
Noncanonical Sequences Involving NHERF1 Interaction with NPT2A Govern Hormone-Regulated Phosphate Transport: Binding Outside the Box


Abstract
1. Introduction
2. NHERF1 (PDZ1) Specifically Binds NPT2A for Hormone-Sensitive Phosphate Transport
2.1. NHERF1 PDZ1 and PDZ2 Domains Are Not Interchangeable
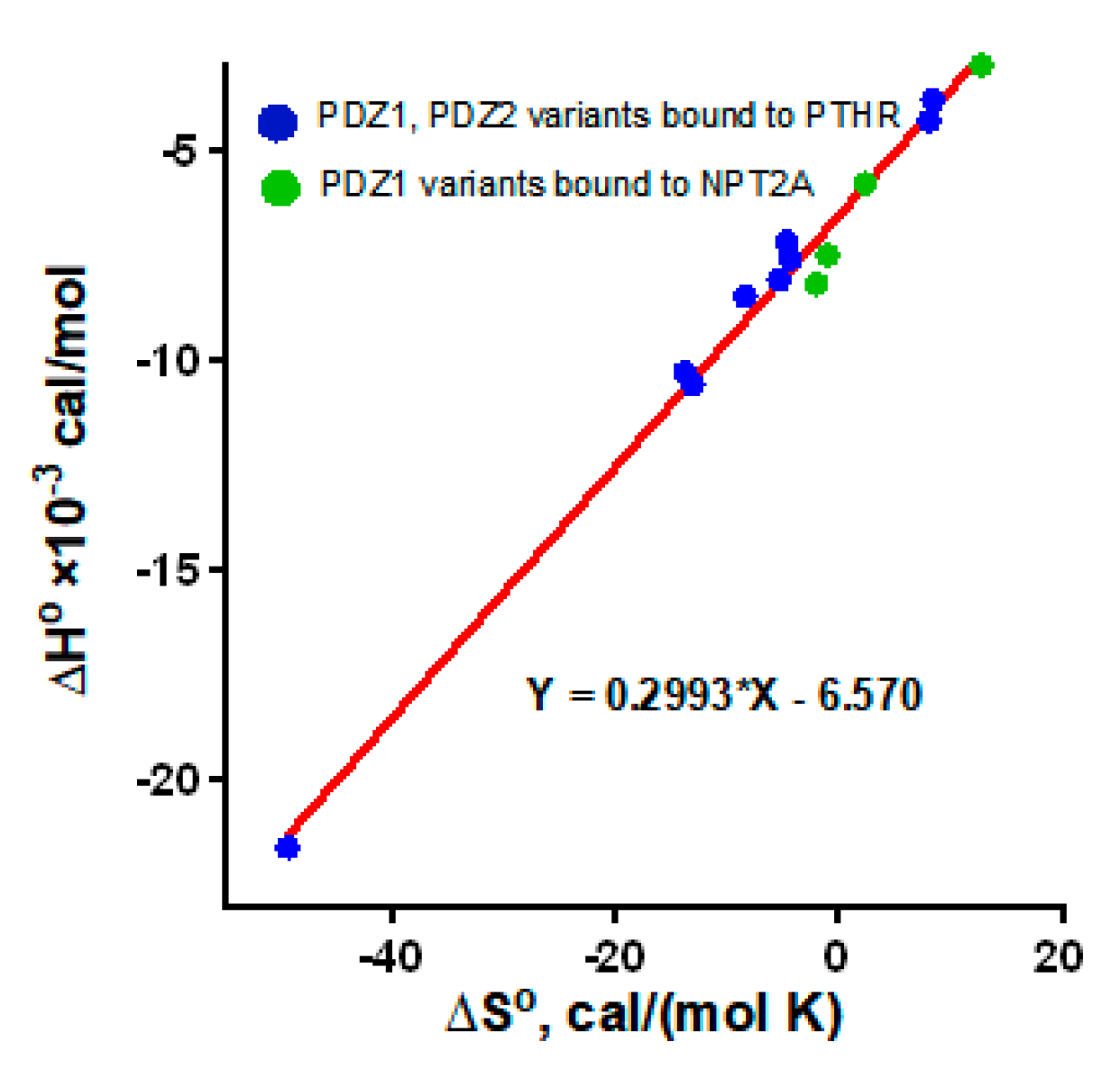
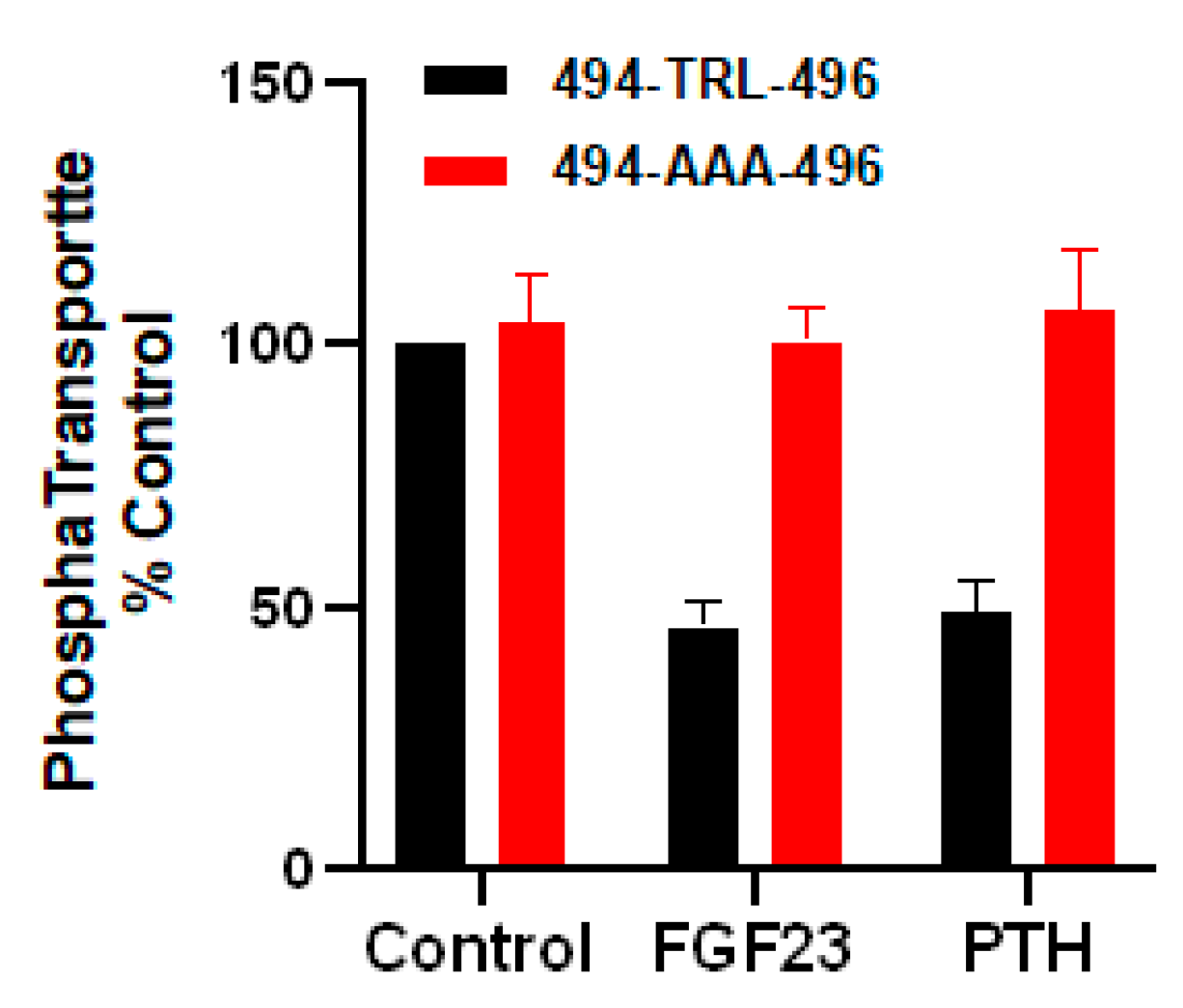
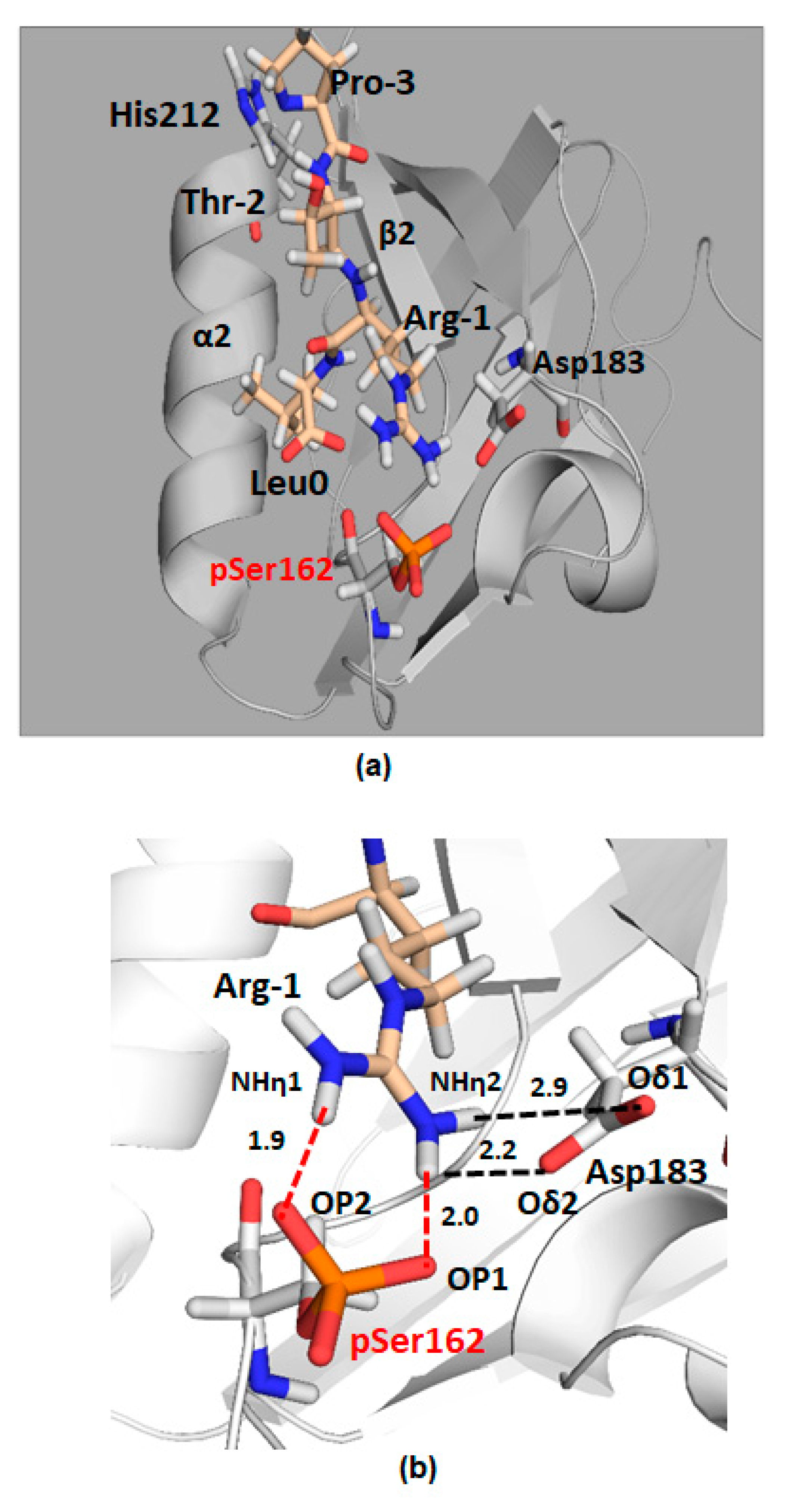
2.2. The Role of the NPT2A Internal -TRL- Motif Remains to Be Explored
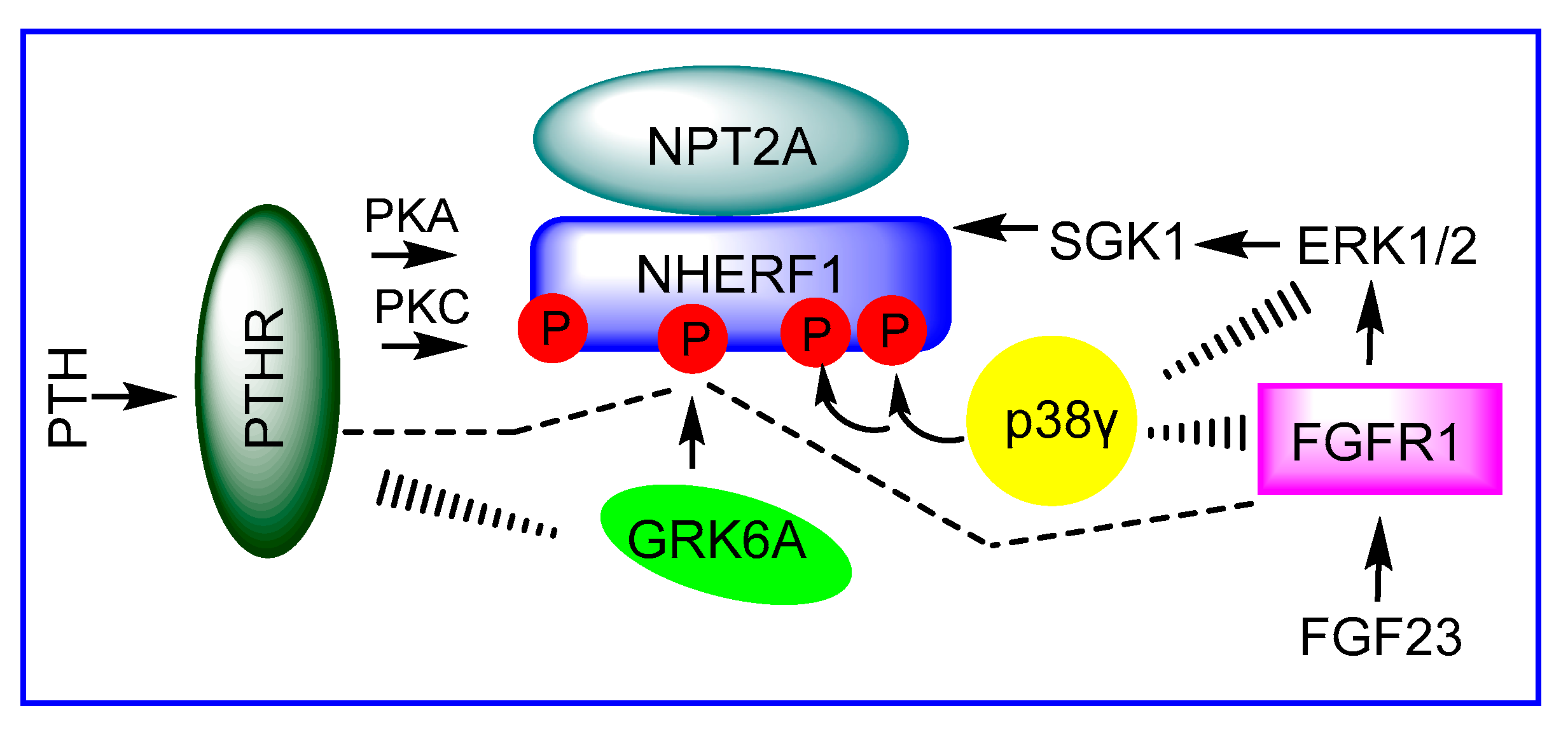
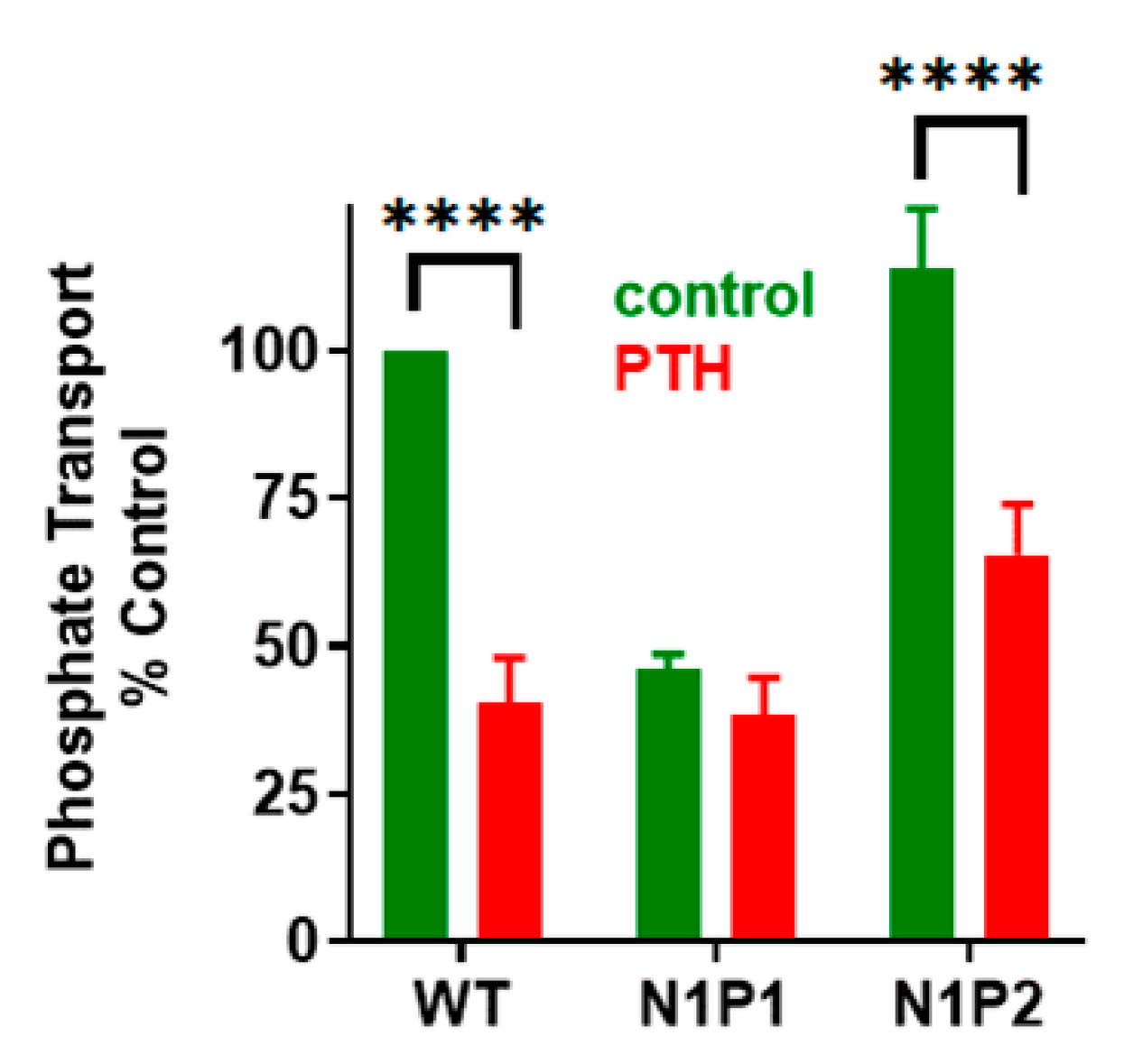
3. NPT2A-Dependent Hormone-Inhibitable Phosphate Transport Requires Association between PDZ2 and GRK6A
4. Conformational Reorganization of NHERF1 Regulates NHERF1-NPT2A Interactions
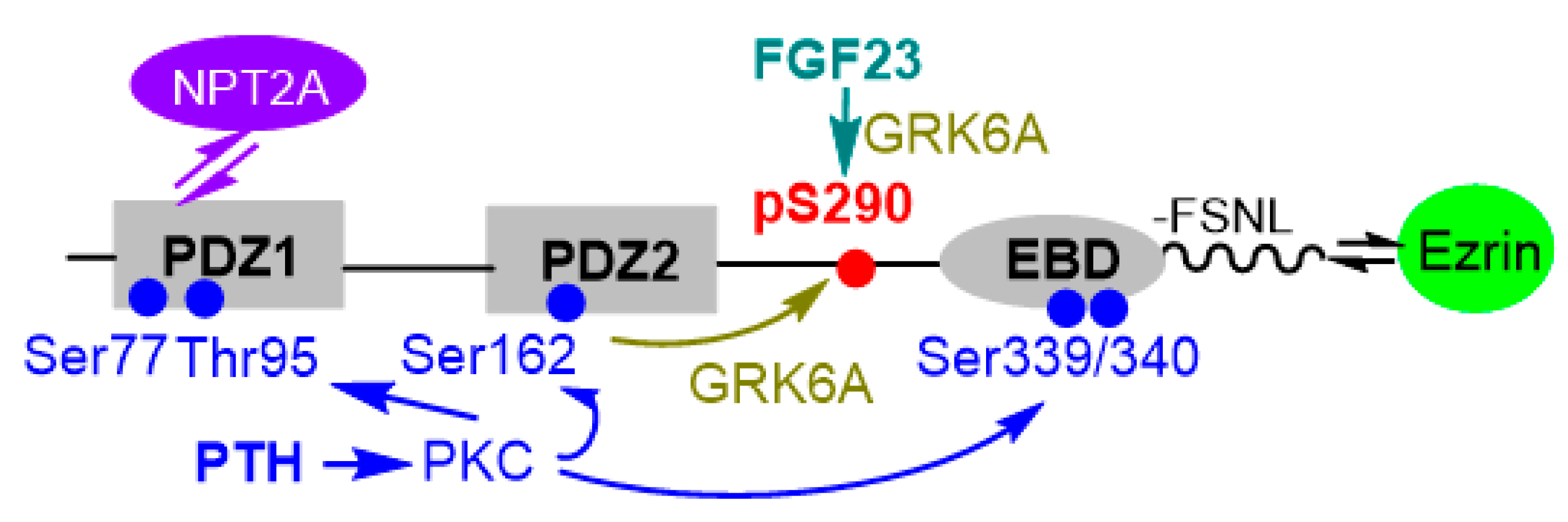
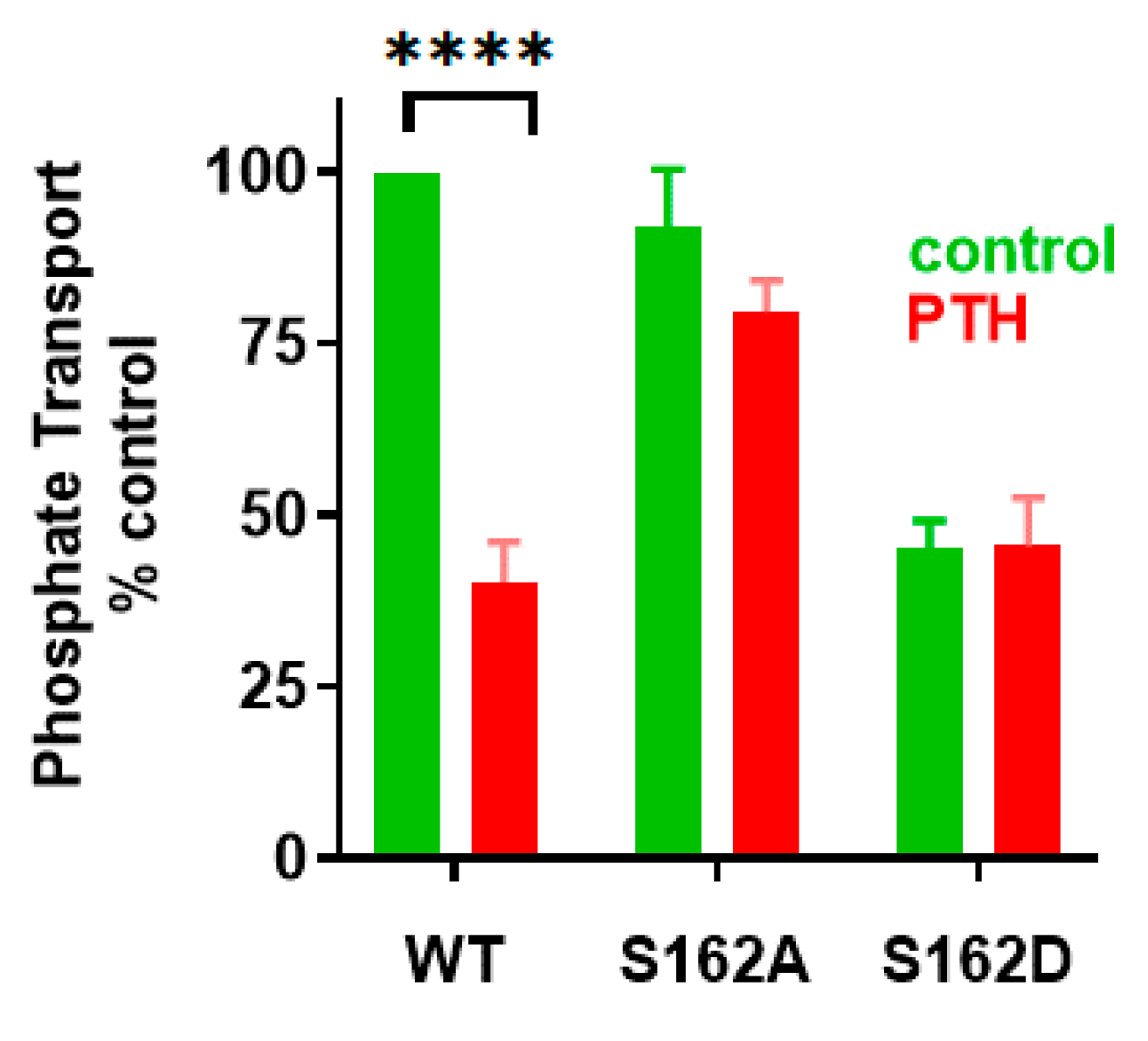
4.1. Phosphorylation of Ser290 Controls NHERF1 Conformation and Interactions with NPT2A
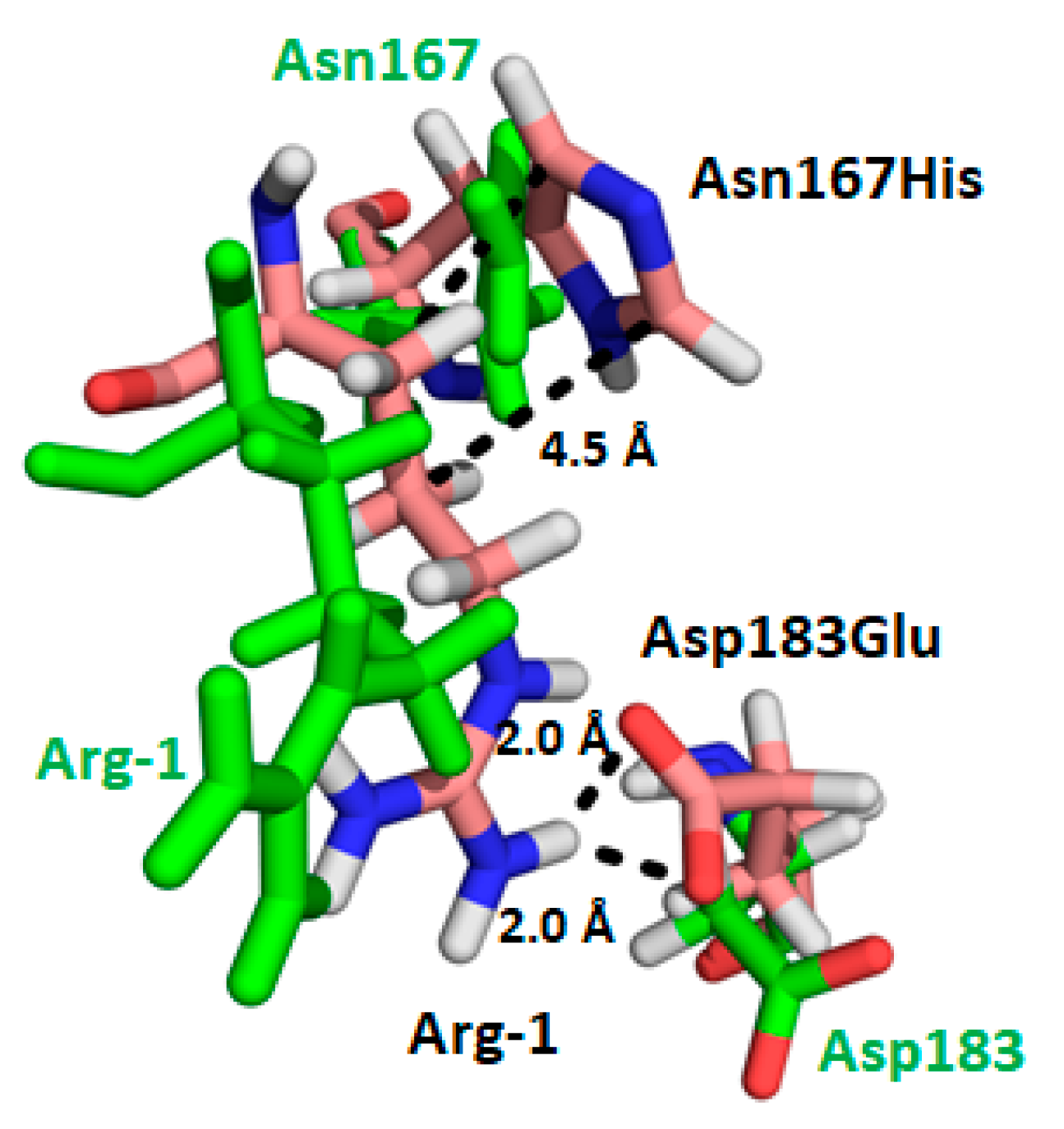
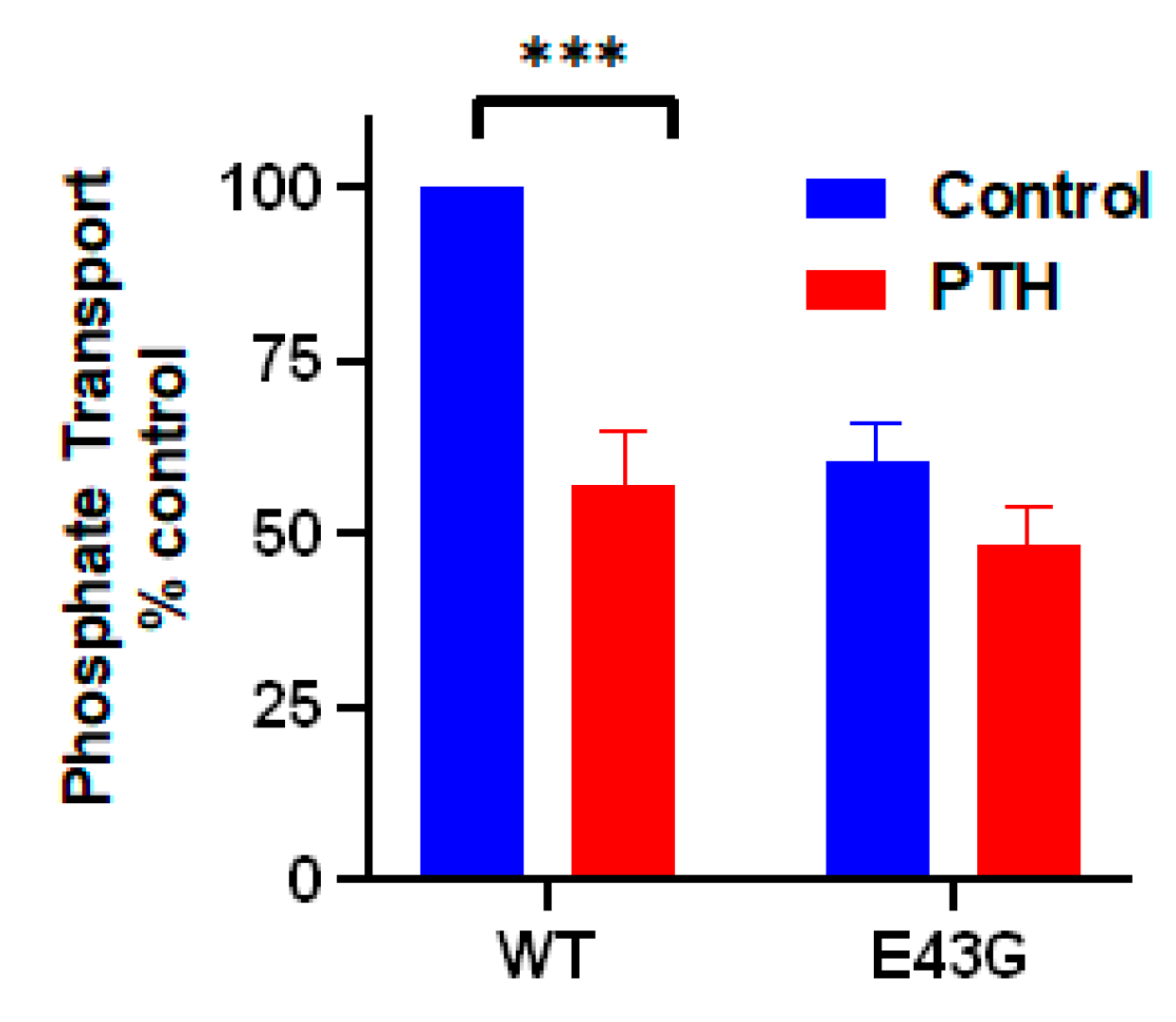
4.2. NHERF1 Disease-Associated Mutations (Leu110Val, Arg153Gln and Glu225Lys) Affect Conformational Landscape and Interaction with NPT2A
5. “Outside the Box” Determinants Are Involved in the Interaction between NHERF1 PDZ Domains and PTHR
6. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Donowitz, M.; Cha, B.; Zachos, N.C.; Brett, C.L.; Sharma, A.; Tse, C.M.; Li, X. NHERF family and NHE3 regulation. J. Physiol. 2005, 567(Pt. 1), 3–11. [Google Scholar] [CrossRef] [PubMed]
- Ardura, J.A.; Friedman, P.A. Regulation of G protein-coupled receptor function by Na+/H+ exchange regulatory factors. Pharmacol. Rev. 2011, 63, 882–900. [Google Scholar] [CrossRef] [PubMed]
- Romero, G.; von Zastrow, M.; Friedman, P.A. Role of PDZ proteins in regulating trafficking, signaling, and function of GPCRs: Means, motif, and opportunity. Adv. Pharmacol. 2011, 62, 279–314. [Google Scholar] [PubMed]
- Wang, B.; Means, C.K.; Yang, Y.M.; Mamonova, T.; Bisello, A.; Altschuler, D.L.; Scott, J.D.; Friedman, P.A. Ezrin-anchored protein kinase A coordinates phosphorylation-dependent disassembly of a NHERF1 ternary complex to regulate hormone-sensitive phosphate transport. J. Biol. Chem. 2012, 287, 24148–24163. [Google Scholar] [CrossRef]
- Wagner, C.A.; Rubio-Aliaga, I.; Biber, J.; Hernando, N. Genetic diseases of renal phosphate handling. Nephrol. Dial. Transplant. 2014, 29 (Suppl. 4), iv45–iv54. [Google Scholar] [CrossRef]
- Hernando, N. NaPi-IIa interacting partners and their (un)known functional roles. Pflugers Arch. 2019, 471, 67–82. [Google Scholar] [CrossRef]
- Levi, M.; Gratton, E.; Forster, I.C.; Hernando, N.; Wagner, C.A.; Biber, J.; Sorribas, V.; Murer, H. Mechanisms of phosphate transport. Nat. Rev. Nephrol. 2019, 15, 482–500. [Google Scholar] [CrossRef]
- Hernando, N.; Gagnon, K.; Lederer, E. Phosphate Transport in Epithelial and Nonepithelial Tissue. Physiol. Rev. 2021, 101, 1–35. [Google Scholar] [CrossRef]
- Karim, Z.; Gerard, B.; Bakouh, N.; Alili, R.; Leroy, C.; Beck, L.; Silve, C.; Planelles, G.; Urena-Torres, P.; Grandchamp, B.; et al. NHERF1 mutations and responsiveness of renal parathyroid hormone. N. Engl. J. Med. 2008, 359, 1128–1135. [Google Scholar] [CrossRef]
- Gordon, R.J.; Li, D.; Doyle, D.; Zaritsky, J.; Levine, M.A. Digenic heterozygous mutations in SLC34A3 and SLC34A1 cause dominant hypophosphatemic rickets with hypercalciuria. J. Clin. Endocrinol. Metab. 2020, 105, 2392–2400. [Google Scholar] [CrossRef]
- Kang, S.J.; Lee, R.; Kim, H.S. Infantile hypercalcemia with novel compound heterozygous mutation in SLC34A1 encoding renal sodium-phosphate cotransporter 2a: A case report. Ann. Pediatr. Endocrinol. Metab. 2019, 24, 64–67. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.A.; Rubio-Aliaga, I.; Hernando, N. Renal phosphate handling and inherited disorders of phosphate reabsorption: An update. Pediatr. Nephrol. 2019, 34, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Shenolikar, S.; Voltz, J.W.; Minkoff, C.M.; Wade, J.B.; Weinman, E.J. Targeted disruption of the mouse NHERF-1 gene promotes internalization of proximal tubule sodium-phosphate cotransporter type IIa and renal phosphate wasting. Proc. Natl. Acad. Sci. USA 2002, 99, 11470–11475. [Google Scholar] [CrossRef] [PubMed]
- Morales, F.C.; Takahashi, Y.; Kreimann, E.L.; Georgescu, M.M. Ezrin-radixin-moesin (ERM)-binding phosphoprotein 50 organizes ERM proteins at the apical membrane of polarized epithelia. Proc. Natl. Acad. Sci. USA 2004, 101, 17705–17710. [Google Scholar] [CrossRef]
- Hernando, N.; Deliot, N.; Gisler, S.M.; Lederer, E.; Weinman, E.J.; Biber, J.; Murer, H. PDZ-domain interactions and apical expression of type IIa Na/Pi cotransporters. Proc. Natl. Acad. Sci. USA 2002, 99, 11957–11962. [Google Scholar] [CrossRef]
- Weinman, E.J.; Steplock, D.; Shenolikar, S.; Biswas, R. Fibroblast growth factor-23-mediated inhibition of renal phosphate transport in mice requires sodium-hydrogen exchanger regulatory factor-1 (NHERF-1) and synergizes with parathyroid hormone. J. Biol. Chem. 2011, 286, 37216–37221. [Google Scholar] [CrossRef]
- Mahon, M.J.; Donowitz, M.; Yun, C.C.; Segre, G.V. Na+/H+ exchanger regulatory factor 2 directs parathyroid hormone 1 receptor signalling. Nature 2002, 417, 858–861. [Google Scholar] [CrossRef]
- Mahon, M.J.; Segre, G.V. Stimulation by parathyroid hormone of a NHERF-1-assembled complex consisting of the parathyroid hormone I receptor, phospholipase C beta, and actin increases intracellular calcium in opossum kidney cells. J. Biol. Chem. 2004, 279, 23550–23558. [Google Scholar] [CrossRef]
- Khundmiri, S.J.; Ahmad, A.; Bennett, R.E.; Weinman, E.J.; Steplock, D.; Cole, J.; Baumann, P.D.; Lewis, J.; Singh, S.; Clark, B.J.; et al. Novel regulatory function for NHERF-1 in Npt2a transcription. Am. J. Physiol. Renal Physiol. 2008, 294, F840–F849. [Google Scholar] [CrossRef]
- Reczek, D.; Berryman, M.; Bretscher, A. Identification of EBP50: A PDZ-containing phosphoprotein that associates with members of the ezrin-radixin-moesin family. J. Cell Biol. 1997, 139, 169–179. [Google Scholar] [CrossRef]
- Reczek, D.; Bretscher, A. The carboxyl-terminal region of EBP50 binds to a site in the amino-terminal domain of ezrin that is masked in the dormant molecule. J. Biol. Chem. 1998, 273, 18452–18458. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Bisello, A.; Yang, Y.; Romero, G.G.; Friedman, P.A. NHERF1 regulates parathyroid hormone receptor membrane retention without affecting recycling. J. Biol. Chem. 2007, 282, 36214–36222. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, S.; Leung, T.; Ladias, J.A. Structural determinants of the Na+/H+ exchanger regulatory factor interaction with the β2 adrenergic and platelet-derived growth factor receptors. J. Biol. Chem. 2002, 277, 18973–18978. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, S.; Leung, T.; Ladias, J.A.A. Structural basis of the Na+/H+ exchanger regulatory factor PDZ1 interaction with the carboxyl-terminal region of the cystic fibrosis transmembrane conductance regulator. J. Biol. Chem. 2001, 276, 19683–19686. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.R.; Wu, Y.N.; Jiang, Y.Y.; Wang, S.; Hou, Y.N.; Guan, X.Q.; Brunzelle, J.; Sirinupong, N.; Sheng, S.J.; Li, C.Y.; et al. Structural insights into neutrophilic migration revealed by the crystal structure of the chemokine receptor CXCR2 in complex with the first PDZ domain of NHERF1. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Holcomb, J.; Jiang, Y.Y.; Guan, X.Q.; Trescott, L.; Lu, G.R.; Hou, Y.N.; Wang, S.; Brunzelle, J.; Sirinupong, N.; Li, C.Y.; et al. Crystal structure of the NHERF1 PDZ2 domain in complex with the chemokine receptor CXCR2 reveals probable modes of PDZ2 dimerization. Biochem. Biophys. Res. Commun. 2014, 448, 169–174. [Google Scholar] [CrossRef]
- Karthikeyan, S.; Leung, T.; Birrane, G.; Webster, G.; Ladias, J.A.A. Crystal structure of the PDZ1 domain of human Na+/H+ exchanger regulatory factor provides insights into the mechanism of carboxyl-terminal leucine recognition by class I PDZ domains. J. Mol. Biol. 2001, 308, 963–973. [Google Scholar] [CrossRef]
- Voltz, J.W.; Brush, M.; Sikes, S.; Steplock, D.; Weinman, E.J.; Shenolikar, S. Phosphorylation of PDZ1 domain attenuates NHERF-1 binding to cellular targets. J. Biol. Chem. 2007, 282, 33879–33887. [Google Scholar] [CrossRef]
- Mamonova, T.; Zhang, Q.; Chandra, M.; Collins, B.M.; Sarfo, E.; Bu, Z.; Xiao, K.; Bisello, A.; Friedman, P.A. Origins of PDZ binding specificity. A computational and experimental study using NHERF1 and the parathyroid hormone receptor. Biochemistry 2017, 56, 2584–2593. [Google Scholar] [CrossRef]
- Ernst, A.; Appleton, B.A.; Ivarsson, Y.; Zhang, Y.; Gfeller, D.; Wiesmann, C.; Sidhu, S.S. A structural portrait of the PDZ domain family. J. Mol. Biol. 2014, 426, 3509–3519. [Google Scholar] [CrossRef]
- McGarvey, J.C.; Xiao, K.; Bowman, S.L.; Mamonova, T.; Zhang, Q.; Bisello, A.; Sneddon, W.B.; Ardura, J.A.; Jean-Alphonse, F.; Vilardaga, J.P.; et al. Actin-Sorting Nexin 27 (SNX27)-Retromer Complex Mediates Rapid Parathyroid Hormone Receptor Recycling. J. Biol. Chem. 2016, 291, 10986–11002. [Google Scholar] [CrossRef] [PubMed]
- Clairfeuille, T.; Mas, C.; Chan, A.S.; Yang, Z.; Tello-Lafoz, M.; Chandra, M.; Widagdo, J.; Kerr, M.C.; Paul, B.; Merida, I.; et al. A molecular code for endosomal recycling of phosphorylated cargos by the SNX27-retromer complex. Nat. Struct. Mol. Biol. 2016, 23, 921–932. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, S.W.; Albertsen, L.; Moran, G.E.; Levesque, B.; Pedersen, S.B.; Bartels, L.; Wapenaar, H.; Ye, F.; Zhang, M.; Bowen, M.E.; et al. Site-specific phosphorylation of PSD-95 PDZ domains reveals fine-tuned regulation of protein-protein interactions. ACS Chem. Biol. 2017, 12, 2313–2323. [Google Scholar] [CrossRef] [PubMed]
- Gogl, G.; Biri-Kovacs, B.; Durbesson, F.; Jane, P.; Nomine, Y.; Kostmann, C.; Bilics, V.; Simon, M.; Remenyi, A.; Vincentelli, R.; et al. Rewiring of RSK-PDZ interactome by linear motif phosphorylation. J. Mol. Biol. 2019, 431, 1234–1249. [Google Scholar] [CrossRef]
- Amacher, J.F.; Brooks, L.; Hampton, T.H.; Madden, D.R. Specificity in PDZ-peptide interaction networks: Computational analysis and review. J. Struct Biol. X 2020, 4, 100022. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Ju, J.H.; Orlova, N.; Khajeh, J.A.; Cowburn, D.; Bu, Z.M. Ligand-induced dynamic changes in extended PDZ domains from NHERF1. J. Mol. Biol. 2013, 425, 2509–2528. [Google Scholar] [CrossRef]
- Wang, C.K.; Pan, L.; Chen, J.; Zhang, M. Extensions of PDZ domains as important structural and functional elements. Protein Cell 2010, 1, 737–751. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Dai, Z.P.; Li, J.Q.; Baxter, S.; Callaway, D.J.E.; Cowburn, D.; Bu, Z.M. A conformational switch in the scaffolding protein NHERF1 controls autoinhibition and complex formation. J. Biol. Chem. 2010, 285, 9981–9994. [Google Scholar] [CrossRef]
- Morales, F.C.; Takahashi, Y.; Momin, S.; Adams, H.; Chen, X.; Georgescu, M.M. NHERF1/EBP50 head-to-tail intramolecular interaction masks association with PDZ domain ligands. Mol. Cell. Biol. 2007, 27, 2527–2537. [Google Scholar] [CrossRef]
- Shenolikar, S.; Minkoff, C.M.; Steplock, D.A.; Evangelista, C.; Liu, M.Z.; Weinman, E.J. N-terminal PDZ domain is required for NHERF dimerization. FEBS Lett. 2001, 489, 233–236. [Google Scholar] [CrossRef]
- Lau, A.G.; Hall, R.A. Oligomerization of NHERF-1 and NHERF-2 PDZ domains: Differential regulation by association with receptor carboxyl-termini and by phosphorylation. Biochemistry 2001, 40, 8572–8580. [Google Scholar] [CrossRef] [PubMed]
- Fouassier, L.; Nichols, M.T.; Gidey, E.; McWilliams, R.R.; Robin, H.; Finnigan, C.; Howell, K.E.; Housset, C.; Doctor, R.B. Protein kinase C regulates the phosphorylation and oligomerization of ERM binding phosphoprotein 50. Exp. Cell Res. 2005, 306, 264–273. [Google Scholar] [CrossRef]
- Li, J.Q.; Poulikakos, P.I.; Dai, Z.P.; Testa, J.R.; Callaway, D.J.E.; Bu, Z.M. Protein kinase c phosphorylation disrupts Na+/H+ exchanger regulatory factor 1 autoinhibition and promotes cystic fibrosis transmembrane conductance regulator macromolecular assembly. J. Biol. Chem. 2007, 282, 27086–27099. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xiao, K.; Paredas, J.M.; Mamonova, T.; Sneddon, W.B.; Liu, H.; Wang, D.; Li, S.; McGarvey, J.C.; Uehling, D.; et al. Parathyroid hormone initiates dynamic NHERF1 phosphorylation cycling and conformational changes that regulate NPT2A-dependent phosphate transport. J. Biol. Chem. 2019, 294, 4546–4571. [Google Scholar] [CrossRef] [PubMed]
- Mamonova, T.; Zhang, Q.; Khajeh, J.A.; Bu, Z.; Bisello, A.; Friedman, P.A. Canonical and noncanonical sites determine NPT2A binding selectivity to NHERF1 PDZ1. PLoS ONE 2015, 10, e0129554. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.A.; Spurney, R.F.; Premont, R.T.; Rahman, N.; Blitzer, J.T.; Pitcher, J.A.; Lefkowitz, R.J. G protein-coupled receptor kinase 6A phosphorylates the Na+/H+ exchanger regulatory factor via a PDZ domain-mediated interaction. J. Biol. Chem. 1999, 274, 24328–24334. [Google Scholar] [CrossRef]
- Cardone, R.A.; Bellizzi, A.; Busco, G.; Weinman, E.J.; Dell’aquila, M.E.; Casavola, V.; Azzariti, A.; Mangia, A.; Paradiso, A.; Reshkin, S.J. The NHERF1 PDZ2 domain regulates PKA-RhoA-p38-mediated NHE1 activation and invasion in breast tumor cells. Mol. Biol. Cell 2007, 18, 1768–1780. [Google Scholar] [CrossRef]
- Salyer, S.; Lesousky, N.; Weinman, E.J.; Clark, B.J.; Lederer, E.D.; Khundmiri, S.J. Dopamine regulation of Na+-K+-ATPase requires the PDZ-2 domain of sodium hydrogen regulatory factor-1 (NHERF-1) in opossum kidney cells. Am. J. Physiol. Cell Physiol. 2011, 300, C425–C434. [Google Scholar] [CrossRef]
- Raghuram, V.; Hormuth, H.; Foskett, J.K. A kinase-regulated mechanism controls CFTR channel gating by disrupting bivalent PDZ domain interactions. Proc. Natl. Acad. Sci. USA 2003, 100, 9620–9625. [Google Scholar] [CrossRef]
- Khundmiri, S.J.; Weinman, E.J.; Steplock, D.; Cole, J.; Ahmad, A.; Baumann, P.D.; Barati, M.; Rane, M.J.; Lederer, E. Parathyroid hormone regulation of NA+,K+-ATPase requires the PDZ 1 domain of sodium hydrogen exchanger regulatory factor-1 in opossum kidney cells. J. Am. Soc. Nephrol. 2005, 16, 2598–2607. [Google Scholar] [CrossRef]
- Weinman, E.J.; Steplock, D.; Shenolikar, S.; Blanpied, T.A. Dynamics of PTH-induced disassembly of Npt2a/NHERF-1 complexes in living OK cells. Am. J. Physiol. Renal Physiol. 2011, 300, F231–F235. [Google Scholar] [CrossRef] [PubMed]
- Pouyssegur, J.; Sardet, C.; Franchi, A.; L’Allemain, G.; Paris, S. A specific mutation abolishing Na+/H+ antiport activity in hamster fibroblasts precludes growth at neutral and acidic pH. Proc. Natl. Acad. Sci. USA 1984, 81, 4833–4837. [Google Scholar] [CrossRef] [PubMed]
- Staudinger, J.; Lu, J.; Olson, E.N. Specific interaction of the PDZ domain protein PICK1 with the COOH terminus of protein kinase C-alpha. J. Biol. Chem. 1997, 272, 32019–32024. [Google Scholar] [CrossRef]
- Mahon, M.J. The parathyroid hormone 1 receptor directly binds to the FERM domain of ezrin, an interaction that supports apical receptor localization and signaling in LLC-PK1 cells. Mol. Endocrinol. 2009, 23, 1691–1701. [Google Scholar] [CrossRef]
- Cunningham, R.; XE, D.; Steplock, D.; Shenolikar, S.; Weinman, E.J. Defective PTH regulation of sodium-dependent phosphate transport in NHERF-1−/− renal proximal tubule cells and wild-type cells adapted to low-phosphate media. Am. J. Physiol. Renal Physiol. 2005, 289, F933–F9338. [Google Scholar] [CrossRef]
- Alshafie, W.; Chappe, F.G.; Li, M.; Anini, Y.; Chappe, V.M. VIP regulates CFTR membrane expression and function in Calu-3 cells by increasing its interaction with NHERF1 and P-ERM in a VPAC1- and PKCepsilon-dependent manner. Am. J. Physiol. Cell Physiol. 2014, 307, C107–C119. [Google Scholar] [CrossRef]
- Raghuram, V.; Mak, D.O.; Foskett, J.K. Regulation of cystic fibrosis transmembrane conductance regulator single-channel gating by bivalent PDZ-domain-mediated interaction. Proc. Natl. Acad. Sci. USA 2001, 98, 1300–1305. [Google Scholar] [CrossRef]
- Li, J.Q.; Dai, Z.P.; Jana, D.; Callaway, D.J.E.; Bu, Z.M. Ezrin controls the macromolecular complexes formed between an adapter protein Na+/H+ exchanger regulatory factor and the cystic fibrosis transmembrane conductance regulator. J. Biol. Chem. 2005, 280, 37634–37643. [Google Scholar] [CrossRef]
- Lee, J.H.; Richter, W.; Namkung, W.; Kim, K.H.; Kim, E.; Conti, M.; Lee, M.G. Dynamic regulation of cystic fibrosis transmembrane conductance regulator by competitive interactions of molecular adaptors. J. Biol. Chem. 2007, 282, 10414–10422. [Google Scholar] [CrossRef]
- Martin, E.R.; Barbieri, A.; Ford, R.C.; Robinson, R.C. In vivo crystals reveal critical features of the interaction between cystic fibrosis transmembrane conductance regulator (CFTR) and the PDZ2 domain of Na(+)/H(+) exchange cofactor NHERF1. J. Biol. Chem. 2020, 295, 4464–4476. [Google Scholar] [CrossRef]
- Rajagopal, A.; Braslavsky, D.; Lu, J.T.; Kleppe, S.; Clement, F.; Cassinelli, H.; Liu, D.S.; Liern, J.M.; Vallejo, G.; Bergada, I.; et al. Exome sequencing identifies a novel homozygous mutation in the phosphate transporter SLC34A1 in hypophosphatemia and nephrocalcinosis. J. Clin. Endocrinol. Metab. 2014, 99, E2451–E2456. [Google Scholar] [CrossRef] [PubMed]
- Mamonova, T.; Kurnikova, M.; Friedman, P.A. Structural basis for NHERF1 PDZ domain binding. Biochemistry 2012, 51, 3110–3120. [Google Scholar] [CrossRef] [PubMed]
- Vistrup-Parry, M.; Sneddon, W.B.; Bach, S.; Strømgaard, K.; Friedman, P.A.; Mamonova, T. Multisite NHERF1 phosphorylation controls GRK6A regulation of hormone-sensitive phosphate transport. J. Biol. Chem. under review.
- Rogerson, D.T.; Sachdeva, A.; Wang, K.; Haq, T.; Kazlauskaite, A.; Hancock, S.M.; Huguenin-Dezot, N.; Muqit, M.M.; Fry, A.M.; Bayliss, R.; et al. Efficient genetic encoding of phosphoserine and its nonhydrolyzable analog. Nat. Chem. Biol. 2015, 11, 496–503. [Google Scholar] [CrossRef]
- Toto, A.; Mattei, A.; Jemth, P.; Gianni, S. Understanding the role of phosphorylation in the binding mechanism of a PDZ domain. Protein Eng. Des. Sel. 2017, 30, 1–5. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Stanley, C.B.; Heller, W.T.; Friedman, P.A.; Bu, Z. Dynamic structure of the full-length scaffolding protein NHERF1 influences signaling complex assembly. J. Biol. Chem. 2019, 294, 11297–11310. [Google Scholar] [CrossRef]
- Iakoucheva, L.M.; Radivojac, P.; Brown, C.J.; O’Connor, T.R.; Sikes, J.G.; Obradovic, Z.; Dunker, A.K. The importance of intrinsic disorder for protein phosphorylation. Nucleic Acids Res. 2004, 32, 1037–1049. [Google Scholar] [CrossRef]
- Darling, A.L.; Uversky, V.N. Intrinsic Disorder and Posttranslational Modifications: The Darker Side of the Biological Dark Matter. Front. Genet. 2018, 9, 158. [Google Scholar] [CrossRef]
- Bhattacherjee, A.; Wallin, S. Exploring protein-peptide binding specificity through computational peptide screening. PLoS Comput. Biol. 2013, 9, e1003277. [Google Scholar] [CrossRef]
- Weinman, E.J.; Steplock, D.; Zhang, Y.; Biswas, R.; Bloch, R.J.; Shenolikar, S. Cooperativity between the phosphorylation of Thr95 and Ser77 of NHERF-1 in the hormonal regulation of renal phosphate transport. J. Biol. Chem. 2010, 285, 25134–25138. [Google Scholar] [CrossRef]
- Sun, C.; Mierke, D.F. Characterization of interactions of Na+/H+ exchanger regulatory factor-1 with the parathyroid hormone receptor and phospholipase C. J. Pept. Res. 2005, 65, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Cushing, P.R.; Fellows, A.; Villone, D.; Boisguerin, P.; Madden, D.R. The relative binding affinities of PDZ partners for CFTR: A biochemical basis for efficient endocytic recycling. Biochemistry 2008, 47, 10084–10098. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PDZ1 | PDZ2 |
|---|---|
| NPT2A (-ATRLa) | PKCα (-QSAV) |
| GRK6A (-PTRL) | GRK6A (-PTRL) |
| CFTR (-DTRL) | CFTR (-DTRL) |
| β2-AR (-DSSL) | TAZ/YAP65 (-LTWL) |
| PTHR (-ETVM) | PTHR (-ETVM) |
| PDGFR (-DSFL) | p38γ (-ETPL) |
| P2Y1 (-DTSL) | SGK1 (-DSFL) |
| PLCβ 1,2b,3 | PLCβ 3 (-NTQL) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mamonova, T.; Friedman, P.A. Noncanonical Sequences Involving NHERF1 Interaction with NPT2A Govern Hormone-Regulated Phosphate Transport: Binding Outside the Box. Int. J. Mol. Sci. 2021, 22, 1087. https://doi.org/10.3390/ijms22031087
Mamonova T, Friedman PA. Noncanonical Sequences Involving NHERF1 Interaction with NPT2A Govern Hormone-Regulated Phosphate Transport: Binding Outside the Box. International Journal of Molecular Sciences. 2021; 22(3):1087. https://doi.org/10.3390/ijms22031087
Chicago/Turabian StyleMamonova, Tatyana, and Peter A. Friedman. 2021. "Noncanonical Sequences Involving NHERF1 Interaction with NPT2A Govern Hormone-Regulated Phosphate Transport: Binding Outside the Box" International Journal of Molecular Sciences 22, no. 3: 1087. https://doi.org/10.3390/ijms22031087
APA StyleMamonova, T., & Friedman, P. A. (2021). Noncanonical Sequences Involving NHERF1 Interaction with NPT2A Govern Hormone-Regulated Phosphate Transport: Binding Outside the Box. International Journal of Molecular Sciences, 22(3), 1087. https://doi.org/10.3390/ijms22031087