Author Contributions
Conceptualization, A.S. and T.K.; methodology, T.K.; validation, H.A., K.I., and T.T.; formal analysis, T.W.; investigation, A.S., H.A., T.T., K.I., T.W., Y.K., and T.K.; data curation, K.I. and Y.K.; writing—original draft preparation, A.S., H.A., T.T., and T.K.; writing—review and editing, T.T. and T.K.; supervision, Y.K.; project administration, T.W.; funding acquisition, T.K. All authors have read and agreed to the published version of the manuscript.
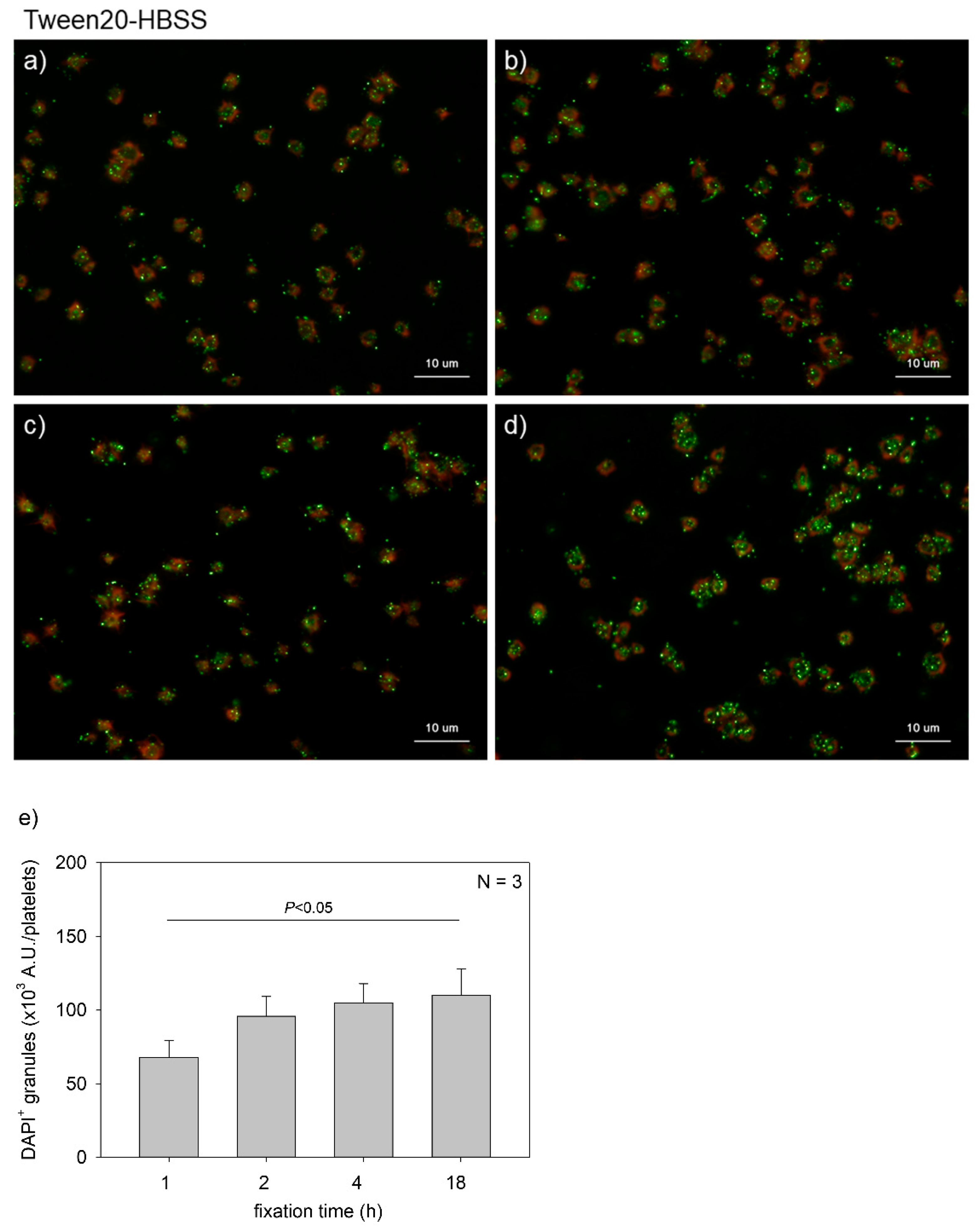
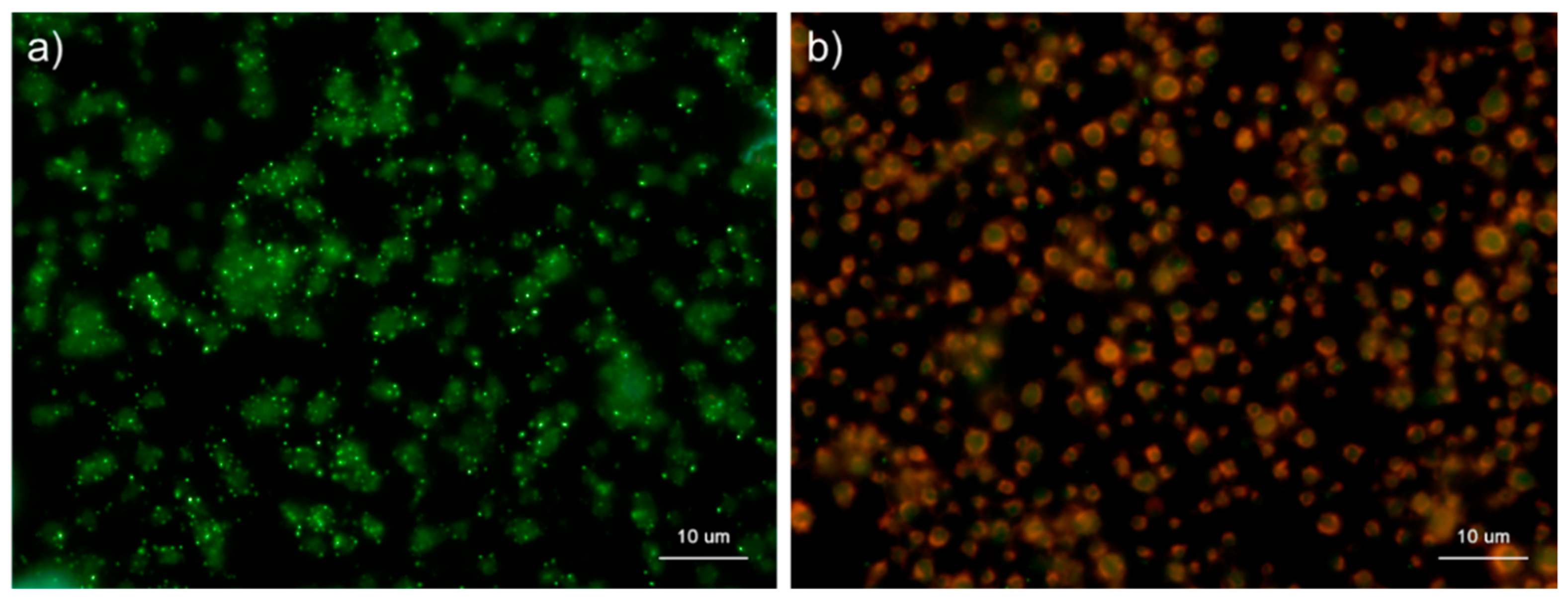
Figure 1.
Effects of fixation time on polyphosphate detection in platelets. Platelets suspended in phosphate-buffered saline were immobilized on glass slides using a Cytospin and fixed in 10% neutral-buffered formalin for (a) 1 h, (b) 2 h, (c) 4 h, or (d) 18 h. Platelets were then treated with 4′,6-diamidino-2-phenylindole (DAPI; green) and phalloidin (red) in 0.1% Tween-20-containing Hanks balanced salt solution and examined using a fluorescence microscope. (e) Quantification of DAPI staining in the fluorescence images (above). A significant difference was obtained via a parametric test but not by using a non-parametric test.
Figure 1.
Effects of fixation time on polyphosphate detection in platelets. Platelets suspended in phosphate-buffered saline were immobilized on glass slides using a Cytospin and fixed in 10% neutral-buffered formalin for (a) 1 h, (b) 2 h, (c) 4 h, or (d) 18 h. Platelets were then treated with 4′,6-diamidino-2-phenylindole (DAPI; green) and phalloidin (red) in 0.1% Tween-20-containing Hanks balanced salt solution and examined using a fluorescence microscope. (e) Quantification of DAPI staining in the fluorescence images (above). A significant difference was obtained via a parametric test but not by using a non-parametric test.
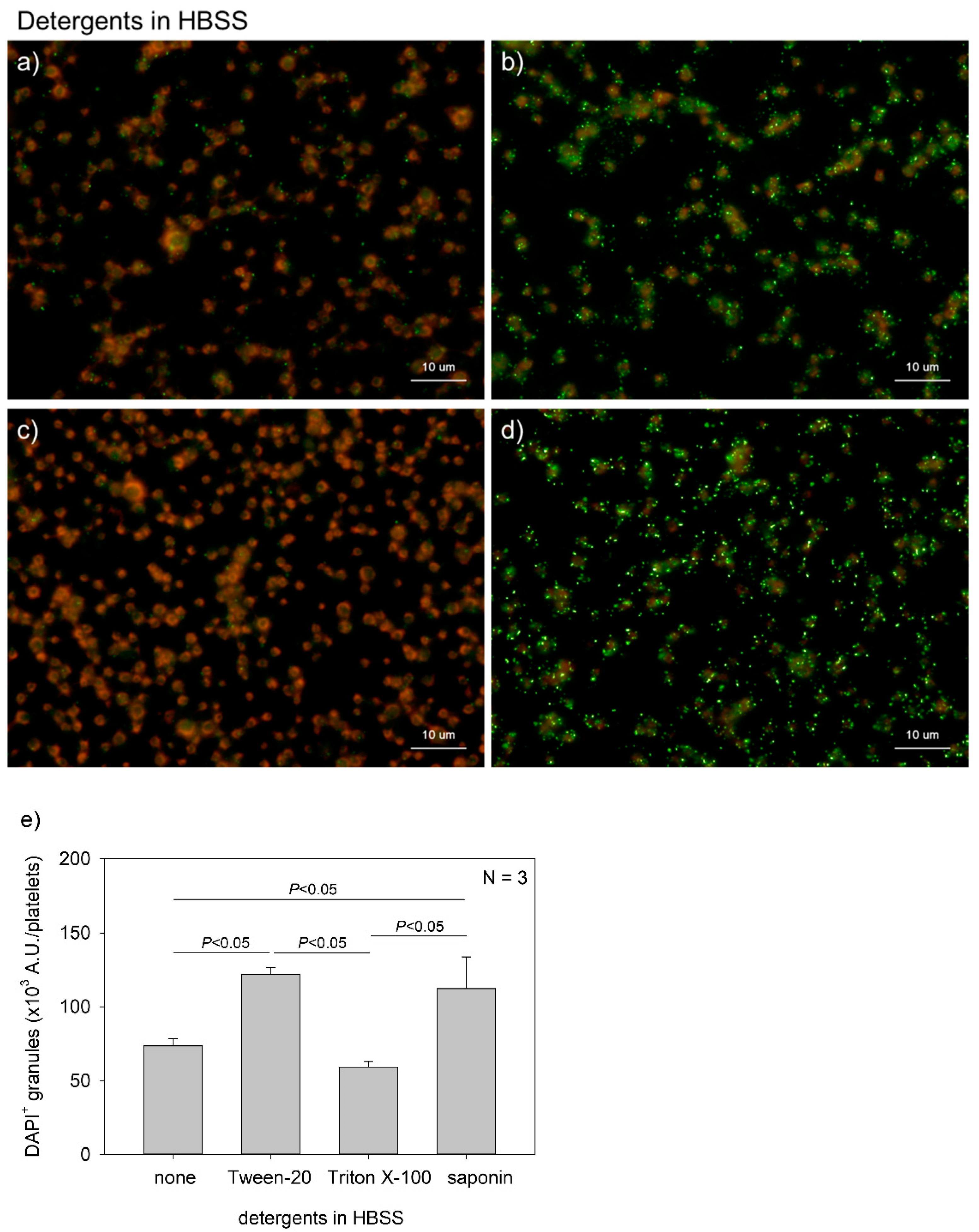
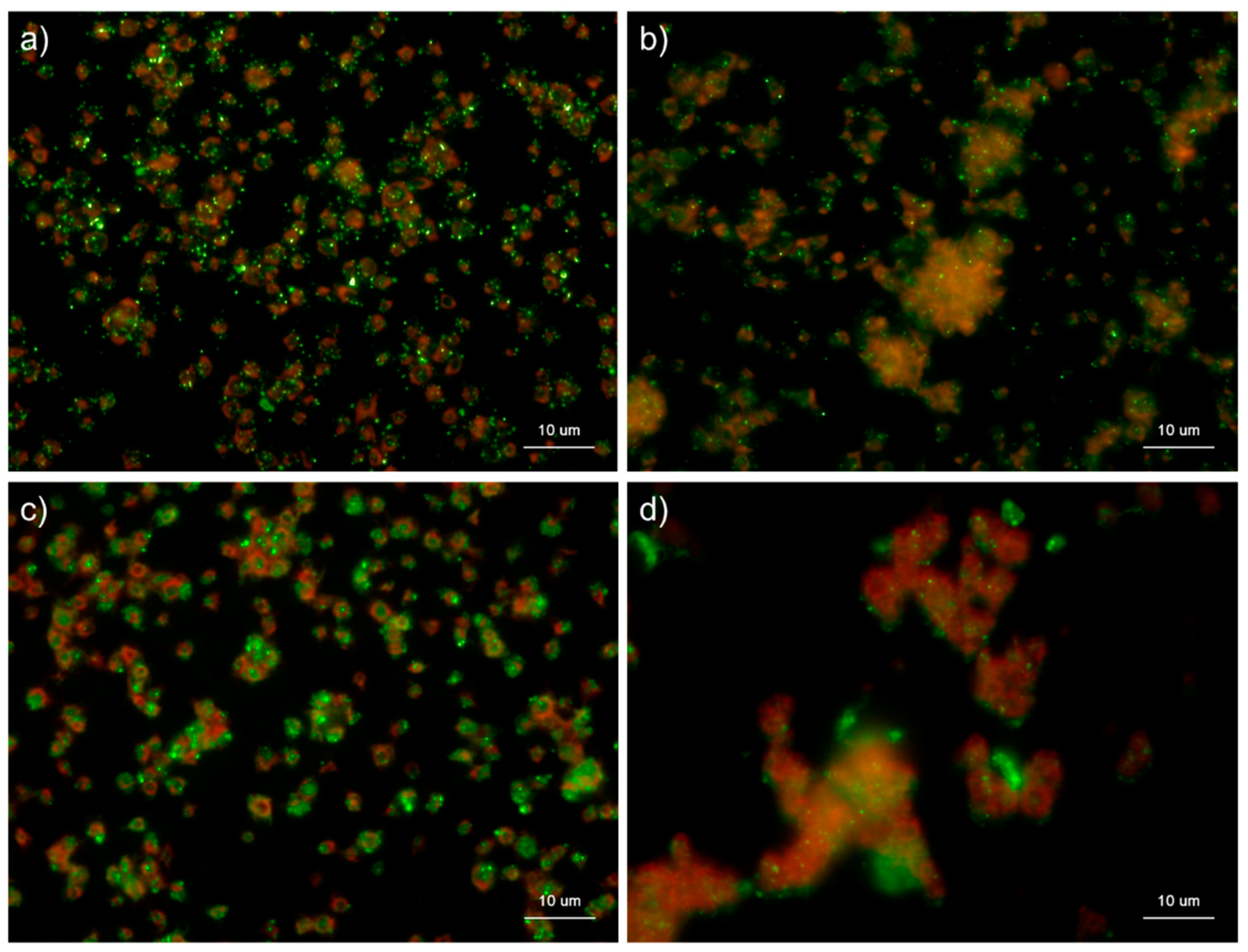
Figure 2.
Effects of detergent types on polyphosphate detection in platelets. Platelets immobilized on glass slides using a Cytospin were fixed in 10% neutral-buffered formalin for 4 h, then treated with 4′,6-diamidino-2-phenylindole (green) and phalloidin (red) in (a) detergent-free Hanks balanced salt solution (HBSS), (b) 0.1% Tween-20-containing HBSS, (c) 0.1% Triton-X-100-containing HBSS, or (d) 0.02% saponin-containing HBSS and examined using a fluorescence microscope. (e) Quantification of DAPI staining in the fluorescence images (above). A significant difference was obtained via a parametric test but not by using a non-parametric test.
Figure 2.
Effects of detergent types on polyphosphate detection in platelets. Platelets immobilized on glass slides using a Cytospin were fixed in 10% neutral-buffered formalin for 4 h, then treated with 4′,6-diamidino-2-phenylindole (green) and phalloidin (red) in (a) detergent-free Hanks balanced salt solution (HBSS), (b) 0.1% Tween-20-containing HBSS, (c) 0.1% Triton-X-100-containing HBSS, or (d) 0.02% saponin-containing HBSS and examined using a fluorescence microscope. (e) Quantification of DAPI staining in the fluorescence images (above). A significant difference was obtained via a parametric test but not by using a non-parametric test.
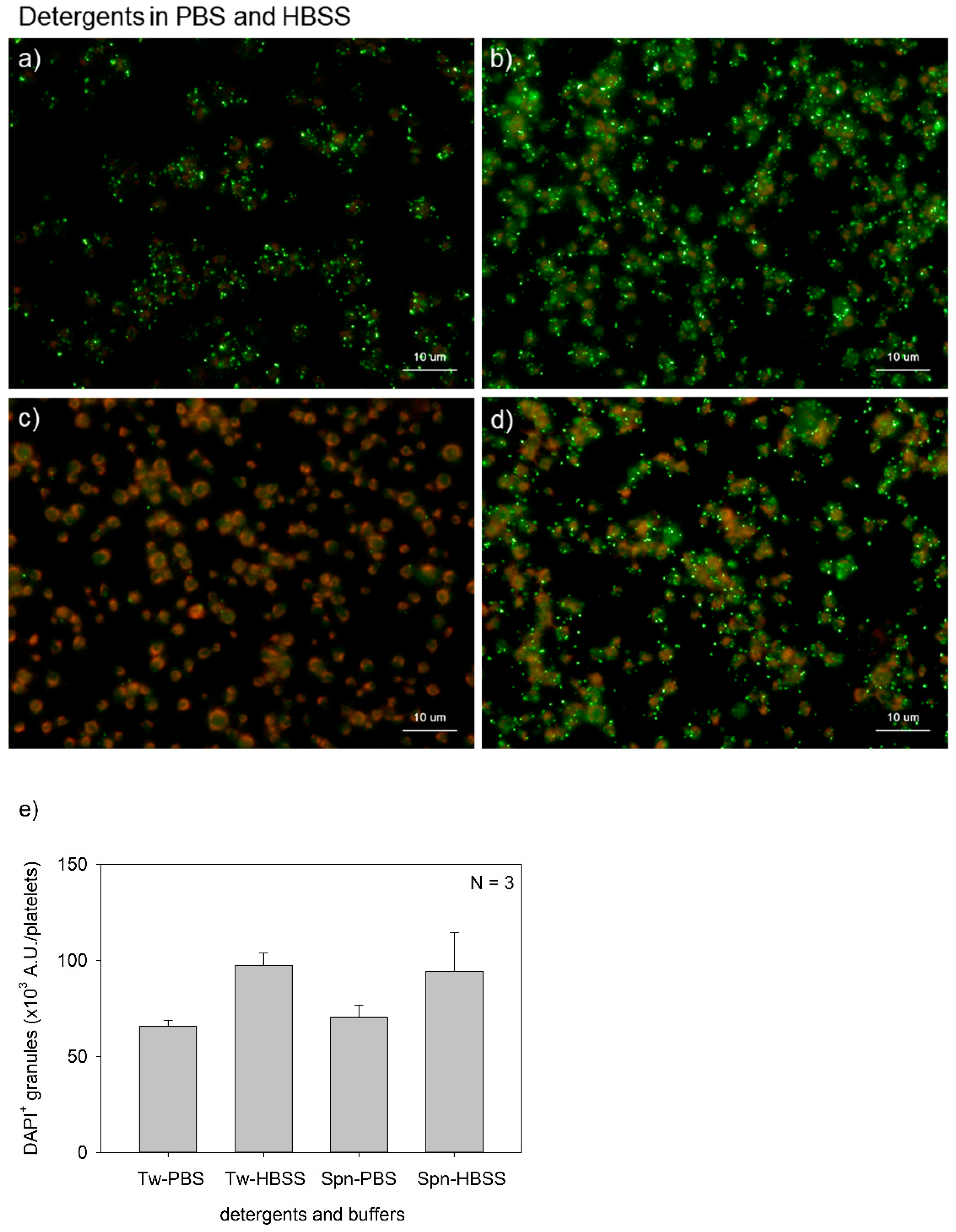
Figure 3.
Effects of buffer types on polyphosphate detection in platelets. Platelets were suspended in phosphate-buffered saline (PBS) and incubated for 60 min at room temperature (22–25 °C). Platelets were then immobilized on glass slides using a Cytospin, fixed in 10% neutral-buffered formalin for 4 h, and then treated with 4′,6-diamidino-2-phenylindole (green) and phalloidin (red) in (a) 0.1% Tween-20-containing PBS, (b) 0.1% Tween-20-containing Hanks balanced salt solution (HBSS), (c) 0.02% saponin-containing PBS, or (d) 0.02% saponin-containing HBSS and examined using a fluorescence microscope. (e) Quantification of DAPI staining in the fluorescence images (above). No significant differences were observed.
Figure 3.
Effects of buffer types on polyphosphate detection in platelets. Platelets were suspended in phosphate-buffered saline (PBS) and incubated for 60 min at room temperature (22–25 °C). Platelets were then immobilized on glass slides using a Cytospin, fixed in 10% neutral-buffered formalin for 4 h, and then treated with 4′,6-diamidino-2-phenylindole (green) and phalloidin (red) in (a) 0.1% Tween-20-containing PBS, (b) 0.1% Tween-20-containing Hanks balanced salt solution (HBSS), (c) 0.02% saponin-containing PBS, or (d) 0.02% saponin-containing HBSS and examined using a fluorescence microscope. (e) Quantification of DAPI staining in the fluorescence images (above). No significant differences were observed.
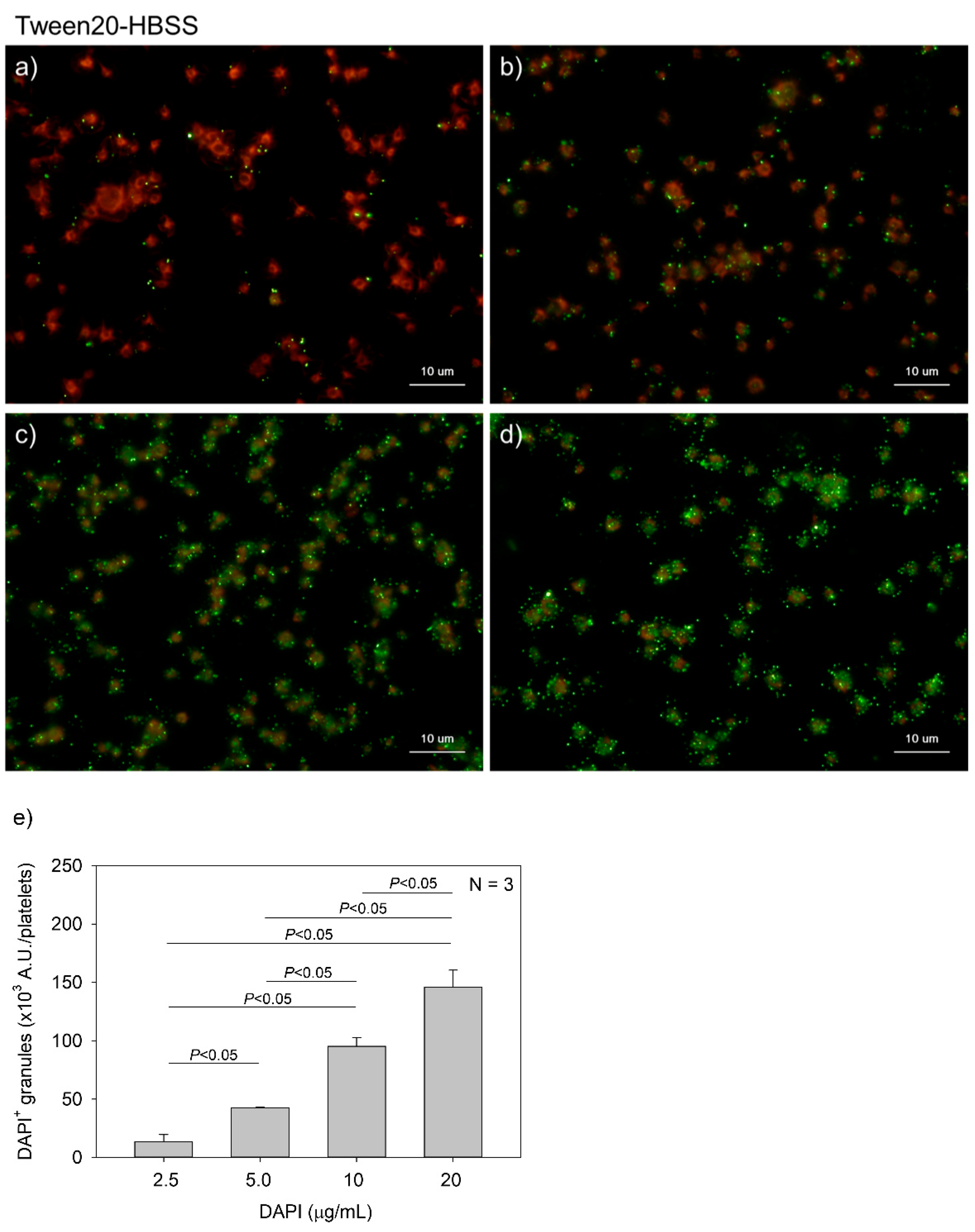
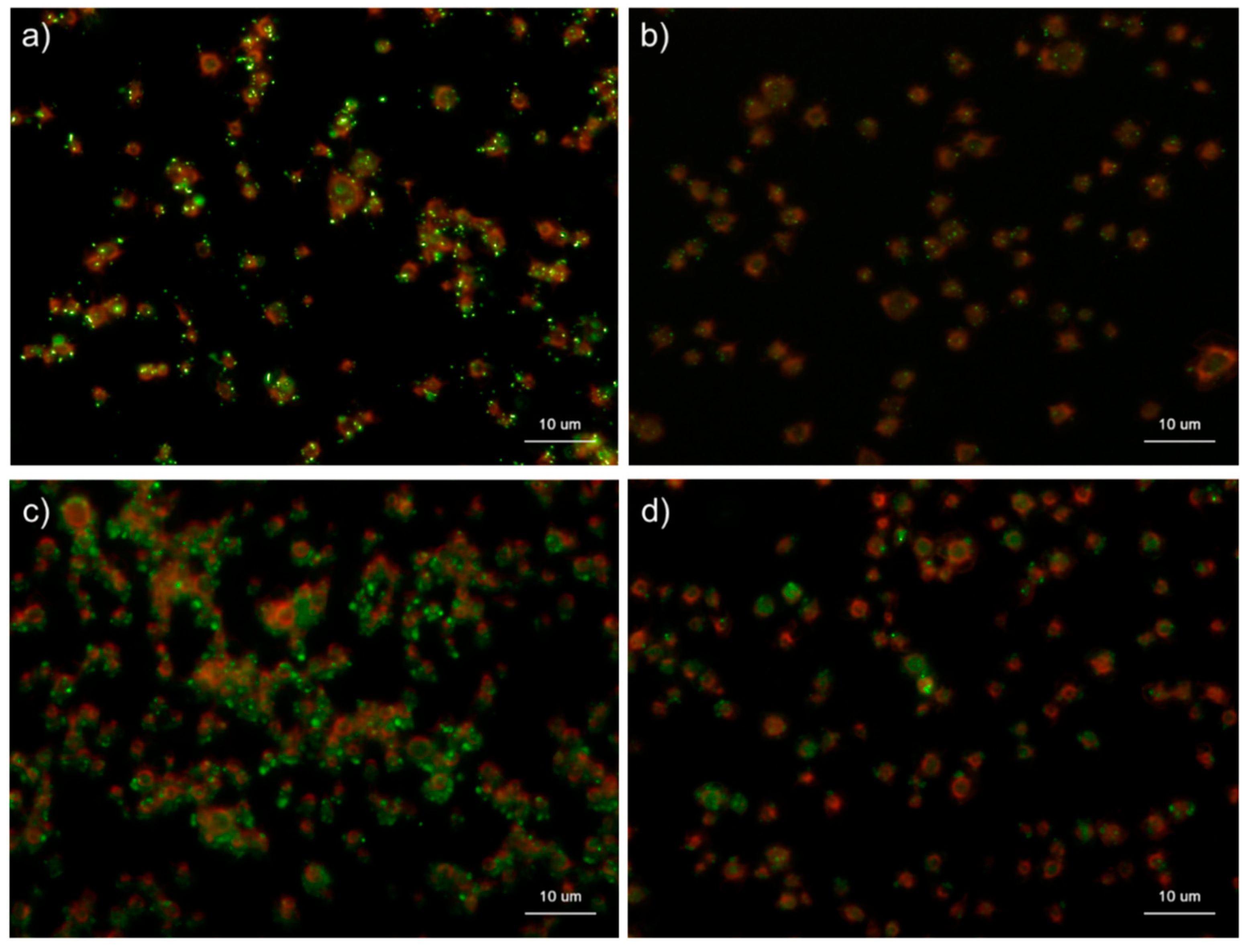
Figure 4.
Effects of 4′,6-diamidino-2-phenylindole (DAPI) concentrations on polyphosphate detection in platelets in 0.1% Tween-20-containing Hanks balanced salt solution (Tw-HBSS). Platelets immobilized on glass slides using a Cytospin were fixed in 10% neutral-buffered formalin for 4 h and then treated with DAPI at (a) 2.5, (b) 5.0, (c) 10, or (d) 20 μg/mL (green) and phalloidin (red) in Tw-HBSS and examined using a fluorescence microscope. (e) Quantitative analysis of the fluorescence images. Significant differences were obtained by a parametric test but not by a non-parametric test.
Figure 4.
Effects of 4′,6-diamidino-2-phenylindole (DAPI) concentrations on polyphosphate detection in platelets in 0.1% Tween-20-containing Hanks balanced salt solution (Tw-HBSS). Platelets immobilized on glass slides using a Cytospin were fixed in 10% neutral-buffered formalin for 4 h and then treated with DAPI at (a) 2.5, (b) 5.0, (c) 10, or (d) 20 μg/mL (green) and phalloidin (red) in Tw-HBSS and examined using a fluorescence microscope. (e) Quantitative analysis of the fluorescence images. Significant differences were obtained by a parametric test but not by a non-parametric test.
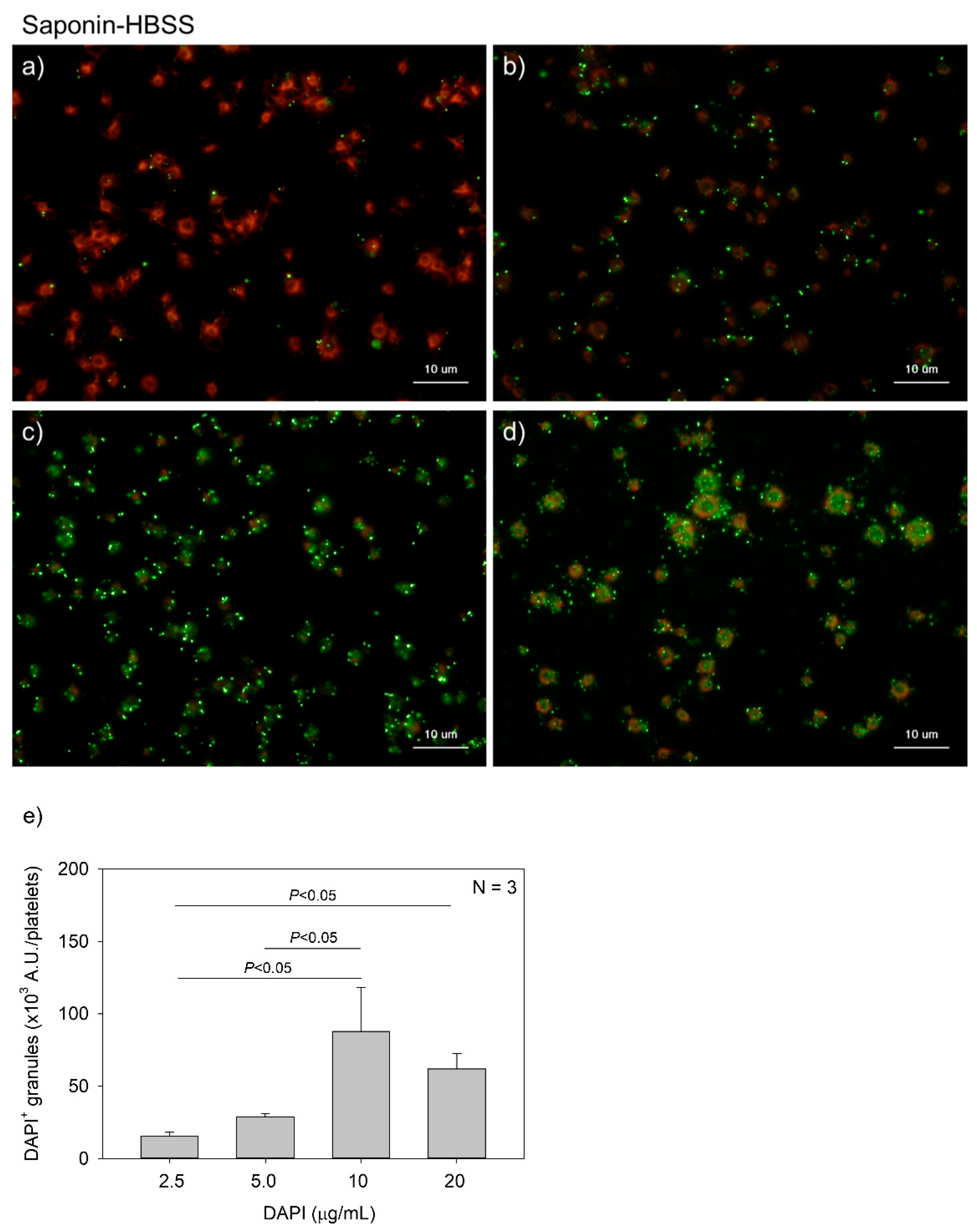
Figure 5.
Effects of 4′,6-diamidino-2-phenylindole (DAPI) concentrations on polyphosphate detection in platelets in 0.02% saponin-containing Hanks balanced salt solution (Spn-HBSS). Platelets immobilized on glass slides using a Cytospin were fixed in 10% neutral-buffered formalin for 4 h and then treated with DAPI at (a) 2.5, (b) 5.0, (c) 10, or (d) 20 μg/mL (green) and phalloidin (red) in Spn-HBSS and examined using a fluorescence microscope. (e) Quantitative analysis of the fluorescence images. Significant differences were obtained by a parametric test but not by a non-parametric test.
Figure 5.
Effects of 4′,6-diamidino-2-phenylindole (DAPI) concentrations on polyphosphate detection in platelets in 0.02% saponin-containing Hanks balanced salt solution (Spn-HBSS). Platelets immobilized on glass slides using a Cytospin were fixed in 10% neutral-buffered formalin for 4 h and then treated with DAPI at (a) 2.5, (b) 5.0, (c) 10, or (d) 20 μg/mL (green) and phalloidin (red) in Spn-HBSS and examined using a fluorescence microscope. (e) Quantitative analysis of the fluorescence images. Significant differences were obtained by a parametric test but not by a non-parametric test.
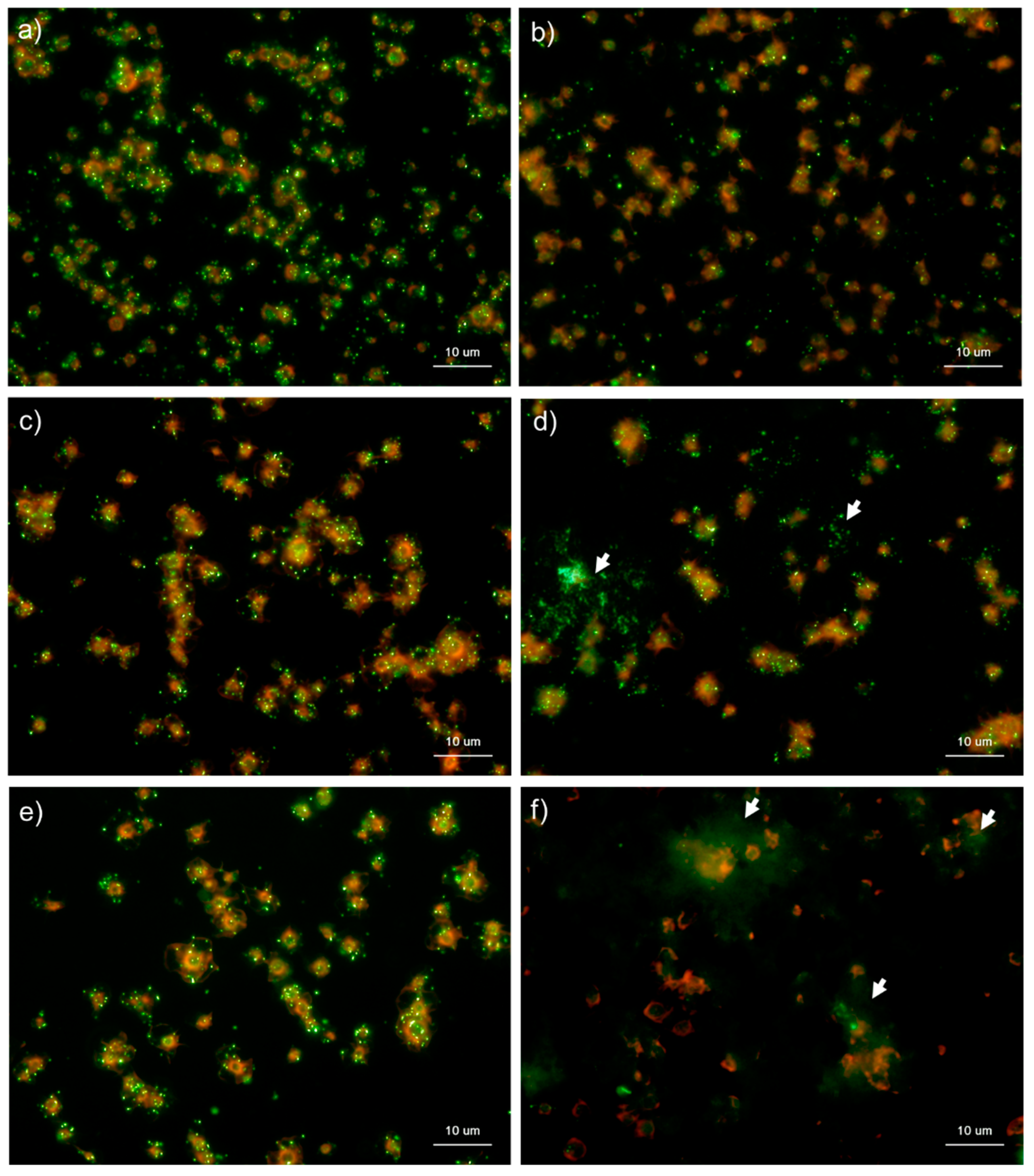
Figure 6.
Effects of CaCl2 on polyphosphate detection in platelets using 4′,6-diamidino-2-phenylindole (DAPI) in 0.1% Tween-20-containing Hanks balanced salt solution (Tw-HBSS). Platelets suspended in phosphate-buffered saline were activated by 0.1% CaCl2 for (b) 15 min, (d) 30 min, or (f) 60 min and then immobilized on glass slides using a Cytospin. Controls (no addition) are shown at (a) 15 min, (c) 30 min, and (e) 60 min. After 4 h of fixation with 10% neutral-buffered formalin, platelets were treated with DAPI (green) and phalloidin (red) in Tw-HBSS and examined using a fluorescence microscope. Arrows indicate possibly diffused polyP. Quantitative analysis could not be performed owing to the aggregation of platelets and diffuse polyP localization.
Figure 6.
Effects of CaCl2 on polyphosphate detection in platelets using 4′,6-diamidino-2-phenylindole (DAPI) in 0.1% Tween-20-containing Hanks balanced salt solution (Tw-HBSS). Platelets suspended in phosphate-buffered saline were activated by 0.1% CaCl2 for (b) 15 min, (d) 30 min, or (f) 60 min and then immobilized on glass slides using a Cytospin. Controls (no addition) are shown at (a) 15 min, (c) 30 min, and (e) 60 min. After 4 h of fixation with 10% neutral-buffered formalin, platelets were treated with DAPI (green) and phalloidin (red) in Tw-HBSS and examined using a fluorescence microscope. Arrows indicate possibly diffused polyP. Quantitative analysis could not be performed owing to the aggregation of platelets and diffuse polyP localization.
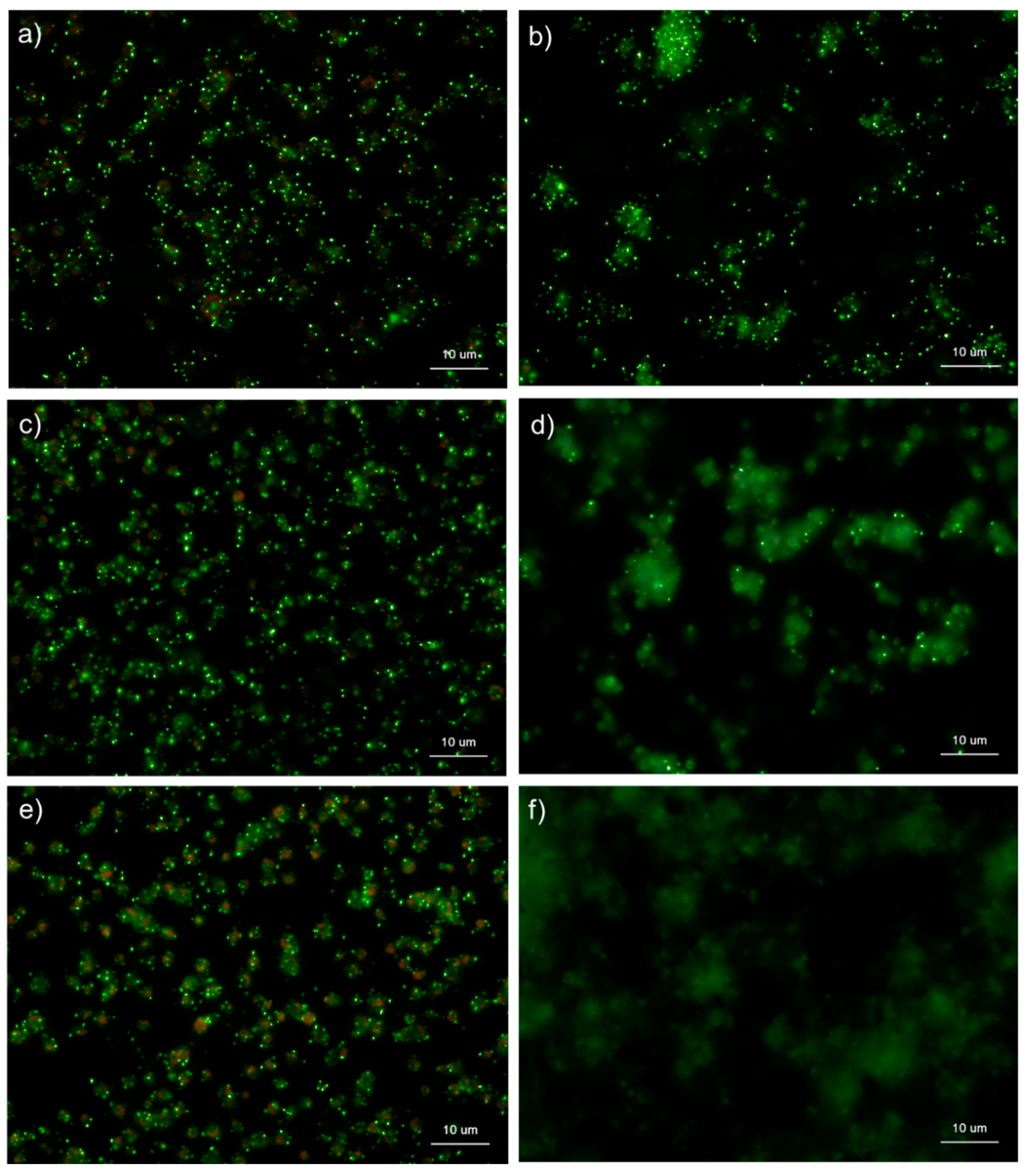
Figure 7.
Effects of CaCl2 on polyphosphate detection in platelets using 4′,6-diamidino-2-phenylindole (DAPI) in 0.02% saponin-containing Hanks balanced salt solution (Spn-HBSS). Platelets suspended in PBS were activated by 0.1% CaCl2 for (b) 15 min, (d) 30 min, or (f) 60 min then immobilized on glass sides using a Cytospin. Controls (no addition) are shown at (a) 15 min, (c) 30 min, and (e) 60 min. After 4 h of fixation with 10% neutral-buffered formalin, platelets were treated with DAPI (green) and phalloidin (red) in Spn-HBSS and examined using a fluorescence microscope. Quantitative analysis could not be performed owing to the aggregation of platelets and diffuse polyP localization.
Figure 7.
Effects of CaCl2 on polyphosphate detection in platelets using 4′,6-diamidino-2-phenylindole (DAPI) in 0.02% saponin-containing Hanks balanced salt solution (Spn-HBSS). Platelets suspended in PBS were activated by 0.1% CaCl2 for (b) 15 min, (d) 30 min, or (f) 60 min then immobilized on glass sides using a Cytospin. Controls (no addition) are shown at (a) 15 min, (c) 30 min, and (e) 60 min. After 4 h of fixation with 10% neutral-buffered formalin, platelets were treated with DAPI (green) and phalloidin (red) in Spn-HBSS and examined using a fluorescence microscope. Quantitative analysis could not be performed owing to the aggregation of platelets and diffuse polyP localization.
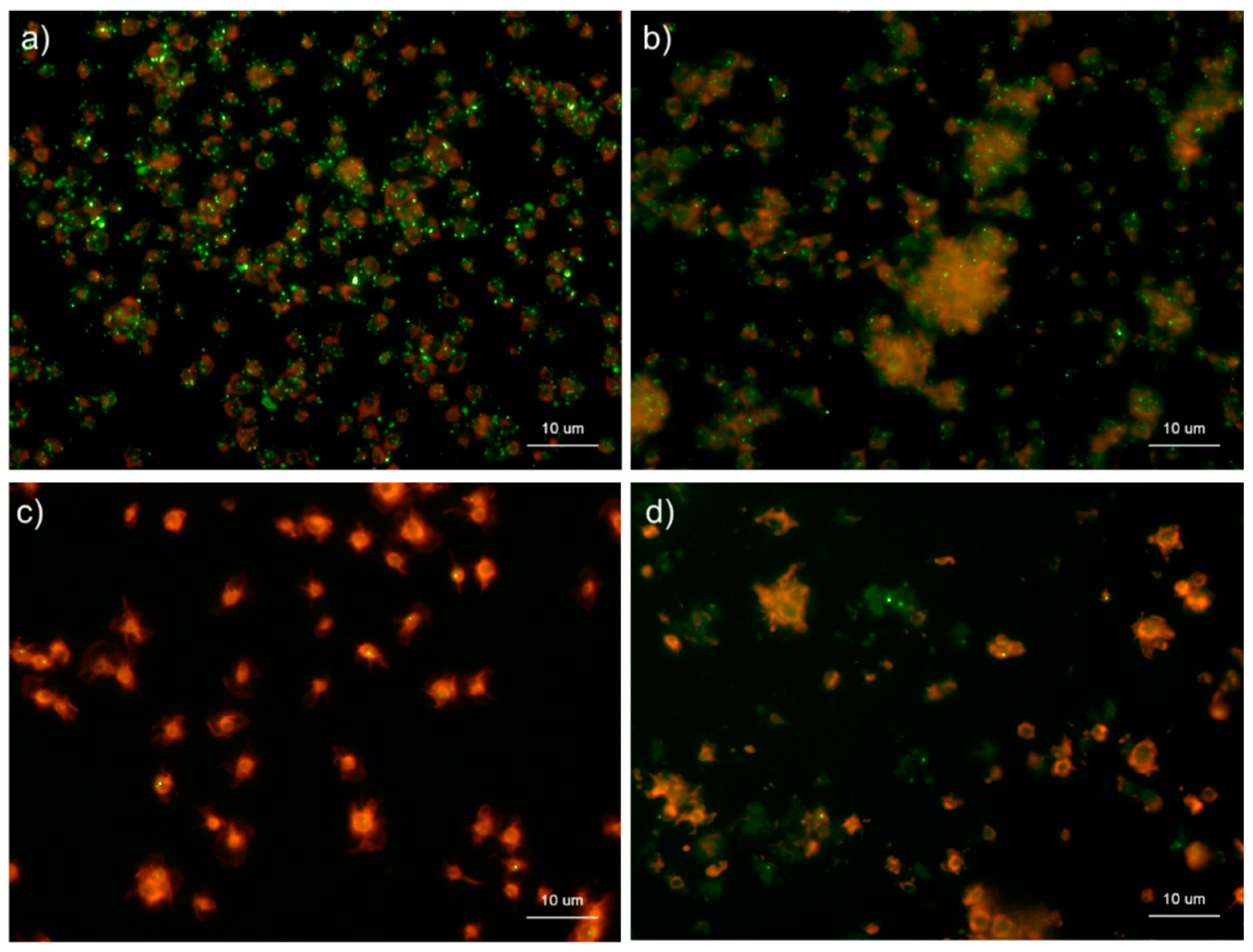
Figure 8.
Effects of fixative types on polyphosphate detection in platelets. Platelets suspended in phosphate-buffered saline were activated by 0.1% CaCl2 for 60 min (b,d), immobilized on glass sides using a Cytospin, and fixed with (a) 10% neutral-buffered formalin or (b) ThromboFix, which was developed and optimized for platelet fixation for the examination of surface markers by flow cytometric analysis. Controls (no addition) are shown in (a) 10% neutral-buffered formalin and (c) ThromboFix. Platelets were treated with 4′,6-diamidino-2-phenylindole (green) and phalloidin (red) in detergent-free Hanks balanced salt solution and examined using a fluorescence microscope. Quantitative analysis could not be performed owing to the aggregation of platelets and diffuse polyP localization.
Figure 8.
Effects of fixative types on polyphosphate detection in platelets. Platelets suspended in phosphate-buffered saline were activated by 0.1% CaCl2 for 60 min (b,d), immobilized on glass sides using a Cytospin, and fixed with (a) 10% neutral-buffered formalin or (b) ThromboFix, which was developed and optimized for platelet fixation for the examination of surface markers by flow cytometric analysis. Controls (no addition) are shown in (a) 10% neutral-buffered formalin and (c) ThromboFix. Platelets were treated with 4′,6-diamidino-2-phenylindole (green) and phalloidin (red) in detergent-free Hanks balanced salt solution and examined using a fluorescence microscope. Quantitative analysis could not be performed owing to the aggregation of platelets and diffuse polyP localization.
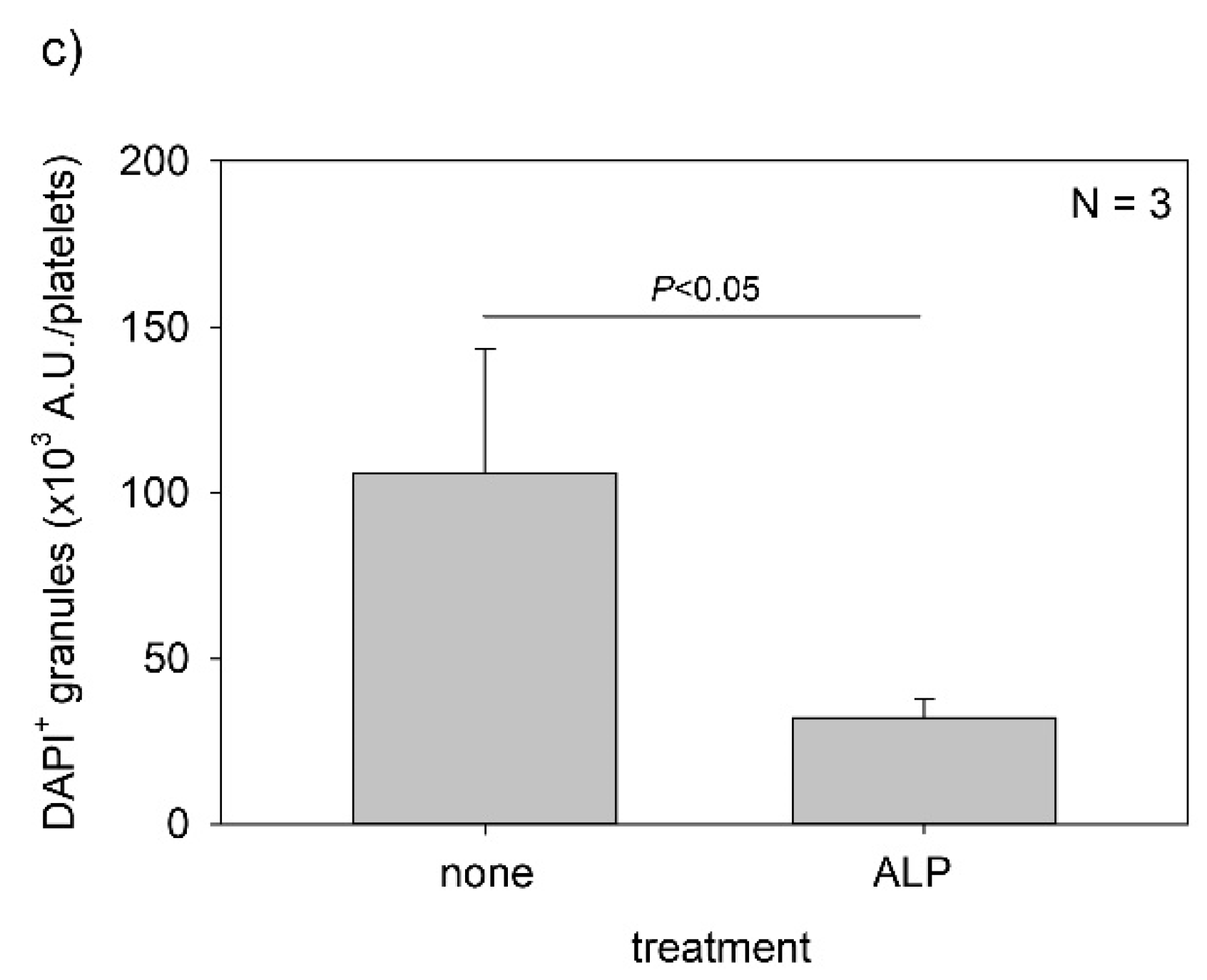
Figure 9.
Effect of alkaline phosphatase (ALP) on polyphosphate detection in platelets. Platelets were immobilized on glass sides using a Cytospin. After 4 h of fixation with 10% neutral-buffered formalin, platelets were treated with (b) 0.04 U/μL ALP in Mg2+-containing alkaline buffer for 24 h, then with 4′,6-diamidino-2-phenylindole (green) and phalloidin (red) in 0.1% Tween-20-containing Hanks balanced salt solution and examined using a fluorescence microscope. The control (no ALP treatment) is shown in (a). (c) Quantitative analysis of the fluorescence images. Significant differences were observed by both a parametric test and a non-parametric test.
Figure 9.
Effect of alkaline phosphatase (ALP) on polyphosphate detection in platelets. Platelets were immobilized on glass sides using a Cytospin. After 4 h of fixation with 10% neutral-buffered formalin, platelets were treated with (b) 0.04 U/μL ALP in Mg2+-containing alkaline buffer for 24 h, then with 4′,6-diamidino-2-phenylindole (green) and phalloidin (red) in 0.1% Tween-20-containing Hanks balanced salt solution and examined using a fluorescence microscope. The control (no ALP treatment) is shown in (a). (c) Quantitative analysis of the fluorescence images. Significant differences were observed by both a parametric test and a non-parametric test.
Figure 10.
Effects of CaCl2 on polyphosphate and serotonin distribution in platelets. Platelets suspended in phosphate-buffered saline (PBS) were activated by 0.1% CaCl2 for 60 min (b,d) then immobilized on glass sides using a Cytospin. After 4 h of fixation with 10% neutral-buffered formalin, platelets were subjected to (b) 4′,6-diamidino-2-phenylindole (green) or phalloidin (red) in 0.1% Tween-20-containing Hanks balanced salt solution or (d) immunocytochemical examination for serotonin using BlockAce-containing 0.1% Tween-20-containing PBS. The controls (no addition) are shown in (a) polyP and (c) serotonin. Quantitative analysis could not be performed owing to the aggregation of platelets and diffuse polyP localization.
Figure 10.
Effects of CaCl2 on polyphosphate and serotonin distribution in platelets. Platelets suspended in phosphate-buffered saline (PBS) were activated by 0.1% CaCl2 for 60 min (b,d) then immobilized on glass sides using a Cytospin. After 4 h of fixation with 10% neutral-buffered formalin, platelets were subjected to (b) 4′,6-diamidino-2-phenylindole (green) or phalloidin (red) in 0.1% Tween-20-containing Hanks balanced salt solution or (d) immunocytochemical examination for serotonin using BlockAce-containing 0.1% Tween-20-containing PBS. The controls (no addition) are shown in (a) polyP and (c) serotonin. Quantitative analysis could not be performed owing to the aggregation of platelets and diffuse polyP localization.
Figure 11.
Effects of ADP on the distribution of polyphosphate and serotonin in platelets. Platelets suspended in phosphate-buffered saline (PBS) were activated by 5 mM of ADP for 60 min then immobilized on glass slides using a Cytospin. After 4 h of fixation with 10% neutral-buffered formalin, platelets were subjected to (b) 4′,6-diamidino-2-phenylindole (green) or phalloidin (red) in 0.1% Tween-20-containing Hanks balanced salt solution or (d) immunocytochemical examination for serotonin using BlockAce-containing 0.1% Tween-20-containing PBS. The controls (no addition) are shown in (a) polyP and (c) serotonin. Quantitative analysis could not be performed owing to the aggregation of platelets and diffuse polyP localization.
Figure 11.
Effects of ADP on the distribution of polyphosphate and serotonin in platelets. Platelets suspended in phosphate-buffered saline (PBS) were activated by 5 mM of ADP for 60 min then immobilized on glass slides using a Cytospin. After 4 h of fixation with 10% neutral-buffered formalin, platelets were subjected to (b) 4′,6-diamidino-2-phenylindole (green) or phalloidin (red) in 0.1% Tween-20-containing Hanks balanced salt solution or (d) immunocytochemical examination for serotonin using BlockAce-containing 0.1% Tween-20-containing PBS. The controls (no addition) are shown in (a) polyP and (c) serotonin. Quantitative analysis could not be performed owing to the aggregation of platelets and diffuse polyP localization.

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}











