
Magnesium Signaling in Plants



Abstract
1. Introduction
2. AK and NDPK in Cell Energetics
3. AK-Mediated Adenylate Equilibrium and Mg2+ Signaling
4. Magnesium and the Adenylate Energy Charge Theory
5. Magnesium Status in Cells
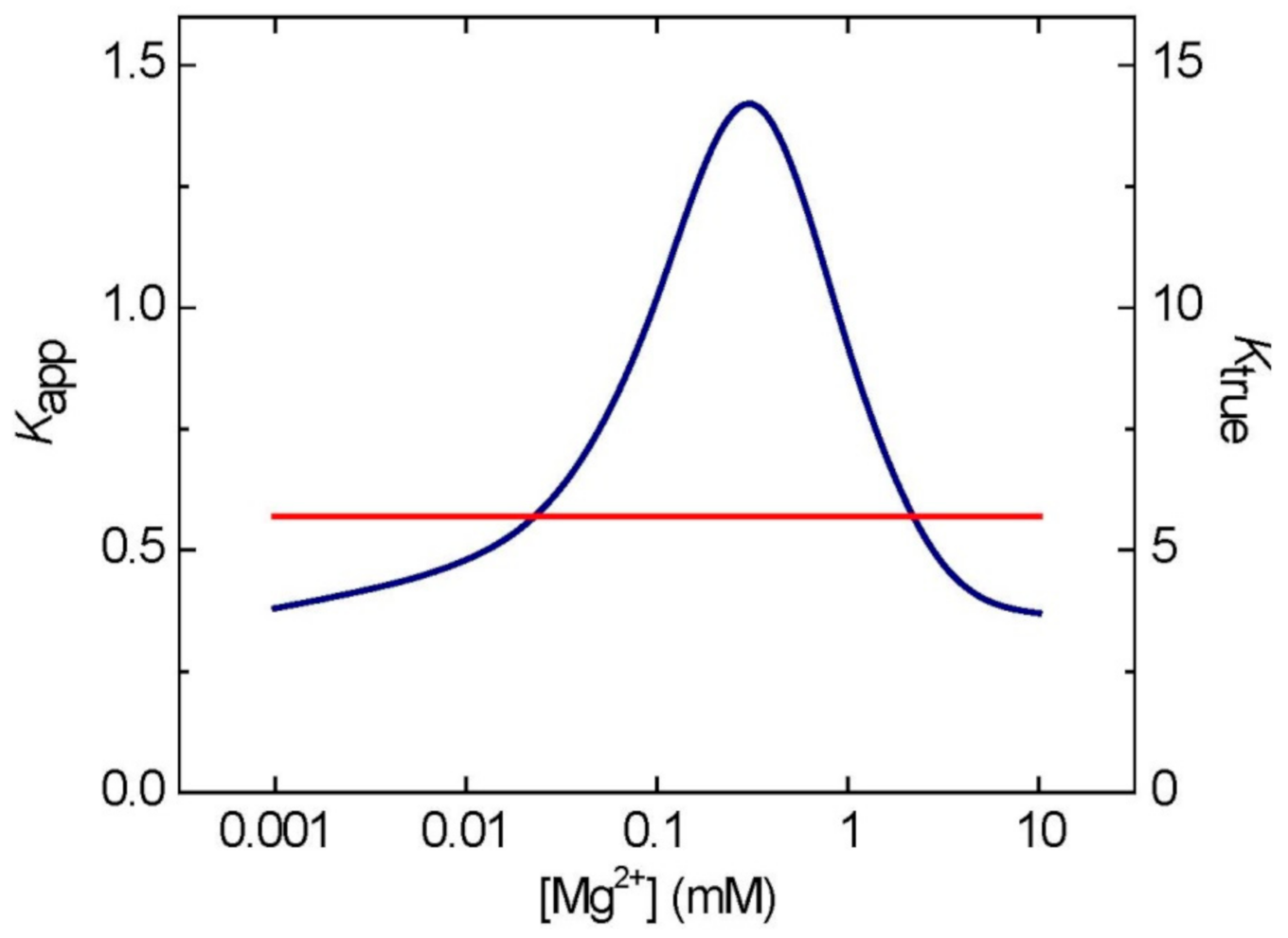
6. Feasibility of the Estimations of [Mg2+] Based on Adenylate Measurements
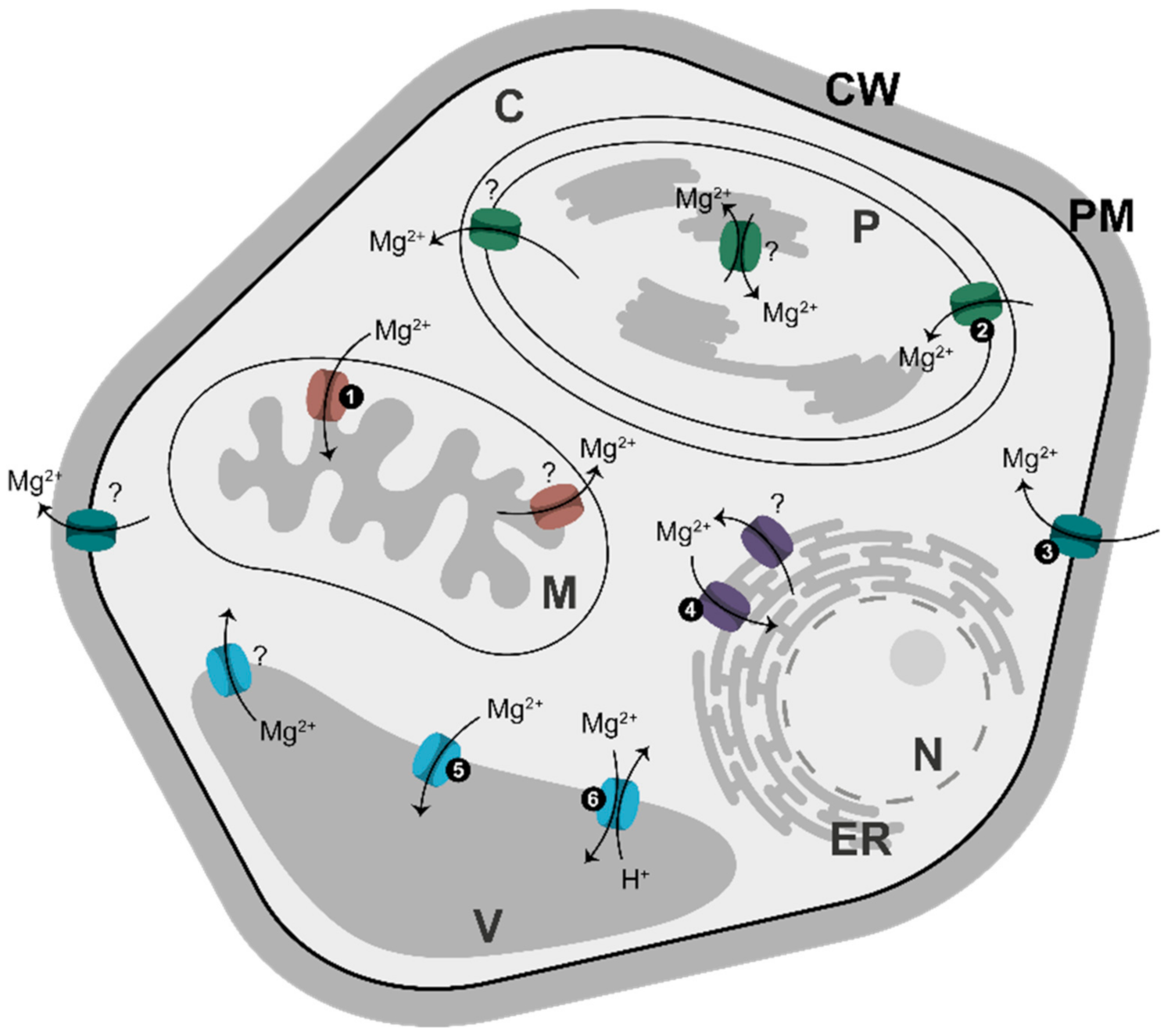
7. Magnesium Translocators
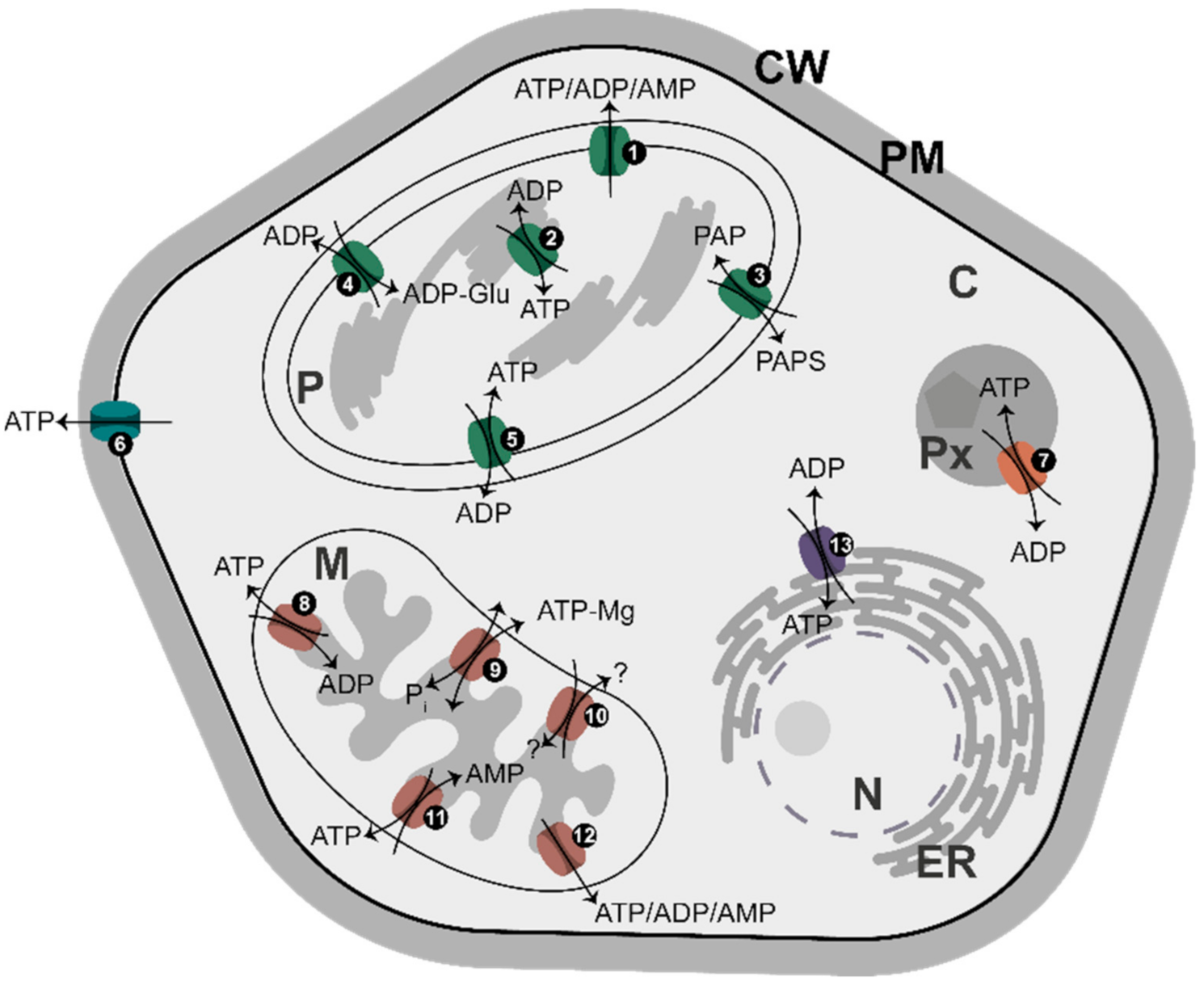
8. Adenylate Translocators
9. Role of [Mg2+] in Metabolism and Signaling
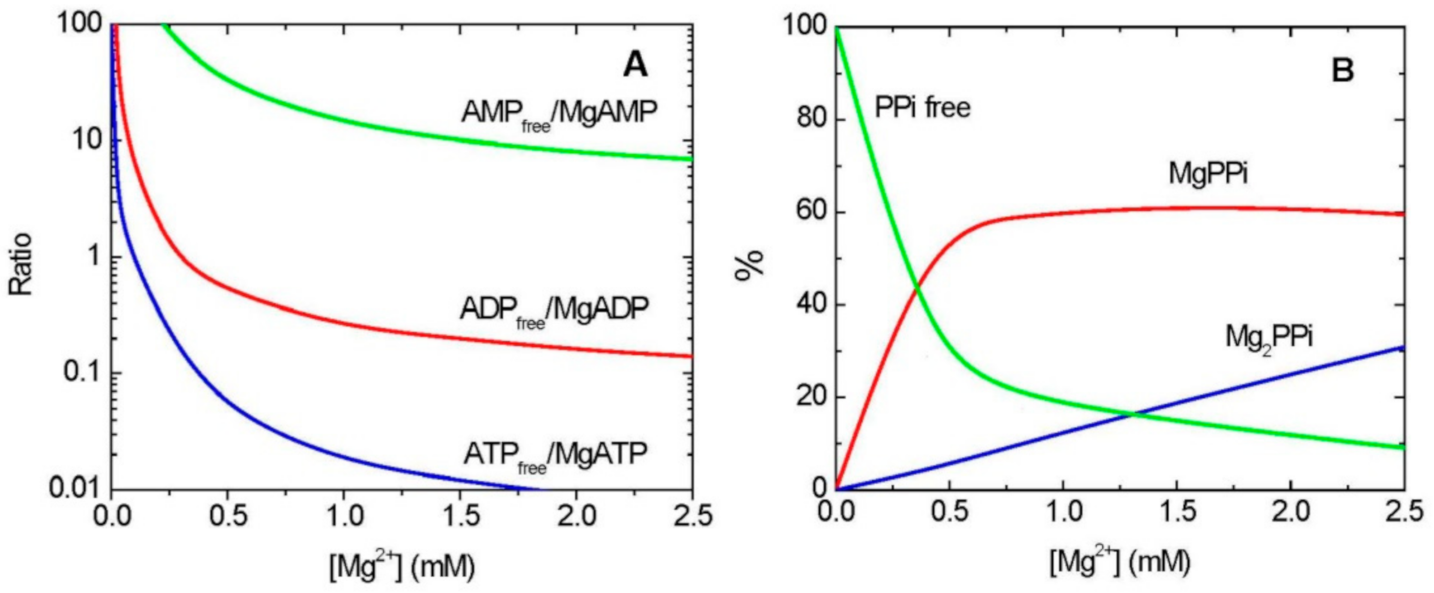
9.1. Mg2+ and Chelation of Adenylates and PPi
9.2. Mg2+ as a Regulator of Enzymatic Activities
9.3. Mg2+ Regulates Energy Metabolism, Adenylate Transport, and Allosteric Regulation
9.4. [Mg2+] under Anoxia
9.5. Magnesium versus Calcium, Sodium and Aluminum
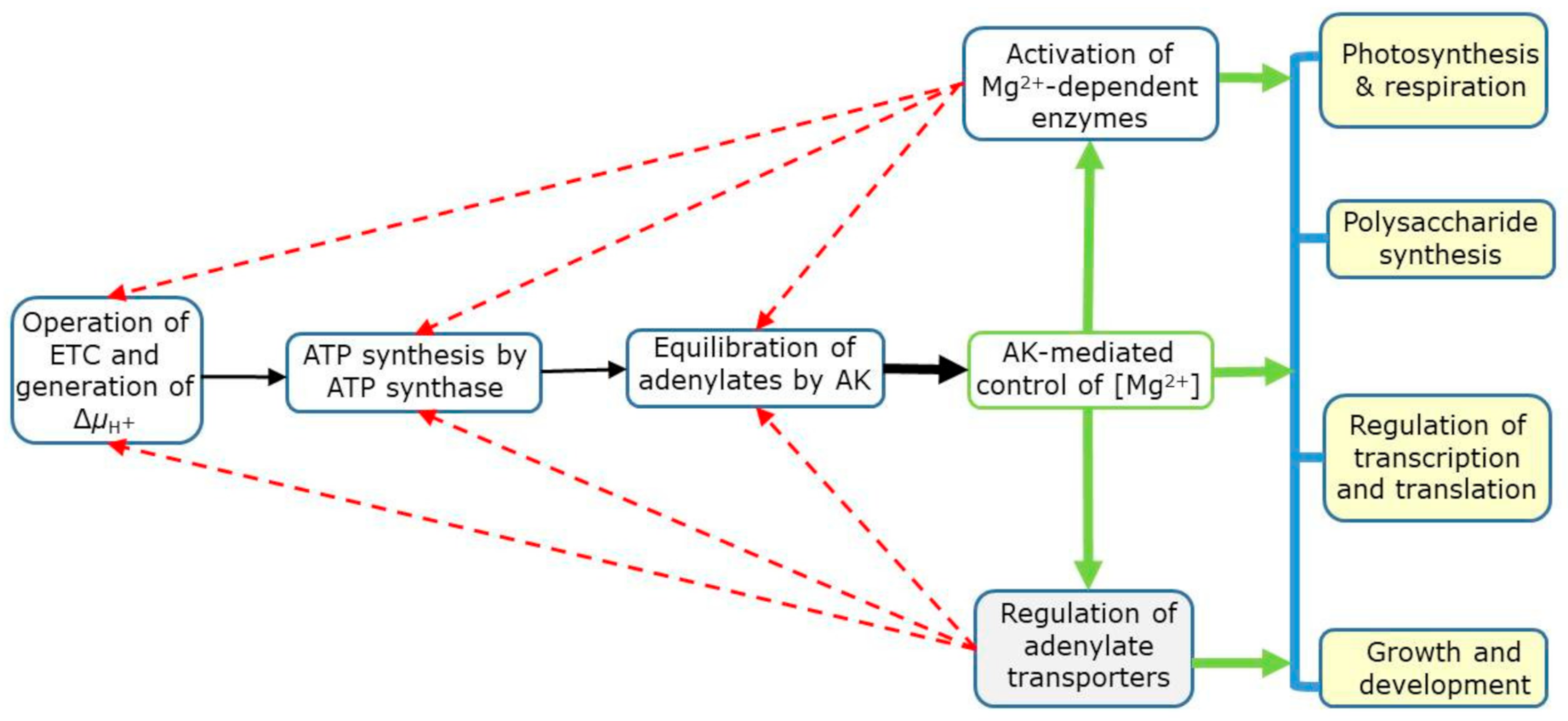
9.6. Summary of Mg2+ Signaling in Plants
10. Does AK Control Mg2+ Signaling in Other Organisms?
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AEC | Adenylate energy charge |
| AK | Adenylate kinase |
| AMPK | AMP-activated protein kinase |
| CBL | Calcineurin B-like protein |
| CIPK | CBL-interacting protein kinase |
| ER | Endoplasmic reticulum |
| IMS | Intermembrane space |
| Kapp | Apparent equilibrium constant |
| Ktrue | True equilibrium constant |
| MGT | Magnesium translocator |
| NDP | Nucleoside diphosphate |
| NMP | Nucleoside monophosphate |
| NTP | Nucleoside triphosphate |
| NDPK | Nucleoside diphosphate kinase |
| Pi | Inorganic phosphate |
| PPase | Pyrophosphatase |
| PPi | Inorganic pyrophosphate |
| SuSy | Sucrose synthase |
| UGPase | UDP-glucose pyrophosphorylase |
References
- Kobayashi, N.I.; Tanoi, K. Critical issues in the study of magnesium transport systems and magnesium deficiency symptoms in plants. Int. J. Mol. Sci. 2015, 16, 23076–23093. [Google Scholar] [CrossRef]
- Hermans, C.; Conn, S.J.; Chen, J.; Xiao, Q.; Verbruggen, N. An update on magnesium homeostasis mechanisms in plants. Metallomics 2013, 5, 1170–1183. [Google Scholar] [CrossRef] [PubMed]
- Dorenstouter, H.; Pieters, G.; Findenegg, G. Distribution of magnesium between chlorophyll and other photosynthetic functions in magnesium deficient “sun”; and “shade”; leaves of poplar. J. Plant Nutr. 1985, 8, 1089–1101. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets—Iron, zinc, cop-per, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.H.; Marschner, H. The mineral nutrition of higher plants. J. Ecol. 1988, 76, 1250. [Google Scholar] [CrossRef]
- Cakmak, I.; Hengeler, C.; Marschner, H. Partitioning of shoot and root dry matter and carbohydrates in bean plants suffering from phosphorus, potassium and magnesium deficiency. J. Exp. Bot. 1994, 45, 1245–1250. [Google Scholar] [CrossRef]
- Hermans, C.; Bourgis, F.; Faucher, M.; Strasser, R.J.; Velrot, S.; Verbruggen, N. Magnesium deficiency in sugar beets alters sugar partitioning and phloem loading in young mature leaves. Planta 2004, 220, 541–549. [Google Scholar] [CrossRef]
- Zhang, B.; Cakmak, I.; Feng, J.; Yu, C.; Chen, X.; Xie, D.; Wu, L.; Song, Z.; Cao, J.; He, Y. Magnesium deficiency reduced the yield and seed germination in wax gourd by affecting the carbohydrate translocation. Front. Plant Sci. 2020, 11, 797. [Google Scholar] [CrossRef]
- Tanoi, K.; Kobayashi, N.I. Leaf senescence by magnesium deficiency. Plants 2015, 4, 756–772. [Google Scholar] [CrossRef]
- Bruggemann, L.I.; Pottosin, I.I.; Schönknecht, G. Cytoplasmic magnesium regulates the fast activating vacuolar cation chan-nel. J. Exp. Bot. 1999, 50, 1547–1552. [Google Scholar] [CrossRef]
- Shaul, O. Magnesium transport and function in plants: The tip of the iceberg. Biometals 2002, 15, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Igamberdiev, A.U.; Kleczkowski, L.A. Optimization of ATP synthase function in mitochondria and chloroplasts via the adenylate kinase equilibrium. Front. Plant Sci. 2015, 6, 10. [Google Scholar] [CrossRef] [PubMed]
- Voon, C.P.; Guan, X.; Sun, Y.; Sahu, A.; Chan, M.N.; Gardeström, P.; Wagner, S.; Fuchs, P.; Nietzel, T.; Versaw, W.K.; et al. ATP compartmentation in plastids and cytosol of Arabidopsis thalianarevealed by fluorescent protein sensing. Proc. Natl. Acad. Sci. USA 2018, 115, E10778–E10787. [Google Scholar] [CrossRef] [PubMed]
- Igamberdiev, A.U.; Kleczkowski, L.A. Thermodynamic buffering, stable non-equilibrium and establishment of the computable structure of plant metabolism. Prog. Biophys. Mol. Biol. 2019, 146, 23–36. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, W.J.; Smithers, G.W. Stability constants for biologically important metal-ligand complexes. Methods Enzymol. 1979, 63, 294–336. [Google Scholar] [CrossRef]
- Purich, D.L.; Fromm, H.J. Studies on factors influencing enzyme responses to adenylate energy charge. J. Biol. Chem. 1972, 247, 249–255. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Kleczkowski, L.A. Membrane potential, adenylate levels and Mg2+ are interconnected via adenylate kinase equilibrium in plant cells. Biochim. Biophys. Acta (BBA) Bioenergy 2003, 1607, 111–119. [Google Scholar] [CrossRef]
- Li, F.Y.; Chaigne-Delalande, B.; Kanellopoulou, C. Second messenger role for Mg2+ revealed by human T-cell immunodeficien-cy. Nature 2011, 475, 471–476. [Google Scholar] [CrossRef]
- Atkinson, D.E. Energy charge of the adenylate pool as a regulatory parameter. Interaction with feedback modifiers. Biochemistry 1968, 7, 4030–4034. [Google Scholar] [CrossRef]
- Halford, N.G.; Paul, M.J. Carbon metabolite sensing and signalling. Plant Biotechnol. J. 2003, 1, 381–398. [Google Scholar] [CrossRef]
- Dzeja, P.P.; Terzic, A. Adenylate kinase and AMP signaling networks: Metabolic monitoring, signal communication and body energy sensing. Int. J. Mol. Sci. 2009, 10, 1729–1772. [Google Scholar] [CrossRef] [PubMed]
- Schlattner, U.; Wagner, E.; Greppin, H.; Bonzon, M. Chloroplast adenylate kinase from tobacco. Purification and partial characterization. Phytochemistry 1996, 42, 589–594. [Google Scholar] [CrossRef]
- Schlattner, U.; Wagner, E.; Greppin, H.; Bonzon, M. Changes in distribution of chloroplast adenylate kinase isoforms during floral induction. Physiol. Plant. 1996, 96, 319–323. [Google Scholar] [CrossRef]
- Carrari, F.; Coll-Garcia, D.; Schauer, N.; Lytovchenko, A.; Palacios-Rojas, N.; Balbo, I.; Rosso, M.; Fernie, A.R. Deficiency of a plastidial adenylate kinase in arabidopsis results in elevated photosynthetic amino acid biosynthesis and enhanced growth. Plant Physiol. 2004, 137, 70–82. [Google Scholar] [CrossRef]
- Lange, P.R.; Geserick, C.; Tischendorf, G.; Zrenner, R. Functions of chloroplastic adenylate kinases in Arabidopsis. Plant Physiol. 2007, 146, 492–504. [Google Scholar] [CrossRef]
- Feng, X.; Yang, R.; Zheng, X.; Zhang, F. Identification of a novel nuclear-localized adenylate kinase 6 from Arabidopsis thaliana as an essential stem growth factor. Plant Physiol. Biochem. 2012, 61, 180–186. [Google Scholar] [CrossRef]
- Deppert, W.R.; Wagner, E. Purification of adenylate kinase from green leaves of barley, maize and Chenopodium rubrum L. J. Plant Physiol. 1995, 145, 17–23. [Google Scholar] [CrossRef]
- Lee, C.P.; Taylor, N.L.; Millar, A.H. Recent Advances in the Composition and Heterogeneity of the Arabidopsis Mitochondrial Proteome. Front. Plant Sci. 2013, 4, 4. [Google Scholar] [CrossRef]
- Noma, T.; Fujisawa, K.; Yamashiro, Y.; Shinohara, M.; Nakazawa, A.; Gondo, T.; Ishihara, T.; Yoshinobu, K. Structure and expression of human mitochondrial adenylate kinase targeted to the mitochondrial matrix. Biochem. J. 2001, 358, 225–232. [Google Scholar] [CrossRef]
- Salvato, F.; Havelund, J.F.; Chen, M.; Rao, R.S.P.; Rogowska-Wrzesinska, A.; Jensen, O.N.; Gang, D.R.; Thelen, J.J.; Møller, I.M. The potato tuber mitochondrial proteome. Plant Physiol. 2013, 164, 637–653. [Google Scholar] [CrossRef]
- Slovak, R.; Setzer, C.; Roiuk, M.; Bertels, J.; Göschl, C.; Jandrasits, K.; Beemster, G.T.; Busch, W. Ribosome assembly factor Adenylate Kinase 6 maintains cell proliferation and cell size homeostasis during root growth. New. Phytol. 2019, 225, 2064–2076. [Google Scholar] [CrossRef] [PubMed]
- Kleczkowski, L.A.; Randall, D.D. Maize leaf adenylate kinase: Purification and partial characterization. Plant Physiol. 1986, 81, 1110–1114. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hamada, M.; Sumida, M.; Kurokawa, Y.; Sunayashiki-Kusuzaki, K.; Okuda, H.; Watanabe, T.; Kuby, S.A. Studies on the adenylate kinase isozymes from the serum and erythrocyte of normal and Duchenne dystrophic patients. Isolation, physicochemical properties, and several comparisons with the Duchenne dystrophic aberrant enzyme. J. Biol. Chem. 1985, 260, 11595–11602. [Google Scholar] [CrossRef]
- Zhang, Y.; Launay, H.; Liu, F.; Lebrun, R.; Gontero, B. Interaction between adenylate kinase 3 and glyceraldehyde-3-phosphate dehydrogenase from Chlamydomonas reinhardtii. FEBS J. 2018, 285, 2495–2503. [Google Scholar] [CrossRef]
- Peña, C.; Hurt, E.; Panse, V.G. Eukaryotic ribosome assembly, transport and quality control. Nat. Struct. Mol. Biol. 2017, 24, 689–699. [Google Scholar] [CrossRef]
- Schlattner, U.; Wagner, E.; Greppin, H.; Bonzon, M. Binding of adenylate kinase to RNA. Biochem. Biophys. Res. Commun. 1995, 217, 509–514. [Google Scholar] [CrossRef]
- Raveneau, M.-P.; Benamar, A.; Macherel, D. Water content, adenylate kinase, and mitochondria drive adenylate balance in dehydrating and imbibing seeds. J. Exp. Bot. 2017, 68, 3501–3512. [Google Scholar] [CrossRef]
- Regierer, B.; Fernie, A.R.; Springer, F.; Perez-Melis, A.; Leisse, A.; Koehl, K.; Willmitzer, L.; Geigenberger, P.; Kossmann, J. Starch content and yield increase as a result of altering adenylate pools in transgenic plants. Nat. Biotechnol. 2002, 20, 1256–1260. [Google Scholar] [CrossRef]
- Kleczkowski, L.A. Is leaf ADP-glucose pyrophosphorylase an allosteric enzyme? Biochim. Biophys. Acta (BBA) Protein Struct. Mol. Enzym. 2000, 1476, 103–108. [Google Scholar] [CrossRef]
- Zala, D.; Schlattner, U.; Desvignes, T.; Bobe, J.; Roux, A.; Chavrier, P.; Boissan, M. The advantage of channeling nucleotides for very processive functions. F1000Research 2017, 6, 724. [Google Scholar] [CrossRef] [PubMed]
- Dorion, S.; Rivoal, E. Clues to the functions of plant NDPK isoforms. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2015, 388, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Kleczkowski, L.A.; Igamberdiev, A.U. Optimization of nucleotide sugar supply for polysaccharide formation via thermodynamic buffering. Biochem. J. 2020, 477, 341–356. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Inouye, M. Adenylate kinase complements nucleoside diphosphate kinase deficiency in nucleotide metabolism. Proc. Natl. Acad. Sci. USA 1996, 93, 5720–5725. [Google Scholar] [CrossRef] [PubMed]
- Kleczkowski, L. Back to the drawing board: Redefining starch synthesis in cereals. Trends Plant Sci. 1996, 1, 363–364. [Google Scholar] [CrossRef]
- Roberts, J.; Aubert, S.; Gout, E.; Bligny, R.; Douce, R. Cooperation and competition between adenylate kinase, nucleoside diphosphokinase, electron transport, and ATP synthase in plant mitochondria studied by 31P-nuclear magnetic resonance. Plant Physiol. 1997, 113, 191–199. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Johansson, M.; Hammargren, J.; Uppsäll, E.; MacKenzie, A.; Knorpp, C. The activities of nucleoside diphosphate kinase and adenylate kinase are influenced by their interaction. Plant Sci. 2008, 174, 192–199. [Google Scholar] [CrossRef]
- Valenti, D.; Vacca, R.A.; Romero-Puertas, M.D.C.; De Gara, L.; Marra, E.; Passarella, S. In the early phase of programmed cell death in Tobacco Bright Yellow 2 cells the mitochondrial adenine nucleotide translocator, adenylate kinase and nucleoside diphosphate kinase are impaired in a reactive oxygen species-dependent manner. Biochim. Biophys. Acta (BBA) Gen. Subj. 2007, 1767, 66–78. [Google Scholar] [CrossRef]
- Stucki, J.W. The thermodynamic-buffer enzymes. Eur. J. Biochem. 1980, 109, 257–267. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Kleczkowski, L.A. Metabolic systems maintain stable non-equilibrium via thermodynamic buffering. BioEssays 2009, 31, 1091–1099. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Kleczkowski, L.A. Magnesium and cell energetics in plants under anoxia. Biochem. J. 2011, 437, 373–379. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Kleczkowski, L.A. Optimization of CO2 fixation in photosynthetic cells via thermodynamic buffering. Biosystems 2011, 103, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Dorion, S.; Clendenning, A.; Rivoal, J. Engineering the expression level of cytosolic nucleoside diphosphate kinase in transgenic Solanum tuberosum roots alters growth, respiration and carbon metabolism. Plant J. 2017, 89, 914–926. [Google Scholar] [CrossRef] [PubMed]
- Rose, I.A. The state of magnesium in cells as estimated from the adenylate kinase equilibrium. Proc. Natl. Acad. Sci. USA 1968, 61, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Kleczkowski, L.A.; Randall, D.D. Equilibration of adenylates by maize leaf adenylate kinase—Effects of magnesium on appar-ent and true equilibria. J. Exp. Bot. 1991, 42, 537–540. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Kleczkowski, L.A. Implications of adenylate kinase-governed equilibrium of adenylates on contents of free magnesium in plant cells and compartments. Biochem. J. 2001, 360, 225–231. [Google Scholar] [CrossRef]
- Blair, J.M. Magnesium, potassium, and the adenylate kinase equilibrium. Magnesium as a feedback signal from the adenine nucleotide pool. Eur. J. Biochem. 1970, 13, 384–390. [Google Scholar] [CrossRef]
- Kholodenko, N.Y.; Kartashov, I.M.; Makarov, A.D. Some kinetic characteristics of chloroplast adenylate kinase. Biochemistry (Moscow) 1983, 48, 411–416. [Google Scholar]
- Zhou, L.; Lacroute, F.; Thornburg, R. Cloning, expression in Escherichia coli, and characterization of Arabidopsis thaliana UMP/CMP kinase. Plant Physiol. 1998, 117, 245–254. [Google Scholar] [CrossRef]
- Purich, D.L.; Fromm, H.J. Additional factors influencing enzyme responses to the adenylate energy charge. J. Biol. Chem. 1973, 248, 461–466. [Google Scholar] [CrossRef]
- Pradet, A.; Raymond, P. Adenine nucleotide ratios and adenylate energy charge in energy metabolism. Annu. Rev. Plant Physiol. 1983, 34, 199–224. [Google Scholar] [CrossRef]
- Waters, B.M. Moving magnesium in plant cells. New Phytol. 2011, 190, 510–513. [Google Scholar] [CrossRef] [PubMed]
- Igamberdiev, A.U.; Kleczkowski, L.A. Equilibration of adenylates in the mitochondrial intermembrane space maintains respi-ration and regulates cytosolic metabolism. J. Exp. Bot. 2006, 57, 2133–2141. [Google Scholar] [CrossRef] [PubMed]
- Bligny, R.; Gout, E. Regulation of respiration by cellular key parameters: Energy demand, ADP, and Mg2+. In Plant Respiration: Metabolic Fluxes and Carbon Balance. Advances in Photosynthesis and Respiration (Including Bioenergy and Related Processes); Springer: Cham, The Netherland, 2017; Volume 43, pp. 19–41. [Google Scholar]
- Yazaki, Y.; Asukagawa, N.; Ishikawa, Y.; Ohta, E.; Sakata, M. Estimation of cytoplasmic free Mg2+ levels and phosphorylation potentials in mung bean root tips by in vivo 31P NMR spectroscopy. Plant Cell Physiol. 1988, 29, 919–924. [Google Scholar] [CrossRef]
- Portis, A.R. Evidence of a low stromal Mg2+ concentration in intact chloroplasts in the dark. Plant Physiol. 1981, 67, 985–989. [Google Scholar] [CrossRef]
- Ishijima, S.; Uchibori, A.; Takagi, H.; Maki, R.; Ohnishi, M. Light-induced increase in free Mg2+ concentration in spinach chlo-roplasts: Measurement of free Mg2+ by using a fluorescent probe and necessity of stromal alkalinization. Arch. Biochem. Biophys. 2003, 412, 126–132. [Google Scholar] [CrossRef]
- Gout, E.; Rébeillé, F.; Douce, R.; Bligny, R. Interplay of Mg2+, ADP, and ATP in the cytosol and mitochondria: Unravelling the role of Mg2+ in cell respiration. Proc. Natl. Acad. Sci. USA 2014, 111, E4560–E4567. [Google Scholar] [CrossRef]
- Bose, J.; Babourina, O.; Shabala, S.; Rengel, Z. Low-pH and aluminum resistance in arabidopsis correlates with high cytosolic magnesium content and increased magnesium uptake by plant roots. Plant Cell Physiol. 2013, 54, 1093–1104. [Google Scholar] [CrossRef]
- Conn, S.J.; Conn, V.; Kaiser, B.N.; Leigh, R.A.; Tyerman, S.; Gilliham, M. Magnesium transporters, MGT2/MRS2-1 and MGT3/MRS2-5, are important for magnesium partitioning within Arabidopsis thaliana mesophyll vacuoles. New Phytol. 2011, 190, 583–594. [Google Scholar] [CrossRef]
- Koch, M.; Winkelmann, M.K.; Hasler, M.; Pawelzik, E.; Naumann, M. Root growth in light of changing magnesium distribution and transport between source and sink tissues in potato (Solanum tuberosum L.). Sci. Rep. 2020, 10, 8796. [Google Scholar] [CrossRef]
- Romani, A. Cellular magnesium homeostasis. Arch. Biochem. Biophys. 2011, 512, 1–23. [Google Scholar] [CrossRef]
- Vishnu, N.; Jadoon, K.M.; Karsten, F.; Groschner, L.N.; Waldeck-Weiermair, M.; Rost, R.; Hallström, S.; Imamura, H.; Graier, W.F.; Malli, R. ATP increases within the lumen of the endoplasmic reticulum upon intracellular Ca2+ release. Mol. Biol. Cell 2014, 25, 368–379. [Google Scholar] [CrossRef] [PubMed]
- DePaoli, M.R.; Hay, J.C.; Graier, W.F.; Malli, R. The enigmatic ATP supply of the endoplasmic reticulum. Biol. Rev. 2018, 94, 610–628. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, H.; Iwasawa, N.; Citterio, D.; Suzuki, Y.; Kubota, T.; Tokuno, K.; Kitamura, Y.; Oka, A.K.; Suzuki, K. Design and Synthesis of highly sensitive and selective fluorescein-derived magnesium fluorescent probes and application to intracellular 3D Mg2+ imaging. J. Am. Chem. Soc. 2004, 126, 16353–16360. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Yu, X.; Li, M.; Liao, N.; Bi, A.; Jiang, Y.; Liu, S.; Gong, Z.; Wenbin, Z. Fluorescent probes for the detection of magnesium ions (Mg2+): From design to application. RSC Adv. 2018, 8, 12573–12587. [Google Scholar] [CrossRef]
- Gupta, R.; Benovic, J.; Rose, Z. The determination of the free magnesium level in the human red blood cell by 31P NMR. J. Biol. Chem. 1978, 253, 6172–6176. [Google Scholar] [CrossRef]
- Romani, A.; Scarpa, A. Regulation of cell magnesium. Arch. Biochem. Biophys. 1992, 298, 1–12. [Google Scholar] [CrossRef]
- Packer, L.; Douce, R. Plant cell membranes. In Methods in Enzymology; Academic Press: London, UK, 1987; Volume 148. [Google Scholar]
- Farré, E.M.; Tiessen, A.; Roessner, U.; Geigenberger, P.; Trethewey, R.N.; Willmitzer, L. Analysis of the compartmentation of glycolytic intermediates, nucleotides, sugars, organic acids, amino acids, and sugar alcohols in potato tubers using a nonaqueous fractionation method. Plant Physiol. 2001, 127, 685–700. [Google Scholar] [CrossRef] [PubMed]
- Nieman, R.H.; Clark, R.A.; Pap, D.; Ogata, G.; Maas, E.V. Effects of salt stress on adenine and uridine nucleotide pools, sugar and acid-soluble phosphate in shoots of pepper and safflower. J. Exp. Bot. 1988, 39, 301–309. [Google Scholar] [CrossRef]
- Díaz-Troya, S.; Roldán, M.; Mallén-Ponce, M.J.; Ortega-Martínez, P.; Florencio, F.J. Lethality caused by ADP-glucose accu-mulation is suppressed by salt-induced carbon flux redirection in cyanobacteria. J. Exp. Bot. 2020, 71, 2005–2017. [Google Scholar] [CrossRef]
- Tang, R.J.; Luan, S. Regulation of calcium and magnesium homeostasis in plants: From transporters to signaling network. Curr. Opin. Plant Biol. 2017, 39, 97–105. [Google Scholar] [CrossRef]
- Chen, Z.C.; Peng, W.T.; Li, J.; Liao, H. Functional dissection and transport mechanism of magnesium in plants. Semin. Cell Dev. Biol. 2018, 74, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Tutone, A.F.; Drummond, R.S.; Gardner, R.C.; Luan, S. A novel family of magnesium transport genes in Arabidopsis. Plant Cell 2001, 13, 2761–2775. [Google Scholar] [CrossRef] [PubMed]
- Schock, I.; Gregan, J.; Steinhauser, S.; Schweyen, R.; Brennicke, A.; Knoop, V. A member of a novel Arabidopsis thaliana gene family of candidate Mg2+ ion transporters complements a yeastmitochondrial group II intron-splicing mutant. Plant J. 2000, 24, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Li, L.-G.; Sokolov, L.N.; Yang, Y.-H.; Li, D.-P.; Ting, J.; Pandy, G.K.; Luan, S. A mitochondrial magnesium transporter functions in arabidopsis pollen development. Mol. Plant 2008, 1, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Kobayashi, N.I.; Tanoi, K.; Iwata, N.; Suzuki, H.; Iwata, R.; Nakanishi, T.M. Expression and functional analysis of the CorA-MRS2-ALR-type magnesium transporter family in rice. Plant Cell Physiol. 2013, 54, 1673–1683. [Google Scholar] [CrossRef]
- Shabala, S.; Hariadi, Y. Effects of magnesium availability on the activity of plasma membrane ion transporters and light-induced responses from broad bean leaf mesophyll. Planta 2005, 221, 56–65. [Google Scholar] [CrossRef]
- Walker, R.B.; Walker, H.M.; Ashworth, P.R. Calcium-magnesium nutrition with special reference to serpentine soils. Plant Physiol. 1955, 30, 214–221. [Google Scholar] [CrossRef]
- Tang, R.-J.; Zhao, F.-G.; Garcia, V.J.; Kleist, T.J.; Yang, L.; Zhang, H.-X.; Luan, S. Tonoplast CBL–CIPK calcium signaling network regulates magnesium homeostasis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2015, 112, 3134–3139. [Google Scholar] [CrossRef]
- Gao, C.; Zhao, Q.; Jiang, L. Vacuoles protect plants from high magnesium stress. Proc. Natl. Acad. Sci. USA 2015, 112, 2931–2932. [Google Scholar] [CrossRef]
- Ma, X.; Li, Q.-H.; Yu, Y.-N.; Qiao, Y.-M.; Haq, S.U.; Gong, Z.-H. The CBL–CIPK pathway in plant response to stress signals. Int. J. Mol. Sci. 2020, 21, 5668. [Google Scholar] [CrossRef]
- Yan, Y.-W.; Mao, D.-D.; Yang, L.; Qi, J.-L.; Zhang, X.-X.; Tang, Q.-L.; Li, Y.-P.; Tang, R.; Luan, S. Magnesium transporter MGT6 plays an essential role in maintaining magnesium homeostasis and regulating high magnesium tolerance in Arabidopsis. Front. Plant Sci. 2018, 9, 274. [Google Scholar] [CrossRef] [PubMed]
- Mao, D.; Chen, J.; Tian, L.; Liu, Z.; Yang, L.; Tang, R.; Li, J.; Lu, C.; Yang, Y.; Shi, J.; et al. Arabidopsis transporter MGT6 mediates magnesium uptake and is required for growth under magnesium limitation. Plant Cell 2014, 26, 2234–2248. [Google Scholar] [CrossRef] [PubMed]
- Shaul, O.; Hilgemann, D.W.; De-Almeida-Engler, J.; Van Montagu, M.; Inzé, D.; Galili, G. Cloning and characterization of a novel Mg2+/H+ exchanger. EMBO J. 1999, 18, 3973–3980. [Google Scholar] [CrossRef] [PubMed]
- Gaash, R.; Elazar, M.; Mizrahi, K.; Avramov-Mor, M.; Berezin, I.; Shaul, O. Phylogeny and a structural model of plant MHX transporters. BMC Plant Biol. 2013, 13, 75. [Google Scholar] [CrossRef] [PubMed]
- David-Assael, O.; Mizrachy-Dagri, T.; Berezin, I.; Brook, E.; Shaul, O.; Saul, H. Expression of AtMHX, an Arabidopsis vacuolar metal transporter, is repressed by the 5’ untranslated region of its gene. J. Exp. Bot. 2005, 56, 1039–1047. [Google Scholar] [CrossRef]
- Vreugdenhil, D.; Aarts, M.G.M.; Koorneef, M. Exploring natural genetic variation to improve plant nutrient content. In Plant Nutritional Genomics; Broadley, M.R., White, P.J., Eds.; Blackwell: Oxford, UK, 2005; pp. 201–219. [Google Scholar]
- Da Fonseca-Pereira, P.; Neri-Silva, R.; Cavalcanti, J.H.F.; Brito, D.S.; Weber, A.P.M.; Araújo, W.L.; Nunes-Nesi, A. Da-ta-mining bioinformatics: Connecting adenylate transport and metabolic responses to stress. Trends Plant Sci. 2018, 23, 961–974. [Google Scholar] [CrossRef] [PubMed]
- Kramer, R. Influence of divalent cations on the reconstituted ADP, ATP exchange. Biochim. Biophys. Acta (BBA) Bioenergy 1980, 592, 615–620. [Google Scholar] [CrossRef]
- Trentmann, O.; Mühlhaus, T.; Zimmer, D.; Sommer, F.; Schroda, M.; Haferkamp, I.; Keller, I.; Pommerrenig, B.; Neuhaus, H.E. Identification of chloroplast envelope proteins with critical importance for cold acclimation. Plant Physiol. 2020, 182, 1239–1255. [Google Scholar] [CrossRef]
- Schlattner, U.; Wagner, E. The adenylate kinase family in plants: Isozyme activity is related to flower induction. Endocytobiosis Cell Res. 2001, 14, 67–73. [Google Scholar]
- Scopes, R.K. Binding of substrates and other anions to yeast phosphoglycerate kinase. Eur. J. Biochem. 1978, 91, 119–129. [Google Scholar] [CrossRef]
- Renz, A.; Stitt, M. Substrate specificity and product inhibition of different forms of fructokinases and hexokinases in devel-oping potato tubers. Planta 1993, 190, 166–175. [Google Scholar] [CrossRef]
- Monasterio, O.; Cárdenas, M.L. Kinetic studies of rat liver hexokinase D (glucokinase) in non-co-operative conditions show an ordered mechanism with MgADP as the last product to be released. Biochem. J. 2003, 371, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Xu, L.; Xu, M.; Wan, B.; Yu, L.; Huang, Q. Role of Mg2+ ions in protein kinase phosphorylation: Insights from molecu-lar dynamics simulations of ATP-kinase complexes. Mol. Simul. 2011, 37, 1143–1150. [Google Scholar] [CrossRef]
- Stitt, M. Pyrophosphate as an energy donor in the cytosol of plant cells: An enigmatic alternative to ATP. Bot. Acta 1998, 111, 167–175. [Google Scholar] [CrossRef]
- Davies, J.M.; Poole, R.J.; Sanders, D. The computed free energy change of hydrolysis of inorganic pyrophosphate and ATP: Apparent significance. For inorganic-pyrophosphate-driven reactions of intermediary metabolism. Biochim. Biophys. Acta (BBA) Bioenergy 1993, 1141, 29–36. [Google Scholar] [CrossRef]
- Gutiérrez-Luna, F.M.; Hernández-Domínguez, E.-E.; Valencia-Turcotte, L.G.; Rodríguez-Sotres, R. Pyrophosphate and pyrophosphatases in plants, their involvement in stress responses and their possible relationship to secondary metabolism. Plant Sci. 2017, 267, 11–19. [Google Scholar] [CrossRef]
- Horder, M. Complex formation of inorganic pyrophosphate with magnesium –influence of ionic strength, supporting me-dium and temperature. Biochim. Biophys. Acta 1974, 358, 319–328. [Google Scholar] [CrossRef]
- Leigh, R.A.; Pope, A.J.; Jennings, I.R.; Sanders, D. Kinetics of the vacuolar H+-pyrophosphatase: The roles of magnesium, pyrophosphate, and their complexes as substrates, activators, and inhibitors. Plant Physiol. 1992, 100, 1698–1705. [Google Scholar] [CrossRef][Green Version]
- Maeshima, M. Vacuolar H+-pyrophosphatase. Biochim. Biophys. Acta 2000, 1465, 37–51. [Google Scholar] [CrossRef]
- Nilima, K.; Vinay, S. V-PPase in plants: An overview. Res. J. Biotechnol. 2008, 3, 57–63. [Google Scholar]
- Fraichard, A.; Trossat, C.; Perotti, E.; Pugin, A. Allosteric regulation by Mg2+ of the vacuolar H+-PPase from Acer pseudoplatanus cells. Ca2+/Mg2+ interactions. Biochimie 1996, 78, 259–266. [Google Scholar] [CrossRef]
- Airas, R.K. Differences in the magnesium dependences of the class I and class II aminoacyl-tRNA synthetases from Escherichia coli. Eur. J. Biochem. 1996, 240, 223–231. [Google Scholar] [CrossRef]
- Grzechowiak, M.; Ruszkowski, M.; Śliwiak, J.; Szpotkowski, K.; Sikorski, M.; Jaskolski, M. Crystal structures of plant inorganic pyrophosphatase, an enzyme with a moonlighting autoproteolytic activity. Biochem. J. 2019, 476, 2297–2319. [Google Scholar] [CrossRef]
- Pérez-Castiñeira, J.R.; Serrano, A. The H+-Translocating inorganic pyrophosphatase from Arabidopsis thaliana is more sensitive to sodium than its Na+-translocating counterpart from Methanosarcina mazei. Front. Plant Sci. 2020, 11, 1240. [Google Scholar] [CrossRef] [PubMed]
- Ferjani, A.; Kawade, K.; Asaoka, M.; Oikawa, A.; Okada, T.; Mochizuki, A.; Maeshima, M.; Hirai, M.Y.; Saito, K.; Tsukaya, H. Pyrophosphate inhibits gluconeogenesis by restricting UDP-glucose formation in vivo. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Decker, D.; Öberg, C.; Kleczkowski, L.A. Identification and characterization of inhibitors of UDP-glucose and UDP-sugar pyrophosphorylases for in vivo studies. Plant J. 2017, 90, 1093–1107. [Google Scholar] [CrossRef] [PubMed]
- Decker, D.; Kleczkowski, L.A. UDP-sugar producing pyrophosphorylases—Distinct and essential enzymes with overlap-ping substrate specificities, providing de novo precursors for glycosylation reactions. Front. Plant Sci. 2019, 9, 1822. [Google Scholar] [CrossRef] [PubMed]
- Delmer, D.P. The purification and properties of sucrose synthetase from etiolated Phaseolus aureus seedlings. J. Biol. Chem. 1972, 247, 3822–3828. [Google Scholar] [CrossRef]
- Takeda, H.; Niikura, M.; Narumi, A.; Aoki, H.; Sasaki, T.; Shimada, H. Phosphorylation of rice sucrose synthase isoforms promotes the activity of sucrose degradation. Plant Biotechnol. 2017, 34, 107–113. [Google Scholar] [CrossRef]
- Saito, H. Outersphere and innersphere coordinated metal ions in an aminoacyl-tRNA synthetase ribozyme. Nucleic Acids Res. 2002, 30, 5151–5159. [Google Scholar] [CrossRef]
- Villa, T.; Pleiss, J.A.; Guthrie, C. Spliceosomal snRNAs: Mg2+-dependent chemistry at the catalytic core? Cell 2002, 109, 149–152. [Google Scholar] [CrossRef]
- Hartwig, A. Role of magnesium in genomic stability. Mutat. Res. Mol. Mech. Mutagen. 2001, 475, 113–121. [Google Scholar] [CrossRef]
- Svetlov, V.; Nudler, E. Basic mechanism of transcription by RNA polymerase II. Biochim. Biophys. Acta (BBA) Bioenergy 2013, 1829, 20–28. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Misra, V.K.; Draper, D.E. On the role of magnesium ions in RNA stability. Biopolymers 1998, 48, 113–135. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Hurry, V.; Krömer, S.; Gardeström, P. The role of mitochondrial electron transport during photosynthetic induction. A study with barley (Hordeum vulgare) protoplasts incubated with rotenone and oligomycin. Physiol. Plant. 1998, 104, 431–439. [Google Scholar] [CrossRef]
- Santarius, K.A.; Heber, U. Changes in the intracellular levels of ATP, ADP, AMP and Pi and regulatory function of the adenylate system in leaf cells during photosynthesis. Biochim. Biophys. Acta (BBA) Biophys. Incl. Photosynth. 1965, 102, 39–54. [Google Scholar] [CrossRef]
- Hampp, R.; Goller, M.; Ziegler, H. Adenylate levels, energy charge, and phosphorylation potential during dark-light and light-dark transition in chloroplasts, mitochondria, and cytosol of mesophyll protoplasts from Avena sativa L. Plant Physiol. 1982, 69, 448–455. [Google Scholar] [CrossRef]
- Stitt, M.; Lilley, R.M.; Heldt, H.W. Adenine nucleotide levels in the cytosol, chloroplasts, and mitochondria of wheat leaf protoplasts. Plant Physiol. 1982, 70, 971–977. [Google Scholar] [CrossRef]
- Hardie, D.G.; Hawley, S.A. AMP-activated protein kinase: The energy charge hypothesis revisited. BioEssays 2001, 23, 1112–1119. [Google Scholar] [CrossRef]
- Broeckx, T.; Hulsmans, S.; Rolland, F. The plant energy sensor: Evolutionary conservation and divergence of SnRK1 structure, regulation, and function. J. Exp. Bot. 2016, 67, 6215–6252. [Google Scholar] [CrossRef]
- Piattoni, C.V.; Bustos, D.M.; Guerrero, S.A.; Iglesias, A.A. Nonphosphorylating glyceraldehyde-3-phosphate dehydrogenase is phosphorylated in wheat endosperm at serine-404 by an SNF1-related Protein kinase allosterically inhibited by ribose-5-phosphate. Plant Physiol. 2011, 156, 1337–1350. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Y.; Zhu, L.; Li, Y.; Guo, M.; Liu, Y.; Cheng, J.; Zhao, J.; Wu, Y. Architectural plasticity of AMPK revealed by electron microscopy and X-ray crystallography. Sci. Rep. 2016, 6, 24191. [Google Scholar] [CrossRef]
- Xiao, B.; Sanders, M.J.; Underwood, E.; Heath, R.J.; Mayer, F.V.; Carmena, D.; Jing, C.; Walker, P.A.; Eccleston, J.F.; Haire, L.F.; et al. Structure of mammalian AMPK and its regulation by ADP. Nat. Cell Biol. 2011, 472, 230–233. [Google Scholar] [CrossRef]
- Lasanthi-Kudahettige, R.; Magneschi, L.; Loreti, E.; Gonzali, S.; Licausi, F.; Novi, G.; Beretta, O.; Vitulli, F.; Alpi, A.; Perata, P. Transcript profiling of the anoxic rice coleoptile. Plant Physiol. 2007, 144, 218–231. [Google Scholar] [CrossRef]
- Lambeck, I.C.; Fischer-Schrader, K.; Niks, D.; Roeper, J.; Chi, J.-C.; Hille, R.; Schwarz, G. Molecular Mechanism of 14-3-3 Protein-mediated Inhibition of Plant Nitrate Reductase. J. Biol. Chem. 2012, 287, 4562–4571. [Google Scholar] [CrossRef] [PubMed]
- Gilli, R.; Lafitte, D.; Lopez, C.; Kilhoffer, M.-C.; Makarov, A.; Briand, A.C.; Haiech, J. Thermodynamic analysis of calcium and magnesium binding to calmodulin. Biochemistry 1998, 37, 5450–5456. [Google Scholar] [CrossRef] [PubMed]
- Igamberdiev, A.U.; Hill, R.D. Elevation of cytosolic Ca2+ in response to energy deficiency in plants: The general mechanism of adaptation to low oxygen stress. Biochem. J. 2018, 475, 1411–1425. [Google Scholar] [CrossRef]
- Sillen, L.G.; Martell, A.E. Stability constants of metal ion complexes. J. Chem. Educ. 1964, 42, 521. [Google Scholar]
- Vincent, A.; Blair, J. The coupling of the adenylate kinase and creatine kinase equilibria. Calculation of substrate and feedback signal levels in muscle. FEBS Lett. 1970, 7, 239–244. [Google Scholar] [CrossRef]
- Malmendal, A.; Linse, S.; Evenäs, J.; Forsén, S.; Drakenberg, T. Battle for the EF-Hands: Magnesium−Calcium Interference in Calmodulin. Biochemistry 1999, 38, 11844–11850. [Google Scholar] [CrossRef]
- Banti, V.; Giuntoli, B.; Gonzali, S.; Loreti, E.; Magneschi, L.; Novi, G.; Paparelli, E.; Parlanti, S.; Pucciariello, C.; Santaniello, A.; et al. Low Oxygen Response Mechanisms in Green Organisms. Int. J. Mol. Sci. 2013, 14, 4734–4761. [Google Scholar] [CrossRef] [PubMed]
- Møller, I.M.; Rasmusson, A.G. The role of NADP in the mitochondrial matrix. Trends Plant Sci. 1998, 3, 21–27. [Google Scholar] [CrossRef]
- Zielinski, R.E. Calmodulin and calmodulin-binding proteins in plants. Annu. Rev. Plant Biol. 1998, 49, 697–725. [Google Scholar] [CrossRef] [PubMed]
- Lemtiri-Chlieh, F.; Arold, S.T.; Gehring, C.A. Mg2+ is a missing link in plant cell Ca2+ signalling and homeostasis—A study on vicia faba guard cells. Int. J. Mol. Sci. 2020, 21, 3771. [Google Scholar] [CrossRef] [PubMed]
- Serrano, R.; Mulet, J.M.; Rios, G.; Marquez, J.A.; De Larrinoa, I.I.F.; Leube, M.P.; Mendizabal, I.; Pascual-Ahuir, A.; Proft, M.; Ros, R.; et al. A glimpse of the mechanisms of ion homeostasis during salt stress. J. Exp. Bot. 1999, 50, 1023–1036. [Google Scholar] [CrossRef]
- Chen, Z.C.; Yamaji, N.; Horie, T.; Che, J.; Li, J.; An, G.; Ma, J.F. A magnesium transporter OsMGT1 plays a critical role in salt tolerance in rice. Plant Physiol. 2017, 174, 1837–1849. [Google Scholar] [CrossRef]
- Jacoby, R.P.; Millar, A.H.; Taylor, N.L. Investigating the role of respiration in plant salinity tolerance by analyzing mitochondrial proteomes from wheat and a salinity-tolerant amphiploid (Wheat × Lophopyrum elongatum). J. Proteome Res. 2013, 12, 4807–4829. [Google Scholar] [CrossRef]
- Chen, Z.C.; Yamaji, N.; Motoyama, R.; Nagamura, Y.; Ma, J.F. Up-regulation of a magnesium transporter gene OsMGT1 is required for conferring aluminum tolerance in rice. Plant Physiol. 2012, 159, 1624–1633. [Google Scholar] [CrossRef]
- Moon, S.; Kim, J.; Bae, E. Structural analyses of adenylate kinases from Antarctic and tropical fishes for understanding cold adaptation of enzymes. Sci. Rep. 2017, 7, 16027. [Google Scholar] [CrossRef]
- Barbour, R.L.; Ribaudo, J.; Chan, S.H. Effect of creatine kinase activity on mitochondrial ADP/ATP transport. Evidence for a functional interaction. J. Biol. Chem. 1984, 259, 8246–8251. [Google Scholar] [CrossRef]
- Golding, E.M.; Teague, W.E.; Dobson, G.P. Adjustment of K’ to varying pH and pMg for the creatine kinase, adenylate kinase and ATP hydrolysis equilibria permitting quantitative bioenergetic assessment. J. Exp. Biol. 1995, 198, 1775–1782. [Google Scholar] [PubMed]
- Williams, R.J. Chemical advances in evolution by and changes in use of space during time. J. Theor. Biol. 2011, 268, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Sakano, K. Revision of biochemical pH-stat: Involvement of alternative pathway metabolisms. Plant Cell Physiol. 1998, 39, 467–473. [Google Scholar] [CrossRef]
- Kunz, S.; Gardeström, P.; Pesquet, E.; Kleczkowski, L.A. Hexokinase 1 is required for glucose-induced repression of bZIP63, At5g22920, and BT2 in Arabidopsis. Front. Plant Sci. 2015, 6, 525. [Google Scholar] [CrossRef] [PubMed]
- Ciereszko, I. Regulatory roles of sugars in plant growth and development. Acta Soc. Bot. Pol. 2018, 87, 3583. [Google Scholar] [CrossRef]
- Freire, M.Á. Phosphorylation and acylation transfer reactions: Clues to a dual origin of metabolism. Biosystems 2020, 198, 104260. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AK Name and Gene | Location | Function | Reference |
|---|---|---|---|
| AK1 (At2g37250) | M (a), P (b) | control of growth | [24,25,30] |
| AK2 (At5g47840) | P | plastid development | [24,25] |
| AK3 (At5g50370) | M (c), C (d) | unknown | [25,28] |
| AK4 (At5g63400) | M | unknown | [28] |
| AK5 (At5g35170) | P | no phenotype for the knockout | [25] |
| AK6 (At5g60340) | N | control of stem growth; | [26] |
| control of root growth; ribosome maturation | [31] | ||
| AK7 (At3g01820) | M | unknown | [25] |
| AK8 (e) (At2g39270) | M | unknown | [25] |
| Compartment | [Mg2+], mM | Method | Reference |
|---|---|---|---|
| Cytosol | 0.25 | 31P-NMR | [67] |
| 0.40 | 31P-NMR | [64] | |
| 0.9 | Ionophore | [68] | |
| 0.2–0.4 | From Kapp of AK | [55,62] | |
| Mitochondria | 2.4 | 31P-NMR | [67] |
| 1.0–3.0 | From Kapp of AK | [55] | |
| Chloroplasts | 0.5–2.0 | Ionophore | [66] |
| 1.0–3.0 | Ionophore | [65] | |
| 0.2–5.0 | From Kapp of AK | [55] | |
| Vacuole | 5–80 (a) | X-ray analysis | [2,69] |
| ER lumen | Unknown | ||
| Peroxisomes | Unknown |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kleczkowski, L.A.; Igamberdiev, A.U. Magnesium Signaling in Plants. Int. J. Mol. Sci. 2021, 22, 1159. https://doi.org/10.3390/ijms22031159
Kleczkowski LA, Igamberdiev AU. Magnesium Signaling in Plants. International Journal of Molecular Sciences. 2021; 22(3):1159. https://doi.org/10.3390/ijms22031159
Chicago/Turabian StyleKleczkowski, Leszek A., and Abir U. Igamberdiev. 2021. "Magnesium Signaling in Plants" International Journal of Molecular Sciences 22, no. 3: 1159. https://doi.org/10.3390/ijms22031159
APA StyleKleczkowski, L. A., & Igamberdiev, A. U. (2021). Magnesium Signaling in Plants. International Journal of Molecular Sciences, 22(3), 1159. https://doi.org/10.3390/ijms22031159