
Effect of 2-Cys Peroxiredoxins Inhibition on Redox Modifications of Bull Sperm Proteins


Abstract
:1. Introduction
2. Results
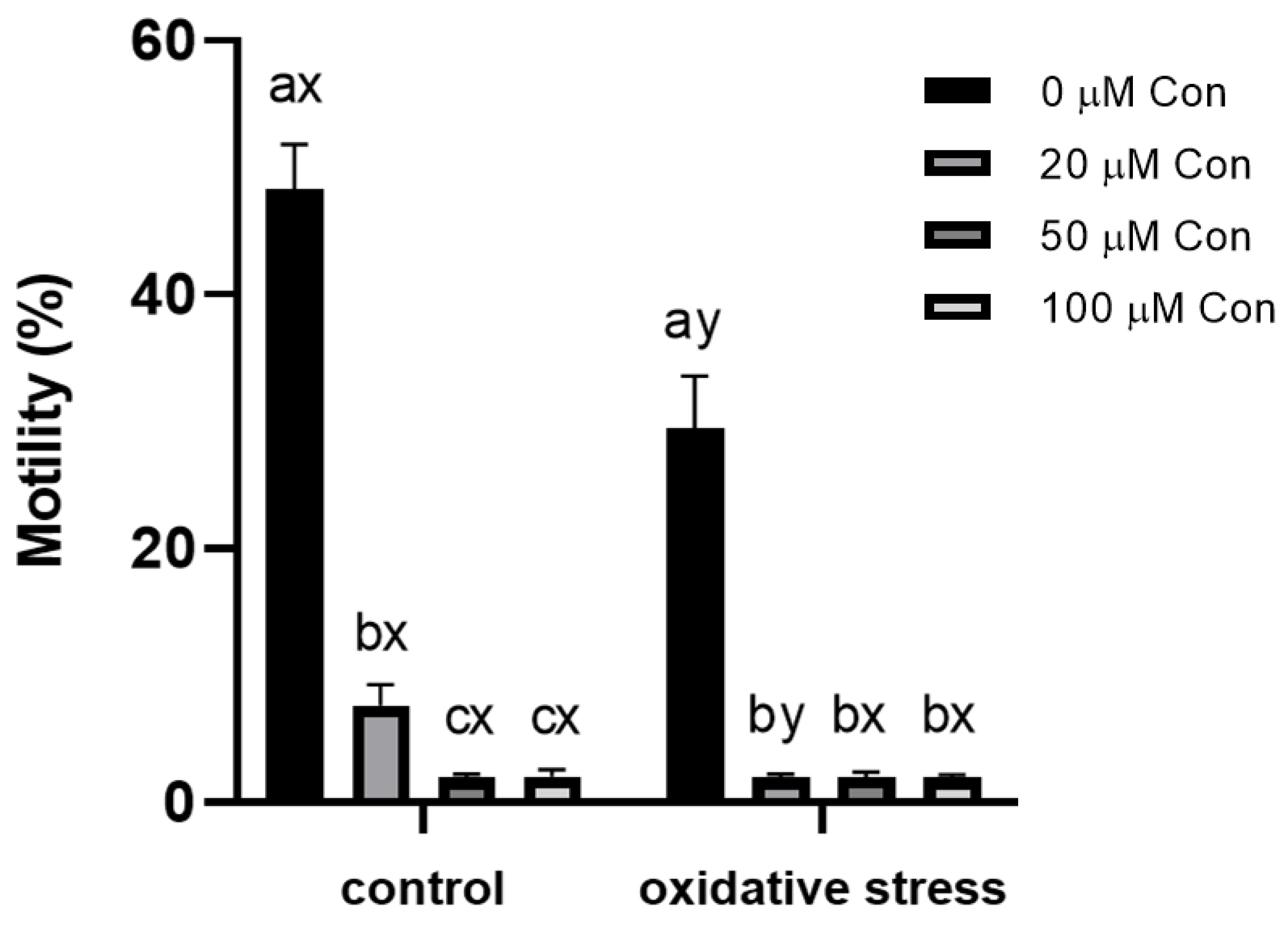
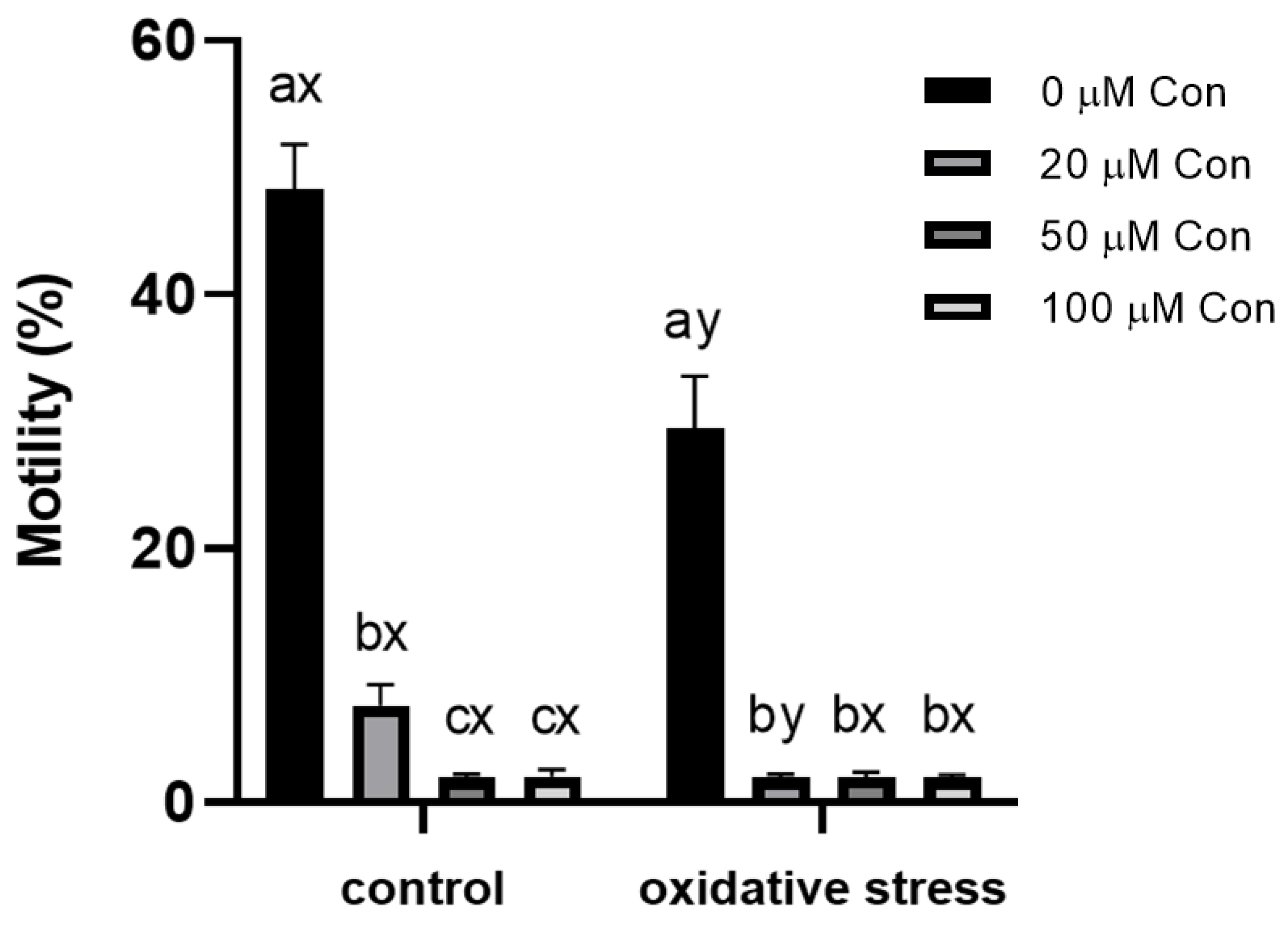
2.1. Sperm Motility
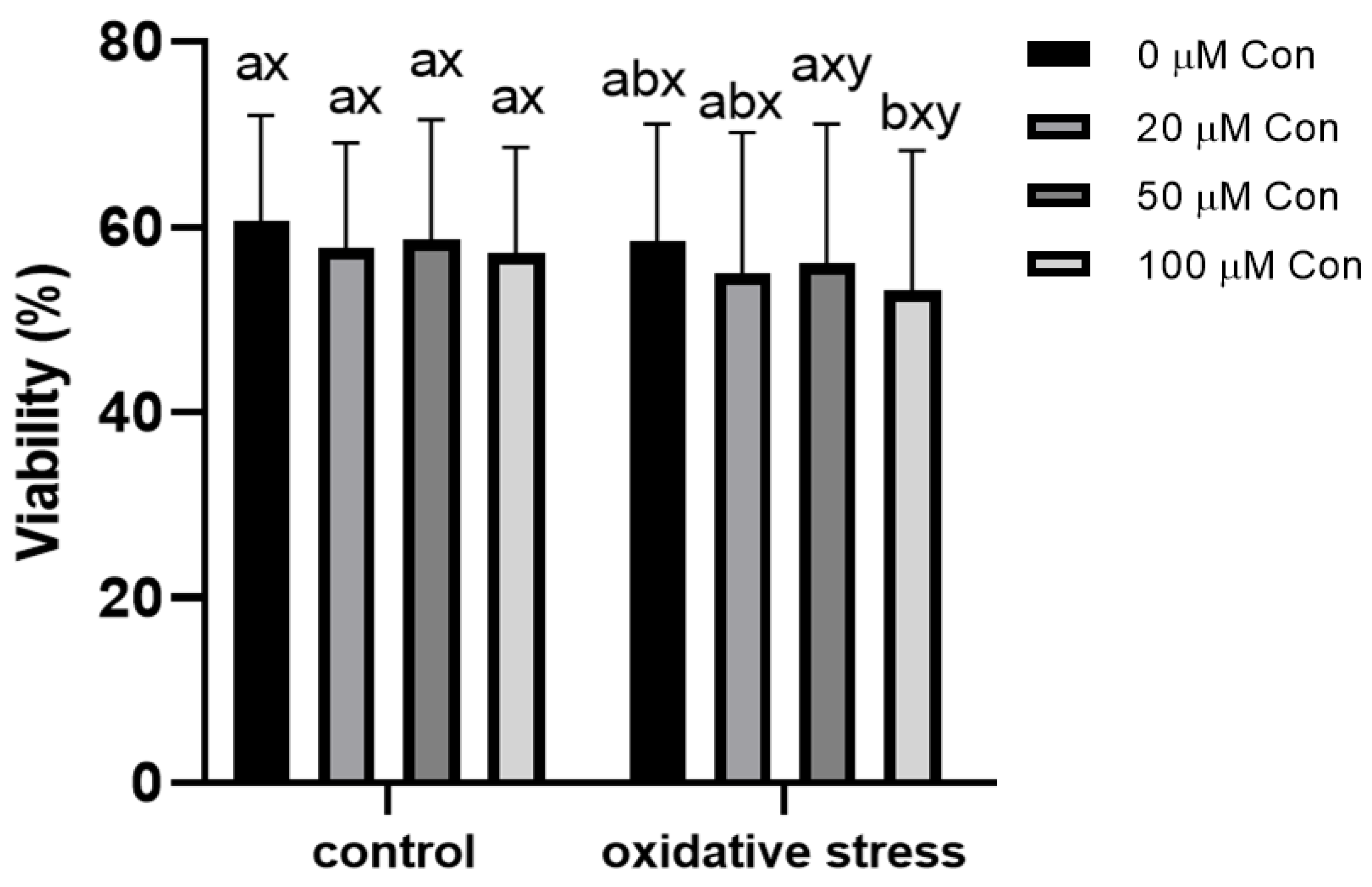
2.2. Sperm Viability
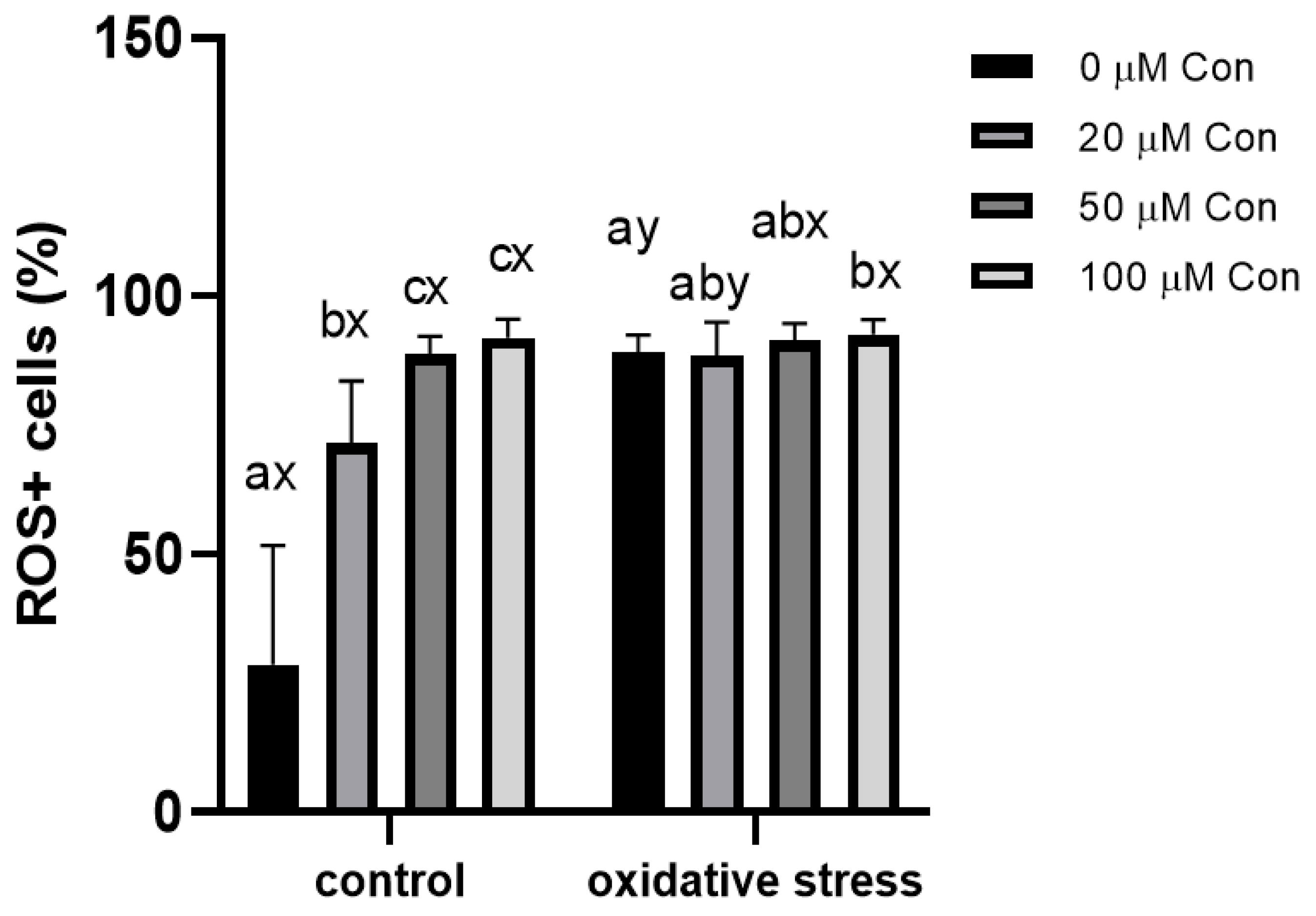
2.3. ROS-Positive Cell Content
2.4. Effect of Conoidin A on the Inhibition of Tyrosine Phosphorylation
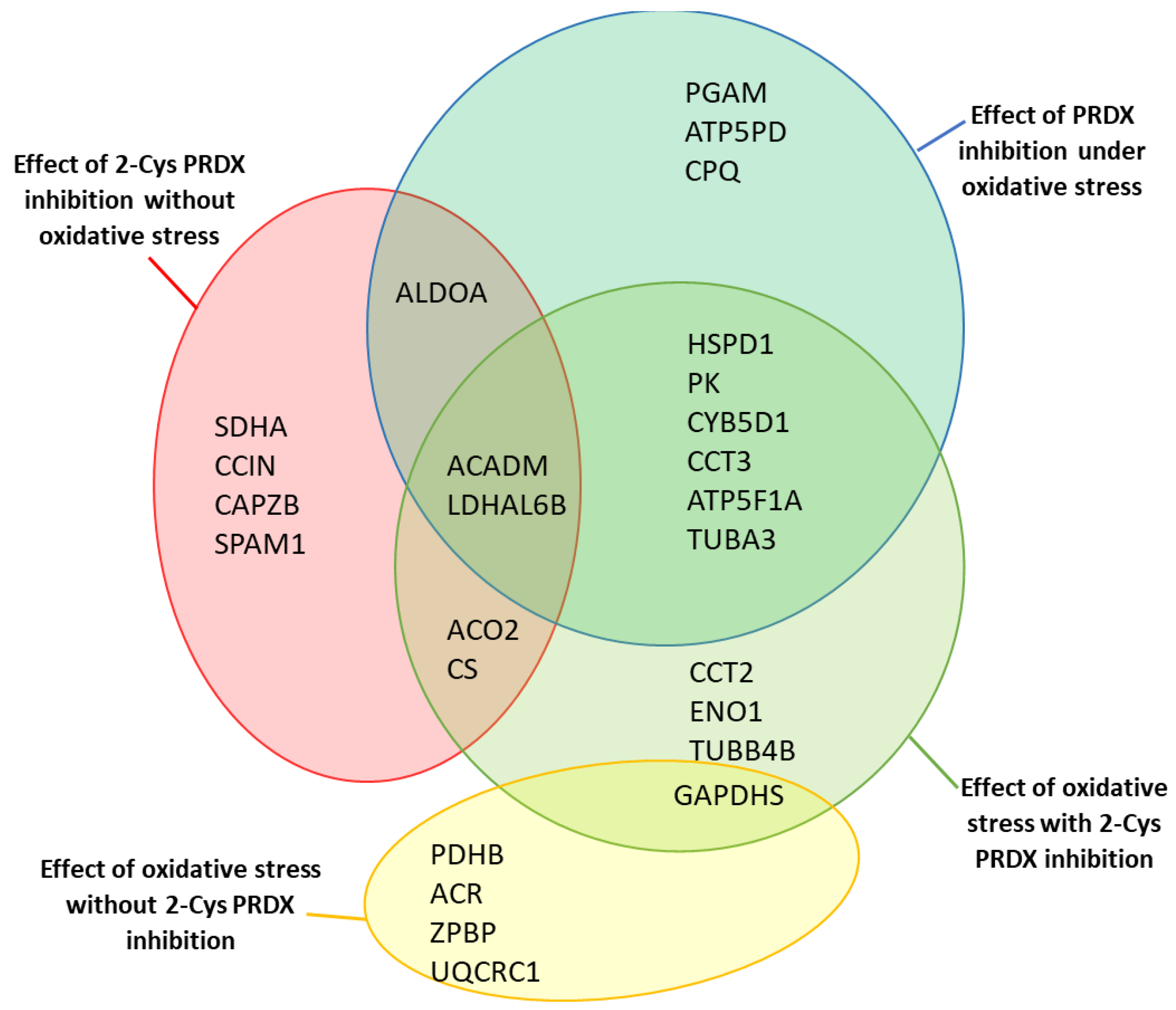
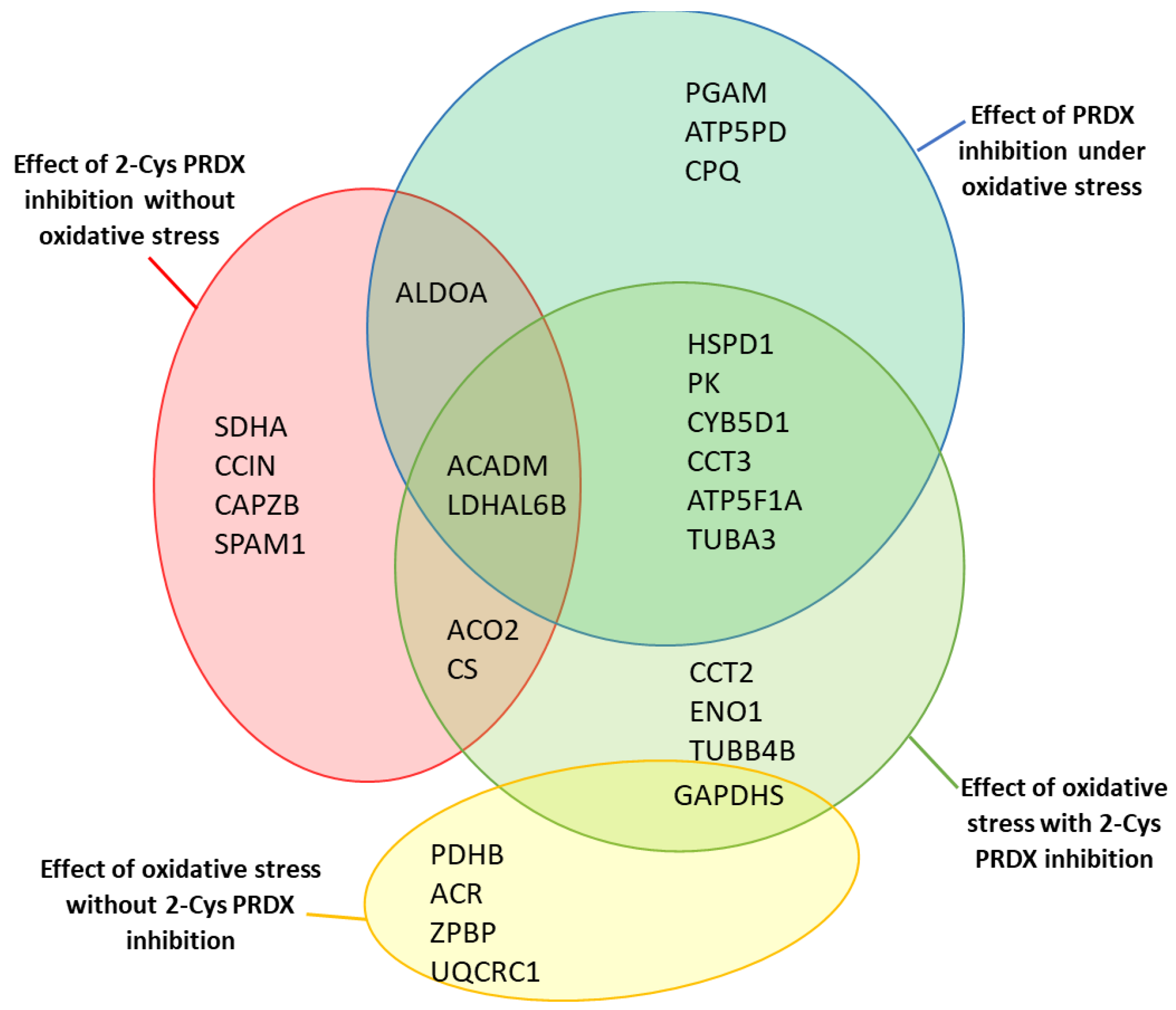
2.5. Identification of Proteins with Changed Levels of oxPTMs
2.6. Effect of 2-Cys PRDX Inhibition on the Level of Protein oxPTMs
2.7. Effect of Oxidative Stress on the Level of Protein oxPTMs
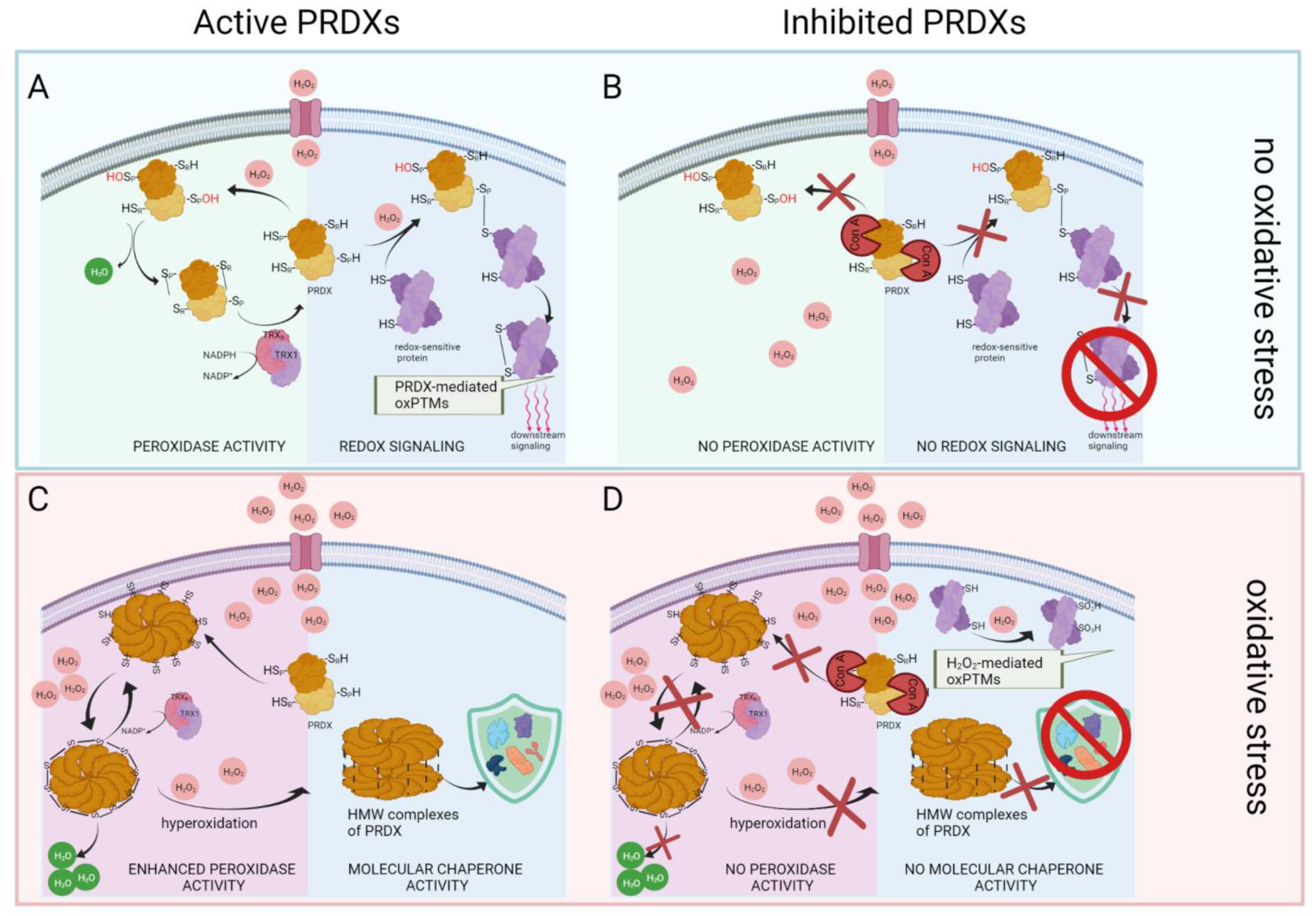
3. Discussion
3.1. Research Assumptions and Rationale for Experimental Design
3.2. Inhibition of 2-Cys PRDXs Affects ROS+ Spermatozoa Content
3.3. Inhibition of 2-Cys PRDXs Affects Sperm Motility without Changing Viability
3.4. Inhibition of PRDX Activity Results in Opposite Changes in the Level of Protein oxPTMs Depending on the Absence or Presence of Oxidative Stress
3.5. Inhibition of 2-Cys PRDXs in Non-Oxidative Conditions Leads to Decrease in Sperm Protein oxPTMs
3.6. Inhibition of 2-Cys PRDXs in Non-Oxidative Conditions Leads to a Decrease in Sperm Protein Tyrosine Phosphorylation
3.7. Oxidative Stress Causes Oxidation of Glycolytic Enzymes That Intensifies after Inhibition of PRDXs
3.8. Inhibition of 2-Cys PRDXs under Oxidative Stress Conditions Leads to an Increase in Sperm Protein oxPTM Level
4. Materials and Methods
4.1. Research Material
4.2. Sperm Preparation
4.3. 2-Cys PRDX Inhibition under Control or Oxidative Stress Conditions
4.4. Sperm Motility and Viability Measurement
4.5. Sperm Oxidative Status Evaluation
4.6. Protein Extraction
4.7. Blocking Free Thiols and Labeling Oxidized Cysteines
4.8. D-PAGE Separation of Fluorescently Labeled Proteins
4.9. Image Acquisition and Identification of Proteins
4.10. Evaluation of Tyrosine Phosphorylation of Bull Sperm
4.11. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chae, H.Z.; Kim, H.J.; Kang, S.W.; Rhee, S.G. Characterization of three isoforms of mammalian peroxiredoxin that reduce peroxides in the presence of thioredoxin. Diabetes Res. Clin. Pr. 1999, 45, 101–112. [Google Scholar] [CrossRef]
- Forshaw, T.E.; Holmila, R.; Nelson, K.J.; Lewis, J.E.; Kemp, M.L.; Tsang, A.W.; Poole, L.B.; Lowther, W.T.; Furdui, C.M. Peroxiredoxins in cancer and response to radiation therapies. Antioxidants 2019, 8, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O′Flaherty, C.; De Souza, A.R. Hydrogen peroxide modifies human sperm peroxiredoxins in a dose-dependent manner. Biol. Reprod. 2010, 84, 238–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastor-Flores, D.; Talwar, D.; Pedre, B.; Dick, T.P. Real-time monitoring of peroxiredoxin oligomerization dynamics in living cells. Proc. Natl. Acad. Sci. USA 2020, 117, 16313–16323. [Google Scholar] [CrossRef] [PubMed]
- Veal, E.A.; Day, A.M.; Morgan, B.A. Hydrogen peroxide sensing and signaling. Mol. Cell 2007, 26, 1–14. [Google Scholar] [CrossRef]
- Elko, E.A.; Manuel, A.M.; White, S.; Zito, E.; van der Vliet, A.; Anathy, V.; Janssen-Heininger, Y.M. Oxidation of peroxiredoxin-4 induces oligomerization and promotes interaction with proteins governing protein folding and endoplasmic reticulum stress. J. Biol. Chem. 2021, 296, 100665. [Google Scholar] [CrossRef] [PubMed]
- Stöcker, S.; Van Laer, K.; Mijuskovic, A.; Dick, T.P. The conundrum of hydrogen peroxide signaling and the emerging role of peroxiredoxins as redox relay hubs. Antioxid. Redox Signal. 2018, 28, 558–573. [Google Scholar] [CrossRef]
- O′Flaherty, C.; Matsushita-Fournier, D. Reactive oxygen species and protein modifications in spermatozoa. Biol. Reprod. 2017, 97, 577–585. [Google Scholar] [CrossRef]
- Sobotta, M.C.; Liou, W.; Stöcker, S.; Talwar, D.; Oehler, M.; Ruppert, T.; Scharf, A.N.D.; Dick, T.P. Peroxiredoxin-2 and STAT3 form a redox relay for H2O2 signaling. Nat. Chem. Biol. 2015, 11, 64–70. [Google Scholar] [CrossRef]
- Fisher, A.B. Peroxiredoxin 6 in the repair of peroxidized cell membranes and cell signaling. Arch. Biochem. Biophys. 2017, 617, 68–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, M.C.; O′Flaherty, C. Peroxiredoxin 6 is the primary antioxidant enzyme for the maintenance of viability and DNA integrity in human spermatozoa. Hum. Reprod. 2018, 33, 1394–1407. [Google Scholar] [CrossRef] [PubMed]
- Scarlata, E.; O′Flaherty, C. Antioxidant enzymes and male fertility: Lessons from knockout models. Antioxid. Redox Signal. 2020, 32, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Bolduc, J.; Koruza, K.; Luo, T.; Pueyo, J.M.; Vo, T.N.; Ezeriņa, D.; Messens, J. Peroxiredoxins wear many hats: Factors that fashion their peroxide sensing personalities. Redox Biol. 2021, 42, 101959. [Google Scholar] [CrossRef]
- Jang, H.H.; Lee, K.O.; Chi, Y.H.; Jung, B.G.; Park, S.K.; Park, J.H.; Lee, J.R.; Lee, S.S.; Moon, J.C.; Yun, J.W.; et al. Two enzymes in one: Two yeast peroxiredoxins display oxidative stress-dependent switching from a peroxidase to a Molecular chaperone function. Cell 2004, 117, 625–635. [Google Scholar] [CrossRef]
- Hall, A.; Karplus, P.A.; Poole, L.B. Typical 2-Cys peroxiredoxins—Structures, mechanisms and functions. FEBS J. 2009, 276, 2469–2477. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Moawad, A.; Morielli, T.; Fernandez, M.C.; O′Flaherty, C. Peroxiredoxins prevent oxidative stress during human sperm capacitation. Mol. Hum. Reprod. 2016, 23, 106–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutovsky, P.; Aarabi, M.; Miranda-Vizuete, A.; Oko, R. Negative biomarker based male fertility evaluation: Sperm phenotypes associated with molecular-level anomalies. Asian J. Androl. 2015, 17, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Manandhar, G.; Miranda-Vizuete, A.; Pedrajas, J.R.; Krause, W.J.; Zimmerman, S.; Sutovsky, M.; Sutovsky, P. Peroxiredoxin 2 and peroxidase enzymatic activity of mammalian spermatozoa. Biol. Reprod. 2009, 80, 1168–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godoy, J.R.; Funke, M.; Ackermann, W.; Haunhorst, P.; Oesteritz, S.; Capani, F.; Elsässer, H.-P.; Lillig, C.H. Redox atlas of the mouse: Immunohistochemical detection of glutaredoxin-, peroxiredoxin-, and thioredoxin-family proteins in various tissues of the laboratory mouse. Biochim. Biophys. Acta BBA Gen. Subj. 2011, 1810, 2–92. [Google Scholar] [CrossRef]
- Ryu, D.-Y.; Kim, K.-U.; Kwon, W.-S.; Rahman, S.; Khatun, A.; Pang, M.-G. Peroxiredoxin activity is a major landmark of male fertility. Sci. Rep. 2017, 7, 17174. [Google Scholar] [CrossRef] [Green Version]
- Moawad, A.R.; Fernandez, M.C.; Scarlata, E.; Dodia, C.; Feinstein, S.I.; Fisher, A.B.; O′Flaherty, C. Deficiency of peroxiredoxin 6 or inhibition of its phospholipase A2 activity impair the in vitro sperm fertilizing competence in mice. Sci. Rep. 2017, 7, 12994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, D.; Pang, W.; Rahman, M.; Park, Y.; Pang, M. Peroxiredoxin 4 directly affects the male fertility outcome in porcine. Theriogenology 2021, 171, 85–93. [Google Scholar] [CrossRef]
- O′Flaherty, C. Redox regulation of mammalian sperm capacitation. Asian J. Androl. 2015, 17, 583–590. [Google Scholar] [CrossRef]
- Haraldsen, J.D.; Liu, G.; Botting, C.H.; Walton, J.G.A.; Storm, J.; Phalen, T.J.; Kwok, L.Y.; Soldati-Favre, D.; Heintz, N.H.; Müller, S.; et al. Identification of conoidin A as a covalent inhibitor of peroxiredoxin II. Org. Biomol. Chem. 2009, 7, 3040–3048. [Google Scholar] [CrossRef] [Green Version]
- Netto, L.E.S.; Antunes, F. The roles of peroxiredoxin and thioredoxin in hydrogen peroxide sensing and in signal transduction. Mol. Cells 2016, 39, 65. [Google Scholar]
- Latimer, H.R.; Veal, E.A. Peroxiredoxins in regulation of MAPK signalling pathways; sensors and barriers to signal transduction. Mol. Cells 2016, 39, 40–45. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Kim, T.J.; Lee, K.-Y. A novel function of peroxiredoxin 1 (Prx-1) in apoptosis signal-regulating kinase 1 (ASK1)-mediated signaling pathway. FEBS Lett. 2008, 582, 1913–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarvis, R.M.; Hughes, S.; Ledgerwood, E.C. Peroxiredoxin 1 functions as a signal peroxidase to receive, transduce, and transmit peroxide signals in mammalian cells. Free Radic. Biol. Med. 2012, 53, 1522–1530. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.-S.; Cho, C.-S.; Park, S.; Yu, S.; Kang, S.W.; Rhee, S.G. Peroxiredoxin III, a mitochondrion-specific peroxidase, regulates apoptotic signaling by mitochondria. J. Biol. Chem. 2004, 279, 41975–41984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, J.A.; Wood, S.T.; Bolduc, J.; Nurmalasari, N.D.; Chubinskaya, S.; Poole, L.; Furdui, C.M.; Nelson, K.J.; Loeser, R.F. Differential peroxiredoxin hyperoxidation regulates MAP kinase signaling in human articular chondrocytes. Free Radic. Biol. Med. 2019, 134, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Somyajit, K.; Gupta, R.; Sedlackova, H.; Neelsen, K.J.; Ochs, F.; Rask, M.-B.; Choudhary, C.; Lukas, J. Redox-sensitive alteration of replisome architecture safeguards genome integrity. Science 2017, 358, 797–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chae, H.Z.; Uhm, T.B.; Rhee, S.G. Dimerization of thiol-specific antioxidant and the essential role of cysteine 47. Proc. Natl. Acad. Sci. USA 1994, 91, 7022–7026. [Google Scholar] [CrossRef] [Green Version]
- O′Flaherty, C.; Beorlegui, N.; Beconi, M.T. Participation of superoxide anion in the capacitation of cryopreserved bovine sperm. Int. J. Androl. 2003, 26, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, P.; O′Flaherty, C.; Beconi, M.; Beorlegui, N. Nitric oxide-induced capacitation of cryopreserved bull spermatozoa and assessment of participating regulatory pathways. Anim. Reprod. Sci. 2005, 85, 231–242. [Google Scholar] [CrossRef]
- De Lamirande, E.; O′Flaherty, C. Sperm activation: Role of reactive oxygen species and kinases. Biochim. Biophys. Acta Proteins Proteom. 2008, 1784, 106–115. [Google Scholar] [CrossRef]
- Fernández, L.G.; Ortega-Ferrusola, C.; Garcia, B.M.; Salido, G.M.; Peña, F.J.; Tapia, J.A. Identification of protein tyrosine phosphatases and dual-specificity phosphatases in mammalian spermatozoa and their role in sperm motility and protein tyrosine phosphorylation. Biol. Reprod. 2009, 80, 1239–1252. [Google Scholar] [CrossRef]
- Foster, D.B.; Van Eyk, J.E.; Marbán, E.; O′Rourke, B. Redox signaling and protein phosphorylation in mitochondria: Progress and prospects. J. Bioenerg. Biomembr. 2009, 41, 159–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, Y.; Okuno, M. Activation of mammalian sperm motility by regulation of microtubule sliding via cyclic adenosine 5′-monophosphate-dependent phosphorylation. Biol. Reprod. 1995, 53, 1081–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stincone, A.; Prigione, A.; Cramer, T.; Wamelink, M.M.C.; Campbell, K.; Cheung, E.; Olin-Sandoval, V.; Grüning, N.-M.; Krüger, A.; Alam, M.T.; et al. The return of metabolism: Biochemistry and physiology of the pentose phosphate pathway. Biol. Rev. 2014, 90, 927–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.; Umehara, T.; Tsujita, N.; Kawai, T.; Goto, M.; Cheng, B.; Zeng, W.; Shimada, M. Itaconate regulates the glycolysis/pentose phosphate pathway transition to maintain boar sperm linear motility by regulating redox homeostasis. Free Radic. Biol. Med. 2020, 159, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Grüning, N.-M.; Rinnerthaler, M.; Bluemlein, K.; Mülleder, M.; Wamelink, M.M.; Lehrach, H.; Jakobs, C.; Breitenbach, M.; Ralser, M. Pyruvate kinase triggers a metabolic feedback loop that controls redox metabolism in respiring cells. Cell Metab. 2011, 14, 415–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grüning, N.-M.; Du, D.; Keller, M.A.; Luisi, B.F.; Ralser, M. Inhibition of triosephosphate isomerase by phosphoenolpyruvate in the feedback-regulation of glycolysis. Open Biol. 2014, 4, 130232. [Google Scholar] [CrossRef] [Green Version]
- Miraglia, E.; Lussiana, C.; Viarisio, D.; Racca, C.; Cipriani, A.; Gazzano, E.; Bosia, A.; Revelli, A.; Ghigo, D. The pentose phosphate pathway plays an essential role in supporting human sperm capacitation. Fertil. Steril. 2010, 93, 2437–2440. [Google Scholar] [CrossRef]
- Qiu, J.-H.; Li, Y.-W.; Xie, H.-L.; Li, Q.; Dong, H.-B.; Sun, M.-J.; Gao, W.-Q.; Tan, J.-H. Effects of glucose metabolism pathways on sperm motility and oxidative status during long-term liquid storage of goat semen. Theriogenology 2016, 86, 839–849. [Google Scholar] [CrossRef] [PubMed]
- Evdokimov, V.V.; Barinova, K.V.; Turovetskii, V.B.; Muronetz, V.I.; Schmalhausen, E.V. Low concentrations of hydrogen peroxide activate the antioxidant defense system in human sperm cells. Biochemistry 2015, 80, 1178–1185. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.; Ford, W. Functional significance of the pentose phosphate pathway and glutathione reductase in the antioxidant defenses of human sperm. Biol. Reprod. 2004, 71, 1309–1316. [Google Scholar] [CrossRef] [PubMed]
- Mostek, A.; Dietrich, M.A.; Słowińska, M.; Ciereszko, A. Cryopreservation of bull semen is associated with carbonylation of sperm proteins. Theriogenology 2017, 92, 95–102. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot No. | Protein Name | Gene Name | Accession Number | Calculated MW/pI | Protein Score/Number of Peptides | No. of Cysteines |
|---|---|---|---|---|---|---|
| 101 | Hyaluronidase | SPAM1 | F1MTV1 NP_001008413.3 | 63268/8.79 | 244/2 | 18 |
| 102 | Hyaluronidase | SPAM1 | F1MTV1 NP_001008413.3 | 63268/8.79 | 353/3 | 18 |
| 103 | Pyruvate kinase | PKM | A5D984 | 58482/7.96 | 215/2 | 10 |
| 104 | T-complex protein 1 subunit beta | CCT2 | Q32BHO | 57781/6.18 | 187/2 | 6 |
| 183 | Aconitate hydratase, mitochondrial | ACO2 | A0A3Q1M6K6 XP_005909496.1 | 85481/7.67 | 164/1 | 13 |
| 184 | Aconitate hydratase, mitochondrial | ACO2 | A0A3Q1M6K6 XP_005909496.1 | 85481/7.67 | 164/1 | 13 |
| 185 | Aconitate hydratase, mitochondrial | ACO2 | P20004 XP_005909496.1 | 86047/7.87 | 244/1 | 13 |
| 286 | T-complex protein 1 subunit gamma | CCT3 | Q3T0K2 ELR54279.1 | 61118/6.38 | 470/3 | 10 |
| 292 | Succinate dehydrogenase (ubiquinone) flavoprotein subunit, mitochondrial | SDHA | G3MY67 XP_027375926.1 | 74101/7.55 | 906/10 | 19 |
| 304 | Pyruvate kinase | PK | A5D984 | 58482/7.96 | 220/2 | 10 |
| 324 | Calicin | CCIN | Q28068 XP_005908650.1 | 67873/8.49 | 414/4 | 18 |
| 341 | Carboxypeptidase Q | CPQ | A0A3Q1LSW9 XP_005215732.1 | 52902/5.83 | 356/3 | 1 |
| 344 | 60 kDa chaperonin | HSPD1 | F1MUZ9 XP_015313986.1 | 61110/5.71 | 330/4 | 3 |
| 347 | 60 kDa chaperonin | HSPD1 | F1MUZ9 XP_015313986.1 | 61110/5.71 | 1010/7 | 3 |
| 349 | Carboxypeptidase Q | CPQ | A0A3Q1MFF3 | 49530/5.52 | 188/2 | 5 |
| 394 | ATP synthase subunit alpha | ATP5F1A | F1MLB8 XP_019842301.1 | 59767/9.21 | 399/2 | 2 |
| 397 | ATP synthase subunit alpha | ATP5F1A | F1MLB8 XP_019842301.1 | 59767/9.21 | 496/4 | 2 |
| 422 | Acrosin | ACR | G5E5C6 | 46149/9.64 | 185/2 | 12 |
| 431 | Alpha-enolase | ENO1 | Q9XSJ4 NP_776474.2 | 47639/6.37 | 291/3 | 6 |
| 444 | Fructose-bisphosphate aldolase | ALDOA | A6QLL8 | 39925/8.45 | 223/2 | 9 |
| 449 | Glyceraldehyde-3-phosphate dehydrogenase, testis-specific | GAPDHS | Q2KJE5 | 43659/8.32 | 152/2 | 5 |
| 454 | Cytochrome b-c1 complex subunit 1, mitochondrial | UQCRC1 | P13272 | 29813/9.04 | 393/3 | 5 |
| 481 | Fructose-bisphosphate aldolase | ALDOA | A6QLL8 XP_006070551.2 | 39925/8.45 | 283/3 | 9 |
| 483 | Citrate synthase, mitochondrial | CS | Q29RK1 | 51968/8.16 | 378/4 | 4 |
| 484 | Phosphoglycerate mutase 2 | PGAM2 | Q32KV0 NP_001033200.1 | 28838/8.99 | 203/2 | 3 |
| 488 | Medium-chain specific acyl-CoA dehydrogenase, mitochondrial | ACADM | Q3SZB4 NP_001068703.1 | 46943/8.31 | 505/4 | 7 |
| 542 | Zona pellucida binding protein | ZPBP | F1N369 | 40313/9.14 | 312/2 | 15 |
| 559 | Pyruvate dehydrogenase E1 component subunit beta, mitochondrial | PDHB | P11966 | 39443/6.21 | 271/3 | 6 |
| 612 | F-actin-capping protein subunit beta | CAPZB | A0A3Q1MG13 XP_005203140.1 | 34782/5.59 | 336/2 | 7 |
| 620 | L-lactate dehydrogenase A-like 6B | LDHAL6B | Q3T056 | 42022/8.91 | 379/4 | 8 |
| 627 | L-lactate dehydrogenase A-like 6B | LDHAL6B | Q3T056 NP_001030352.1 | 42022/8.91 | 379/4 | 8 |
| 725 | Cytochrome b5 heme-binding domain-containing protein | CYB5D1 | A0A3Q1LQK0 | 26704/5.36 | 194/3 | 1 |
| 775 | ATP synthase subunit d, mitochondrial | ATP5PD | P13620 | 18738/5.99 | 436/5 | 1 |
| 801 | Tubulin alpha-3 chain isoform X3 | TUBA3 | XP_024833647.1 | 46605/4.99 | 828/7 | 9 |
| 802 | Tubulin beta-4B chain | TUBB4B | NP_001029835.1 | 50255/4.79 | 498/6 | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mostek-Majewska, A.; Janta, A.; Majewska, A.; Ciereszko, A. Effect of 2-Cys Peroxiredoxins Inhibition on Redox Modifications of Bull Sperm Proteins. Int. J. Mol. Sci. 2021, 22, 12888. https://doi.org/10.3390/ijms222312888
Mostek-Majewska A, Janta A, Majewska A, Ciereszko A. Effect of 2-Cys Peroxiredoxins Inhibition on Redox Modifications of Bull Sperm Proteins. International Journal of Molecular Sciences. 2021; 22(23):12888. https://doi.org/10.3390/ijms222312888
Chicago/Turabian StyleMostek-Majewska, Agnieszka, Anna Janta, Anna Majewska, and Andrzej Ciereszko. 2021. "Effect of 2-Cys Peroxiredoxins Inhibition on Redox Modifications of Bull Sperm Proteins" International Journal of Molecular Sciences 22, no. 23: 12888. https://doi.org/10.3390/ijms222312888
APA StyleMostek-Majewska, A., Janta, A., Majewska, A., & Ciereszko, A. (2021). Effect of 2-Cys Peroxiredoxins Inhibition on Redox Modifications of Bull Sperm Proteins. International Journal of Molecular Sciences, 22(23), 12888. https://doi.org/10.3390/ijms222312888