
EGFR Transgene Stimulates Spontaneous Formation of MCF7 Breast Cancer Cells Spheroids with Partly Loss of HER3 Receptor

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
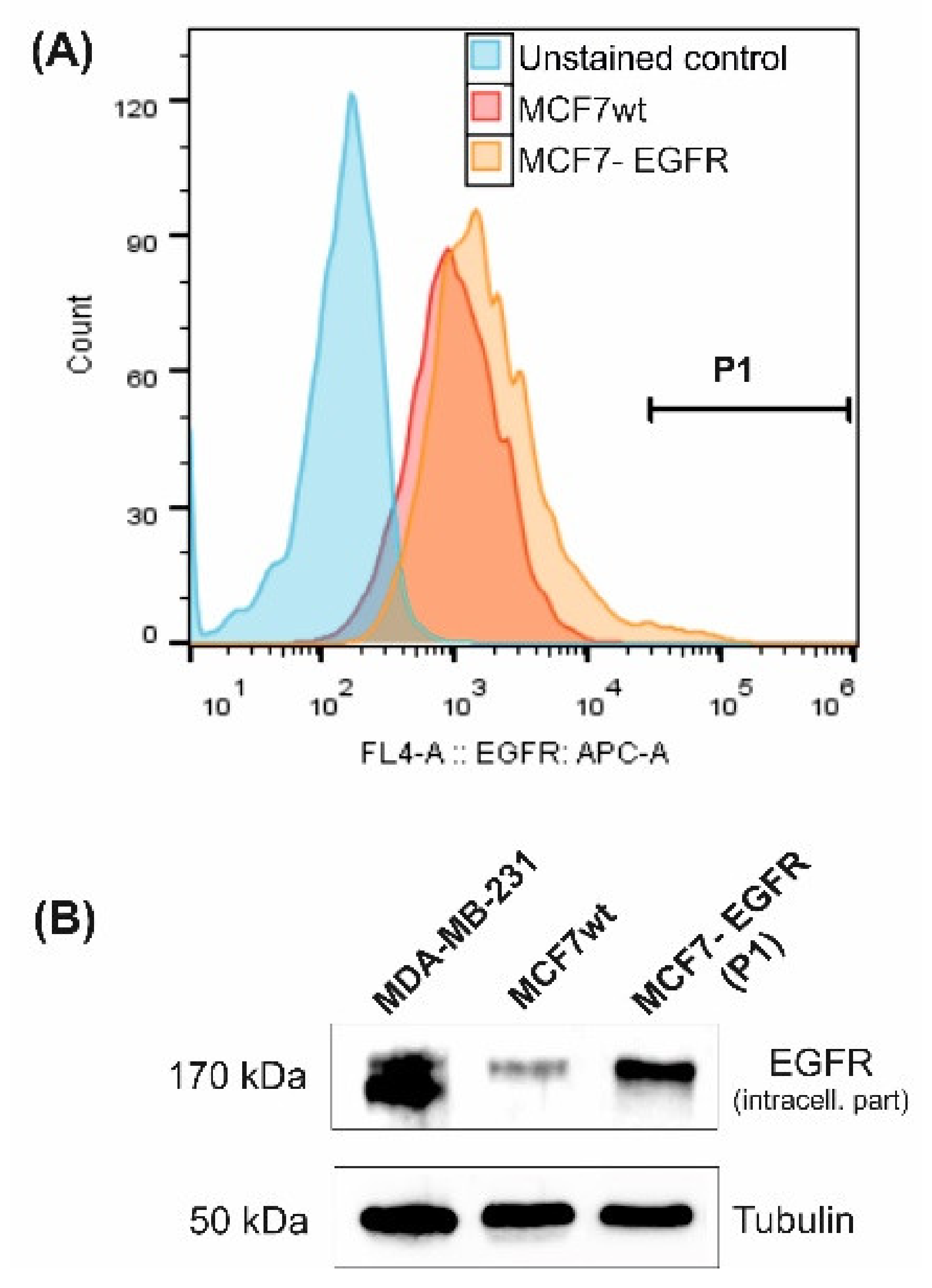
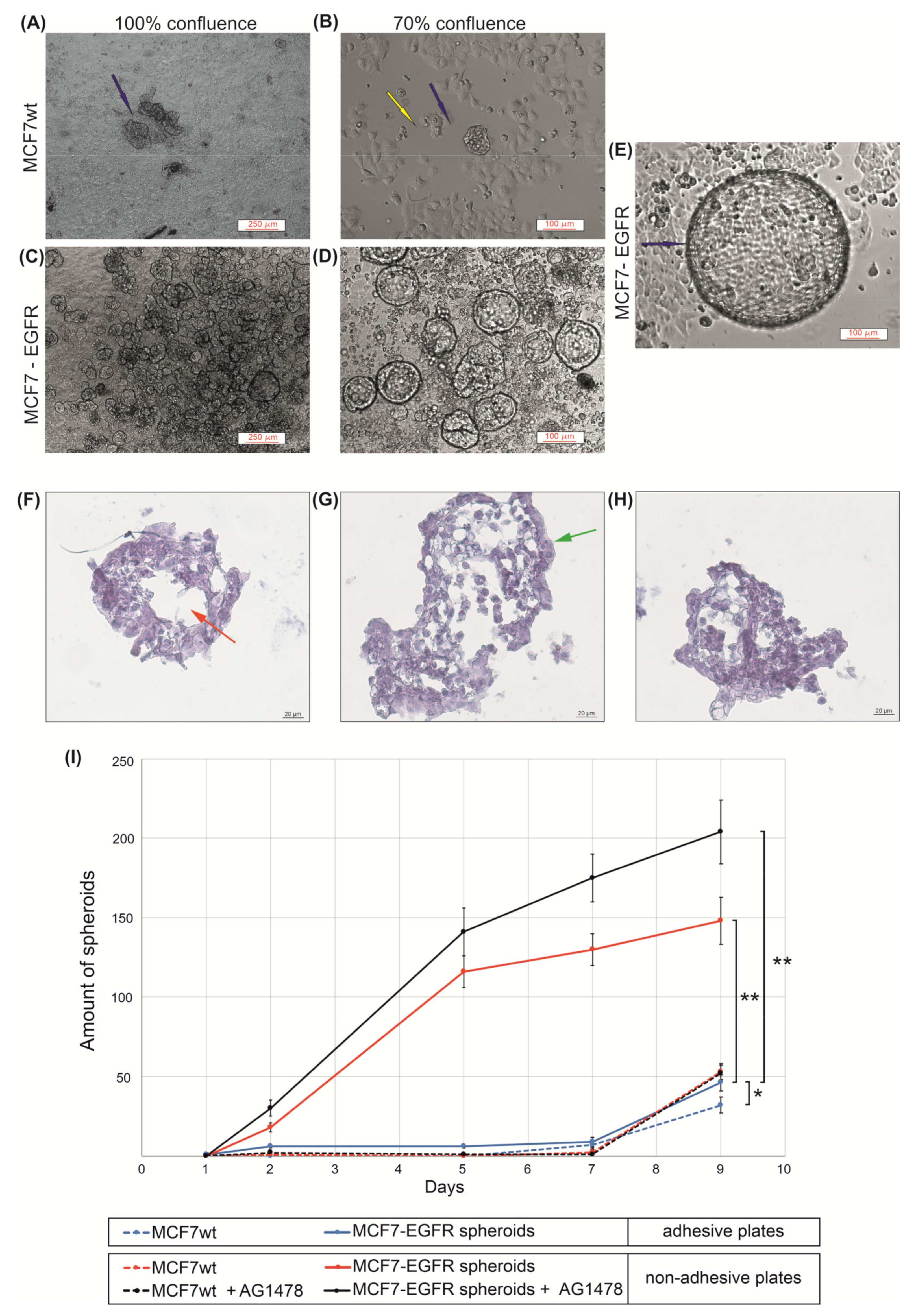
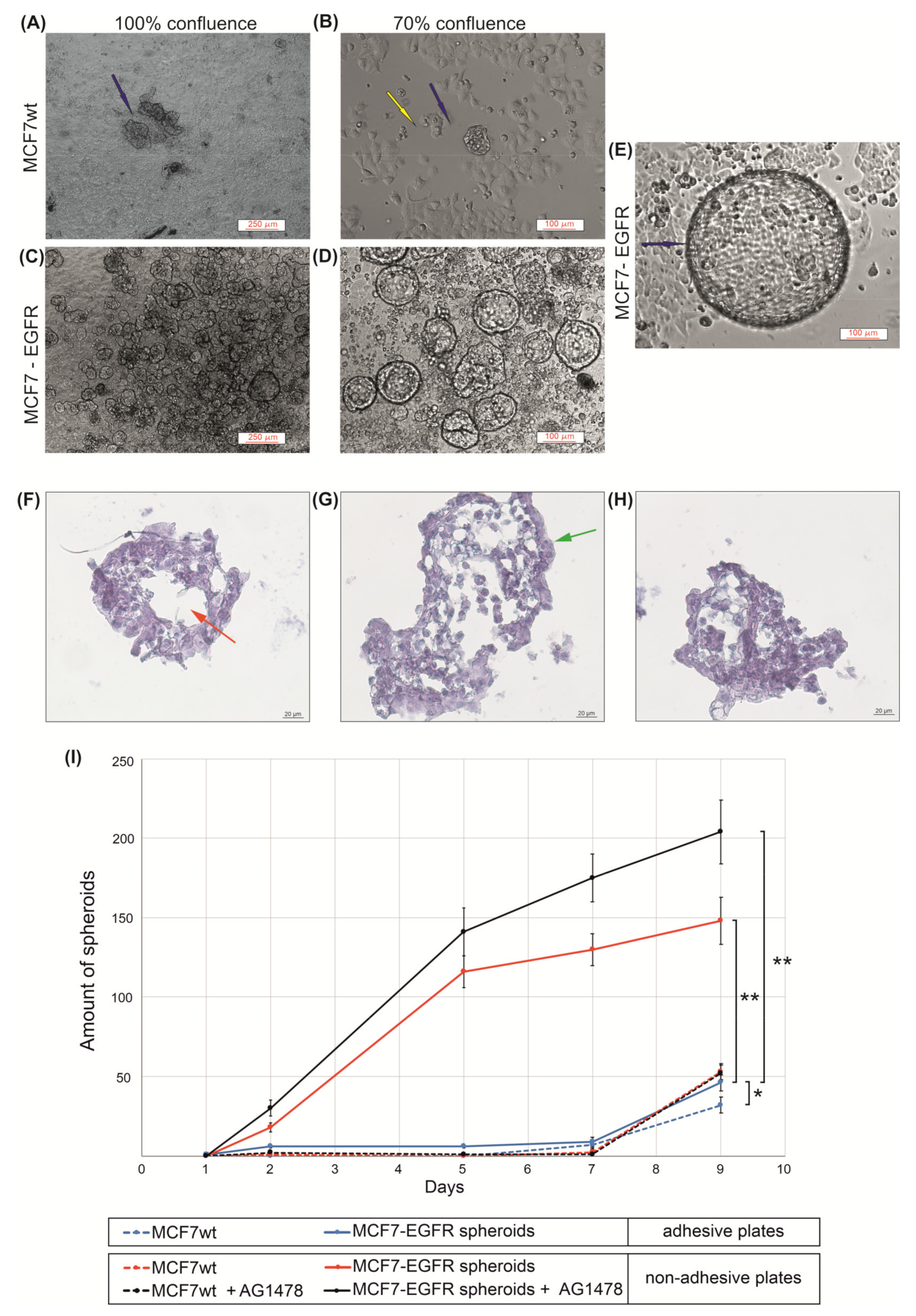
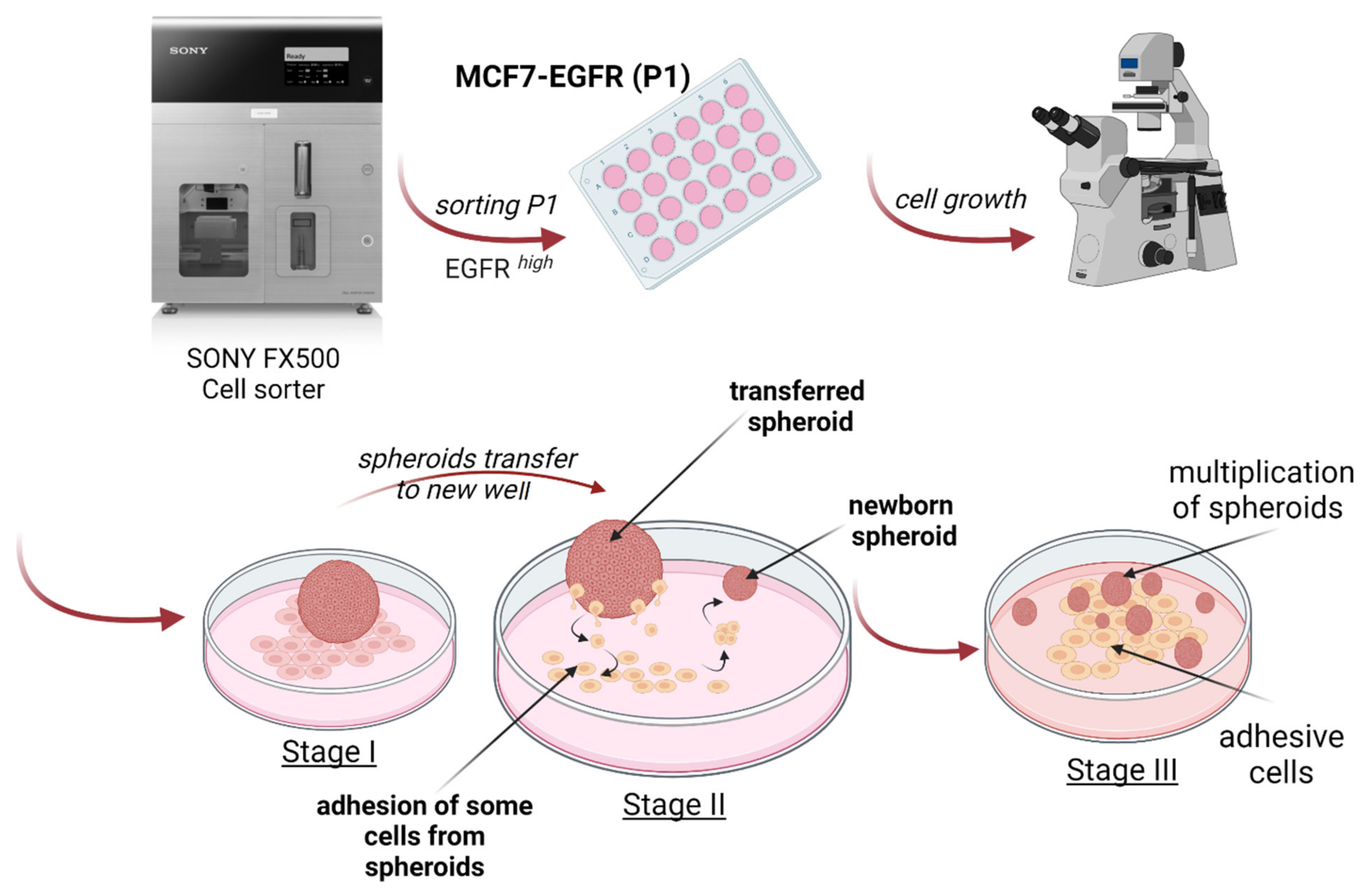
2.1. MCF7-EGFR Cell Cultivation and Spheroid Formation
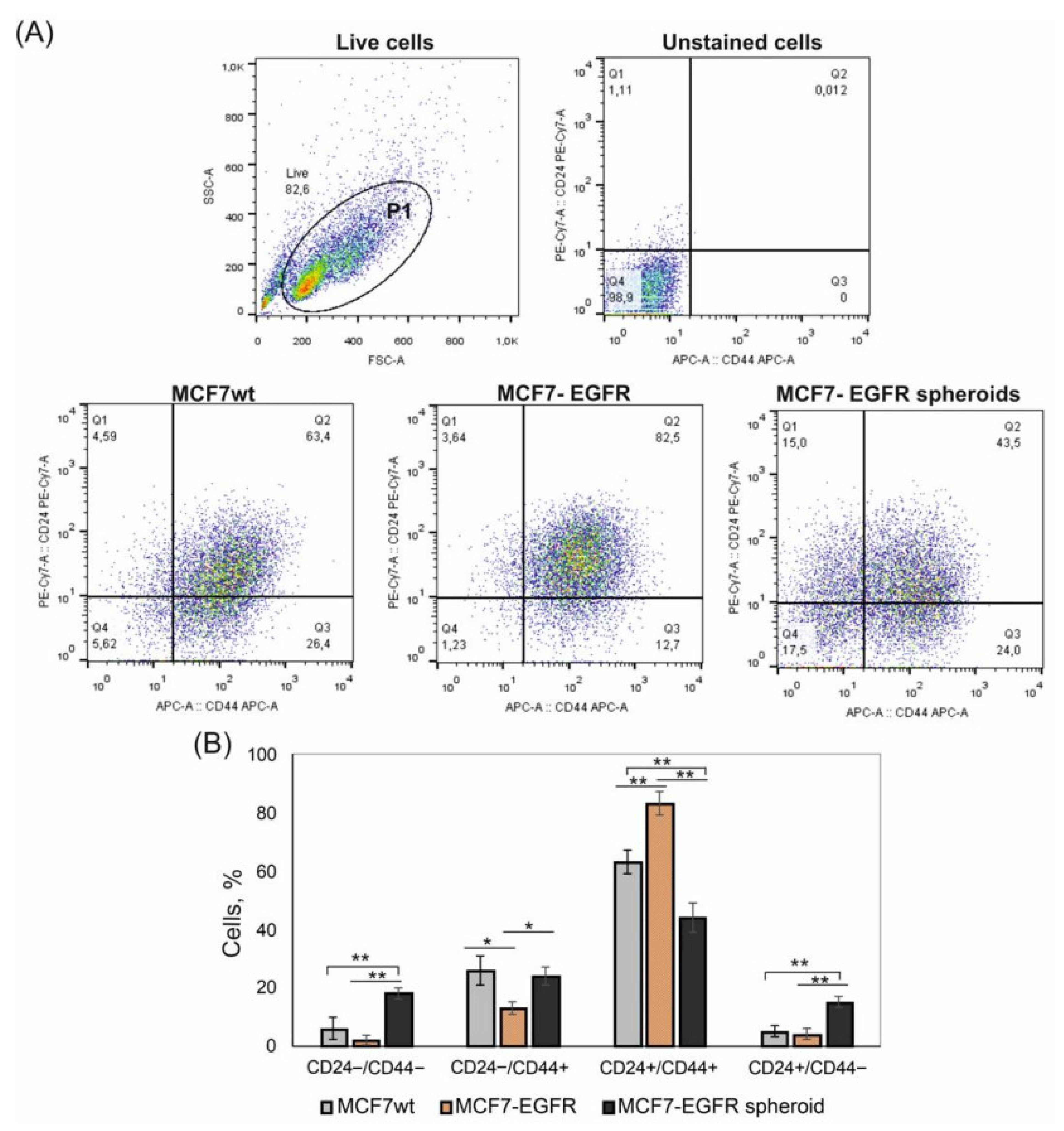
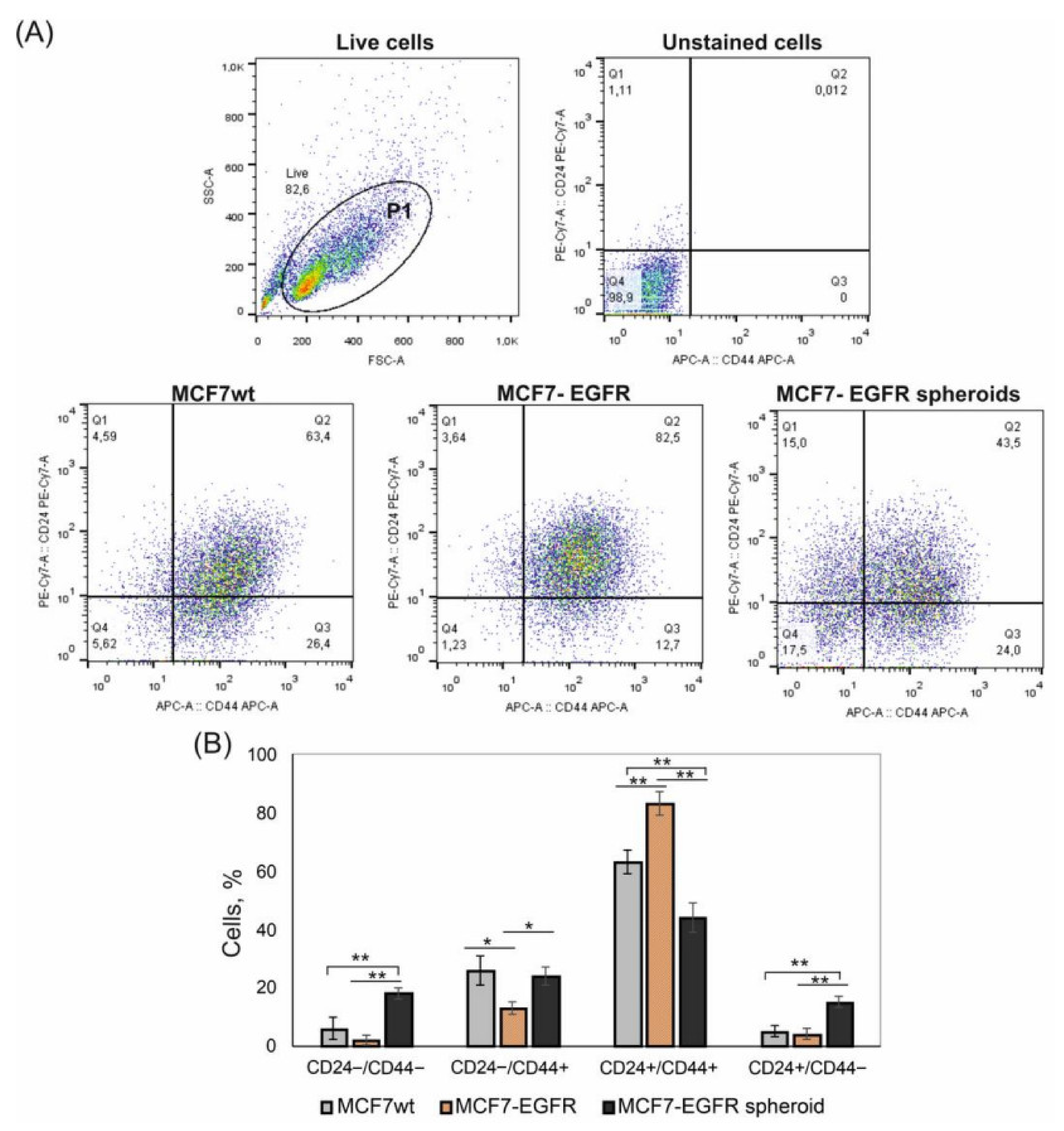
2.2. CD24/CD44 Content in MCF7-EGFR Spheroids
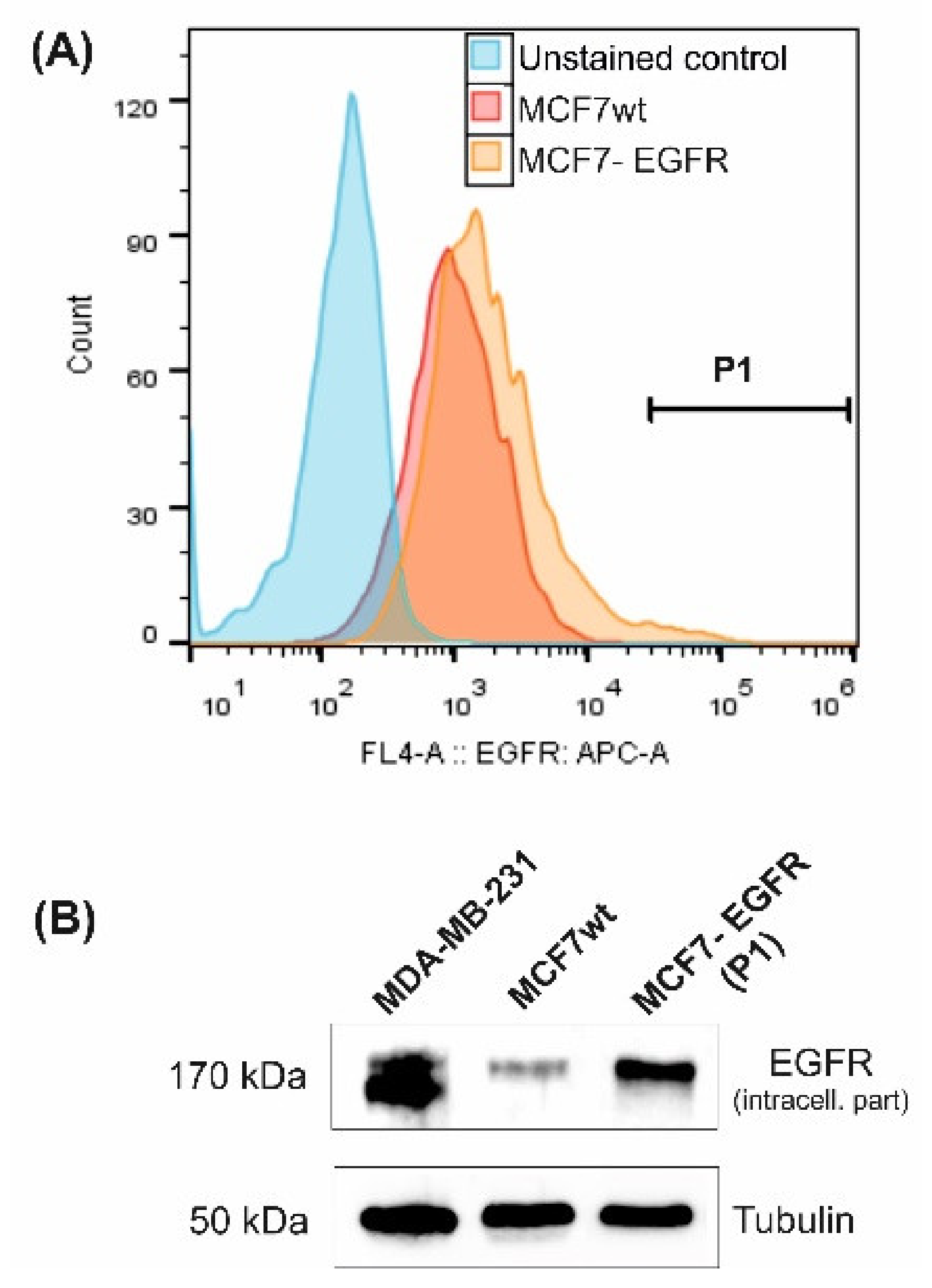
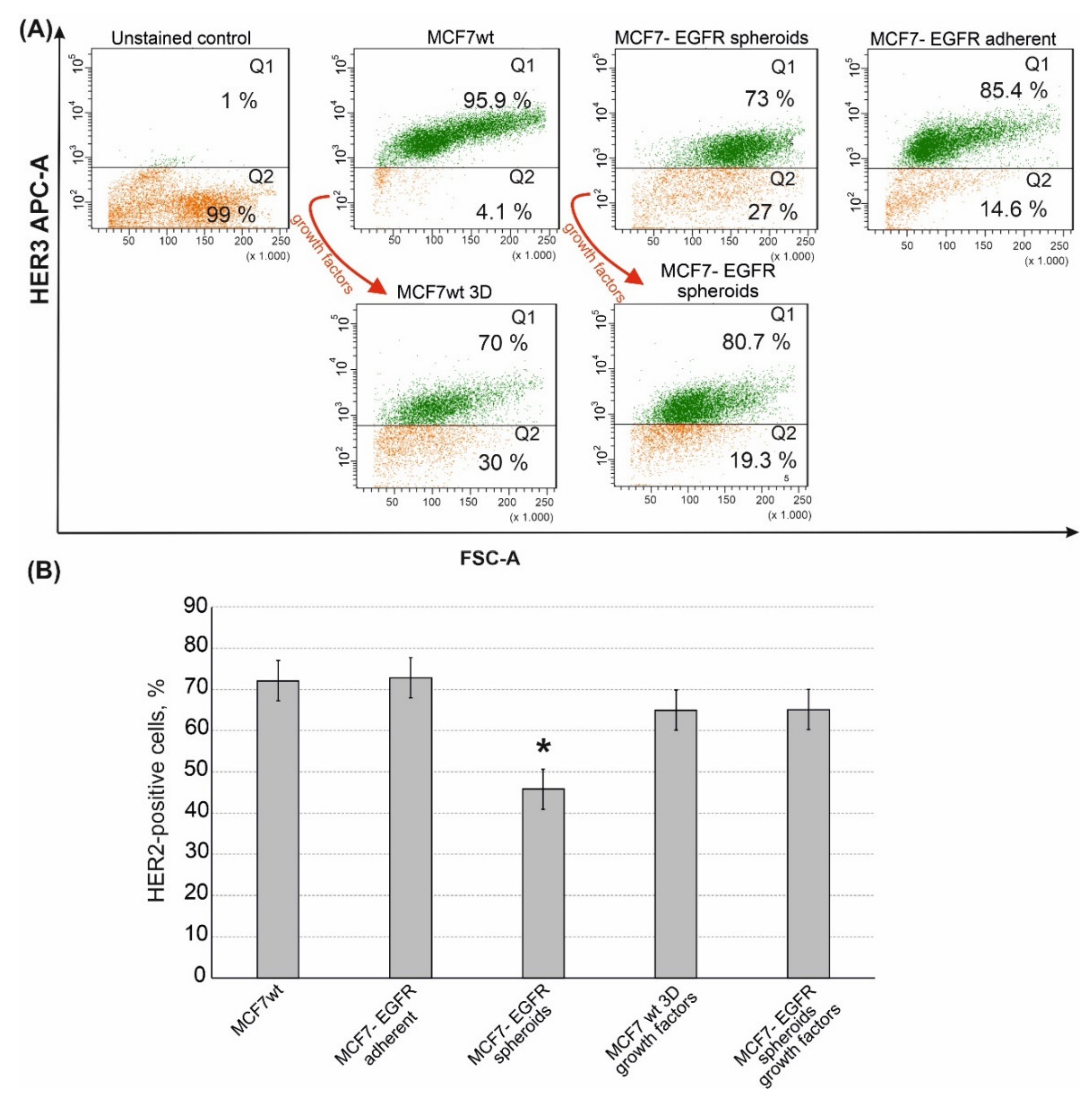
2.3. Sphere-Forming Ability Leads to the Partial Loss of HER3 Receptor
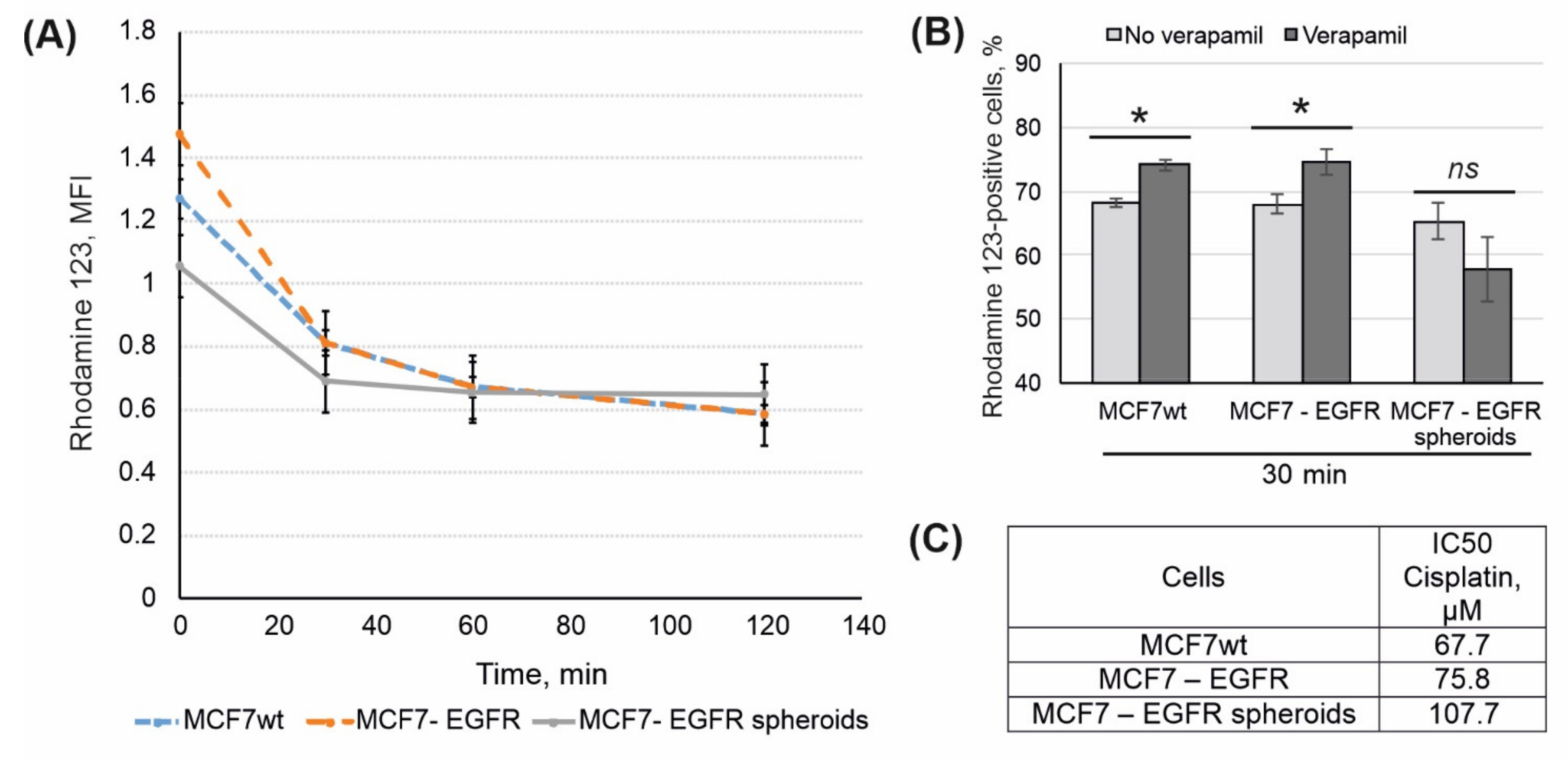
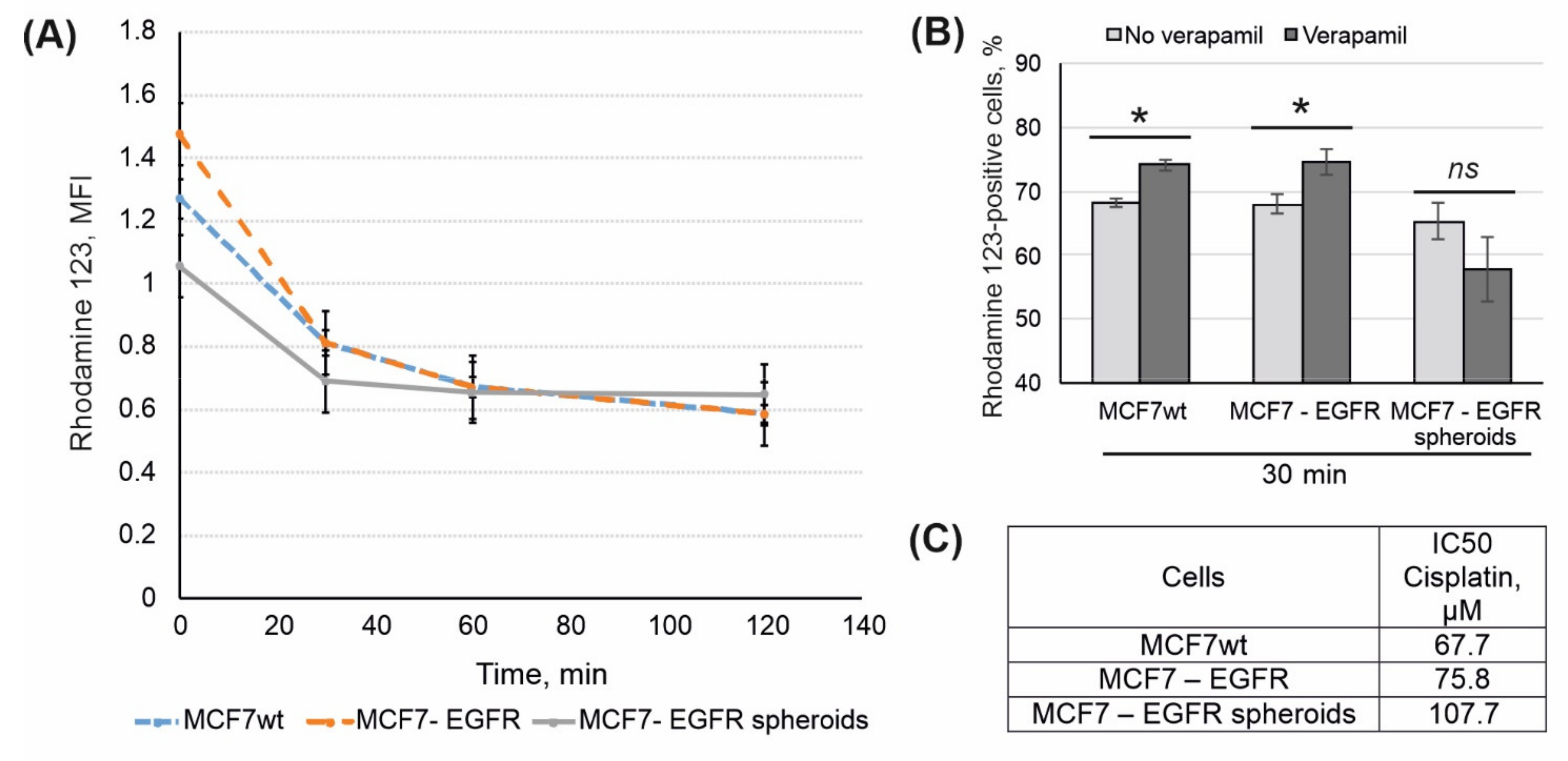
2.4. Rhodamine 123 Efflux
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. MCF7-EGFR Cells Construction
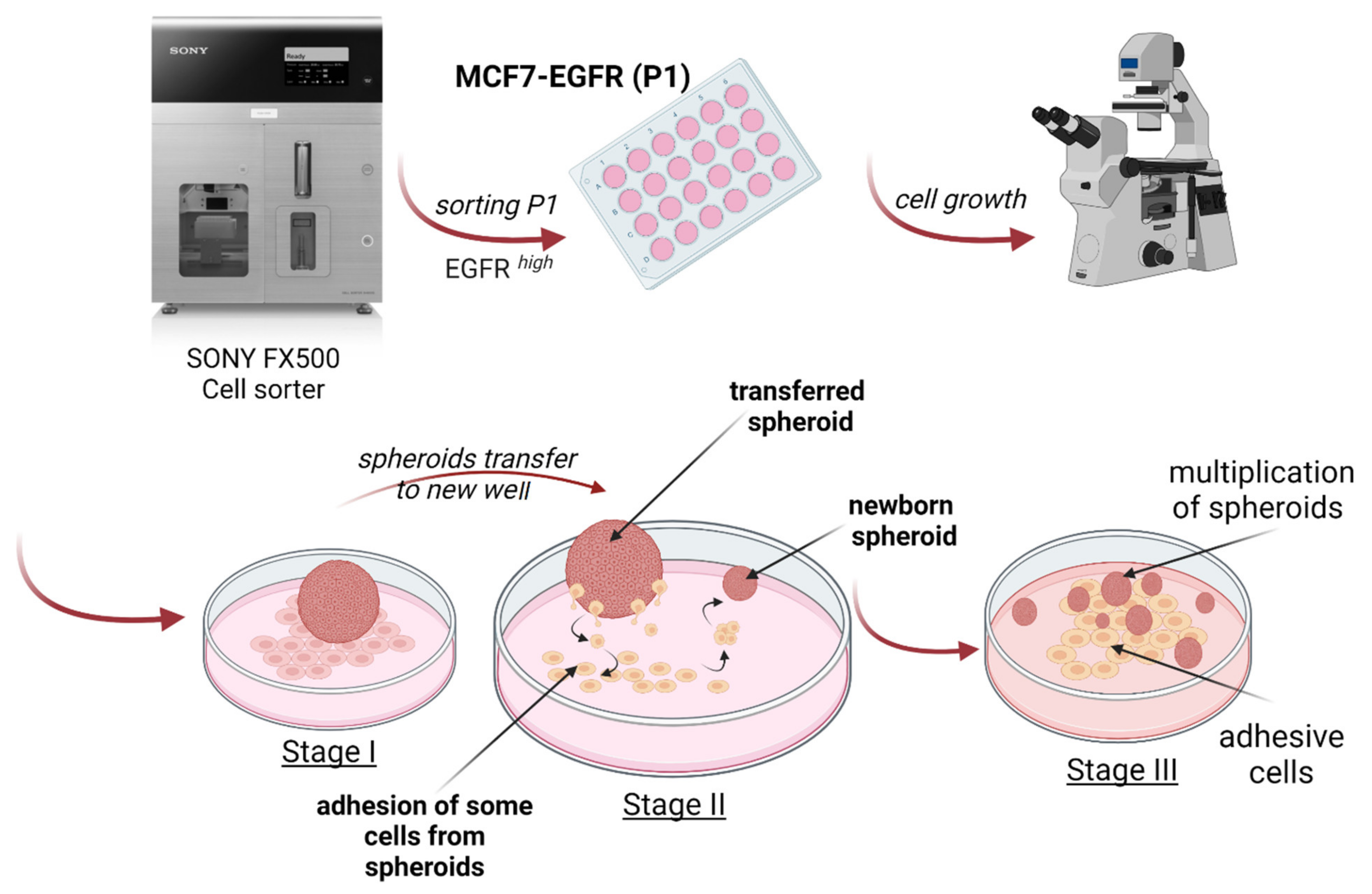
4.3. Spheroids Formation and Counting
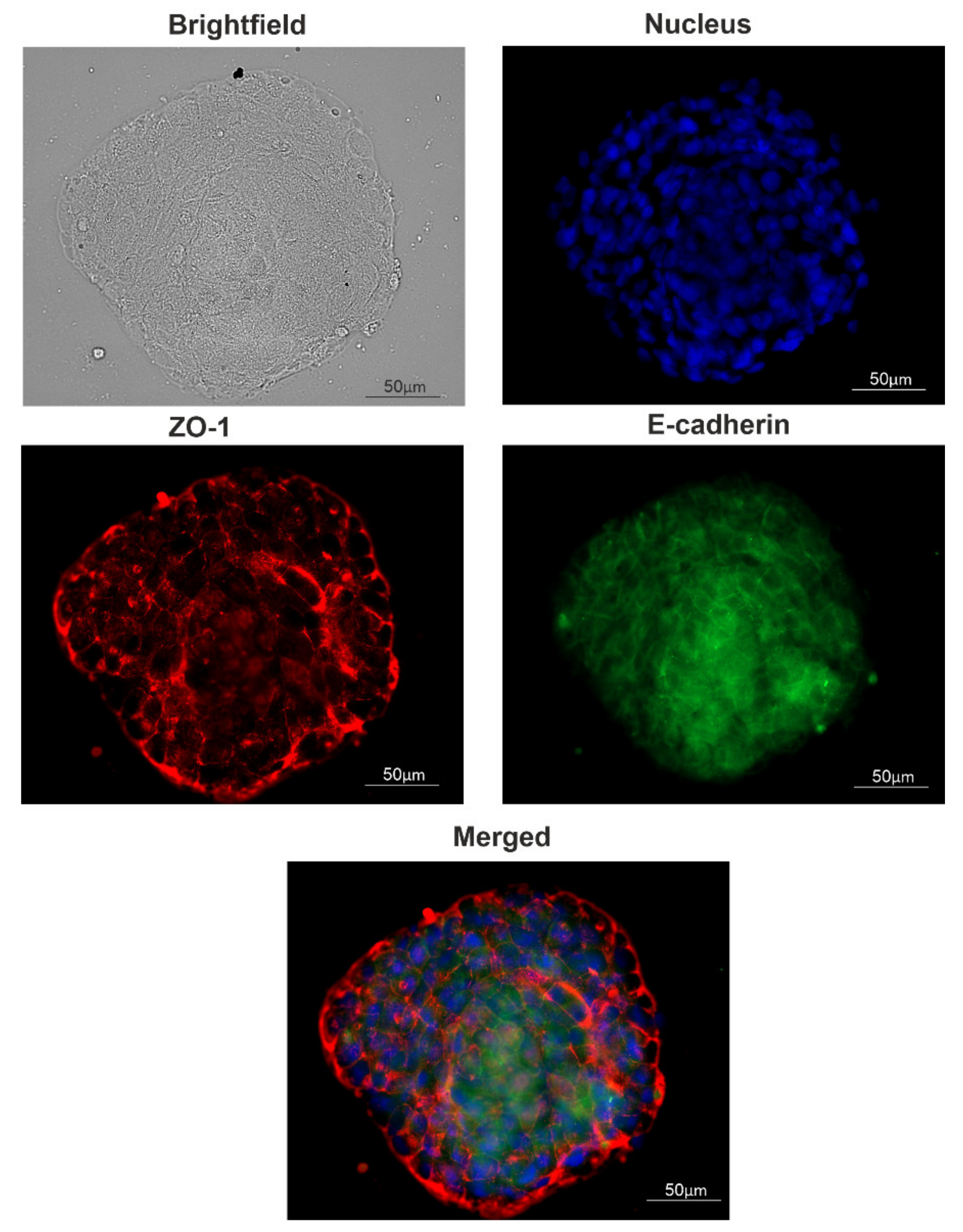
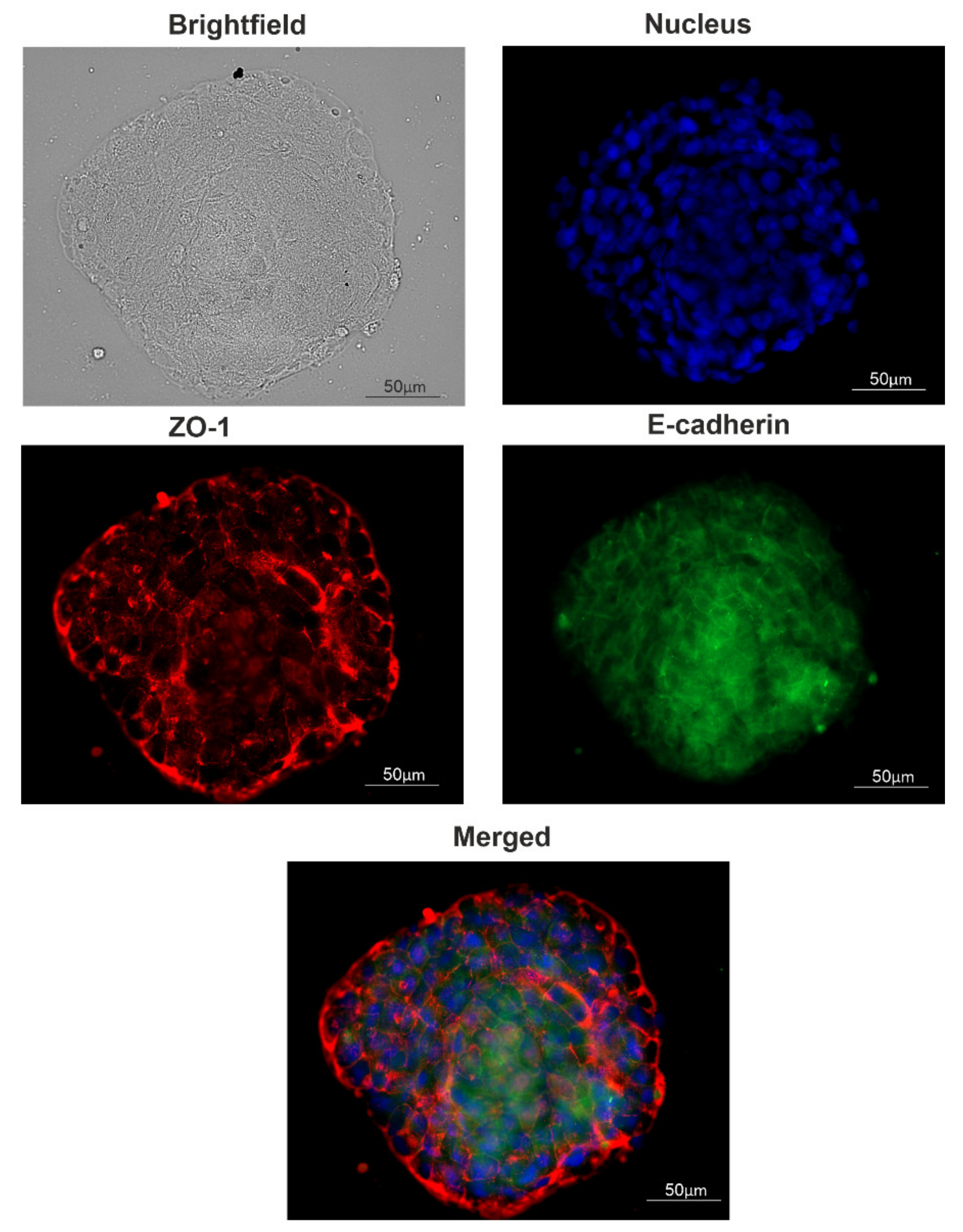
4.4. Immunocytochemistry of Spheroids
4.5. Histological Analysis of Spheroids
4.6. Cell Killing Assay
4.7. Western Blot
4.8. Flow Cytometry
4.9. Rhodamine 123 Efflux
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaprin, A.D.; Starinsky, V.V.; Shahzadova, A.O. The State of Cancer Care for the Population of Russia in 2019; National Medical Research Radiological Centre of the Ministry of Health of the Russian Federation: Moscow, Russia, 2020; ISBN 978-5-85502-255-1. [Google Scholar]
- Białkowska, K.; Komorowski, P.; Bryszewska, M.; Miłowska, K. Spheroids as a Type of Three-Dimensional Cell Cultures—Examples of Methods of Preparation and the Most Important Application. Int. J. Mol. Sci. 2020, 21, 6225. [Google Scholar] [CrossRef]
- Lohberger, B.; Glaenzer, D.; Eck, N.; Steinecker-Frohnwieser, B.; Leithner, A.; Rinner, B.; Kerschbaum-Gruber, S.; Georg, D.; Lohberger, B.; Glaenzer, D.; et al. Effects of a Combined Therapy of Bortezomib and Ionizing Radiation on Chondrosarcoma Three-Dimensional Spheroid Cultures. Oncol. Lett. 2021, 21, 1–10. [Google Scholar] [CrossRef]
- Edmondson, R.; Broglie, J.J.; Adcock, A.F.; Yang, L. Three-Dimensional Cell Culture Systems and Their Applications in Drug Discovery and Cell-Based Biosensors. ASSAY Drug Dev. Technol. 2014, 12, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Lilly, G.D.; Doty, R.C.; Podsiadlo, P.; Kotov, N.A. In Vitro Toxicity Testing of Nanoparticles in 3D Cell Culture. Small 2009, 5, 1213–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravi, M.; Paramesh, V.; Kaviya, S.R.; Anuradha, E.; Solomon, F.D.P. 3D Cell Culture Systems: Advantages and Applications. J. Cell. Physiol. 2015, 230, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Denes, V.; Lakk, M.; Makarovskiy, A.; Jakso, P.; Szappanos, S.; Graf, L.; Mandel, L.; Karadi, I.; Geck, P. Metastasis Blood Test by Flow Cytometry: In Vivo Cancer Spheroids and the Role of Hypoxia. Int. J. Cancer 2015, 136, 1528–1536. [Google Scholar] [CrossRef]
- Froehlich, K.; Haeger, J.-D.; Heger, J.; Pastuschek, J.; Photini, S.M.; Yan, Y.; Lupp, A.; Pfarrer, C.; Mrowka, R.; Schleußner, E.; et al. Generation of Multicellular Breast Cancer Tumor Spheroids: Comparison of Different Protocols. J. Mammary Gland Biol. Neoplasia 2016, 21, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Clouthier, S.G.; Deol, Y.; Liu, S.; Nagrath, S.; Azizi, E.; Wicha, M.S. Breast Cancer Stem Cells: Current Advances and Clinical Implications. In Mammary Stem Cells: Methods and Protocols; del Vivanco, M., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2015; pp. 1–49. ISBN 978-1-4939-2519-3. [Google Scholar]
- Geng, S.-Q.; Alexandrou, A.T.; Li, J.J. Breast Cancer Stem Cells: Multiple Capacities in Tumor Metastasis. Cancer Lett. 2014, 349, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Najafi, M.; Farhood, B.; Mortezaee, K. Cancer Stem Cells (CSCs) in Cancer Progression and Therapy. J. Cell. Physiol. 2019, 234, 8381–8395. [Google Scholar] [CrossRef]
- Dragu, D.L.; Necula, L.G.; Bleotu, C.; Diaconu, C.C.; Chivu-Economescu, M. Therapies Targeting Cancer Stem Cells: Current Trends and Future Challenges. World J. Stem Cells 2015, 7, 1185–1201. [Google Scholar] [CrossRef]
- Zhao, L.; Si, C.-S.; Yu, Y.; Lu, J.-W.; Zhuang, Y. Depletion of DNA Damage Binding Protein 2 Sensitizes Triple-Negative Breast Cancer Cells to Poly ADP-Ribose Polymerase Inhibition by Destabilizing Rad51. Cancer Sci. 2019, 110, 3543–3552. [Google Scholar] [CrossRef]
- Boesch, M.; Zeimet, A.G.; Rumpold, H.; Gastl, G.; Sopper, S.; Wolf, D. Drug Transporter-Mediated Protection of Cancer Stem Cells From Ionophore Antibiotics. STEM CELLS Transl. Med. 2015, 4, 1028–1032. [Google Scholar] [CrossRef]
- Najafi, M.; Mortezaee, K.; Majidpoor, J. Cancer Stem Cell (CSC) Resistance Drivers. Life Sci. 2019, 234, 116781. [Google Scholar] [CrossRef] [PubMed]
- Kelm, J.M.; Timmins, N.E.; Brown, C.J.; Fussenegger, M.; Nielsen, L.K. Method for Generation of Homogeneous Multicellular Tumor Spheroids Applicable to a Wide Variety of Cell Types. Biotechnol. Bioeng. 2003, 83, 173–180. [Google Scholar] [CrossRef]
- Braunholz, D.; Saki, M.; Niehr, F.; Öztürk, M.; Puértolas, B.B.; Konschak, R.; Budach, V.; Tinhofer, I. Spheroid Culture of Head and Neck Cancer Cells Reveals an Important Role of EGFR Signalling in Anchorage Independent Survival. PLoS ONE 2016, 11, e0163149. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z. ErbB Receptors and Cancer. Methods Mol. Biol. 2017, 1652, 3–35. [Google Scholar] [CrossRef] [PubMed]
- McGaffin, K.R.; Acktinson, L.E.; Chrysogelos, S.A. Growth and EGFR Regulation in Breast Cancer Cells by Vitamin D and Retinoid Compounds. Breast Cancer Res. Treat. 2004, 86, 55–73. [Google Scholar] [CrossRef]
- Kim, J.; Jung, J.; Lee, S.-J.; Lee, J.-S.; Park, M.-J. Cancer Stem-like Cells Persist in Established Cell Lines through Autocrine Activation of EGFR Signaling. Oncol. Lett. 2012, 3, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Gazit, A.; Chen, J.; App, H.; McMahon, G.; Hirth, P.; Chen, I.; Levitzki, A. Tyrphostins IV—Highly Potent Inhibitors of EGF Receptor Kinase. Structure-Activity Relationship Study of 4-Anilidoquinazolines. Bioorganic Med. Chem. 1996, 4, 1203–1207. [Google Scholar] [CrossRef]
- Xie, G.; Yao, Q.; Liu, Y.; Du, S.; Liu, A.; Guo, Z.; Sun, A.; Ruan, J.; Chen, L.; Ye, C.; et al. IL-6-Induced Epithelial-Mesenchymal Transition Promotes the Generation of Breast Cancer Stem-like Cells Analogous to Mammosphere Cultures. Int. J. Oncol. 2012, 40, 1171–1179. [Google Scholar] [CrossRef] [Green Version]
- Bahmad, H.F.; Cheaito, K.; Chalhoub, R.M.; Hadadeh, O.; Monzer, A.; Ballout, F.; El-Hajj, A.; Mukherji, D.; Liu, Y.-N.; Daoud, G.; et al. Sphere-Formation Assay: Three-Dimensional in Vitro Culturing of Prostate Cancer Stem/Progenitor Sphere-Forming Cells. Front. Oncol. 2018, 8, 347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vantangoli, M.M.; Madnick, S.J.; Huse, S.M.; Weston, P.; Boekelheide, K. MCF-7 Human Breast Cancer Cells Form Differentiated Microtissues in Scaffold-Free Hydrogels. PLoS ONE 2015, 10, e0135426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulze, L.; Congiu, T.; Brevini, T.A.L.; Grimaldi, A.; Tettamanti, G.; D’Antona, P.; Baranzini, N.; Acquati, F.; Ferraro, F.; de Eguileor, M. MCF7 Spheroid Development: New Insight about Spatio/Temporal Arrangements of TNTs, Amyloid Fibrils, Cell Connections, and Cellular Bridges. Int. J. Mol. Sci. 2020, 21, 5400. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, D.S.; Tevis, K.M.; Blessing, W.A.; Colson, Y.L.; Zaman, M.H.; Grinstaff, M.W. Breast Cancer Spheroids Reveal a Differential Cancer Stem Cell Response to Chemotherapeutic Treatment. Sci. Rep. 2017, 7, 10382. [Google Scholar] [CrossRef]
- Normanno, N.; De Luca, A.; Bianco, C.; Strizzi, L.; Mancino, M.; Maiello, M.R.; Carotenuto, A.; De Feo, G.; Caponigro, F.; Salomon, D.S. Epidermal Growth Factor Receptor (EGFR) Signaling in Cancer. Gene 2006, 366, 2–16. [Google Scholar] [CrossRef]
- Hanawa, M.; Suzuki, S.; Dobashi, Y.; Yamane, T.; Kono, K.; Enomoto, N.; Ooi, A. EGFR Protein Overexpression and Gene Amplification in Squamous Cell Carcinomas of the Esophagus. Int. J. Cancer 2006, 118, 1173–1180. [Google Scholar] [CrossRef]
- Masuda, H.; Zhang, D.; Bartholomeusz, C.; Doihara, H.; Hortobagyi, G.N.; Ueno, N.T. Role of Epidermal Growth Factor Receptor in Breast Cancer. Breast Cancer Res. Treat. 2012, 136, 331–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otani, T.; Furuse, M. Tight Junction Structure and Function Revisited. Trends Cell Biol. 2020, 30, 805–817. [Google Scholar] [CrossRef]
- Morrison, B.J.; Steel, J.C.; Morris, J.C. Sphere Culture of Murine Lung Cancer Cell Lines Are Enriched with Cancer Initiating Cells. PLoS ONE 2012, 7, e49752. [Google Scholar] [CrossRef] [PubMed]
- Nushtaeva, A.A.; Stepanov, G.A.; Semenov, D.V.; Juravlev, E.S.; Balahonova, E.A.; Gerasimov, A.V.; Sidorov, S.V.; Savelyev, E.I.; Kuligina, E.V.; Richter, V.A.; et al. Characterization of Primary Normal and Malignant Breast Cancer Cell and Their Response to Chemotherapy and Immunostimulatory Agents. BMC Cancer 2018, 18, 728. [Google Scholar] [CrossRef]
- Qiao, X.; Zhang, Y.; Sun, L.; Ma, Q.; Yang, J.; Ai, L.; Xue, J.; Chen, G.; Zhang, H.; Ji, C.; et al. Association of Human Breast Cancer CD44-/CD24- Cells with Delayed Distant Metastasis. eLife 2021, 10, e65418. [Google Scholar] [CrossRef] [PubMed]
- Litviakov, N.; Ibragimova, M.; Tsyganov, M.; Kazantseva, P.; Deryusheva, I.; Pevzner, A.; Doroshenko, A.; Garbukov, E.; Tarabanovskaya, N.; Slonimskaya, E. Amplifications of Stemness Genes and the Capacity of Breast Tumors for Metastasis. Oncotarget 2020, 11, 1988–2001. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Chen, Y.; Yao, Y.; Zhang, H.; Wang, T. Increased Invasion and Tumorigenicity Capacity of CD44+/CD24- Breast Cancer MCF7 Cells in Vitro and in Nude Mice. Cancer Cell Int. 2013, 13, 62. [Google Scholar] [CrossRef] [Green Version]
- Senbanjo, L.T.; Chellaiah, M.A. CD44: A Multifunctional Cell Surface Adhesion Receptor Is a Regulator of Progression and Metastasis of Cancer Cells. Front. Cell Dev. Biol. 2017, 5, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nushtaeva, A.A.; Karpushina, A.A.; Ermakov, M.S.; Gulyaeva, L.F.; Gerasimov, A.V.; Sidorov, S.V.; Gayner, T.A.; Yunusova, A.Y.; Tkachenko, A.V.; Richter, V.A.; et al. Establishment of Primary Human Breast Cancer Cell Lines Using “Pulsed Hypoxia” Method and Development of Metastatic Tumor Model in Immunodeficient Mice. Cancer Cell Int. 2019, 19, 46. [Google Scholar] [CrossRef]
- Chun, S.-Y.; Kwon, Y.-S.; Nam, K.-S.; Kim, S. Lapatinib Enhances the Cytotoxic Effects of Doxorubicin in MCF-7 Tumorspheres by Inhibiting the Drug Efflux Function of ABC Transporters. Biomed. Pharm. 2015, 72, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Koval, O.; Kochneva, G.; Tkachenko, A.; Troitskaya, O.; Sivolobova, G.; Grazhdantseva, A.; Nushtaeva, A.; Kuligina, E.; Richter, V. Recombinant Vaccinia Viruses Coding Transgenes of Apoptosis-Inducing Proteins Enhance Apoptosis But Not Immunogenicity of Infected Tumor Cells. Biomed Res. Int. 2017, 2017, 3620510. [Google Scholar] [CrossRef] [Green Version]
- Simmons, A.; Alberola-Ila, J. Retroviral Transduction of T Cells and T Cell Precursors. Methods Mol. Biol. 2016, 1323, 99–108. [Google Scholar] [CrossRef] [Green Version]
- O’Doherty, U.; Swiggard, W.J.; Malim, M.H. Human Immunodeficiency Virus Type 1 Spinoculation Enhances Infection through Virus Binding. J. Virol. 2000, 74, 10074–10080. [Google Scholar] [CrossRef] [Green Version]
- Mokhtari, R.B.; Qorri, B.; Sambi, M.; Baluch, N.; Kumar, S.; Das, B.; Szewczuk, M.R.; Yeger, H.; Cheng, H.-L.M. 3D Multicellular Stem-Like Human Breast Tumor Spheroids Enhance Tumorigenicity of Orthotopic Xenografts in Athymic Nude Rat Model. Cancers 2021, 13, 2784. [Google Scholar] [CrossRef]
- Tazzari, P.L.; Cappellini, A.; Ricci, F.; Evangelisti, C.; Papa, V.; Grafone, T.; Martinelli, G.; Conte, R.; Cocco, L.; McCubrey, J.A.; et al. Multidrug Resistance-Associated Protein 1 Expression Is under the Control of the Phosphoinositide 3 Kinase/Akt Signal Transduction Network in Human Acute Myelogenous Leukemia Blasts. Leukemia 2007, 21, 427–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Troitskaya, O.; Novak, D.; Nushtaeva, A.; Savinkova, M.; Varlamov, M.; Ermakov, M.; Richter, V.; Koval, O. EGFR Transgene Stimulates Spontaneous Formation of MCF7 Breast Cancer Cells Spheroids with Partly Loss of HER3 Receptor. Int. J. Mol. Sci. 2021, 22, 12937. https://doi.org/10.3390/ijms222312937
Troitskaya O, Novak D, Nushtaeva A, Savinkova M, Varlamov M, Ermakov M, Richter V, Koval O. EGFR Transgene Stimulates Spontaneous Formation of MCF7 Breast Cancer Cells Spheroids with Partly Loss of HER3 Receptor. International Journal of Molecular Sciences. 2021; 22(23):12937. https://doi.org/10.3390/ijms222312937
Chicago/Turabian StyleTroitskaya, Olga, Diana Novak, Anna Nushtaeva, Maria Savinkova, Mikhail Varlamov, Mikhail Ermakov, Vladimir Richter, and Olga Koval. 2021. "EGFR Transgene Stimulates Spontaneous Formation of MCF7 Breast Cancer Cells Spheroids with Partly Loss of HER3 Receptor" International Journal of Molecular Sciences 22, no. 23: 12937. https://doi.org/10.3390/ijms222312937
APA StyleTroitskaya, O., Novak, D., Nushtaeva, A., Savinkova, M., Varlamov, M., Ermakov, M., Richter, V., & Koval, O. (2021). EGFR Transgene Stimulates Spontaneous Formation of MCF7 Breast Cancer Cells Spheroids with Partly Loss of HER3 Receptor. International Journal of Molecular Sciences, 22(23), 12937. https://doi.org/10.3390/ijms222312937