
Transient Hyperglycemia and Hypoxia Induce Memory Effects in AngiomiR Expression Profiles of Feto-Placental Endothelial Cells


Abstract
:1. Introduction
2. Results
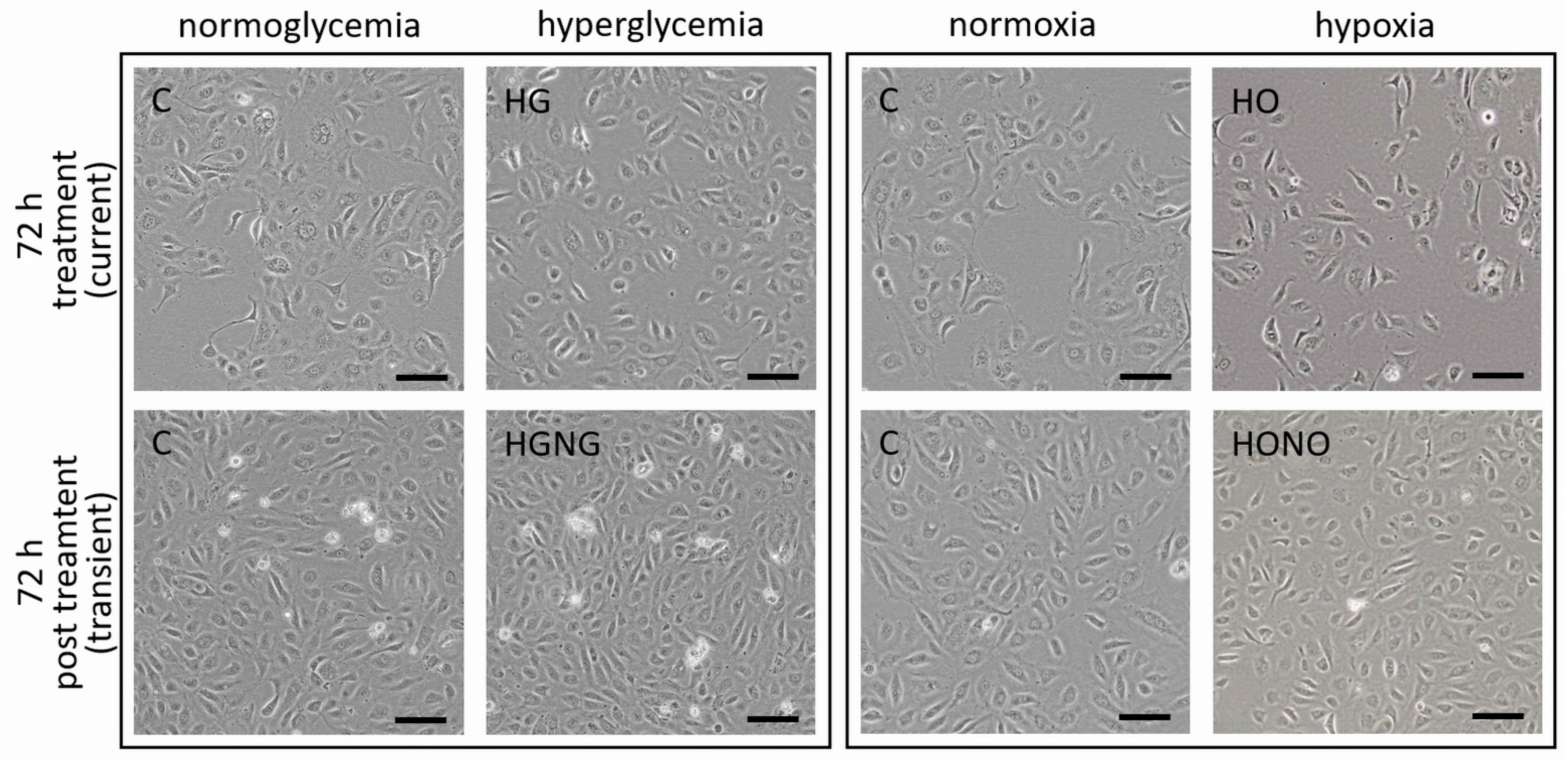
2.1. Hyperglycemia and Hypoxia Did Not Alter Microscopical Growing Pattern of fpEC
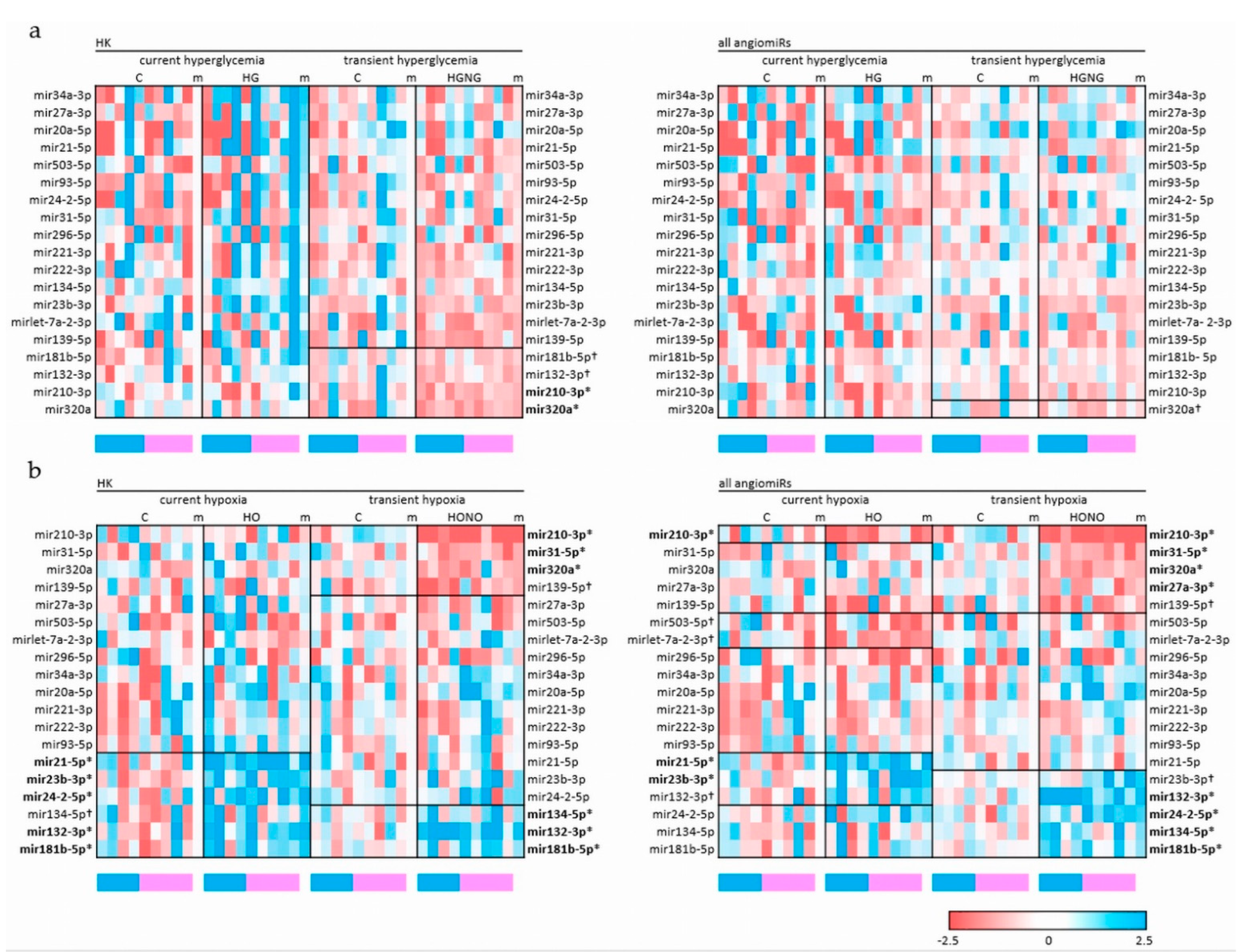
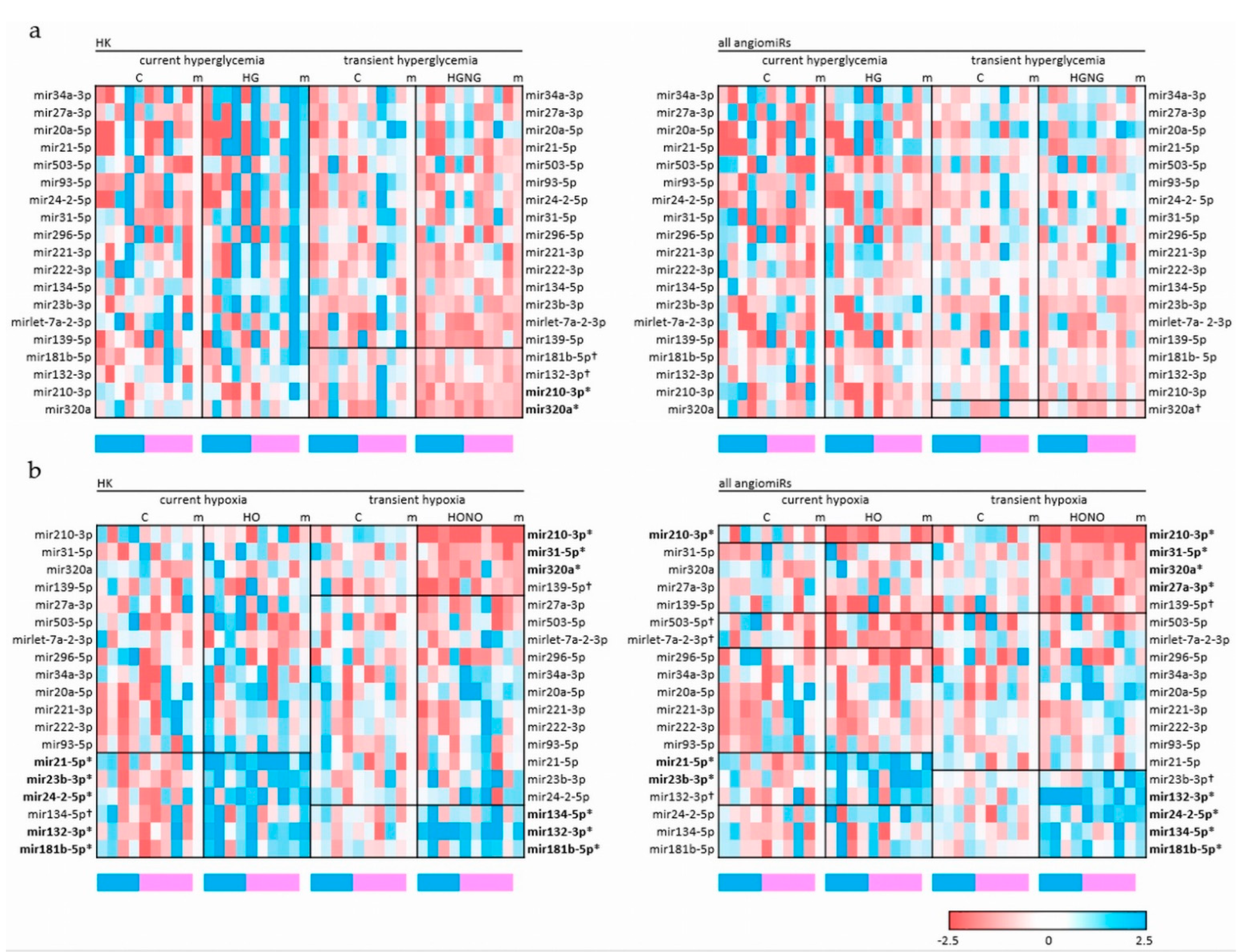
2.2. Current and Transient Effects of Hyperglycemia and Hypoxia on AngiomiRs
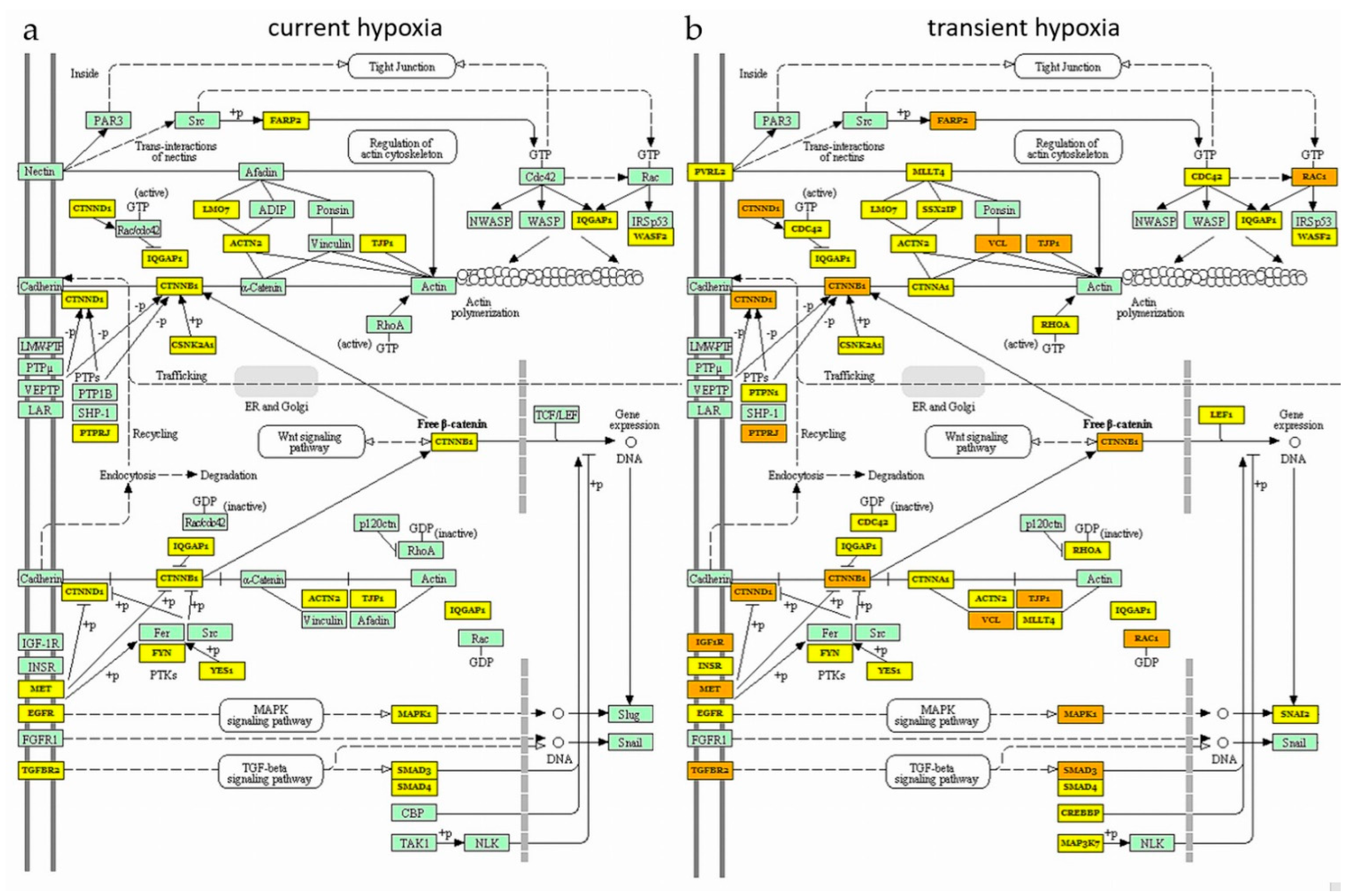
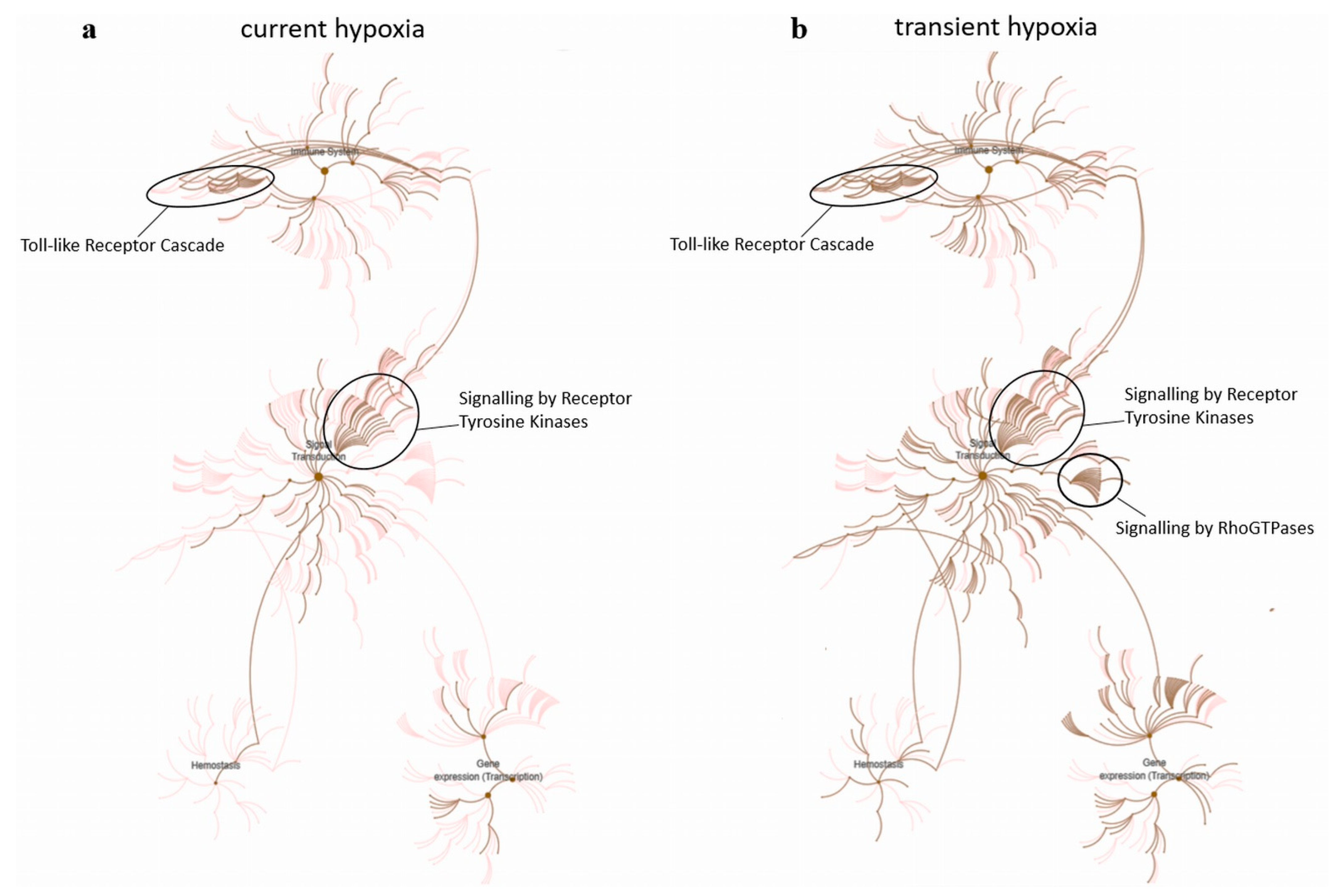
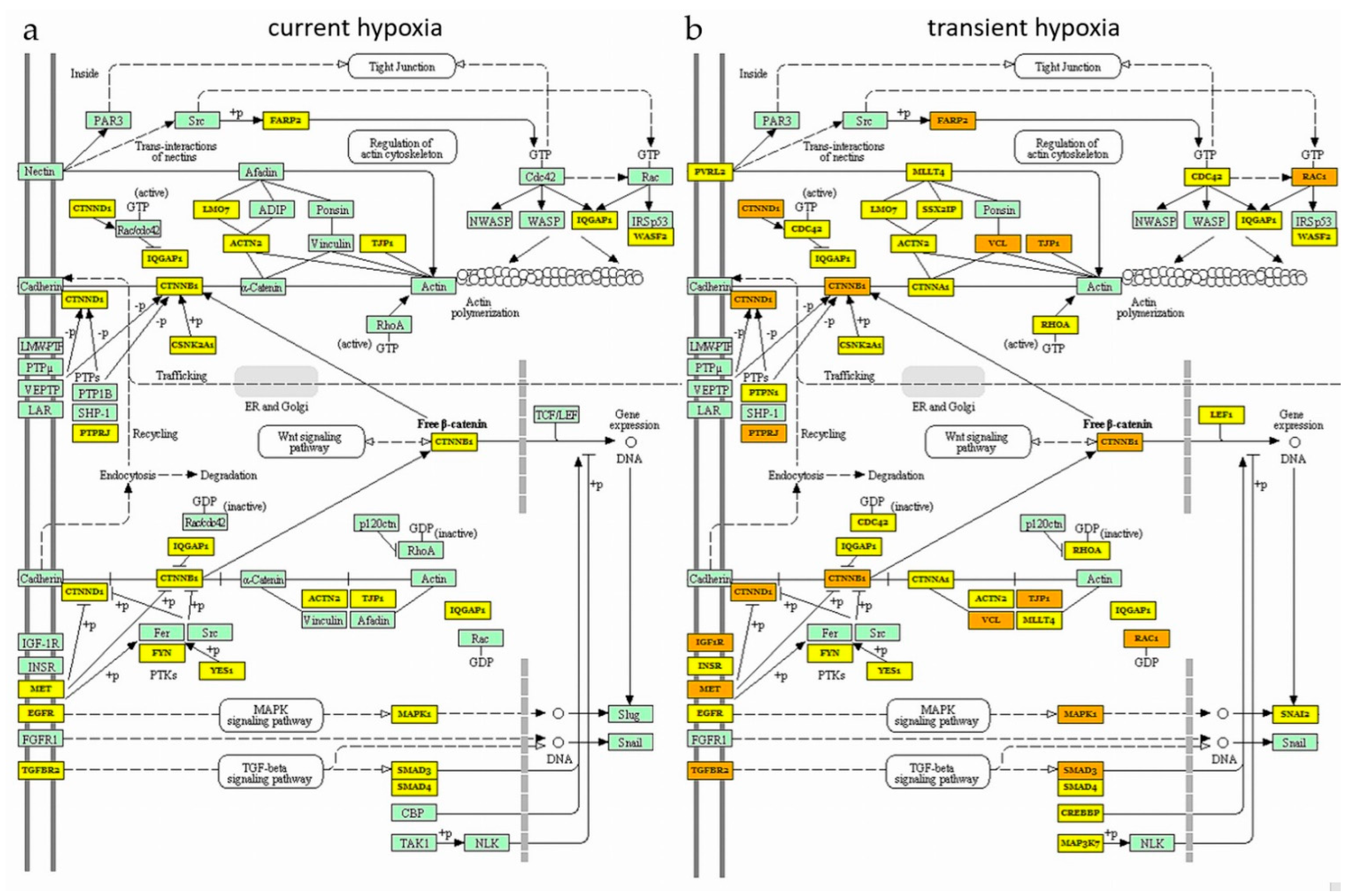
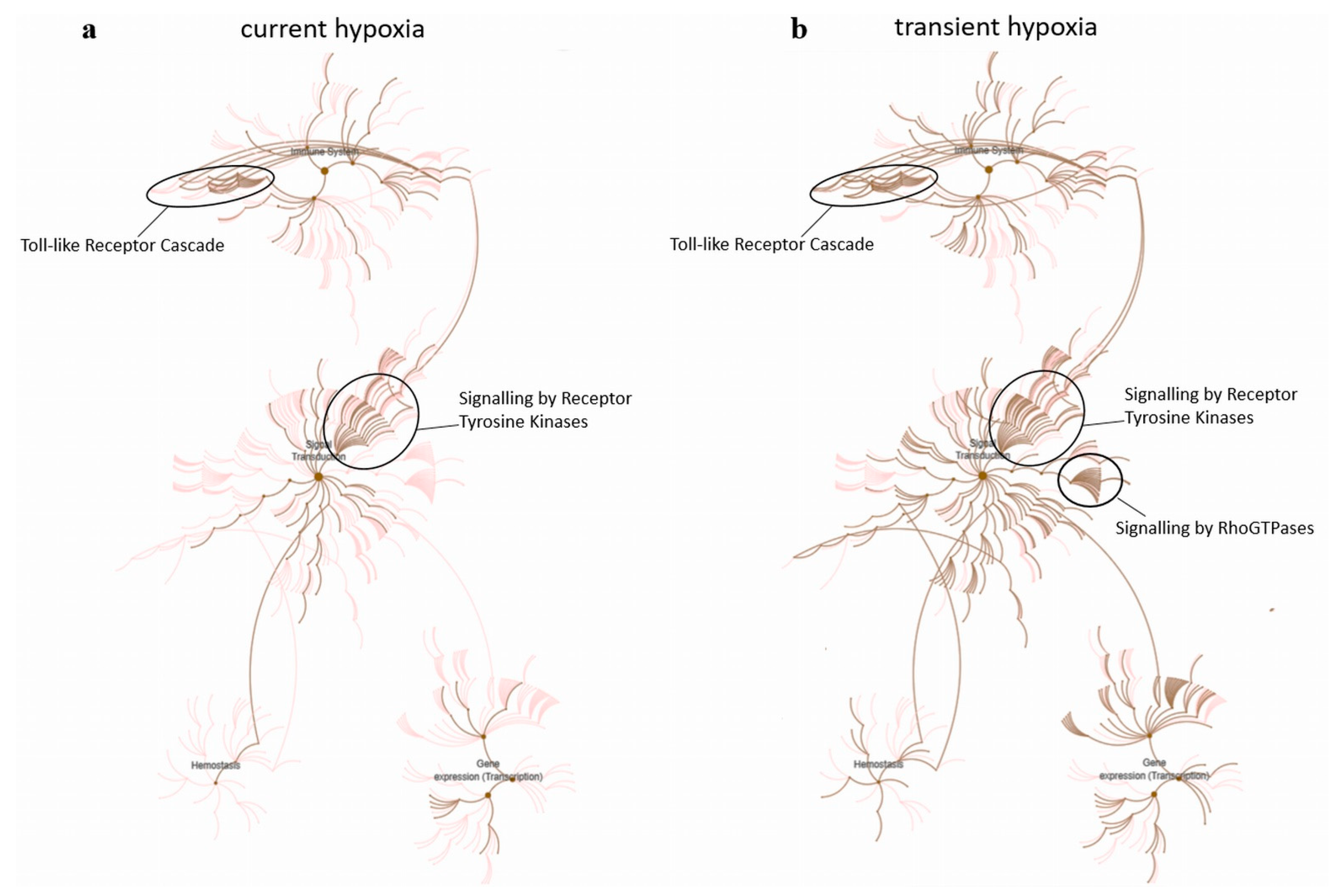
2.3. Current and Transient Effects of Hypoxia on AngiomiR-Regulated Functional Pathways
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Isolation of Human Feto-Placental Endothelial Cells (fpEC)
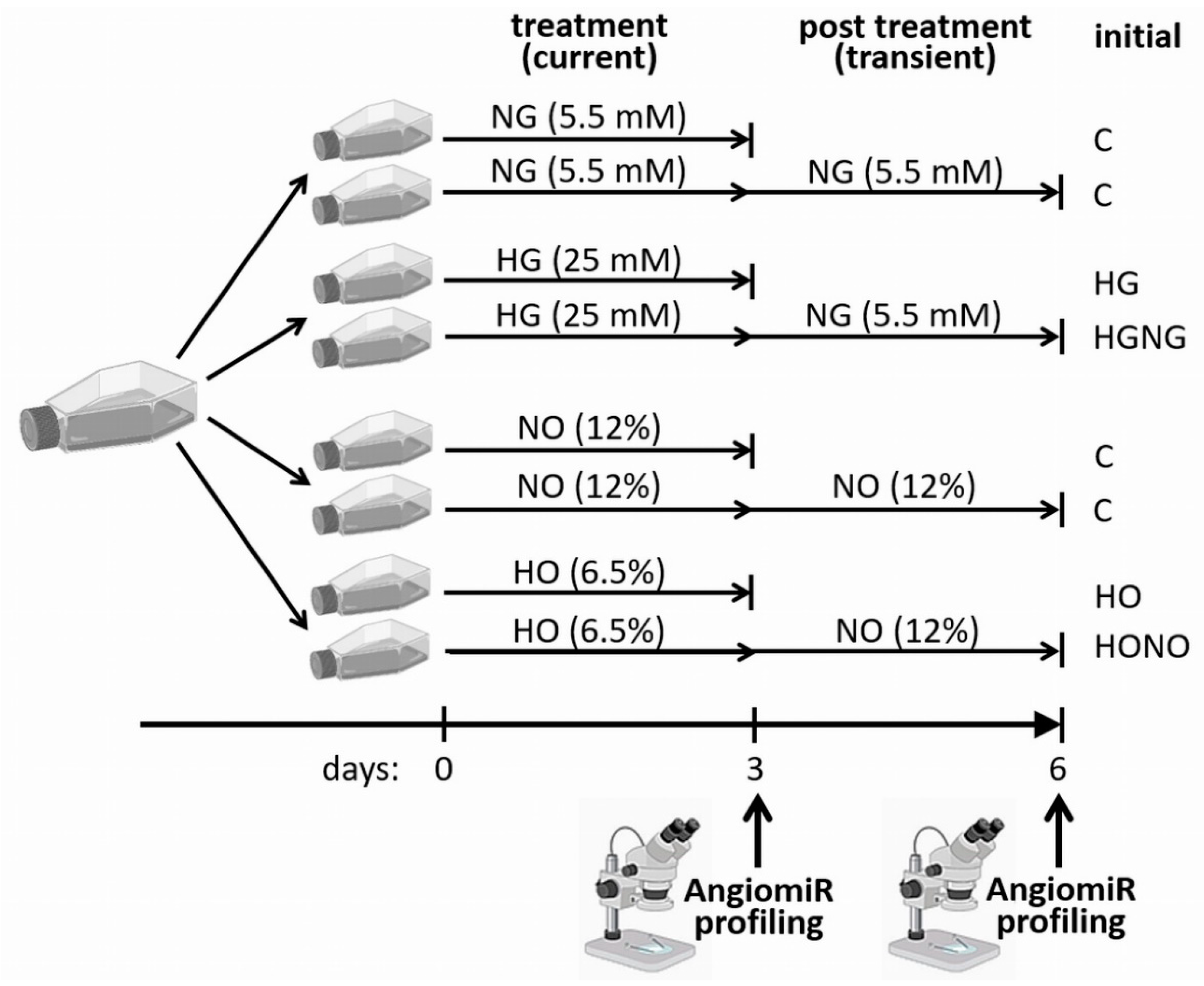
4.3. Treatments
4.4. Cell Imaging
4.5. MiRNA Isolation and cDNA Synthesis
4.6. RT-qPCR for AngiomiR Expression
4.7. Pathway Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tisi, D.K.; Burns, D.H.; Luskey, G.W.; Koski, K.G. Fetal exposure to altered amniotic fluid glucose, insulin, and insulin-like growth factor-binding protein 1 occurs before screening for gestational diabetes mellitus. Diabetes Care 2011, 34, 139–144. [Google Scholar] [CrossRef] [Green Version]
- Escobar, J.; Teramo, K.; Stefanovic, V.; Andersson, S.; Asensi, M.A.; Arduini, A.; Cubells, E.; Sastre, J.; Vento, M. Amniotic fluid oxidative and nitrosative stress biomarkers correlate with fetal chronic hypoxia in diabetic pregnancies. Neonatology 2013, 103, 193–198. [Google Scholar] [CrossRef]
- Taricco, E.; Radaelli, T.; Rossi, G.; Nobile de Santis, M.S.; Bulfamante, G.P.; Avagliano, L.; Cetin, I. Effects of gestational diabetes on fetal oxygen and glucose levels in vivo. BJOG 2009, 116, 1729–1735. [Google Scholar] [CrossRef]
- Teramo, K.A.; Hiilesmaa, V.K.; Schwartz, R.; Clemons, G.K.; Widness, J.A. Amniotic fluid and cord plasma erythropoietin levels in pregnancies complicated by preeclampsia, pregnancy-induced hypertension and chronic hypertension. J. Perinat. Med. 2004, 32, 240–247. [Google Scholar] [CrossRef]
- Salafia, C.M.; Minior, V.K.; Lopez-Zeno, J.A.; Whittington, S.S.; Pezzullo, J.C.; Vintzileos, A.M. Relationship between placental histologic features and umbilical cord blood gases in preterm gestations. Am. J. Obstet. Gynecol. 1995, 173, 1058–1064. [Google Scholar] [CrossRef]
- Giacco, F.; Brownlee, M. Oxidative stress and diabetic complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef] [Green Version]
- Bakker, W.; Eringa, E.C.; Sipkema, P.; van Hinsbergh, V.W. Endothelial dysfunction and diabetes: Roles of hyperglycemia, impaired insulin signaling and obesity. Cell Tissue Res. 2009, 335, 165–189. [Google Scholar] [CrossRef] [Green Version]
- Mochol, J.; Gawrys, J.; Gajecki, D.; Szahidewicz-Krupska, E.; Martynowicz, H.; Doroszko, A. Cardiovascular Disorders Triggered by Obstructive Sleep Apnea-A Focus on Endothelium and Blood Components. Int. J. Mol. Sci. 2021, 22, 5139. [Google Scholar] [CrossRef]
- Song, Y.; Xing, H.; He, Y.; Zhang, Z.; Shi, G.; Wu, S.; Liu, Y.; Harrington, E.O.; Sellke, F.W.; Feng, J. Inhibition of mitochondrial reactive oxygen species improves coronary endothelial function after cardioplegic hypoxia/reoxygenation. J. Thorac. Cardiovasc. Surg. 2021. [Google Scholar] [CrossRef]
- Frost, A.L.; Suriano, K.; Aye, C.Y.L.; Leeson, P.; Lewandowski, A.J. The Immediate and Long-Term Impact of Preeclampsia on Offspring Vascular and Cardiac Physiology in the Preterm Infant. Front. Pediatr. 2021, 9, 625726. [Google Scholar] [CrossRef] [PubMed]
- Sallam, N.A.; Palmgren, V.A.C.; Singh, R.D.; John, C.M.; Thompson, J.A. Programming of Vascular Dysfunction in the Intrauterine Milieu of Diabetic Pregnancies. Int. J. Mol. Sci. 2018, 19, 3665. [Google Scholar] [CrossRef] [Green Version]
- Cvitic, S.; Novakovic, B.; Gordon, L.; Ulz, C.M.; Muhlberger, M.; Diaz-Perez, F.I.; Joo, J.E.; Svendova, V.; Schimek, M.G.; Trajanoski, S.; et al. Human fetoplacental arterial and venous endothelial cells are differentially programmed by gestational diabetes mellitus, resulting in cell-specific barrier function changes. Diabetologia 2018, 61, 2398–2411. [Google Scholar] [CrossRef] [Green Version]
- Brodowski, L.; Zindler, T.; von Hardenberg, S.; Schroder-Heurich, B.; von Kaisenberg, C.S.; Frieling, H.; Hubel, C.A.; Dork, T.; von Versen-Hoynck, F. Preeclampsia-Associated Alteration of DNA Methylation in Fetal Endothelial Progenitor Cells. Front. Cell Dev. Biol. 2019, 7, 32. [Google Scholar] [CrossRef] [Green Version]
- King, G.L.; Kunisaki, M.; Nishio, Y.; Inoguchi, T.; Shiba, T.; Xia, P. Biochemical and molecular mechanisms in the development of diabetic vascular complications. Diabetes 1996, 45 (Suppl. S3), S105–S108. [Google Scholar] [CrossRef]
- Fioretto, P.; Kim, Y.; Mauer, M. Diabetic nephropathy as a model of reversibility of established renal lesions. Curr. Opin. Nephrol. Hypertens. 1998, 7, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Pepin, M.E.; Schiano, C.; Miceli, M.; Benincasa, G.; Mansueto, G.; Grimaldi, V.; Soricelli, A.; Wende, A.R.; Napoli, C. The human aortic endothelium undergoes dose-dependent DNA methylation in response to transient hyperglycemia. Exp. Cell Res. 2021, 400, 112485. [Google Scholar] [CrossRef]
- Paneni, F.; Mocharla, P.; Akhmedov, A.; Costantino, S.; Osto, E.; Volpe, M.; Luscher, T.F.; Cosentino, F. Gene silencing of the mitochondrial adaptor p66(Shc) suppresses vascular hyperglycemic memory in diabetes. Circ. Res. 2012, 111, 278–289. [Google Scholar] [CrossRef] [Green Version]
- El-Osta, A.; Brasacchio, D.; Yao, D.; Pocai, A.; Jones, P.L.; Roeder, R.G.; Cooper, M.E.; Brownlee, M. Transient high glucose causes persistent epigenetic changes and altered gene expression during subsequent normoglycemia. J. Exp. Med. 2008, 205, 2409–2417. [Google Scholar] [CrossRef]
- Brasacchio, D.; Okabe, J.; Tikellis, C.; Balcerczyk, A.; George, P.; Baker, E.K.; Calkin, A.C.; Brownlee, M.; Cooper, M.E.; El-Osta, A. Hyperglycemia induces a dynamic cooperativity of histone methylase and demethylase enzymes associated with gene-activating epigenetic marks that coexist on the lysine tail. Diabetes 2009, 58, 1229–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nangaku, M.; Hirakawa, Y.; Mimura, I.; Inagi, R.; Tanaka, T. Epigenetic Changes in the Acute Kidney Injury-to-Chronic Kidney Disease Transition. Nephron 2017, 137, 256–259. [Google Scholar] [CrossRef]
- Saxena, K.; Jolly, M.K. Acute vs. Chronic vs. Cyclic Hypoxia: Their Differential Dynamics, Molecular Mechanisms, and Effects on Tumor Progression. Biomolecules 2019, 9, 339. [Google Scholar] [CrossRef] [Green Version]
- Hartel, F.V.; Holl, M.; Arshad, M.; Aslam, M.; Gunduz, D.; Weyand, M.; Micoogullari, M.; Abdallah, Y.; Piper, H.M.; Noll, T. Transient hypoxia induces ERK-dependent anti-apoptotic cell survival in endothelial cells. Am. J. Physiol. Cell Physiol. 2010, 298, C1501–C1509. [Google Scholar] [CrossRef] [Green Version]
- Liu, O.H.; Kiema, M.; Beter, M.; Yla-Herttuala, S.; Laakkonen, J.P.; Kaikkonen, M.U. Hypoxia-Mediated Regulation of Histone Demethylases Affects Angiogenesis-Associated Functions in Endothelial Cells. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 2665–2677. [Google Scholar] [CrossRef]
- Gulyaeva, L.F.; Kushlinskiy, N.E. Regulatory mechanisms of microRNA expression. J. Transl. Med. 2016, 14, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, F.; Yang, Z.; Li, G. Role of specific microRNAs for endothelial function and angiogenesis. Biochem. Biophys. Res. Commun. 2009, 386, 549–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Olson, E.N. AngiomiRs--key regulators of angiogenesis. Curr. Opin. Genet. Dev. 2009, 19, 205–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monticelli, S.; Natoli, G. Short-term memory of danger signals and environmental stimuli in immune cells. Nat. Immunol. 2013, 14, 777–784. [Google Scholar] [CrossRef]
- Costantino, S.; Paneni, F.; Luscher, T.F.; Cosentino, F. MicroRNA profiling unveils hyperglycaemic memory in the diabetic heart. Eur. Heart J. 2016, 37, 572–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, X.; Liao, Y.; Chen, L.; Liu, G.; Feng, Y.; Zeng, T.; Zhang, J. The MicroRNAs in the Pathogenesis of Metabolic Memory. Endocrinology 2015, 156, 3157–3168. [Google Scholar] [CrossRef]
- Yuan, S.Y.; Breslin, J.W.; Perrin, R.; Gaudreault, N.; Guo, M.; Kargozaran, H.; Wu, M.H. Microvascular permeability in diabetes and insulin resistance. Microcirculation 2007, 14, 363–373. [Google Scholar] [CrossRef]
- Simionescu, M. Implications of early structural-functional changes in the endothelium for vascular disease. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 266–274. [Google Scholar] [CrossRef]
- Liu, R.; Guan, S.; Gao, Z.; Wang, J.; Xu, J.; Hao, Z.; Zhang, Y.; Yang, S.; Guo, Z.; Yang, J.; et al. Pathological Hyperinsulinemia and Hyperglycemia in the Impaired Glucose Tolerance Stage Mediate Endothelial Dysfunction Through miR-21, PTEN/AKT/eNOS, and MARK/ET-1 Pathways. Front. Endocrinol. 2021, 12, 644159. [Google Scholar] [CrossRef] [PubMed]
- Rask-Madsen, C.; King, G.L. Vascular complications of diabetes: Mechanisms of injury and protective factors. Cell Metab. 2013, 17, 20–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flynn, M.C.; Kraakman, M.J.; Tikellis, C.; Lee, M.K.S.; Hanssen, N.M.J.; Kammoun, H.L.; Pickering, R.J.; Dragoljevic, D.; Al-Sharea, A.; Barrett, T.J.; et al. Transient Intermittent Hyperglycemia Accelerates Atherosclerosis by Promoting Myelopoiesis. Circ. Res. 2020, 127, 877–892. [Google Scholar] [CrossRef]
- Nallamshetty, S.; Chan, S.Y.; Loscalzo, J. Hypoxia: A master regulator of microRNA biogenesis and activity. Free Radic. Biol. Med. 2013, 64, 20–30. [Google Scholar] [CrossRef] [Green Version]
- Bertero, T.; Rezzonico, R.; Pottier, N.; Mari, B. Impact of MicroRNAs in the Cellular Response to Hypoxia. Int. Rev. Cell Mol. Biol. 2017, 333, 91–158. [Google Scholar] [CrossRef]
- Greco, S.; Martelli, F. MicroRNAs in Hypoxia Response. Antioxid. Redox Signal. 2014, 21, 1164–1166. [Google Scholar] [CrossRef] [Green Version]
- Yao, C.; Shi, X.; Zhang, Z.; Zhou, S.; Qian, T.; Wang, Y.; Ding, F.; Gu, X.; Yu, B. Hypoxia-Induced Upregulation of miR-132 Promotes Schwann Cell Migration After Sciatic Nerve Injury by Targeting PRKAG3. Mol. Neurobiol. 2016, 53, 5129–5139. [Google Scholar] [CrossRef]
- Zhou, C.H.; Zhang, X.P.; Liu, F.; Wang, W. Modeling the interplay between the HIF-1 and p53 pathways in hypoxia. Sci. Rep. 2015, 5, 13834. [Google Scholar] [CrossRef] [Green Version]
- Pranzini, E.; Leo, A.; Rapizzi, E.; Ramazzotti, M.; Magherini, F.; Giovannelli, L.; Caselli, A.; Cirri, P.; Taddei, M.L.; Paoli, P. miR-210-3p mediates metabolic adaptation and sustains DNA damage repair of resistant colon cancer cells to treatment with 5-fluorouracil. Mol. Carcinog. 2019, 58, 2181–2192. [Google Scholar] [CrossRef]
- Prado, M.S.G.; de Jesus, M.L.; de Goes, T.C.; Mendonca, L.S.O.; Kaneto, C.M. Downregulation of circulating miR-320a and target gene prediction in patients with diabetic retinopathy. BMC Res. Notes 2020, 13, 155. [Google Scholar] [CrossRef]
- Jo, S.; Xu, G.; Jing, G.; Chen, J.; Shalev, A. Human Glucagon Expression Is under the Control of miR-320a. Endocrinology 2021, 162, bqaa238. [Google Scholar] [CrossRef]
- Strutz, J.; Cvitic, S.; Hackl, H.; Kashofer, K.; Appel, H.M.; Thuringer, A.; Desoye, G.; Koolwijk, P.; Hiden, U. Gestational diabetes alters microRNA signatures in human feto-placental endothelial cells depending on fetal sex. Clin. Sci. 2018, 132, 2437–2449. [Google Scholar] [CrossRef]
- He, L.; Wang, X.; Jin, Y.; Xu, W.; Guan, Y.; Wu, J.; Han, S.; Liu, G. Identification and validation of the miRNA-mRNA regulatory network in fetoplacental arterial endothelial cells of gestational diabetes mellitus. Bioengineered 2021, 12, 3503–3515. [Google Scholar] [CrossRef]
- Humphries, D.E.; Lee, S.L.; Fanburg, B.L.; Silbert, J.E. Effects of hypoxia and hyperoxia on proteoglycan production by bovine pulmonary artery endothelial cells. J. Cell. Physiol. 1986, 126, 249–253. [Google Scholar] [CrossRef]
- Aslam, M.; Schluter, K.D.; Rohrbach, S.; Rafiq, A.; Nazli, S.; Piper, H.M.; Noll, T.; Schulz, R.; Gunduz, D. Hypoxia-reoxygenation-induced endothelial barrier failure: Role of RhoA, Rac1 and myosin light chain kinase. J. Physiol. 2013, 591, 461–473. [Google Scholar] [CrossRef]
- Wojciak-Stothard, B.; Tsang, L.Y.; Haworth, S.G. Rac and Rho play opposing roles in the regulation of hypoxia/reoxygenation-induced permeability changes in pulmonary artery endothelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2005, 288, L749–L760. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbach, H.; da Silva, A.M.; Calin, G.; Pantel, K. Data Normalization Strategies for MicroRNA Quantification. Clin. Chem. 2015, 61, 1333–1342. [Google Scholar] [CrossRef]
- Leopold, B.; Strutz, J.; Weiss, E.; Gindlhuber, J.; Birner-Gruenberger, R.; Hackl, H.; Appel, H.M.; Cvitic, S.; Hiden, U. Outgrowth, proliferation, viability, angiogenesis and phenotype of primary human endothelial cells in different purchasable endothelial culture media: Feed wisely. Histochem. Cell Biol. 2019, 152, 377–390. [Google Scholar] [CrossRef] [Green Version]
- Lang, I.; Schweizer, A.; Hiden, U.; Ghaffari-Tabrizi, N.; Hagendorfer, G.; Bilban, M.; Pabst, M.A.; Korgun, E.T.; Dohr, G.; Desoye, G. Human fetal placental endothelial cells have a mature arterial and a juvenile venous phenotype with adipogenic and osteogenic differentiation potential. Differentiation 2008, 76, 1031–1043. [Google Scholar] [CrossRef]
- Arikan, G.M.; Scholz, H.S.; Petru, E.; Haeusler, M.C.; Haas, J.; Weiss, P.A. Cord blood oxygen saturation in vigorous infants at birth: What is normal? BJOG 2000, 107, 987–994. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Babion, I.; Snoek, B.C.; van de Wiel, M.A.; Wilting, S.M.; Steenbergen, R.D.M. A Strategy to Find Suitable Reference Genes for miRNA Quantitative PCR Analysis and Its Application to Cervical Specimens. J. Mol. Diagn. 2017, 19, 625–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wotschofsky, Z.; Meyer, H.A.; Jung, M.; Fendler, A.; Wagner, I.; Stephan, C.; Busch, J.; Erbersdobler, A.; Disch, A.C.; Mollenkopf, H.J.; et al. Reference genes for the relative quantification of microRNAs in renal cell carcinomas and their metastases. Anal. Biochem. 2011, 417, 233–241. [Google Scholar] [CrossRef]
- Marabita, F.; de Candia, P.; Torri, A.; Tegner, J.; Abrignani, S.; Rossi, R.L. Normalization of circulating microRNA expression data obtained by quantitative real-time RT-PCR. Brief. Bioinform. 2016, 17, 204–212. [Google Scholar] [CrossRef] [Green Version]
- Rice, J.; Roberts, H.; Rai, S.N.; Galandiuk, S. Housekeeping genes for studies of plasma microRNA: A need for more precise standardization. Surgery 2015, 158, 1345–1351. [Google Scholar] [CrossRef]
- Hijmans, J.G.; Stockelman, K.; Levy, M.; Brewster, L.M.; Bammert, T.D.; Greiner, J.J.; Connick, E.; DeSouza, C.A. Effects of HIV-1 gp120 and TAT-derived microvesicles on endothelial cell function. J. Appl. Physiol. 2019, 126, 1242–1249. [Google Scholar] [CrossRef]
- Anand, S.; Majeti, B.K.; Acevedo, L.M.; Murphy, E.A.; Mukthavaram, R.; Scheppke, L.; Huang, M.; Shields, D.J.; Lindquist, J.N.; Lapinski, P.E.; et al. MicroRNA-132-mediated loss of p120RasGAP activates the endothelium to facilitate pathological angiogenesis. Nat. Med. 2010, 16, 909–914. [Google Scholar] [CrossRef]
- Wang, H.W.; Su, S.H.; Wang, Y.L.; Chang, S.T.; Liao, K.H.; Lo, H.H.; Chiu, Y.L.; Hsieh, T.H.; Huang, T.S.; Lin, C.S.; et al. MicroRNA-134 Contributes to Glucose-Induced Endothelial Cell Dysfunction and This Effect Can Be Reversed by Far-Infrared Irradiation. PLoS ONE 2016, 11, e0147067. [Google Scholar] [CrossRef]
- Walz, J.M.; Wecker, T.; Zhang, P.P.; Cakir, B.; Gruening, B.; Agostini, H.; Reuer, T.; Ludwig, F.; Boneva, S.; Faerber, L.; et al. Impact of angiogenic activation and inhibition on miRNA profiles of human retinal endothelial cells. Exp. Eye Res. 2019, 181, 98–104. [Google Scholar] [CrossRef]
- Wang, L.; Liu, W.X.; Huang, X.G. MicroRNA-199a-3p inhibits angiogenesis by targeting the VEGF/PI3K/AKT signalling pathway in an in vitro model of diabetic retinopathy. Exp. Mol. Pathol. 2020, 116, 104488. [Google Scholar] [CrossRef]
- Guo, Y.; Du, F.; Tan, Y.L.; Luo, J.; Xiong, D.; Song, W.T. VEGF-mediated angiogenesis in retinopathy of prematurity is co-regulated by miR-17-5p and miR-20a-5p. Biochem. Cell Biol. 2021, 99, 414–423. [Google Scholar] [CrossRef]
- Mirzaei Bavil, F.; Karimi-Sales, E.; Alihemmati, A.; Alipour, M.R. Effect of ghrelin on hypoxia-related cardiac angiogenesis: Involvement of miR-210 signalling pathway. Arch. Physiol. Biochem. 2019, 9, 1–6. [Google Scholar] [CrossRef]
- Penaloza, E.; Soto-Carrasco, G.; Krause, B.J. MiR-21-5p directly contributes to regulating eNOS expression in human artery endothelial cells under normoxia and hypoxia. Biochem. Pharmacol. 2020, 182, 114288. [Google Scholar] [CrossRef]
- Poliseno, L.; Tuccoli, A.; Mariani, L.; Evangelista, M.; Citti, L.; Woods, K.; Mercatanti, A.; Hammond, S.; Rainaldi, G. MicroRNAs modulate the angiogenic properties of HUVECs. Blood 2006, 108, 3068–3071. [Google Scholar] [CrossRef]
- Zhou, Q.; Gallagher, R.; Ufret-Vincenty, R.; Li, X.; Olson, E.N.; Wang, S. Regulation of angiogenesis and choroidal neovascularization by members of microRNA-23~27~24 clusters. Proc. Natl. Acad. Sci. USA 2011, 108, 8287–8292. [Google Scholar] [CrossRef] [Green Version]
- Jia, L.; Zhou, X.; Huang, X.; Xu, X.; Jia, Y.; Wu, Y.; Yao, J.; Wu, Y.; Wang, K. Maternal and umbilical cord serum-derived exosomes enhance endothelial cell proliferation and migration. FASEB J. 2018, 32, 4534–4543. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Yu, M.; Yin, W.; Liang, B.; Li, A.; Li, J.; Li, X.; Zhao, S.; Liu, F. Development of a novel RNAi therapy: Engineered miR-31 exosomes promoted the healing of diabetic wounds. Bioact. Mater. 2021, 6, 2841–2853. [Google Scholar] [CrossRef]
- Zampetaki, A.; Willeit, P.; Burr, S.; Yin, X.; Langley, S.R.; Kiechl, S.; Klein, R.; Rossing, P.; Chaturvedi, N.; Mayr, M. Angiogenic microRNAs Linked to Incidence and Progression of Diabetic Retinopathy in Type 1 Diabetes. Diabetes 2016, 65, 216–227. [Google Scholar] [CrossRef] [Green Version]
- Popov, T.M.; Giragosyan, S.; Petkova, V.; Stancheva, G.; Marinov, T.; Belitova, M.; Rangachev, J.; Popova, D.; Kaneva, R.P. Proangiogenic signature in advanced laryngeal carcinoma after microRNA expression profiling. Mol. Biol. Rep. 2020, 47, 5651–5655. [Google Scholar] [CrossRef]
- Liang, L.; Zhao, L.; Zan, Y.; Zhu, Q.; Ren, J.; Zhao, X. MiR-93-5p enhances growth and angiogenesis capacity of HUVECs by down-regulating EPLIN. Oncotarget 2017, 8, 107033–107043. [Google Scholar] [CrossRef] [Green Version]
- Vlachos, I.S.; Zagganas, K.; Paraskevopoulou, M.D.; Georgakilas, G.; Karagkouni, D.; Vergoulis, T.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-miRPath v3.0: Deciphering microRNA function with experimental support. Nucleic Acids Res. 2015, 43, W460–W466. [Google Scholar] [CrossRef]
- Licursi, V.; Conte, F.; Fiscon, G.; Paci, P. MIENTURNET: An interactive web tool for microRNA-target enrichment and network-based analysis. BMC Bioinform. 2019, 20, 545. [Google Scholar] [CrossRef] [Green Version]
- Kehl, T.; Kern, F.; Backes, C.; Fehlmann, T.; Stockel, D.; Meese, E.; Lenhof, H.P.; Keller, A. miRPathDB 2.0: A novel release of the miRNA Pathway Dictionary Database. Nucleic Acids Res. 2020, 48, D142–D147. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normalized to HK | Normalized to all angiomiRs | ||||
|---|---|---|---|---|---|
| HG mean ± SD | HGNG mean ± SD | HG Mean ± SD | HGNG mean ± SD | ||
| Exposure to current and transient hyperglycemia | let-7a-2-3p | 1.09 ± 0.50 | 0.68 ± 0.23 | 0.83 ± 0.55 | 0.89 ± 0.41 |
| mir132-3p | 1.02 ± 0.34 | 0.74 ± 0.15 † | 0.94 ± 0.31 | 0.89 ± 0.16 | |
| mir134-5p | 1.01 ± 0.46 | 0.85 ± 0.24 | 0.92 ± 0.23 | 0.88 ± 0.11 | |
| mir139-5p | 1.49 ± 1.43 | 0.68 ± 0.19 | 0.84 ± 0.67 | 0.78 ± 0.30 | |
| mir181b-5p | 1.45 ± 1.05 | 0.82 ± 0.17 † | 0.89 ± 0.24 | 0.93 ± 0.16 | |
| mir20a-5p | 1.41 ± 1.54 | 1.16 ± 0.59 | 1.11 ± 1.04 | 1.33 ± 0.49 | |
| mir210-3p | 1.21 ± 1.34 | 0.68 ± 0.14 ** | 0.77 ± 0.46 | 0.84 ± 0.25 | |
| mir21-5p | 1.40 ± 1.04 | 0.81 ± 0.37 | 1.04 ± 0.67 | 0.91 ± 0.33 | |
| mir221-3p | 1.19 ± 0.80 | 0.79 ± 0.34 | 1.14 ± 0.44 | 0.96 ± 0.36 | |
| mir222-3p | 1.13 ± 0.67 | 0.79 ± 0.20 | 1.01 ± 0.37 | 1.09 ± 0.30 | |
| mir23b-3p | 1.40 ± 0.71 | 0.77 ± 0.23 | 1.09 ± 0.52 | 0.77 ± 0.23 | |
| mir24-2-5p | 1.17 ± 0.96 | 0.88 ± 0.48 | 0.80 ± 0.57 | 1.01 ± 0.42 | |
| mir27a-3p | 1.54 ± 0.87 | 0.90 ± 0.33 | 1.02 ± 0.44 | 1.05 ± 0.26 | |
| mir296-5p | 1.02 ± 0.89 | 0.91 ± 0.45 | 1.07 ± 0.87 | 0.91 ± 0.45 | |
| mir31-5p | 1.16 ± 0.86 | 0.94 ± 0.45 | 0.77 ± 0.48 | 1.07 ± 0.34 | |
| mir320a | 1.01 ± 0.39 | 0.65 ± 0.12 * | 0.85 ± 0.41 | 0.83 ± 0.21 † | |
| mir34a-3p | 1.65 ± 1.10 | 0.87 ± 0.41 | 1.27 ± 0.67 | 1.01 ± 0.35 | |
| mir503-5p | 1.38 ± 1.05 | 0.88 ± 0.49 | 0.75 ± 0.46 | 1.10 ± 0.68 | |
| mir93-5p | 1.29 ± 0.92 | 0.82 ± 0.26 | 0.90 ± 0.31 | 0.99 ± 0.18 | |
| Normalized to HK | Normalized to all angiomiRs | ||||
| HO mean ± SD | HONO mean ± SD | HO mean ± SD | HONO mean ± SD | ||
| Exposure to current and transient hypoxia | let-7a-2-3p | 0.85 ± 0.34 | 1.19 ± 0.54 | 0.61 ± 0.27 † | 1.15 ± 0.39 |
| mir132-3p | 1.95 ± 1.45 * | 1.83 ± 0.55 ** | 1.47 ± 0.73 † | 1.83 ± 0.65 ** | |
| mir134-5p | 1.60 ± 1.42 † | 1.34 ± 0.61 * | 1.09 ± 0.69 | 1.41 ± 0.66 * | |
| mir139-5p | 1.15 ± 1.03 | 0.66 ± 0.35 † | 0.85 ± 0.83 | 0.70 ± 0.35 † | |
| mir181b-5p | 1.71 ± 1.06 * | 1.34 ± 0.44 * | 1.21 ± 0.53 | 1.36 ± 0.38 * | |
| mir20a-5p | 1.33 ± 0.60 | 1.27 ± 0.47 | 0.99 ± 0.41 | 1.53 ± 0.72 | |
| mir210-3p | 0.81 ± 0.48 | 0.30 ± 0.24 *** | 0.55 ± 0.35 * | 0.28 ± 0.20 *** | |
| mir21-5p | 2.79 ± 1.93 ** | 1.00 ± 0.43 | 2.13 ± 1.03 ** | 1.02 ± 0.33 | |
| mir221-3p | 1.19 ± 0.46 | 1.05 ± 0.63 | 0.87 ± 0.34 | 1.01 ± 0.35 | |
| mir222-3p | 1.13 ± 0.31 | 1.01 ± 0.52 | 0.83 ± 0.25 | 0.99 ± 0.34 | |
| mir23b-3p | 1.56 ± 0.46 ** | 1.45 ± 0.97 | 1.63 ± 0.70 *** | 1.61 ± 1.10 † | |
| mir24-2-5p | 2.04 ± 1.14 * | 1.46 ± 0.91 | 1.48 ± 0.65 | 1.56 ± 0.52 * | |
| mir27a-3p | 1.22 ± 0.79 | 0.83 ± 0.48 | 1.03 ± 0.41 | 0.81 ± 0.35 * | |
| mir296-5p | 0.98 ± 0.55 | 0.80 ± 0.48 | 0.69 ± 0.29 | 0.84 ± 0.58 | |
| mir31-5p | 1.15 ± 0.47 | 0.70 ± 0.20 * | 1.04 ± 0.65 | 0.74 ± 0.27 *** | |
| mir320a | 1.11 ± 0.60 | 0.69 ± 0.13 ** | 1.11 ± 0.60 | 0.69 ± 0.13 * | |
| mir34a-3p | 1.09 ± 0.58 | 1.24 ± 0.38 | 0.79 ± 0.40 | 1.31 ± 0.37 | |
| mir503-5p | 1.09 ± 0.77 | 0.88 ± 0.35 | 0.74 ± 0.44 † | 0.88 ± 0.44 | |
| mir93-5p | 1.26 ± 0.39 | 1.04 ± 0.44 | 0.93 ± 0.29 | 1.05 ± 0.28 | |
| DIANA miRPath | Mienturnet | miRPathDB | ||
| Current hypoxia | HO normalized to HK: | |||
| Proteoglycans in cancer | x | x | ||
| FOXO signalling pathway | x | x | ||
| HO normalized to all angiomiRs: | ||||
| Proteoglycans in cancer | x | x | x | |
| FOXO signalling pathway | x | x | ||
| Cell cycle | x | x | x | |
| HIF-1 signalling pathway | x | x | ||
| Estrogen signalling pathway | x | x | ||
| Ras signalling pathway | x | x | ||
| Neurotrophin signalling pathway | ||||
| P53 signalling pathway | x | x | ||
| MAPK signalling pathway | x | x | ||
| Prolactin signalling pathway | x | x | ||
| Progesterone mediated oocyte maturation | x | x | ||
| Transient hypoxia | HONO normalized to HK: | |||
| Proteoglycans in cancer | x | x | x | |
| HIF-1 signalling pathway | x | x | ||
| Adherens junctions | x | x | ||
| HONO normalized to all angiomiRs: | ||||
| Proteoglycans in cancer | x | x | x | |
| HIF-1 signalling pathway | x | x | ||
| Adherens junctions | x | x | ||
| Estrogen signalling pathway | x | x | x | |
| Ras signalling pathway | x | x | ||
| FOXO signalling pathway | x | x | ||
| Neurotrophin signalling pathway | x | x | ||
| P53 signalling pathway | x | x | ||
| Protein processing in ER | x | x | ||
| miRNA | Qiagen Primer Assay ID | Reference |
|---|---|---|
| AngiomiRs: | ||
| let-7a-2-3p | Hs_let-7a-2*_2 | [62] |
| miR-132-3p | Hs_miR-132_1 | [63,64] |
| miR-134-5p | Hs_miR-134_2 | [64] |
| miR-139-5p | Hs_miR-139_1 | [65] |
| miR-181b-5p | Hs_miR-181b_1 | [62] |
| miR-199a-3p | Hs_miR-199a-3p_1 | [66] |
| miR-20a-5p | Hs_miR-20a_1 | [67,68] |
| miR-210-3p | Hs_miR-210_1 | [68] |
| miR-21-5p | Hs_miR-21_2 | [69] |
| miR-221-3p | Hs_miR-221_1 | [70] |
| miR-222-3p | Hs_miR-222_2 | [70] |
| miR-23b-3p | Hs_miR-23b_2 | [71] |
| miR-24-2-5p | Hs_miR-24-2*_1 | [71] |
| miR-27a-3p | Hs_miR-27a_1 | [71] |
| miR-296-5p | Hs_miR-296-5p_1 | [72] |
| miR-31-5p | Hs_miR-31_1 | [73] |
| miR-320a | Hs_miR-320a_1 | [74] |
| miR-34a_3p | Hs_miR-34a*_1 | [62] |
| miR-503-5p | Hs_miR-503_2 | [71] |
| miR-93-5p | Hs_miR-93_1 | [72] |
| Housekeeping RNAs: | ||
| RNU6-6P | Hs_RNU6-2_11 | [60] |
| miR-191-5p | Hs_miR-191_1 | [59,61] |
| mir-28-3p | Hs_miR-28-3p_1 | [59,61] |
| mir-30b | Hs_miR-30b_1 | [58] |
| miR-423-3p | Hs_miR_423_1 | [58] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Strutz, J.; Baumann, K.; Weiss, E.; Hiden, U. Transient Hyperglycemia and Hypoxia Induce Memory Effects in AngiomiR Expression Profiles of Feto-Placental Endothelial Cells. Int. J. Mol. Sci. 2021, 22, 13378. https://doi.org/10.3390/ijms222413378
Strutz J, Baumann K, Weiss E, Hiden U. Transient Hyperglycemia and Hypoxia Induce Memory Effects in AngiomiR Expression Profiles of Feto-Placental Endothelial Cells. International Journal of Molecular Sciences. 2021; 22(24):13378. https://doi.org/10.3390/ijms222413378
Chicago/Turabian StyleStrutz, Jasmin, Kathrin Baumann, Elisa Weiss, and Ursula Hiden. 2021. "Transient Hyperglycemia and Hypoxia Induce Memory Effects in AngiomiR Expression Profiles of Feto-Placental Endothelial Cells" International Journal of Molecular Sciences 22, no. 24: 13378. https://doi.org/10.3390/ijms222413378
APA StyleStrutz, J., Baumann, K., Weiss, E., & Hiden, U. (2021). Transient Hyperglycemia and Hypoxia Induce Memory Effects in AngiomiR Expression Profiles of Feto-Placental Endothelial Cells. International Journal of Molecular Sciences, 22(24), 13378. https://doi.org/10.3390/ijms222413378