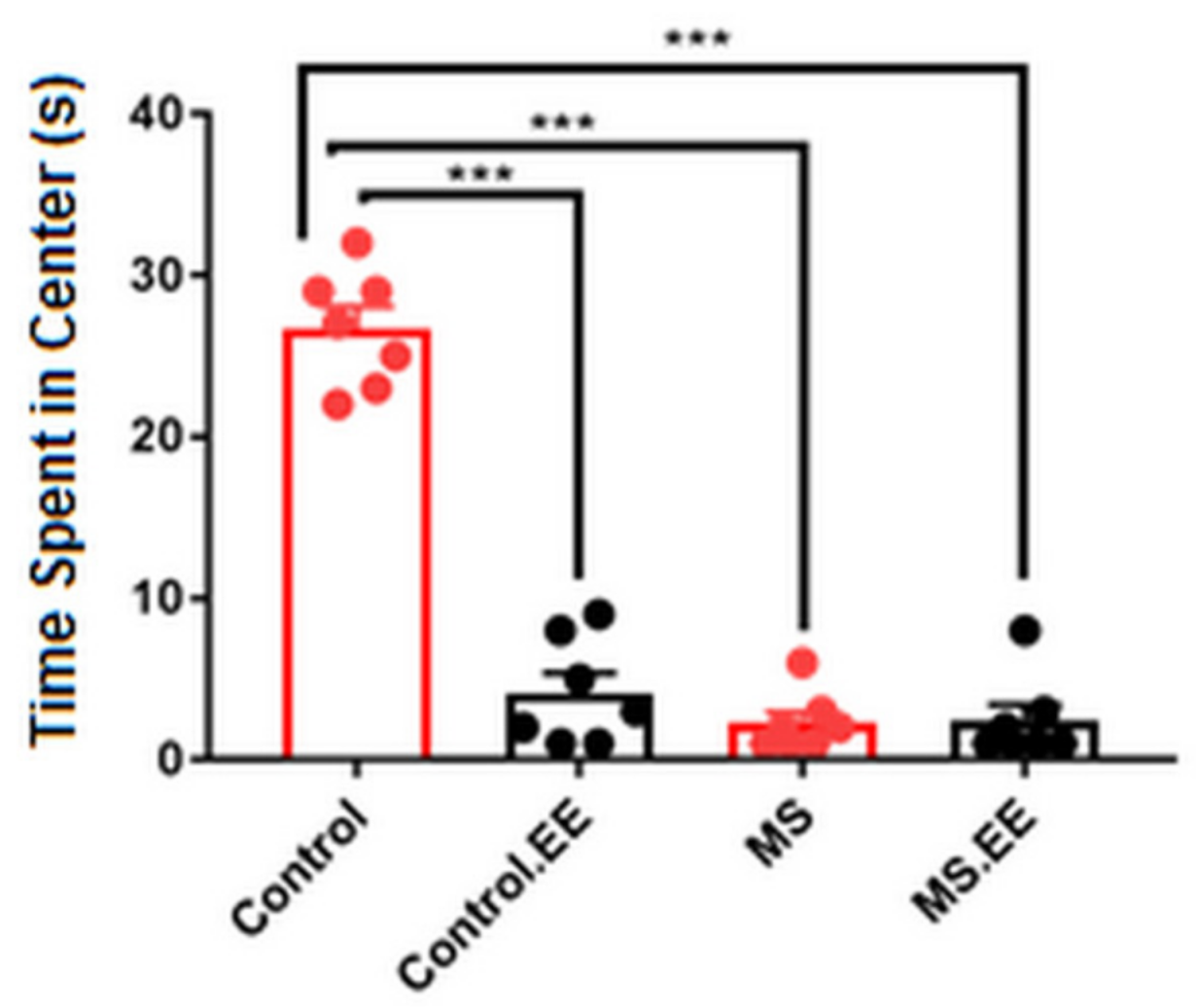
2.2.3. Anxiety-Like Behavior
2.3. Motor Behavior
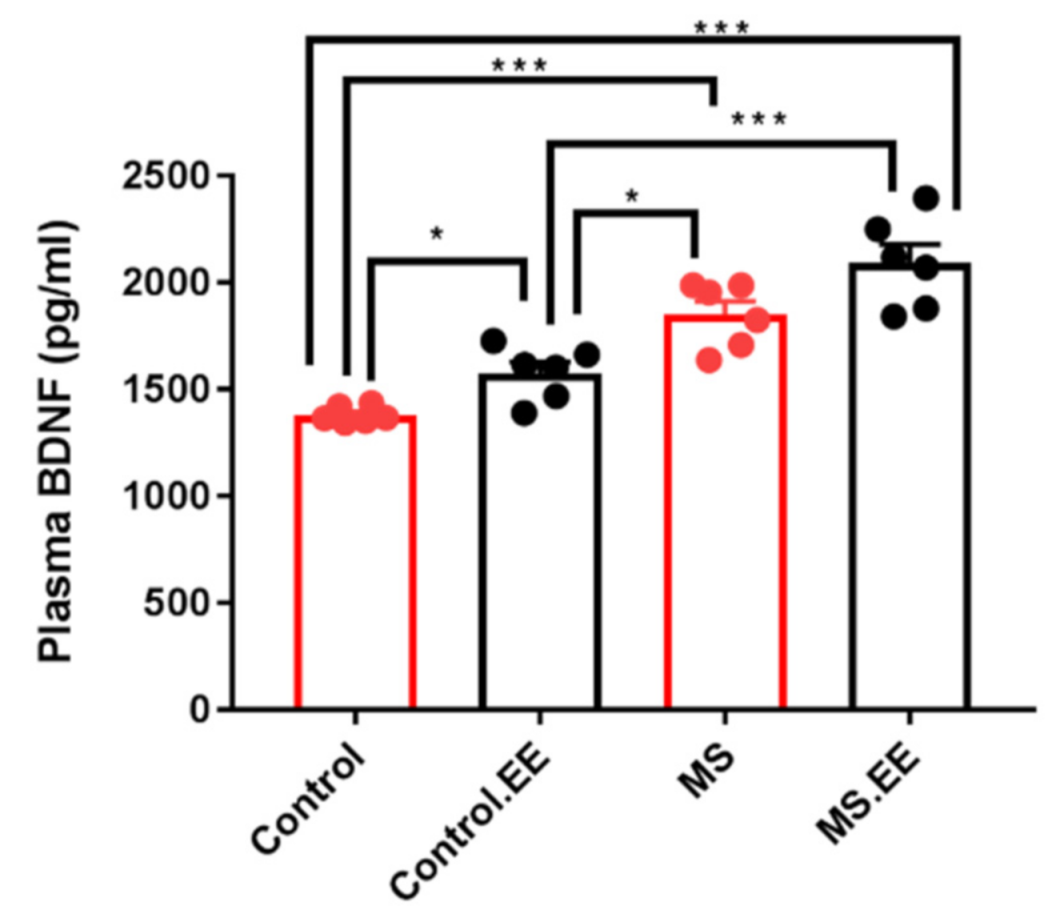
Plasma BDNF Level
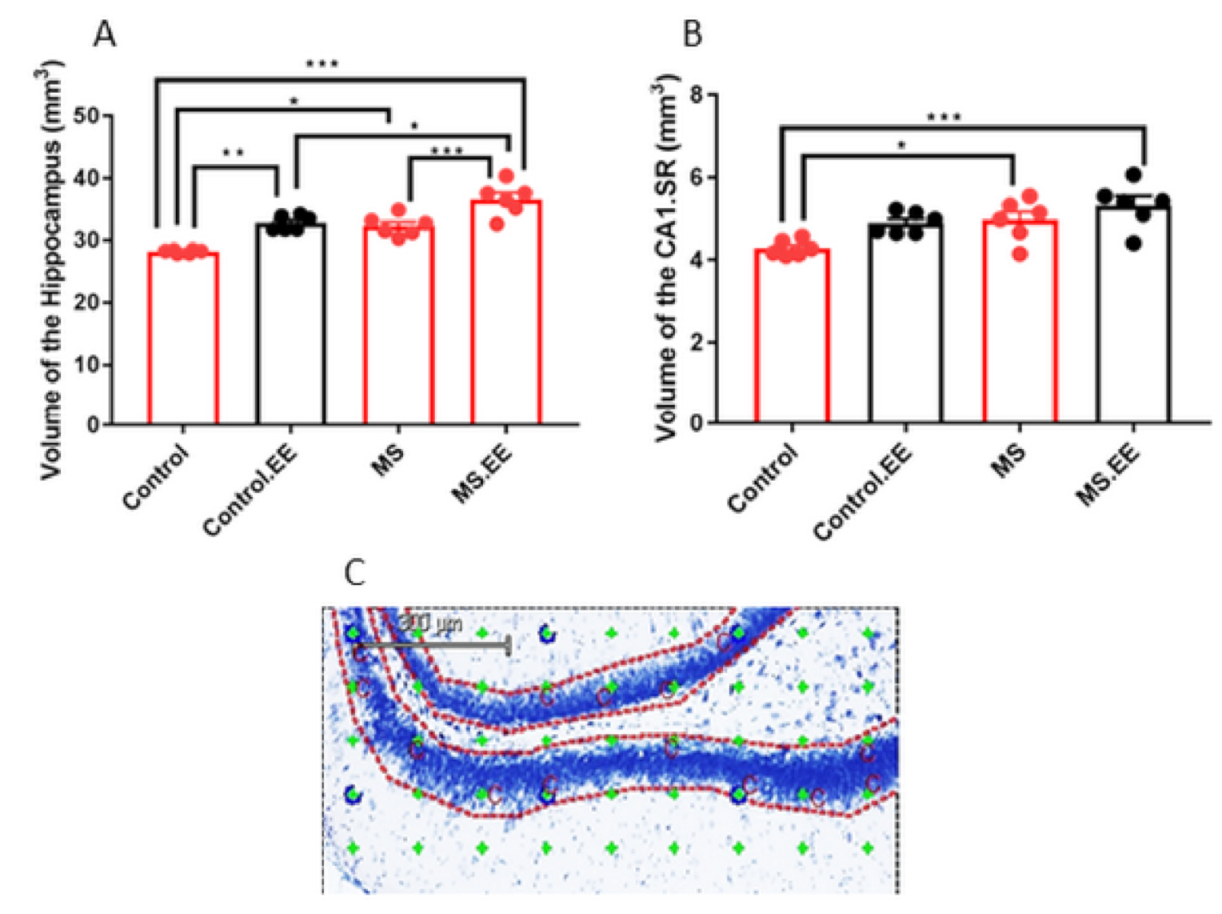
2.4. Quantitative Stereological Measurements
2.4.1. Effects of MS and EE on the Volume of the Hippocampus and Its Sub-Regions
2.4.2. Effects of MS and EE on the Number of Pyramidal Neurons and Volume of the Infra-Limbic and Pre-Limbic Areas
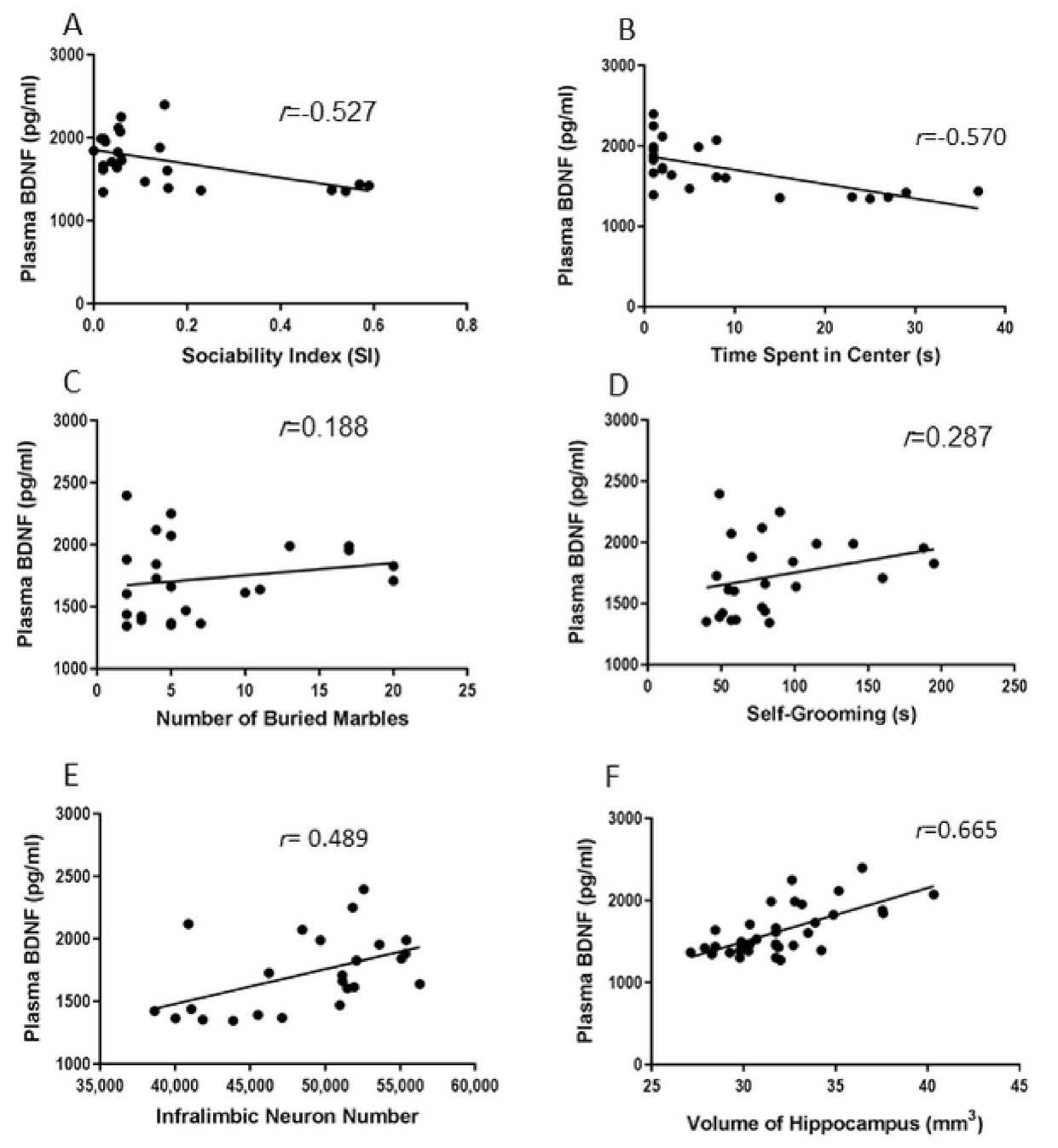
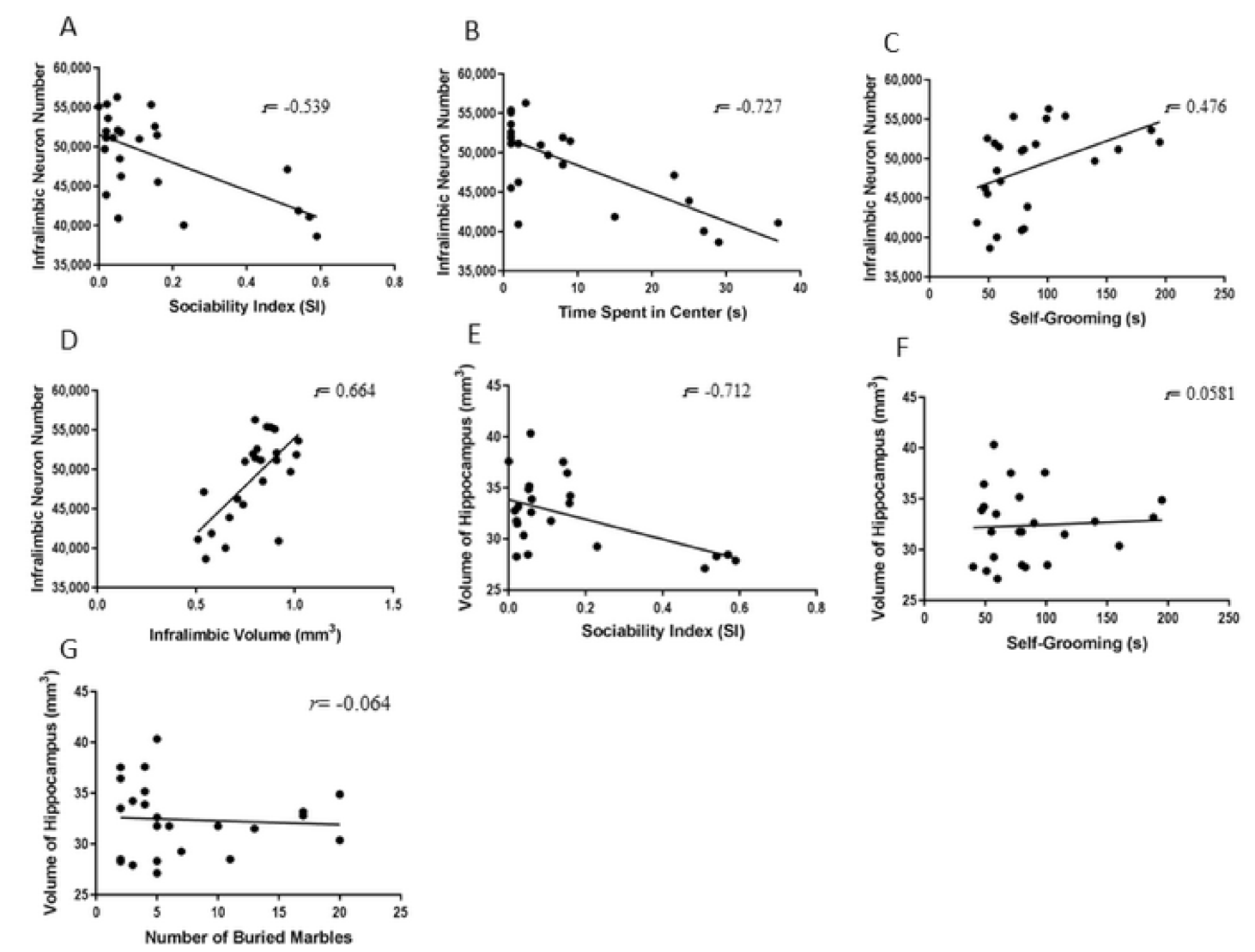
2.5. Correlations between Behavioral and Biological Parameters
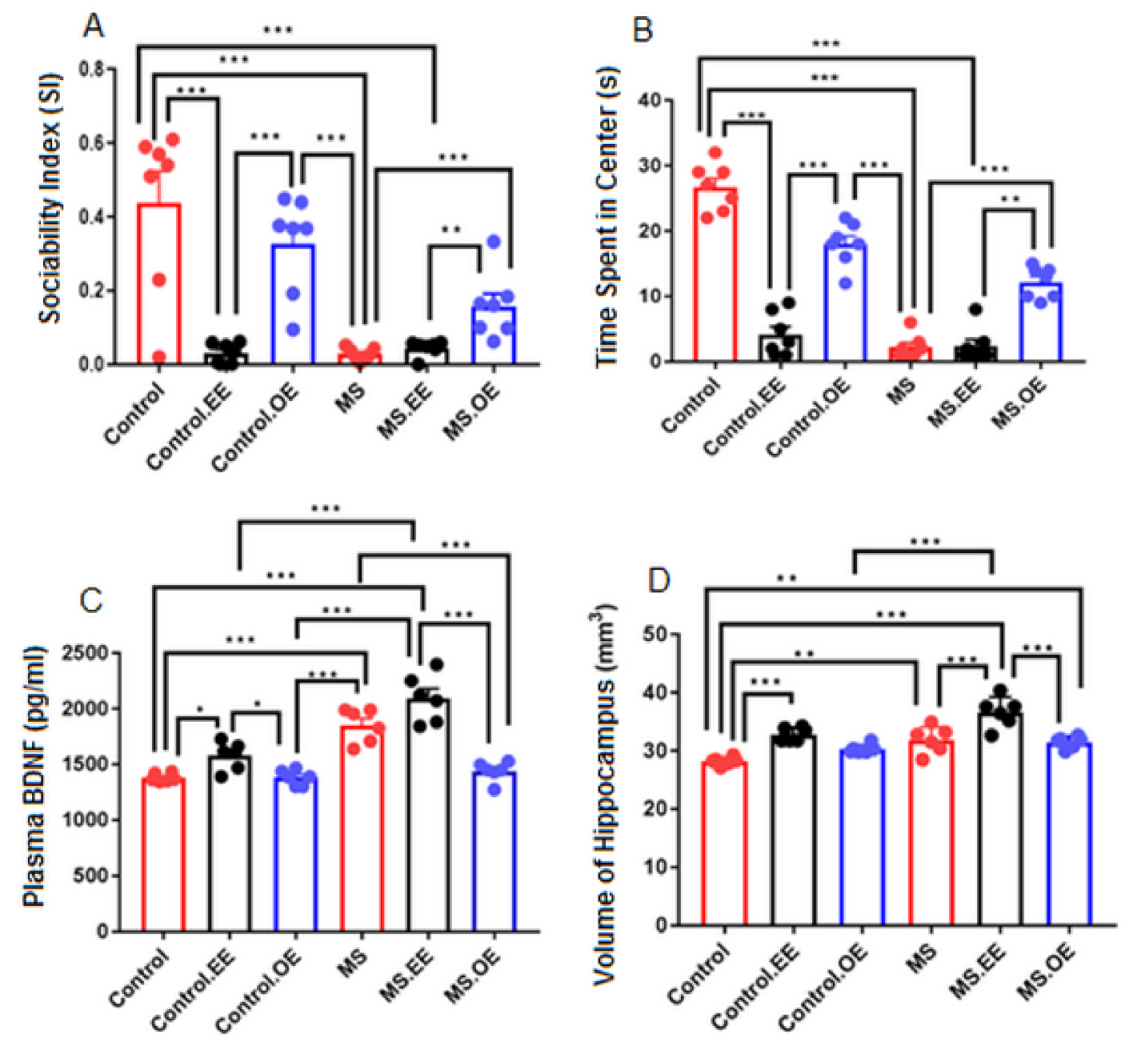
2.6. Results of the Supplementary Experiment (Treatment with both EE and Oxytocin)
3. Discussion
4. Material and Methods
4.1. Animals and Maternal Separation
4.2. Environmental Enrichment
4.3. Supplementary Experiment
4.4. Behavioral Tests
4.4.1. Three-Chamber Test
4.4.2. Open Field Test
4.4.3. Marble Burying Test
4.4.4. Tissue Preparation
4.4.5. Plasma BDNF Level Assay
4.5. Three-D Quantitative Measurements of the Brain
Volume Estimation
4.6. Estimation of the Pyramidal Neuron Number
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cirulli, F. Role of environmental factors on brain development and nerve growth factor expression. Physiol. Behav. 2001, 73, 321–330. [Google Scholar] [CrossRef]
- Hair, N.L.; Hanson, J.L.; Wolfe, B.L.; Pollak, S.D. Association of Child Poverty, Brain Development, and Academic Achievement. JAMA Pediatrics 2015, 169, 822–829. [Google Scholar] [CrossRef] [PubMed]
- Carr, C.P.; Martins, C.M.; Stingel, A.M.; Lemgruber, V.B.; Juruena, M.F. The role of early life stress in adult psychiatric disorders: A systematic review according to childhood trauma subtypes. J. Nerv. Ment. Dis. 2013, 201, 1007–1020. [Google Scholar] [CrossRef] [PubMed]
- Loman, M.M.; Gunnar, M.R. Early experience and the development of stress reactivity and regulation in children. Neurosci. Biobehav. Rev. 2010, 34, 867–876. [Google Scholar] [CrossRef] [PubMed]
- Levine, S. Infantile experience and resistance to physiological stress. Science 1957, 126, 405. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Zhang, C.; Jiang, Y.H.; Brouwer, C.R. Systematic reconstruction of autism biology from massive genetic mutation profiles. Sci. Adv. 2018, 4, e1701799. [Google Scholar] [CrossRef]
- Kleijer, K.T.E.; Huguet, G.; Tastet, J.; Bourgeron, T.; Burbach, J.P.H. Anatomy and Cell Biology of Autism Spectrum Disorder: Lessons from Human Genetics. Adv. Anat. Embryol. Cell Biol. 2017, 224, 1–25. [Google Scholar] [CrossRef]
- Steffenburg, S.; Gillberg, C.; Hellgren, L.; Andersson, L.; Gillberg, I.C.; Jakobsson, G.; Bohman, M. A twin study of autism in Denmark, Finland, Iceland, Norway and Sweden. J. Child Psychol. PsychiatryAllied Discip. 1989, 30, 405–416. [Google Scholar] [CrossRef]
- Hallmayer, J.; Cleveland, S.; Torres, A.; Phillips, J.; Cohen, B.; Torigoe, T.; Miller, J.; Fedele, A.; Collins, J.; Smith, K.; et al. Genetic heritability and shared environmental factors among twin pairs with autism. Arch. Gen. Psychiatry 2011, 68, 1095–1102. [Google Scholar] [CrossRef]
- Sadeghi, S.; Pouretemad, H.; Khosrowabadi, R.; Fathabadi, J.; Nikbakht, S. Behavioral and electrophysiological evidence for parent training in young children with autism symptoms and excessive screen-time. Asian J. Psychiatry 2019, 45, 7–12. [Google Scholar] [CrossRef]
- Sayed Javad Javaheri, E.S.; Bigdeli, M.R.; Zibaii, M.I.; Dargahi, L.; Pouretemad, H.R. Optogenetic Stimulation of the Anterior Cingulate Cortex Ameliorates Autistic-Like Behaviors in Rats Induced by Neonatal Isolation, Caudate Putamen as a Site for Alteration. Neuromolecular Med. 2019, 21, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Tan, T.; Wang, W.; Xu, H.; Huang, Z.; Wang, Y.T.; Dong, Z. Low-Frequency rTMS Ameliorates Autistic-Like Behaviors in Rats Induced by Neonatal Isolation Through Regulating the Synaptic GABA Transmission. Front. Cell. Neurosci. 2018, 12, 46. [Google Scholar] [CrossRef] [PubMed]
- Lyall, K.; Schmidt, R.J.; Hertz-Picciotto, I. Maternal lifestyle and environmental risk factors for autism spectrum disorders. Int. J. Epidemiol. 2014, 43, 443–464. [Google Scholar] [CrossRef] [PubMed]
- Meaney, M.J.; Mitchell, J.B.; Aitken, D.H.; Bhatnagar, S.; Bodnoff, S.R.; Iny, L.J.; Sarrieau, A. The effects of neonatal handling on the development of the adrenocortical response to stress: Implications for neuropathology and cognitive deficits in later life. Psychoneuroendocrinology 1991, 16, 85–103. [Google Scholar] [CrossRef]
- Dawson, G. Early behavioral intervention, brain plasticity, and the prevention of Autism Spectrum Disorder. Dev. Psychopathol. 2008, 20, 775–803. [Google Scholar] [CrossRef]
- Hannan, A.J. Environmental enrichment and brain repair: Harnessing the therapeutic effects of cognitive stimulation and physical activity to enhance experience-dependent plasticity. Neuropathol. Appl. Neurobiol. 2014, 40, 13–25. [Google Scholar] [CrossRef]
- Garbugino, L.; Centofante, E.; D'Amato, F.R. Early Social Enrichment Improves Social Motivation and Skills in a Monogenic Mouse Model of Autism, the Oprm1 (-/-) Mouse. Neural Plast. 2016, 2016, 5346161. [Google Scholar] [CrossRef]
- Kerr, B.; Silva, P.A.; Walz, K.; Young, J.I. Unconventional transcriptional response to environmental enrichment in a mouse model of Rett syndrome. PloS ONE 2010, 5, e11534. [Google Scholar] [CrossRef]
- Reynolds, S.; Urruela, M.; Devine, D.P. Effects of environmental enrichment on repetitive behaviors in the BTBR T+tf/J mouse model of autism. Autism Res. Off. J. Int. Soc. Autism Res. 2013, 6, 337–343. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Hara, Y.; Ago, Y.; Takano, E.; Hasebe, S.; Nakazawa, T.; Hashimoto, H.; Matsuda, T.; Takuma, K. Environmental enrichment attenuates behavioral abnormalities in valproic acid-exposed autism model mice. Behav. Brain Res. 2017, 333, 67–73. [Google Scholar] [CrossRef]
- Favre, M.R.; La Mendola, D.; Meystre, J.; Christodoulou, D.; Cochrane, M.J.; Markram, H.; Markram, K. Predictable enriched environment prevents development of hyper-emotionality in the VPA rat model of autism. Front. Neurosci. 2015, 9, 127. [Google Scholar] [CrossRef] [PubMed]
- Hulbert, S.W.; Bey, A.L.; Jiang, Y.H. Environmental enrichment has minimal effects on behavior in the Shank3 complete knockout model of autism spectrum disorder. Brain Behav. 2018, 8, e01107. [Google Scholar] [CrossRef] [PubMed]
- Lacaria, M.; Spencer, C.; Gu, W.; Paylor, R.; Lupski, J.R. Enriched rearing improves behavioral responses of an animal model for CNV-based autistic-like traits. Hum. Mol. Genet. 2012, 21, 3083–3096. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, M.; Pouretemad, H.; Roghani, M.; Wegener, G.; Ardalan, M. Autistic-like behaviours and associated brain structural plasticity are modulated by oxytocin in maternally separated rats. Behav. Brain Res. 2020, 393, 112756. [Google Scholar] [CrossRef] [PubMed]
- Businaro, R.; Corsi, M.; Azzara, G.; Di Raimo, T.; Laviola, G.; Romano, E.; Ricci, L.; Maccarrone, M.; Aronica, E.; Fuso, A.; et al. Interleukin-18 modulation in autism spectrum disorders. J. Neuroinflammation 2016, 13, 2. [Google Scholar] [CrossRef]
- Meng, F.T.; Zhao, J.; Ni, R.J.; Fang, H.; Zhang, L.F.; Zhang, Z.; Liu, Y.J. Beneficial effects of enriched environment on behaviors were correlated with decreased estrogen and increased BDNF in the hippocampus of male mice. Neuro Endocrinol. Lett. 2015, 36, 490–497. [Google Scholar]
- Caldeira, M.V.; Melo, C.V.; Pereira, D.B.; Carvalho, R.F.; Carvalho, A.L.; Duarte, C.B. BDNF regulates the expression and traffic of NMDA receptors in cultured hippocampal neurons. Mol. Cell. Neurosci. 2007, 35, 208–219. [Google Scholar] [CrossRef]
- Caldeira, M.V.; Melo, C.V.; Pereira, D.B.; Carvalho, R.; Correia, S.S.; Backos, D.S.; Carvalho, A.L.; Esteban, J.A.; Duarte, C.B. Brain-derived neurotrophic factor regulates the expression and synaptic delivery of alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid receptor subunits in hippocampal neurons. J. Biol. Chem. 2007, 282, 12619–12628. [Google Scholar] [CrossRef]
- Ninan, I. Oxytocin suppresses basal glutamatergic transmission but facilitates activity-dependent synaptic potentiation in the medial prefrontal cortex. J. Neurochem. 2011, 119, 324–331. [Google Scholar] [CrossRef]
- Niwa, M.; Jaaro-Peled, H.; Tankou, S.; Seshadri, S.; Hikida, T.; Matsumoto, Y.; Cascella, N.G.; Kano, S.; Ozaki, N.; Nabeshima, T.; et al. Adolescent stress-induced epigenetic control of dopaminergic neurons via glucocorticoids. Science 2013, 339, 335–339. [Google Scholar] [CrossRef]
- Crawley, J.N. Designing mouse behavioral tasks relevant to autistic-like behaviors. Ment. Retard. Dev. Disabil. Res. Rev. 2004, 10, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Pasciuto, E.; Borrie, S.C.; Kanellopoulos, A.K.; Santos, A.R.; Cappuyns, E.; D’Andrea, L.; Pacini, L.; Bagni, C. Autism Spectrum Disorders: Translating human deficits into mouse behavior. Neurobiol. Learn. Mem. 2015, 124, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Ardalan, M.; Rafati, A.H.; Nyengaard, J.R.; Wegener, G. Rapid antidepressant effect of ketamine correlates with astroglial plasticity in the hippocampus. Br. J. Pharm. 2017, 174, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Anzabi, M.; Ardalan, M.; Iversen, N.K.; Rafati, A.H.; Hansen, B.; Ostergaard, L. Hippocampal Atrophy Following Subarachnoid Hemorrhage Correlates with Disruption of Astrocyte Morphology and Capillary Coverage by AQP4. Front. Cell Neurosci. 2018, 12, 19. [Google Scholar] [CrossRef] [PubMed]
- Ardalan, M.; Wegener, G.; Polsinelli, B.; Madsen, T.M.; Nyengaard, J.R. Neurovascular plasticity of the hippocampus one week after a single dose of ketamine in genetic rat model of depression. Hippocampus 2016, 26, 1414–1423. [Google Scholar] [CrossRef]
- Christensen, J.; Gronborg, T.K.; Sorensen, M.J.; Schendel, D.; Parner, E.T.; Pedersen, L.H.; Vestergaard, M. Prenatal valproate exposure and risk of autism spectrum disorders and childhood autism. JAMA 2013, 309, 1696–1703. [Google Scholar] [CrossRef]
- Kirsten, T.B.; Chaves-Kirsten, G.P.; Chaible, L.M.; Silva, A.C.; Martins, D.O.; Britto, L.R.; Dagli, M.L.; Torrao, A.S.; Palermo-Neto, J.; Bernardi, M.M. Hypoactivity of the central dopaminergic system and autistic-like behavior induced by a single early prenatal exposure to lipopolysaccharide. J. Neurosci. Res. 2012, 90, 1903–1912. [Google Scholar] [CrossRef]
- Meaney, M.J.; Szyf, M. Maternal care as a model for experience-dependent chromatin plasticity? Trends Neurosci. 2005, 28, 456–463. [Google Scholar] [CrossRef]
- Weaver, I.C.; Champagne, F.A.; Brown, S.E.; Dymov, S.; Sharma, S.; Meaney, M.J.; Szyf, M. Reversal of maternal programming of stress responses in adult offspring through methyl supplementation: Altering epigenetic marking later in life. J. Neurosci. Off. J. Soc. Neurosci. 2005, 25, 11045–11054. [Google Scholar] [CrossRef]
- Nagasawa, M.; Okabe, S.; Mogi, K.; Kikusui, T. Oxytocin and mutual communication in mother-infant bonding. Front. Hum. Neurosci. 2012, 6, 31. [Google Scholar] [CrossRef]
- Wu, X.; Bai, Y.; Tan, T.; Li, H.; Xia, S.; Chang, X.; Zhou, Z.; Zhou, W.; Li, T.; Wang, Y.T.; et al. Lithium ameliorates autistic-like behaviors induced by neonatal isolation in rats. Front. Behav. Neurosci. 2014, 8, 234. [Google Scholar] [CrossRef] [PubMed]
- Alleva, E.; Francia, N. Psychiatric vulnerability: Suggestions from animal models and role of neurotrophins. Neurosci. Biobehav. Rev. 2009, 33, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Cirulli, F.; Francia, N.; Branchi, I.; Antonucci, M.T.; Aloe, L.; Suomi, S.J.; Alleva, E. Changes in plasma levels of BDNF and NGF reveal a gender-selective vulnerability to early adversity in rhesus macaques. Psychoneuroendocrinology 2009, 34, 172–180. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cirulli, F.; Francia, N.; Berry, A.; Aloe, L.; Alleva, E.; Suomi, S.J. Early life stress as a risk factor for mental health: Role of neurotrophins from rodents to non-human primates. Neurosci. Biobehav. Rev. 2009, 33, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Klein, A.B.; Williamson, R.; Santini, M.A.; Clemmensen, C.; Ettrup, A.; Rios, M.; Knudsen, G.M.; Aznar, S. Blood BDNF concentrations reflect brain-tissue BDNF levels across species. Int. J. Neuropsychopharmacol. 2011, 14, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.-Y.; Feng, J.-C.; Cao, C.; Wu, H.-T.; Loh, Y.P.; Cheng, Y. Association of Peripheral Blood Levels of Brain-Derived Neurotrophic Factor With Autism Spectrum Disorder in Children: A Systematic Review and Meta-analysis. JAMA Pediatrics 2016, 170, 1079–1086. [Google Scholar] [CrossRef]
- Enstrom, A.M.; Lit, L.; Onore, C.E.; Gregg, J.P.; Hansen, R.L.; Pessah, I.N.; Hertz-Picciotto, I.; Van de Water, J.A.; Sharp, F.R.; Ashwood, P. Altered gene expression and function of peripheral blood natural killer cells in children with autism. Brain Behav. Immun. 2009, 23, 124–133. [Google Scholar] [CrossRef]
- Binder, D.K.; Scharfman, H.E. Brain-derived neurotrophic factor. Growth Factors 2004, 22, 123–131. [Google Scholar] [CrossRef]
- Gabriel, P.; Mastracchio, T.A.; Bordner, K.; Jeffrey, R. Impact of enriched environment during adolescence on adult social behavior, hippocampal synaptic density and dopamine D2 receptor expression in rats. Physiol. Behav. 2020, 226, 113133. [Google Scholar] [CrossRef]
- Moreira, F.P.; Fabiao, J.D.; Bittencourt, G.; Wiener, C.D.; Jansen, K.; Oses, J.P.; Quevedo Lde, A.; Souza, L.D.; Crispim, D.; Portela, L.V.; et al. The Met allele of BDNF Val66Met polymorphism is associated with increased BDNF levels in generalized anxiety disorder. Psychiatr. Genet. 2015, 25, 201–207. [Google Scholar] [CrossRef]
- Zeev, B.; Vecsler, M. The common BDNF polymorphism may be a modifier of disease severity in Rett syndrome (vol 72, pg 1242, 2009). Neurology 2009, 73, 161. [Google Scholar] [CrossRef]
- Nithianantharajah, J.; Hannan, A.J. Enriched environments, experience-dependent plasticity and disorders of the nervous system. Nat. Rev.. Neurosci. 2006, 7, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Olsson, I.A.; Dahlborn, K. Improving housing conditions for laboratory mice: A review of "environmental enrichment". Lab. Anim. 2002, 36, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Duffy, S.N.; Craddock, K.J.; Abel, T.; Nguyen, P.V. Environmental enrichment modifies the PKA-dependence of hippocampal LTP and improves hippocampus-dependent memory. Learn. Mem. (Cold Spring Harb. N.Y.) 2001, 8, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Fox, C.; Merali, Z.; Harrison, C. Therapeutic and protective effect of environmental enrichment against psychogenic and neurogenic stress. Behav. Brain Res. 2006, 175, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Nithianantharajah, J.; Barkus, C.; Murphy, M.; Hannan, A.J. Gene-environment interactions modulating cognitive function and molecular correlates of synaptic plasticity in Huntington's disease transgenic mice. Neurobiol. Dis. 2008, 29, 490–504. [Google Scholar] [CrossRef] [PubMed]
- Pang, T.Y.; Du, X.; Zajac, M.S.; Howard, M.L.; Hannan, A.J. Altered serotonin receptor expression is associated with depression-related behavior in the R6/1 transgenic mouse model of Huntington's disease. Hum. Mol. Genet. 2009, 18, 753–766. [Google Scholar] [CrossRef]
- Bechard, A.R.; Cacodcar, N.; King, M.A.; Lewis, M.H. How does environmental enrichment reduce repetitive motor behaviors? Neuronal activation and dendritic morphology in the indirect basal ganglia pathway of a mouse model. Behav. Brain Res. 2016, 299, 122–131. [Google Scholar] [CrossRef]
- Ronzoni, G.; Antón, M.; Mora, F.; Segovia, G.; Del Arco, A. Infralimbic cortex controls the activity of the hypothalamus–pituitary–adrenal axis and the formation of aversive memory: Effects of environmental enrichment. Behav. Brain Res. Sreetestcontent1 2016, 297, 338–344. [Google Scholar] [CrossRef]
- Zajac, M.S.; Pang, T.Y.; Wong, N.; Weinrich, B.; Leang, L.S.; Craig, J.M.; Saffery, R.; Hannan, A.J. Wheel running and environmental enrichment differentially modify exon-specific BDNF expression in the hippocampus of wild-type and pre-motor symptomatic male and female Huntington's disease mice. Hippocampus 2010, 20, 621–636. [Google Scholar] [CrossRef]
- Prasetyo, S.; Yolanda, S.; Ivanali, K. Effect of continuous environmental enrichment and aerobic exercise on rat plasma and hippocampal brain-derived neurotrophic factor. J. Nat. Sci. Biol. Med. 2019, 10, 20–23. [Google Scholar] [CrossRef]
- Hu, Y.S.; Long, N.; Pigino, G.; Brady, S.T.; Lazarov, O. Molecular mechanisms of environmental enrichment: Impairments in Akt/GSK3beta, neurotrophin-3 and CREB signaling. PloS ONE 2013, 8, e64460. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mansouri, M.; Pouretemad, H.; Wegener, G.; Roghani, M.; Afshari, M.; Mallard, C.; Ardalan, M. Dual Profile of Environmental Enrichment and Autistic-Like Behaviors in the Maternal Separated Model in Rats. Int. J. Mol. Sci. 2021, 22, 1173. https://doi.org/10.3390/ijms22031173
Mansouri M, Pouretemad H, Wegener G, Roghani M, Afshari M, Mallard C, Ardalan M. Dual Profile of Environmental Enrichment and Autistic-Like Behaviors in the Maternal Separated Model in Rats. International Journal of Molecular Sciences. 2021; 22(3):1173. https://doi.org/10.3390/ijms22031173
Chicago/Turabian StyleMansouri, Monireh, Hamidreza Pouretemad, Gregers Wegener, Mehrdad Roghani, Masoud Afshari, Carina Mallard, and Maryam Ardalan. 2021. "Dual Profile of Environmental Enrichment and Autistic-Like Behaviors in the Maternal Separated Model in Rats" International Journal of Molecular Sciences 22, no. 3: 1173. https://doi.org/10.3390/ijms22031173
APA StyleMansouri, M., Pouretemad, H., Wegener, G., Roghani, M., Afshari, M., Mallard, C., & Ardalan, M. (2021). Dual Profile of Environmental Enrichment and Autistic-Like Behaviors in the Maternal Separated Model in Rats. International Journal of Molecular Sciences, 22(3), 1173. https://doi.org/10.3390/ijms22031173