
The Framework for Human Host Immune Responses to Four Types of Parasitic Infections and Relevant Key JAK/STAT Signaling

 , and
, and
Abstract
1. Introduction
2. Host Immunological Pathways for Different Types of Parasitic Infections
2.1. Intracellular Protozoa and TH1/TH1-like Immunity
2.2. Extracellular Protozoa and TH22/TH17 Immunity
2.3. Helminths (Endoparasites) and Eradicable TH2a Immunity
2.4. Parasitic Insects and Arachnids (Ectoparasites) and Eradicable TH2b Immunity
2.5. Parasites and Tolerable TH9 Immunity
3. Other Host Immunological Pathways: Immunity against Viruses
4. Future Perspectives
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hu, W.C. A Framework of All Discovered Immunological Pathways and Their Roles for Four Specific Types of Pathogens and Hypersensitivities. Front. Immunol. 2020, 11, 1992. [Google Scholar] [CrossRef]
- Hu, W.C. Human immune responses to Plasmodium falciparum infection: Molecular evidence for a suboptimal THαβ and TH17 bias over ideal and effective traditional TH1 immune response. Malar. J. 2013, 12, 392. [Google Scholar] [CrossRef]
- Gao, Q.; Liang, X.; Shaikh, A.S.; Zang, J.; Xu, W.; Zhang, Y. JAK/STAT Signal Transduction: Promising Attractive Targets for Immune, Inflammatory and Hematopoietic Diseases. Curr. Drug Targets 2018, 19, 487–500. [Google Scholar] [CrossRef]
- Morris, R.; Kershaw, N.J.; Babon, J.J. The molecular details of cytokine signaling via the JAK/STAT pathway. Protein Sci. 2018, 27, 1984–2009. [Google Scholar] [CrossRef] [PubMed]
- Owen, K.L.; Brockwell, N.K.; Parker, B.S. JAK-STAT Signaling: A Double-Edged Sword of Immune Regulation and Cancer Progression. Cancers 2019, 11, 2002. [Google Scholar] [CrossRef]
- Seif, F.; Khoshmirsafa, M.; Aazami, H.; Mohsenzadegan, M.; Sedighi, G.; Bahar, M. The role of JAK-STAT signaling pathway and its regulators in the fate of T helper cells. Cell Commun. Signal 2017, 15, 23. [Google Scholar] [CrossRef]
- Villarino, A.V.; Kanno, Y.; O’Shea, J.J. Mechanisms and consequences of Jak-STAT signaling in the immune system. Nat. Immunol. 2017, 18, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Xin, P.; Xu, X.; Deng, C.; Liu, S.; Wang, Y.; Zhou, X.; Ma, H.; Wei, D.; Sun, S. The role of JAK/STAT signaling pathway and its inhibitors in diseases. Int. Immunopharmacol. 2020, 80, 106210. [Google Scholar] [CrossRef]
- Yao, Y.; Chen, C.L.; Yu, D.; Liu, Z. Roles of follicular helper and regulatory T cells in allergic diseases and allergen immunotherapy. Allergy 2021, 76, 456–470. [Google Scholar] [CrossRef]
- Crotty, S. T Follicular Helper Cell Biology: A Decade of Discovery and Diseases. Immunity 2019, 50, 1132–1148. [Google Scholar] [CrossRef] [PubMed]
- Scheeren, F.A.; Diehl, S.A.; Smit, L.A.; Beaumont, T.; Naspetti, M.; Bende, R.J.; Blom, B.; Karube, K.; Ohshima, K.; van Noesel, C.J.; et al. IL-21 is expressed in Hodgkin lymphoma and activates STAT5: Evidence that activated STAT5 is required for Hodgkin lymphomagenesis. Blood 2008, 111, 4706–4715. [Google Scholar] [CrossRef] [PubMed]
- Spolski, R.; Gromer, D.; Leonard, W.J. The gamma c family of cytokines: Fine-tuning signals from IL-2 and IL-21 in the regulation of the immune response. F1000Res 2017, 6, 1872. [Google Scholar] [CrossRef]
- Burchill, M.A.; Yang, J.; Vogtenhuber, C.; Blazar, B.R.; Farrar, M.A. IL-2 receptor beta-dependent STAT5 activation is required for the development of Foxp3+ regulatory T cells. J. Immunol. 2007, 178, 280–290. [Google Scholar] [CrossRef]
- Kirken, R.A.; Rui, H.; Malabarba, M.G.; Howard, O.M.; Kawamura, M.; O’Shea, J.J.; Farrar, W.L. Activation of JAK3, but not JAK1, is critical for IL-2-induced proliferation and STAT5 recruitment by a COOH-terminal region of the IL-2 receptor beta-chain. Cytokine 1995, 7, 689–700. [Google Scholar] [CrossRef]
- Romero, P.; Zippelius, A.; Kurth, I.; Pittet, M.J.; Touvrey, C.; Iancu, E.M.; Corthesy, P.; Devevre, E.; Speiser, D.E.; Rufer, N. Four functionally distinct populations of human effector-memory CD8+ T lymphocytes. J. Immunol. 2007, 178, 4112–4119. [Google Scholar] [CrossRef]
- Krovi, S.H.; Gapin, L. Invariant Natural Killer T Cell Subsets-More Than Just Developmental Intermediates. Front. Immunol. 2018, 9, 1393. [Google Scholar] [CrossRef] [PubMed]
- Tuttle, K.D.; Krovi, S.H.; Zhang, J.; Bedel, R.; Harmacek, L.; Peterson, L.K.; Dragone, L.L.; Lefferts, A.; Halluszczak, C.; Riemondy, K.; et al. TCR signal strength controls thymic differentiation of iNKT cell subsets. Nat. Commun. 2018, 9, 2650. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Huntington, N.D.; Belz, G.T.; Seillet, C. Type 1 Innate Lymphoid Cell Biology: Lessons Learnt from Natural Killer Cells. Front. Immunol. 2016, 7, 426. [Google Scholar] [CrossRef]
- Fuchs, A.; Vermi, W.; Lee, J.S.; Lonardi, S.; Gilfillan, S.; Newberry, R.D.; Cella, M.; Colonna, M. Intraepithelial type 1 innate lymphoid cells are a unique subset of IL-12- and IL-15-responsive IFN-gamma-producing cells. Immunity 2013, 38, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Xuan, W.; Qu, Q.; Zheng, B.; Xiong, S.; Fan, G.H. The chemotaxis of M1 and M2 macrophages is regulated by different chemokines. J. Leukoc. Biol. 2015, 97, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Tomiyama, H.; Takata, H.; Matsuda, T.; Takiguchi, M. Phenotypic classification of human CD8+ T cells reflecting their function: Inverse correlation between quantitative expression of CD27 and cytotoxic effector function. Eur. J. Immunol. 2004, 34, 999–1010. [Google Scholar] [CrossRef]
- Kemp, R.A.; Backstrom, B.T.; Ronchese, F. The phenotype of type 1 and type 2 CD8+ T cells activated in vitro is affected by culture conditions and correlates with effector activity. Immunology 2005, 115, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Gracie, J.A.; Bradley, J.A. Interleukin-12 induces interferon-gamma-dependent switching of IgG alloantibody subclass. Eur. J. Immunol. 1996, 26, 1217–1221. [Google Scholar] [CrossRef] [PubMed]
- Cabantous, S.; Poudiougou, B.; Traore, A.; Keita, M.; Cisse, M.B.; Doumbo, O.; Dessein, A.J.; Marquet, S. Evidence that interferon-gamma plays a protective role during cerebral malaria. J. Infect. Dis. 2005, 192, 854–860. [Google Scholar] [CrossRef] [PubMed]
- Sedegah, M.; Finkelman, F.; Hoffman, S.L. Interleukin 12 induction of interferon gamma-dependent protection against malaria. Proc. Natl. Acad. Sci. USA 1994, 91, 10700–10702. [Google Scholar] [CrossRef] [PubMed]
- Castellano, L.R.; Filho, D.C.; Argiro, L.; Dessein, H.; Prata, A.; Dessein, A.; Rodrigues, V. Th1/Th2 immune responses are associated with active cutaneous leishmaniasis and clinical cure is associated with strong interferon-gamma production. Hum. Immunol. 2009, 70, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Orellana, M.A.; Schreiber, R.D.; Remington, J.S. Interferon-gamma: The major mediator of resistance against Toxoplasma gondii. Science 1988, 240, 516–518. [Google Scholar] [CrossRef]
- Ehigiator, H.N.; McNair, N.; Mead, J.R. Cryptosporidium parvum: The contribution of Th1-inducing pathways to the resolution of infection in mice. Exp. Parasitol. 2007, 115, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Gowda, D.C.; Wu, X. Parasite Recognition and Signaling Mechanisms in Innate Immune Responses to Malaria. Front. Immunol. 2018, 9, 3006. [Google Scholar] [CrossRef] [PubMed]
- Lima, T.S.; Lodoen, M.B. Mechanisms of Human Innate Immune Evasion by Toxoplasma gondii. Front. Cell Infect. Microbiol. 2019, 9, 103. [Google Scholar] [CrossRef]
- Mukhopadhyay, D.; Arranz-Solís, D.; Saeij, J.P.J. Influence of the Host and Parasite Strain on the Immune Response During Toxoplasma Infection. Front. Cell Infect. Microbiol. 2020, 10, 580425. [Google Scholar] [CrossRef]
- Rodríguez-Serrato, M.A.; Salinas-Carmona, M.C.; Limón-Flores, A.Y. Immune response to Leishmania mexicana: The host-parasite relationship. Pathog. Dis. 2020, 78. [Google Scholar] [CrossRef]
- Sasai, M.; Pradipta, A.; Yamamoto, M. Host immune responses to Toxoplasma gondii. Int. Immunol. 2018, 30, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Yam, X.Y.; Preiser, P.R. Host immune evasion strategies of malaria blood stage parasite. Mol. Biosyst. 2017, 13, 2498–2508. [Google Scholar] [CrossRef] [PubMed]
- Floss, D.M.; Klocker, T.; Schroder, J.; Lamertz, L.; Mrotzek, S.; Strobl, B.; Hermanns, H.; Scheller, J. Defining the functional binding sites of interleukin 12 receptor beta1 and interleukin 23 receptor to Janus kinases. Mol. Biol. Cell 2016, 27, 2301–2316. [Google Scholar] [CrossRef] [PubMed]
- Kohlhuber, F.; Rogers, N.C.; Watling, D.; Feng, J.; Guschin, D.; Briscoe, J.; Witthuhn, B.A.; Kotenko, S.V.; Pestka, S.; Stark, G.R.; et al. A JAK1/JAK2 chimera can sustain alpha and gamma interferon responses. Mol. Cell Biol. 1997, 17, 695–706. [Google Scholar] [CrossRef]
- Bernink, J.H.; Peters, C.P.; Munneke, M.; te Velde, A.A.; Meijer, S.L.; Weijer, K.; Hreggvidsdottir, H.S.; Heinsbroek, S.E.; Legrand, N.; Buskens, C.J.; et al. Human type 1 innate lymphoid cells accumulate in inflamed mucosal tissues. Nat. Immunol. 2013, 14, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.K.; Colonna, M. Innate Lymphoid Cells in Mucosal Immunity. Front. Immunol. 2019, 10, 861. [Google Scholar] [CrossRef] [PubMed]
- Barsheshet, Y.; Wildbaum, G.; Levy, E.; Vitenshtein, A.; Akinseye, C.; Griggs, J.; Lira, S.A.; Karin, N. CCR8(+)FOXp3(+) Treg cells as master drivers of immune regulation. Proc. Natl. Acad. Sci. USA 2017, 114, 6086–6091. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, Y.; Zhang, J.B.; Fakher, M.; Best, A.M.; Schenkein, H.A.; Barbour, S.E.; Tew, J.G. Non-redundant roles for interleukin-1 alpha and interleukin-1 beta in regulating human IgG2. J. Periodontol. 2001, 72, 1332–1339. [Google Scholar] [CrossRef]
- Cai, C.W.; Blase, J.R.; Zhang, X.; Eickhoff, C.S.; Hoft, D.F. Th17 Cells Are More Protective Than Th1 Cells Against the Intracellular Parasite Trypanosoma cruzi. PLoS Pathog. 2016, 12, e1005902. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Barroso, L.; Lyerly, D.M.; Petri, W.A., Jr.; Houpt, E.R. CD4+ and CD8+ T cell- and IL-17-mediated protection against Entamoeba histolytica induced by a recombinant vaccine. Vaccine 2011, 29, 772–777. [Google Scholar] [CrossRef]
- Suryawanshi, A.; Cao, Z.; Sampson, J.F.; Panjwani, N. IL-17A-mediated protection against Acanthamoeba keratitis. J. Immunol. 2015, 194, 650–663. [Google Scholar] [CrossRef] [PubMed]
- Makinde, H.M.; Zariffard, R.; Mirmonsef, P.; Novak, R.M.; Jarrett, O.; Landay, A.L.; Spear, G.T. IL-22 Levels are Associated withTrichomonas vaginalisInfection in the Lower Genital Tract. Am. J. Reprod. Immunol. 2013, 70, 38–44. [Google Scholar] [CrossRef]
- Singer, S.M. Control of Giardiasis by Interleukin-17 in Humans and Mice—Are the Questions All Answered? Clin. Vaccine Immunol. 2016, 23, 2–5. [Google Scholar] [CrossRef]
- Belkaid, Y.; Harrison, O.J. Homeostatic Immunity and the Microbiota. Immunity 2017, 46, 562–576. [Google Scholar] [CrossRef]
- Mercer, F.; Johnson, P.J. Trichomonas vaginalis: Pathogenesis, Symbiont Interactions, and Host Cell Immune Responses. Trends Parasitol. 2018, 34, 683–693. [Google Scholar] [CrossRef]
- Uribe-Querol, E.; Rosales, C. Immune Response to the Enteric Parasite Entamoeba histolytica. Physiology (Bethesda) 2020, 35, 244–260. [Google Scholar] [CrossRef]
- Kaptein, A.; Paillard, V.; Saunders, M. Dominant negative stat3 mutant inhibits interleukin-6-induced Jak-STAT signal transduction. J. Biol. Chem. 1996, 271, 5961–5964. [Google Scholar] [CrossRef]
- Backert, I.; Koralov, S.B.; Wirtz, S.; Kitowski, V.; Billmeier, U.; Martini, E.; Hofmann, K.; Hildner, K.; Wittkopf, N.; Brecht, K.; et al. STAT3 activation in Th17 and Th22 cells controls IL-22-mediated epithelial host defense during infectious colitis. J. Immunol. 2014, 193, 3779–3791. [Google Scholar] [CrossRef]
- Yu, C.Y.; Wang, L.; Khaletskiy, A.; Farrar, W.L.; Larner, A.; Colburn, N.H.; Li, J.J. STAT3 activation is required for interleukin-6 induced transformation in tumor-promotion sensitive mouse skin epithelial cells. Oncogene 2002, 21, 3949–3960. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lee, P.W.; Smith, A.J.; Yang, Y.; Selhorst, A.J.; Liu, Y.; Racke, M.K.; Lovett-Racke, A.E. IL-23R-activated STAT3/STAT4 is essential for Th1/Th17-mediated CNS autoimmunity. JCI Insight 2017, 2. [Google Scholar] [CrossRef]
- Pei, C.; Zhao, C.; Wang, A.J.; Fan, A.X.; Grinchuk, V.; Smith, A.; Sun, R.; Xie, Y.; Lu, N.; Urban, J.F., Jr.; et al. Critical Role for Interleukin-25 in Host Protective Th2 Memory Response against Heligmosomoides polygyrus bakeri. Infect. Immun. 2016, 84, 3328–3337. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Guo, L.; Qiu, J.; Chen, X.; Hu-Li, J.; Siebenlist, U.; Williamson, P.R.; Urban, J.F., Jr.; Paul, W.E. IL-25-responsive, lineage-negative KLRG1(hi) cells are multipotential ‘inflammatory’ type 2 innate lymphoid cells. Nat. Immunol. 2015, 16, 161–169. [Google Scholar] [CrossRef]
- Fort, M.M.; Cheung, J.; Yen, D.; Li, J.; Zurawski, S.M.; Lo, S.; Menon, S.; Clifford, T.; Hunte, B.; Lesley, R.; et al. IL-25 induces IL-4, IL-5, and IL-13 and Th2-associated pathologies in vivo. Immunity 2001, 15, 985–995. [Google Scholar] [CrossRef]
- Camelo, A.; Rosignoli, G.; Ohne, Y.; Stewart, R.A.; Overed-Sayer, C.; Sleeman, M.A.; May, R.D. IL-33, IL-25, and TSLP induce a distinct phenotypic and activation profile in human type 2 innate lymphoid cells. Blood Adv. 2017, 1, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Klechevsky, E.; Morita, R.; Liu, M.; Cao, Y.; Coquery, S.; Thompson-Snipes, L.; Briere, F.; Chaussabel, D.; Zurawski, G.; Palucka, A.K.; et al. Functional specializations of human epidermal Langerhans cells and CD14+ dermal dendritic cells. Immunity 2008, 29, 497–510. [Google Scholar] [CrossRef]
- Mesnil, C.; Raulier, S.; Paulissen, G.; Xiao, X.; Birrell, M.A.; Pirottin, D.; Janss, T.; Starkl, P.; Ramery, E.; Henket, M.; et al. Lung-resident eosinophils represent a distinct regulatory eosinophil subset. J. Clin. Investig. 2016, 126, 3279–3295. [Google Scholar] [CrossRef] [PubMed]
- Vliagoftis, H.; Lacy, P.; Luy, B.; Adamko, D.; Hollenberg, M.; Befus, D.; Moqbel, R. Mast cell tryptase activates peripheral blood eosinophils to release granule-associated enzymes. Int. Arch. Allergy Immunol. 2004, 135, 196–204. [Google Scholar] [CrossRef]
- Ranganath, S.; Murphy, K.M. Structure and specificity of GATA proteins in Th2 development. Mol. Cell Biol. 2001, 21, 2716–2725. [Google Scholar] [CrossRef]
- Zon, L.I.; Yamaguchi, Y.; Yee, K.; Albee, E.A.; Kimura, A.; Bennett, J.C.; Orkin, S.H.; Ackerman, S.J. Expression of mRNA for the GATA-binding proteins in human eosinophils and basophils: Potential role in gene transcription. Blood 1993, 81, 3234–3241. [Google Scholar] [CrossRef]
- van der Bruggen, T.; Caldenhoven, E.; Kanters, D.; Coffer, P.; Raaijmakers, J.A.; Lammers, J.W.; Koenderman, L. Interleukin-5 signaling in human eosinophils involves JAK2 tyrosine kinase and Stat1 alpha. Blood 1995, 85, 1442–1448. [Google Scholar] [CrossRef]
- Pazdrak, K.; Stafford, S.; Alam, R. The activation of the Jak-STAT 1 signaling pathway by IL-5 in eosinophils. J. Immunol. 1995, 155, 397–402. [Google Scholar] [PubMed]
- Klion, A.D.; Nutman, T.B. The role of eosinophils in host defense against helminth parasites. J. Allergy Clin. Immunol. 2004, 113, 30–37. [Google Scholar] [CrossRef]
- Mitre, E.; Masure, D.; Vlaminck, J.; Wang, T.; Chiers, K.; Van den Broeck, W.; Vercruysse, J.; Geldhof, P. A Role for Eosinophils in the Intestinal Immunity against Infective Ascaris suum Larvae. PLoS Negl. Trop. Dis. 2013, 7, e2138. [Google Scholar] [CrossRef]
- Wright, V.; Bickle, Q. Immune responses following experimental human hookworm infection. Clin. Exp. Immunol. 2005, 142, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Minciullo, P.L.; Cascio, A.; Isola, S.; Gangemi, S. Different clinical allergological features of Taenia solium infestation. Clin. Mol. Allergy 2016, 14, 18. [Google Scholar] [CrossRef]
- Schroeder, J.C.; Jones, D.; Maranich, A. Peripheral Eosinophilia Found in Pediatric Enterobius vermicularis Infections. Clin. Pediatr. 2019, 58, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.B.; Seo, J.W.; Lee, J.H.; Choi, B.S.; Park, S.G. Evaluation of the prevalence and clinical impact of toxocariasis in patients with eosinophilia of unknown origin. Korean J. Intern. Med. 2017, 32, 523–529. [Google Scholar] [CrossRef]
- Bakhtiar, N.M.; Spotin, A.; Mahami-Oskouei, M.; Ahmadpour, E.; Rostami, A. Recent advances on innate immune pathways related to host-parasite cross-talk in cystic and alveolar echinococcosis. Parasites Vectors 2020, 13, 232. [Google Scholar] [CrossRef] [PubMed]
- Gazzinelli-Guimaraes, P.H.; Nutman, T.B. Helminth parasites and immune regulation. F1000Res 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Joardar, N.; Mondal, C.; Sinha Babu, S.P. A review on the interactions between dendritic cells, filarial parasite and parasite-derived molecules in regulating the host immune responses. Scand. J. Immunol. 2021, 93, e13001. [Google Scholar] [CrossRef] [PubMed]
- Prodjinotho, U.F.; Lema, J.; Lacorcia, M.; Schmidt, V.; Vejzagic, N.; Sikasunge, C.; Ngowi, B.; Winkler, A.S.; Prazeres da Costa, C. Host immune responses during Taenia solium Neurocysticercosis infection and treatment. PLoS Negl. Trop. Dis. 2020, 14, e0008005. [Google Scholar] [CrossRef] [PubMed]
- Malabarba, M.G.; Kirken, R.A.; Rui, H.; Koettnitz, K.; Kawamura, M.; O’Shea, J.J.; Kalthoff, F.S.; Farrar, W.L. Activation of JAK3, but not JAK1, is critical to interleukin-4 (IL4) stimulated proliferation and requires a membrane-proximal region of IL4 receptor alpha. J. Biol. Chem. 1995, 270, 9630–9637. [Google Scholar] [CrossRef]
- Palmer-Crocker, R.L.; Hughes, C.C.; Pober, J.S. IL-4 and IL-13 activate the JAK2 tyrosine kinase and Stat6 in cultured human vascular endothelial cells through a common pathway that does not involve the gamma c chain. J. Clin. Investig. 1996, 98, 604–609. [Google Scholar] [CrossRef] [PubMed]
- Ogata, N.; Kouro, T.; Yamada, A.; Koike, M.; Hanai, N.; Ishikawa, T.; Takatsu, K. JAK2 and JAK1 Constitutively Associate With an Interleukin-5 (IL-5) Receptor α and βc Subunit, Respectively, and Are Activated Upon IL-5 Stimulation. Blood 1998, 91, 2264–2271. [Google Scholar] [CrossRef]
- Hsu, C.L.; Neilsen, C.V.; Bryce, P.J. IL-33 is produced by mast cells and regulates IgE-dependent inflammation. PLoS ONE 2010, 5, e11944. [Google Scholar] [CrossRef]
- Sakakibara, Y.; Wada, T.; Muraoka, M.; Matsuda, Y.; Toma, T.; Yachie, A. Basophil activation by mosquito extracts in patients with hypersensitivity to mosquito bites. Cancer Sci. 2015, 106, 965–971. [Google Scholar] [CrossRef] [PubMed]
- Karasuyama, H.; Tabakawa, Y.; Ohta, T.; Wada, T.; Yoshikawa, S. Crucial Role for Basophils in Acquired Protective Immunity to Tick Infestation. Front. Physiol. 2018, 9, 1769. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, R.E.; Schemmer, K.R. The role of basophils in the immunopathogenesis of hypersensitivity to fleas (Ctenocephalides felis) in dogs. Vet. Immunol. Immunopathol. 1987, 15, 203–213. [Google Scholar] [CrossRef]
- Erdmann, S.M.; Sachs, B.; Kwiecien, R.; Moll-Slodowy, S.; Sauer, I.; Merk, H.F. The basophil activation test in wasp venom allergy: Sensitivity, specificity and monitoring specific immunotherapy. Allergy 2004, 59, 1102–1109. [Google Scholar] [CrossRef]
- Cichocka-Jarosz, E.; Dorynska, A.; Pietrzyk, J.J.; Spiewak, R. Laboratory markers of mast cell and basophil activation in monitoring rush immunotherapy in bee venom-allergic children. Immunotherapy 2011, 3, 1013–1017. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Liu, H.Y.; Jaeson, M.I.; Deimel, L.P.; Ranasinghe, C. Unique IL-13Ralpha2/STAT3 mediated IL-13 regulation detected in lung conventional dendritic cells, 24 h post viral vector vaccination. Sci. Rep. 2020, 10, 1017. [Google Scholar] [CrossRef]
- He, H.; Guttman-Yassky, E. JAK Inhibitors for Atopic Dermatitis: An Update. Am. J. Clin. Dermatol. 2019, 20, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Lee, J.B.; Liu, B.; Ohta, S.; Wang, P.Y.; Kartashov, A.V.; Mugge, L.; Abonia, J.P.; Barski, A.; Izuhara, K.; et al. Induction of Interleukin-9-Producing Mucosal Mast Cells Promotes Susceptibility to IgE-Mediated Experimental Food Allergy. Immunity 2015, 43, 788–802. [Google Scholar] [CrossRef] [PubMed]
- Verma, M.; Liu, S.; Michalec, L.; Sripada, A.; Gorska, M.M.; Alam, R. Experimental asthma persists in IL-33 receptor knockout mice because of the emergence of thymic stromal lymphopoietin-driven IL-9(+) and IL-13(+) type 2 innate lymphoid cell subpopulations. J. Allergy Clin. Immunol. 2018, 142, 793–803.e8. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, K.; Hwang, Y.; Nikolaev, A.; Atreya, R.; Dornhoff, H.; Steiner, S.; Lehr, H.A.; Wirtz, S.; Vieth, M.; Waisman, A.; et al. TH9 cells that express the transcription factor PU.1 drive T cell-mediated colitis via IL-9 receptor signaling in intestinal epithelial cells. Nat. Immunol. 2014, 15, 676–686. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.C. The Central THalphabeta Immunity Associated Cytokine: IL-10 Has a Strong Anti-Tumor Ability Toward Established Cancer Models In Vivo and Toward Cancer Cells In Vitro. Front. Oncol. 2021, 11, 655554. [Google Scholar] [CrossRef]
- Geginat, J.; Vasco, M.; Gerosa, M.; Tas, S.W.; Pagani, M.; Grassi, F.; Flavell, R.A.; Meroni, P.; Abrignani, S. IL-10 producing regulatory and helper T-cells in systemic lupus erythematosus. Semin. Immunol. 2019, 44, 101330. [Google Scholar] [CrossRef]
- Gauzzi, M.C.; Velazquez, L.; McKendry, R.; Mogensen, K.E.; Fellous, M.; Pellegrini, S. Interferon-alpha-dependent activation of Tyk2 requires phosphorylation of positive regulatory tyrosines by another kinase. J. Biol. Chem. 1996, 271, 20494–20500. [Google Scholar] [CrossRef]
- Caldenhoven, E.; van Dijk, T.B.; Solari, R.; Armstrong, J.; Raaijmakers, J.A.; Lammers, J.W.; Koenderman, L.; de Groot, R.P. STAT3beta, a splice variant of transcription factor STAT3, is a dominant negative regulator of transcription. J. Biol. Chem. 1996, 271, 13221–13227. [Google Scholar] [CrossRef]
- Mogensen, T.H. IRF and STAT Transcription Factors—From Basic Biology to Roles in Infection, Protective Immunity, and Primary Immunodeficiencies. Front. Immunol. 2018, 9, 3047. [Google Scholar] [CrossRef]
- McCall, M.B.; Sauerwein, R.W. Interferon-γ-central mediator of protective immune responses against the pre-erythrocytic and blood stage of malaria. J. Leukoc. Biol. 2010, 88, 1131–1143. [Google Scholar] [CrossRef]
- Nishiyama, S.; Pradipta, A.; Ma, J.S.; Sasai, M.; Yamamoto, M. T cell-derived interferon-γ is required for host defense to Toxoplasma gondii. Parasitol. Int. 2020, 75, 102049. [Google Scholar] [CrossRef] [PubMed]
- Urban, J.F., Jr.; Katona, I.M.; Paul, W.E.; Finkelman, F.D. Interleukin 4 is important in protective immunity to a gastrointestinal nematode infection in mice. Proc. Natl. Acad. Sci. USA 1991, 88, 5513–5517. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, H.; Renauld, J.C.; Van Snick, J.; Grencis, R.K. Interleukin-9 enhances resistance to the intestinal nematode Trichuris muris. Infect. Immun. 1998, 66, 3832–3840. [Google Scholar] [CrossRef]
- Cartron, A.M.; Nguyen, T.H.; Roh, Y.S.; Kwatra, M.M.; Kwatra, S.G. Janus kinase inhibitors for atopic dermatitis: A promising treatment modality. Clin. Exp. Dermatol. 2021, 46, 820–824. [Google Scholar] [CrossRef]
- Farne, H.A.; Wilson, A.; Powell, C.; Bax, L.; Milan, S.J. Anti-IL5 therapies for asthma. Cochrane Database Syst Rev. 2017, 9, Cd010834. [Google Scholar] [CrossRef] [PubMed]
- Leckie, M.J.; ten Brinke, A.; Khan, J.; Diamant, Z.; O’Connor, B.J.; Walls, C.M.; Mathur, A.K.; Cowley, H.C.; Chung, K.F.; Djukanovic, R.; et al. Effects of an interleukin-5 blocking monoclonal antibody on eosinophils, airway hyper-responsiveness, and the late asthmatic response. Lancet 2000, 356, 2144–2148. [Google Scholar] [CrossRef]
- Corren, J.; Lemanske, R.F.; Hanania, N.A.; Korenblat, P.E.; Parsey, M.V.; Arron, J.R.; Harris, J.M.; Scheerens, H.; Wu, L.C.; Su, Z.; et al. Lebrikizumab treatment in adults with asthma. N. Engl. J. Med. 2011, 365, 1088–1098. [Google Scholar] [CrossRef] [PubMed]
- Matucci, A.; Vultaggio, A.; Maggi, E.; Kasujee, I. Is IgE or eosinophils the key player in allergic asthma pathogenesis? Are we asking the right question? Respir. Res. 2018, 19, 113. [Google Scholar] [CrossRef]



{kind=link}
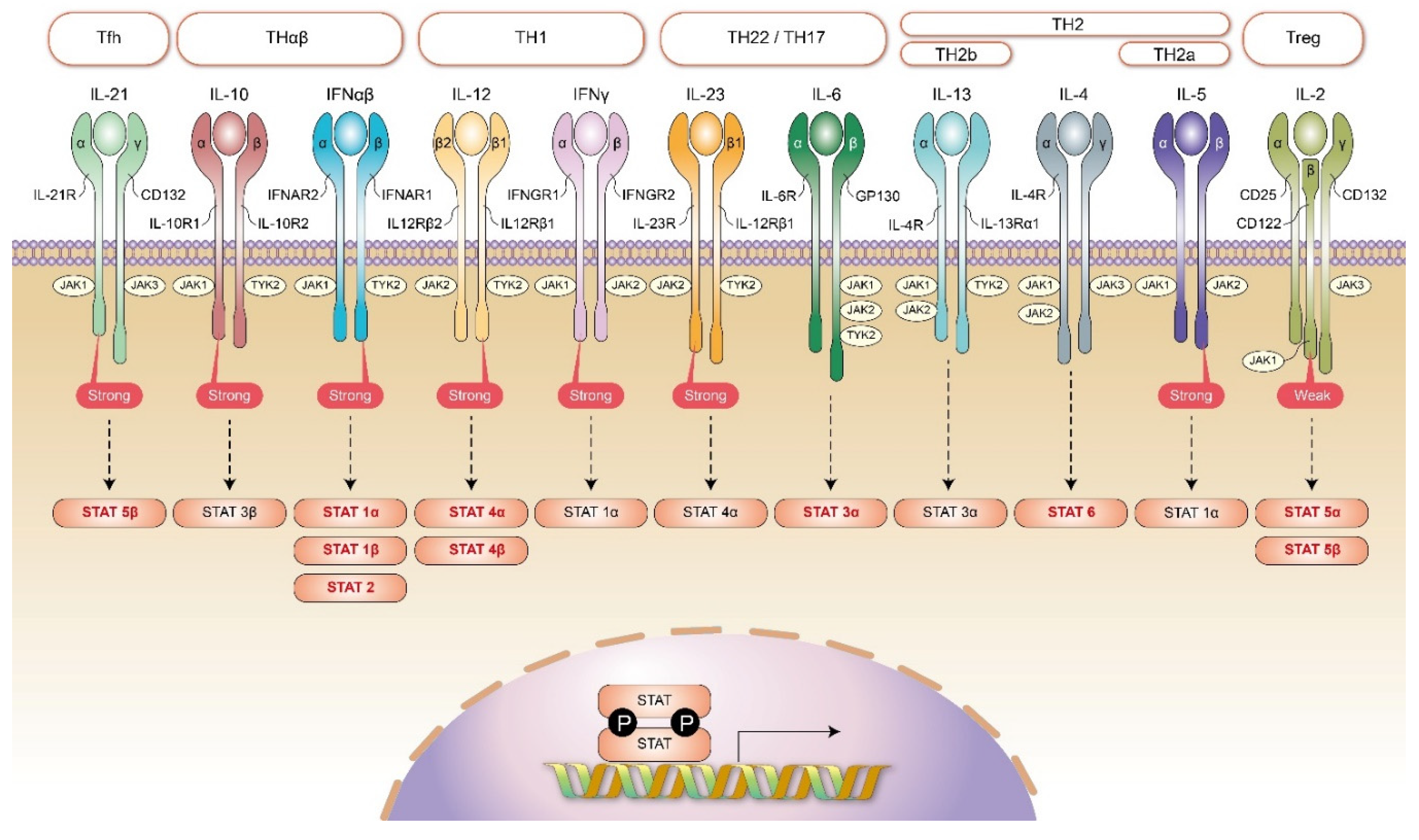{kind=link}
| Immune Pathways | Cytokines | Transcription Factors | Innate Lymphoid Cells | Effector Cells | CD4 T Cells | B Cells | NKT Cells | Pathogens | Autoimmunity |
|---|---|---|---|---|---|---|---|---|---|
| TH1 eradicable immunity | IFNg, IL-12 | STAT1, STAT4, T-bet | ILC1 | Macrophages M1 | IFNg CD4 T cells | IgG3 | iNKT1 | Intracellular protozoa, bacteria, and fungi | Type 4 DTH |
| TH1-like tolerable immunity | IFNg, TGFβ | STAT1, STAT4, STAT5 | ILC1 | Macrophages M2 | IFNg/TGFβCD4 T cells | IgA1 | iNKT1 | Intracellular protozoa, bacteria, and fungi | Type 4 DTH |
| TH22 eradicable immunity | IL-1, IL-6, TNFα | STAT3, AHR | ILC3 NCR+ | Neutrophils N1 | IL-22 CD4 T cells | IgG2 | iNKT17 | Extracellular protozoa, bacteria, and fungi | Type 3 immune complex |
| TH17 tolerable immunity | IL-6, TGFβ | STAT3, STAT5, RORγt | ILC3 NCR- | Neutrophils N2 | IL-17 CD4 T cells | IgA2 | iNKT17 | Extracellular protozoa, bacteria, and fungi | Type 3 immune complex |
| TH2a eradicable immunity | IL-4, IL-5 | STAT6, STAT1, GATA1/3 | IL-25 iILC2 | Eosinophils (iEOS), mast cells (MCt) | IL-5 CD4 T cells | IgG4 | iNKT2 | Helminths | Type 1 allergy IgG4 related |
| TH2b eradicable immunity | IL-4, IL-13 | STAT6, STAT3, GATA2/3 | IL-33 nILC2 | Basophils, mast cells (MCct) | IL-4 /IL-13 CD4 T cells | IgE | iNKT2 | Insects | Type 1 allergy IgE related |
| TH9 tolerable immunity | IL-4, TGFβ | STAT6, STAT5, PU.1 | TSLP ILC2 | Eosinophils (rEOS), basophils, mast cells (MMC9) | IL-9 CD4 T cells | IgA2 | iNKT2 | Helminths and Insects | Type 1 allergy |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, T.-H.; Tsai, K.-W.; Wu, Y.-J.; Liao, M.-T.; Lu, K.-C.; Hu, W.-C. The Framework for Human Host Immune Responses to Four Types of Parasitic Infections and Relevant Key JAK/STAT Signaling. Int. J. Mol. Sci. 2021, 22, 13310. https://doi.org/10.3390/ijms222413310
Wen T-H, Tsai K-W, Wu Y-J, Liao M-T, Lu K-C, Hu W-C. The Framework for Human Host Immune Responses to Four Types of Parasitic Infections and Relevant Key JAK/STAT Signaling. International Journal of Molecular Sciences. 2021; 22(24):13310. https://doi.org/10.3390/ijms222413310
Chicago/Turabian StyleWen, Tsung-Han, Kuo-Wang Tsai, Yan-Jun Wu, Min-Tser Liao, Kuo-Cheng Lu, and Wan-Chung Hu. 2021. "The Framework for Human Host Immune Responses to Four Types of Parasitic Infections and Relevant Key JAK/STAT Signaling" International Journal of Molecular Sciences 22, no. 24: 13310. https://doi.org/10.3390/ijms222413310
APA StyleWen, T.-H., Tsai, K.-W., Wu, Y.-J., Liao, M.-T., Lu, K.-C., & Hu, W.-C. (2021). The Framework for Human Host Immune Responses to Four Types of Parasitic Infections and Relevant Key JAK/STAT Signaling. International Journal of Molecular Sciences, 22(24), 13310. https://doi.org/10.3390/ijms222413310