
Cardiomyogenic Differentiation Potential of Human Dilated Myocardium-Derived Mesenchymal Stem/Stromal Cells: The Impact of HDAC Inhibitor SAHA and Biomimetic Matrices

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
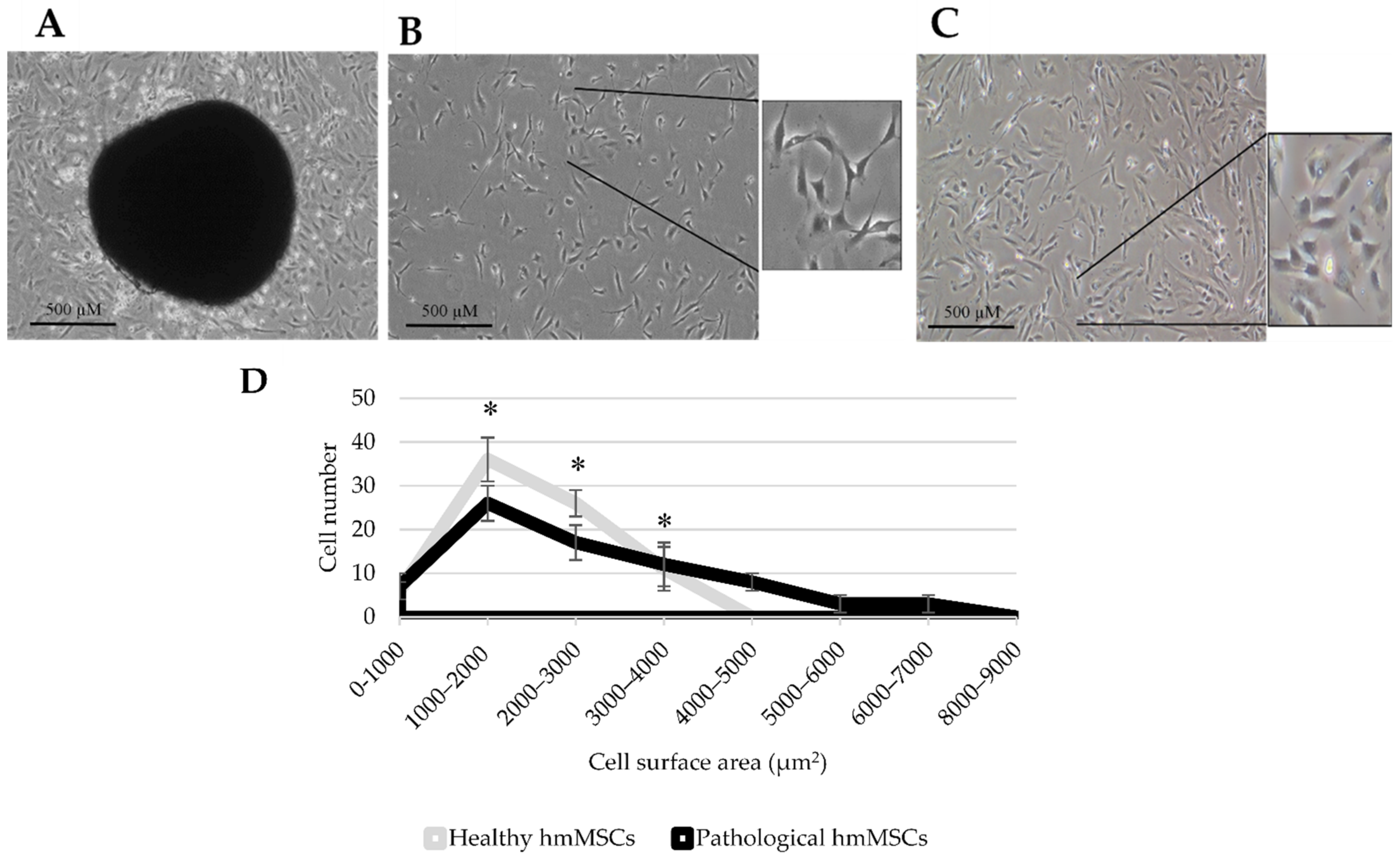
2.1. Cell Isolation and Morphology
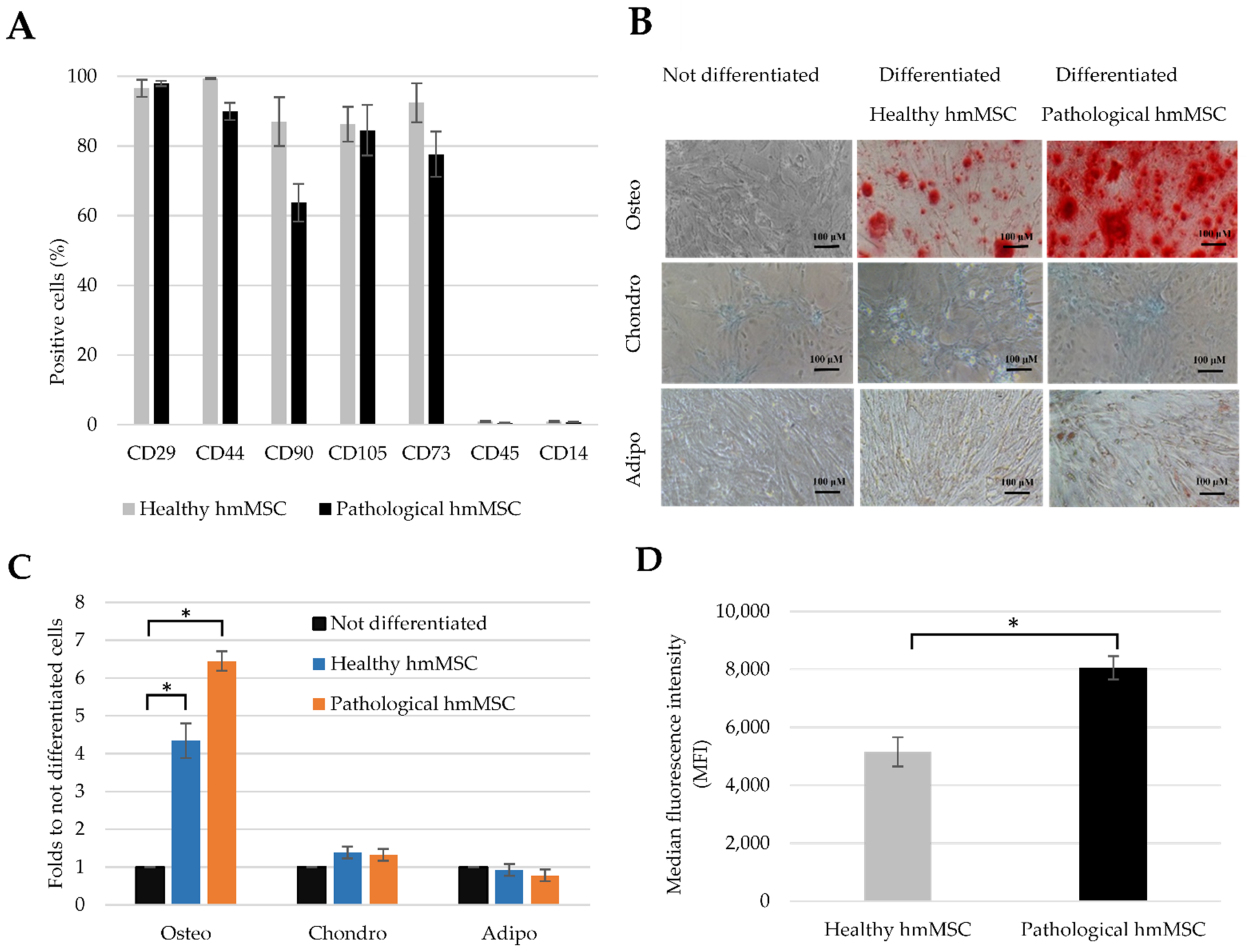
2.2. Characterization of Healthy and Dilated Myocardium-Derived hmMSC
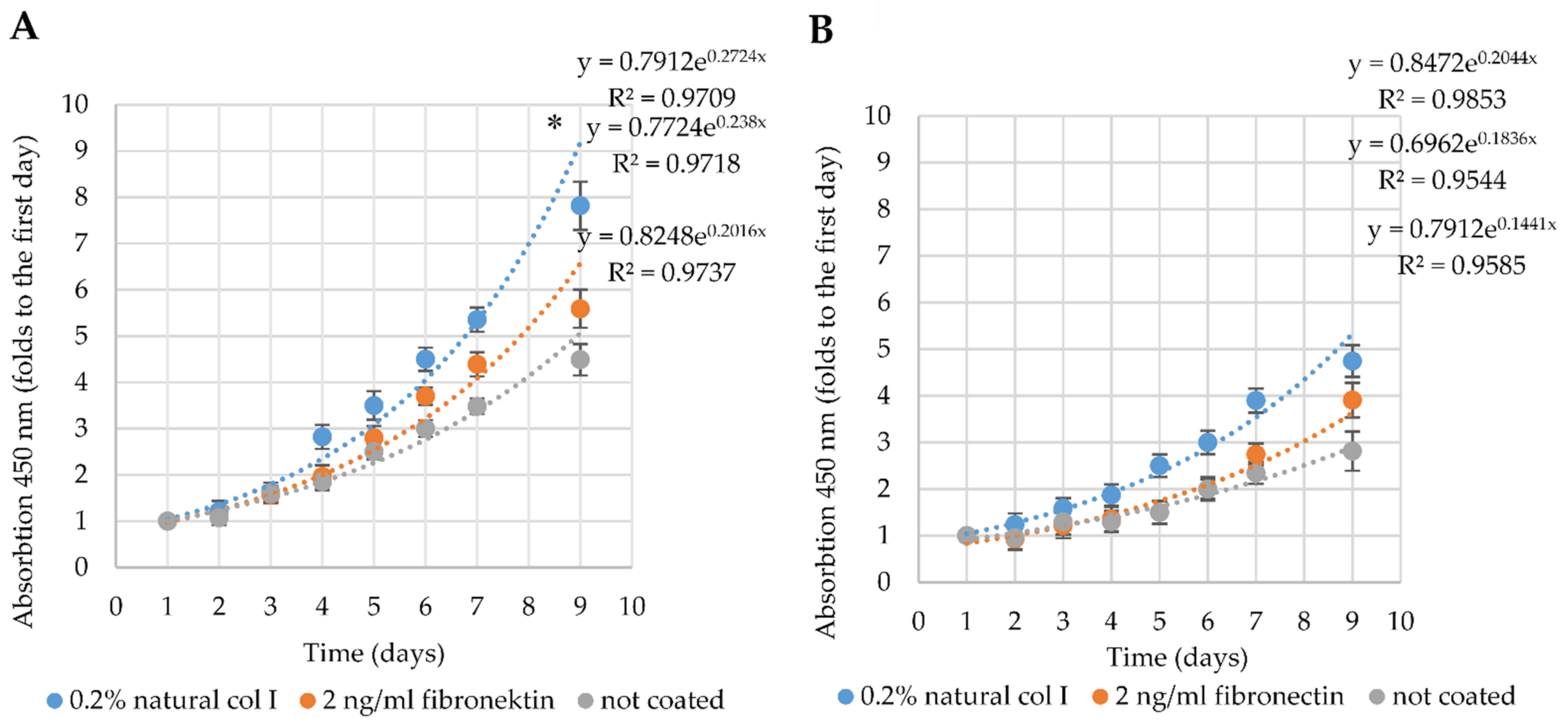
2.3. Proliferation and Cardiomyogenic Differentiation of hmMSC on ECM Components-Precoated Plastic Surfaces
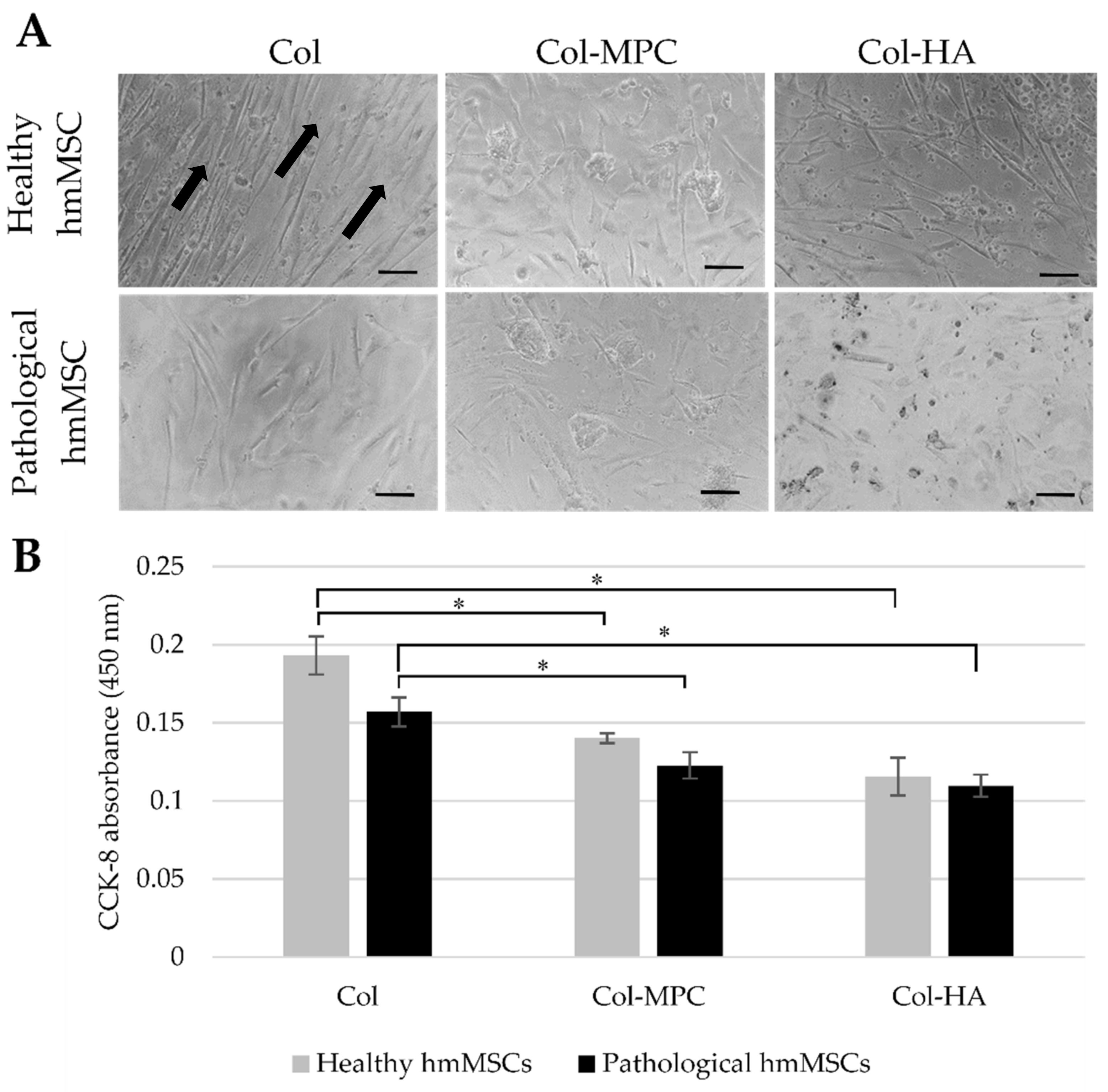
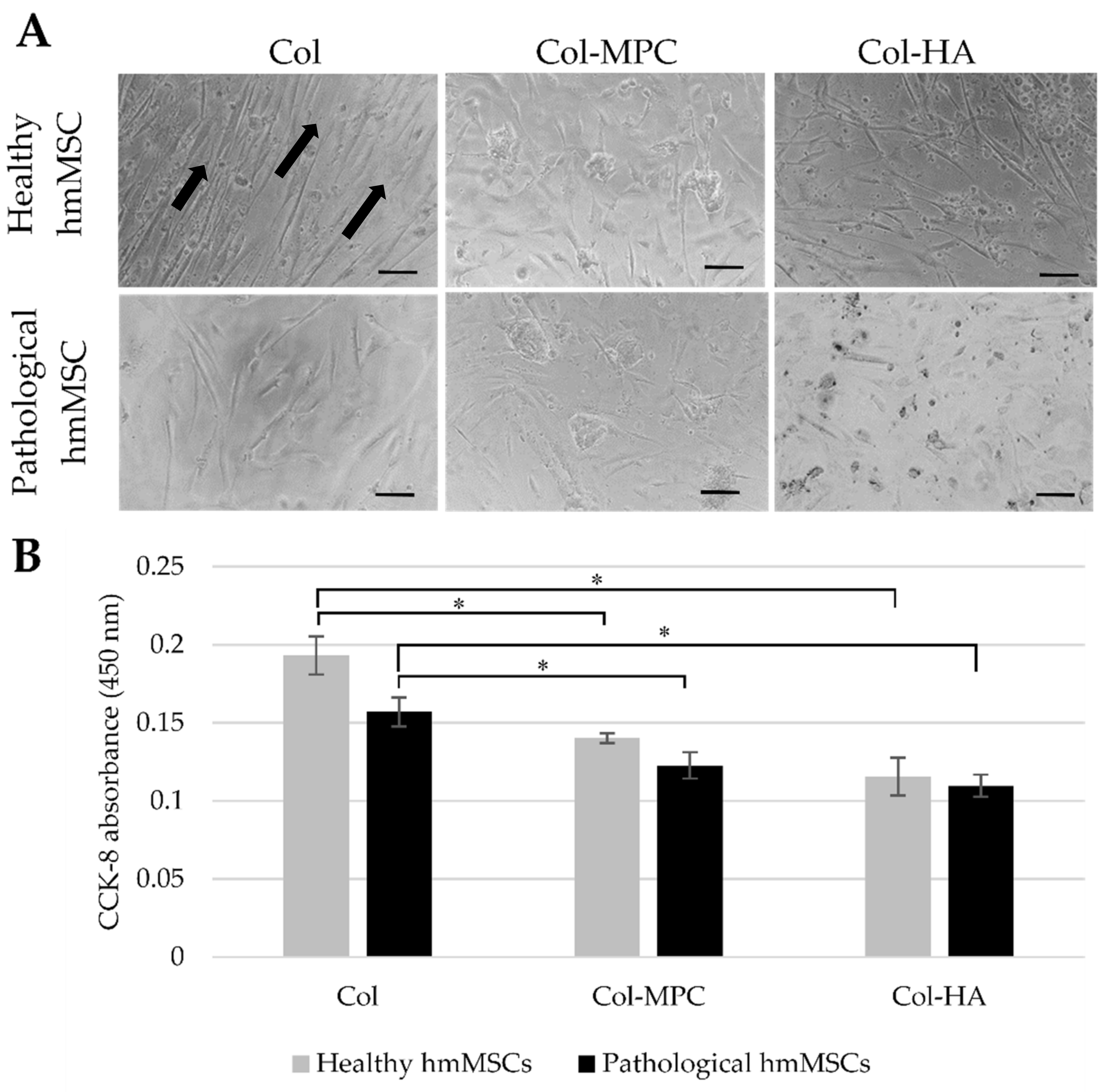
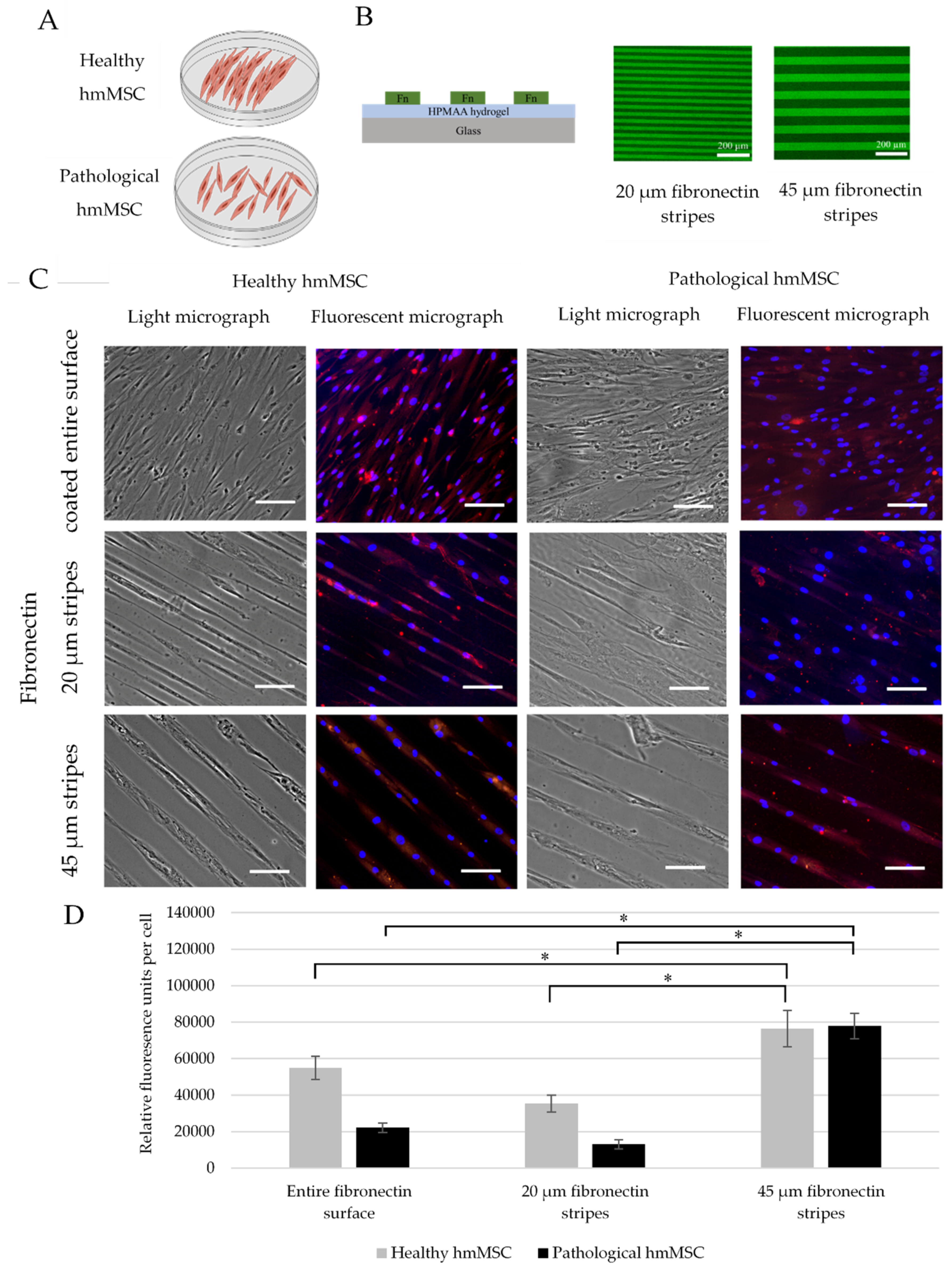
2.4. The Attachment and Growth of Healthy and Dilated Myocardium-Derived hmMSC on Hybrid Collagen I Hydrogels
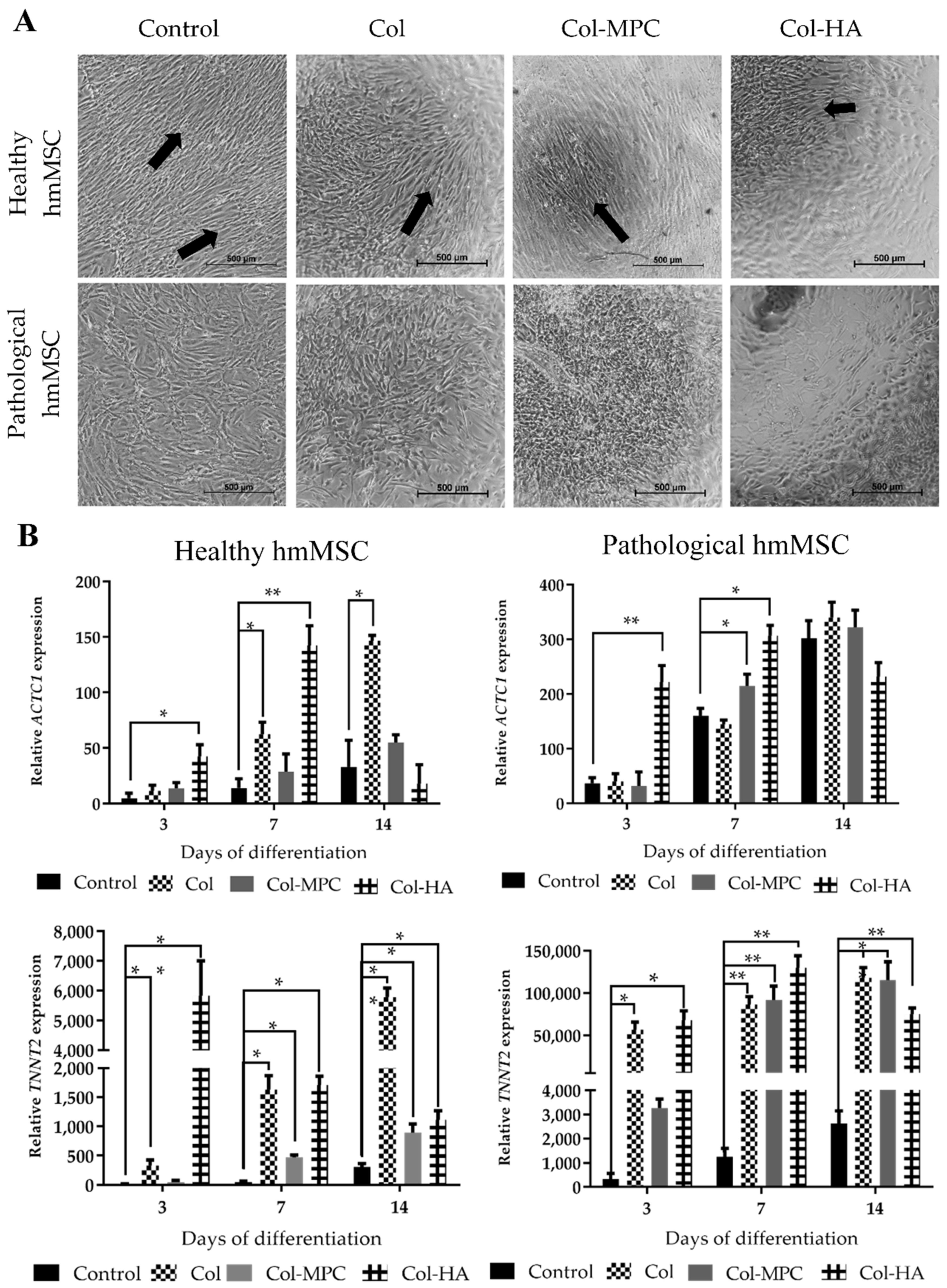
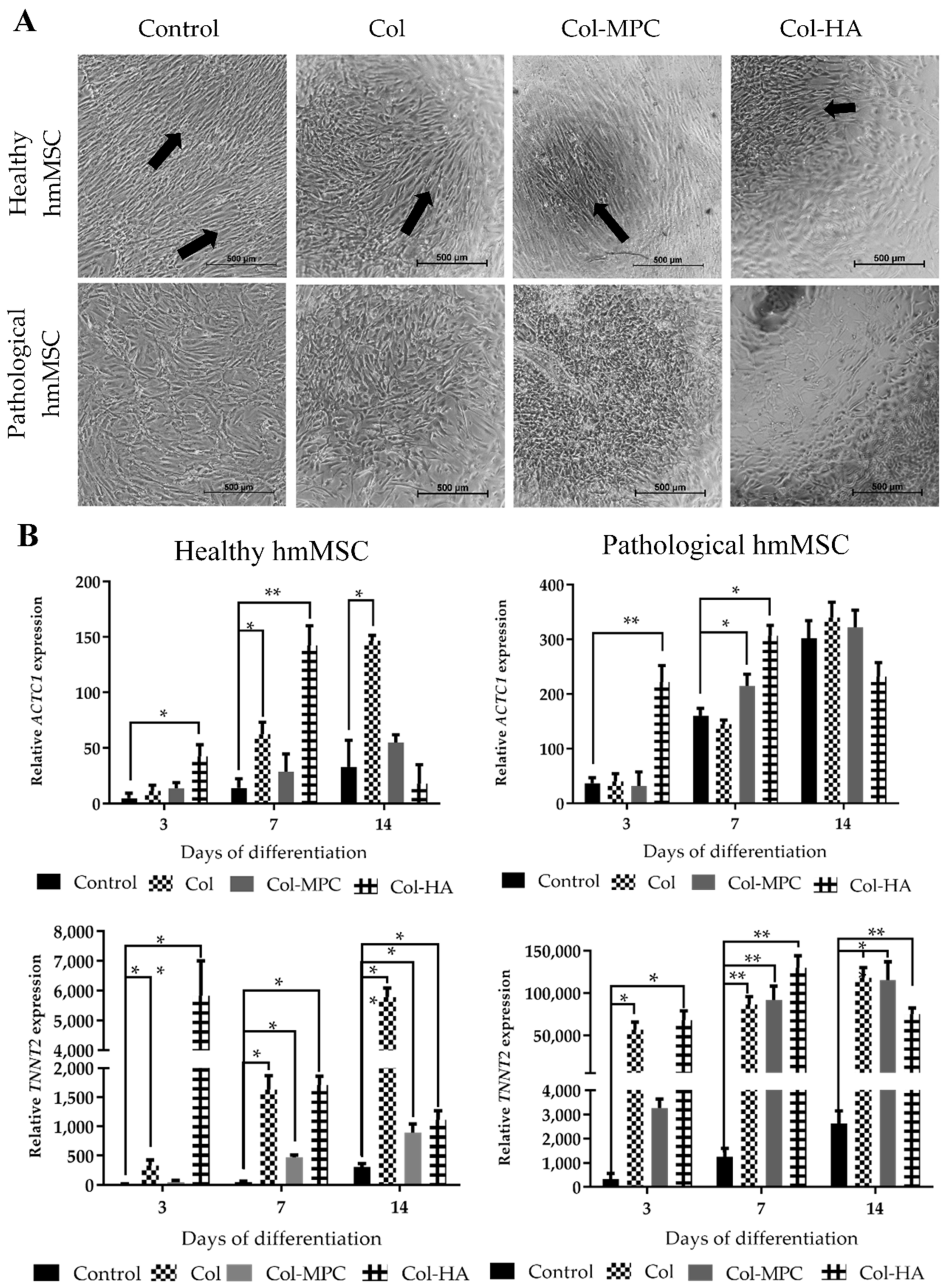
2.5. The Differentiation to Cardiomyogenic Direction of Healthy and Pathological hmMSC on Hybrid Collagen Type I Hydrogels
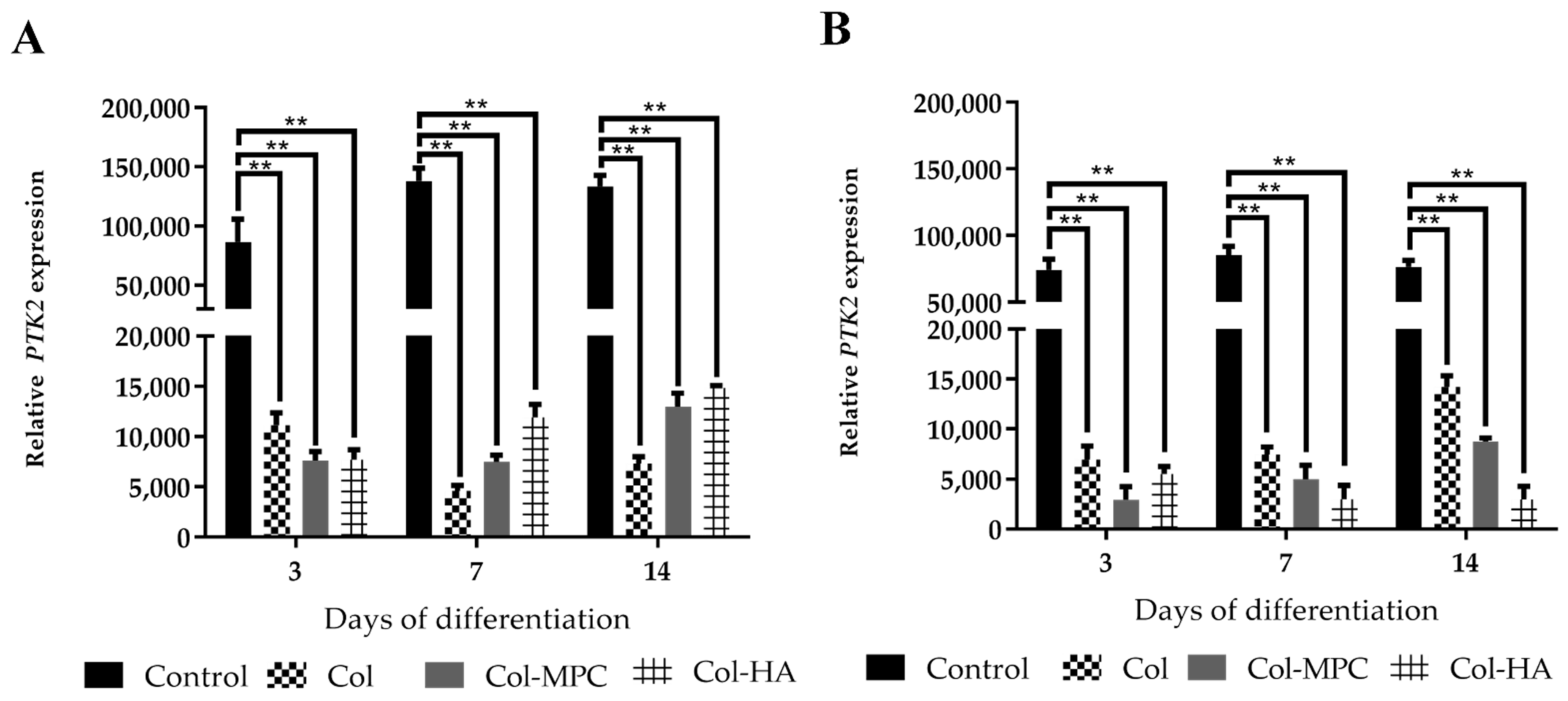
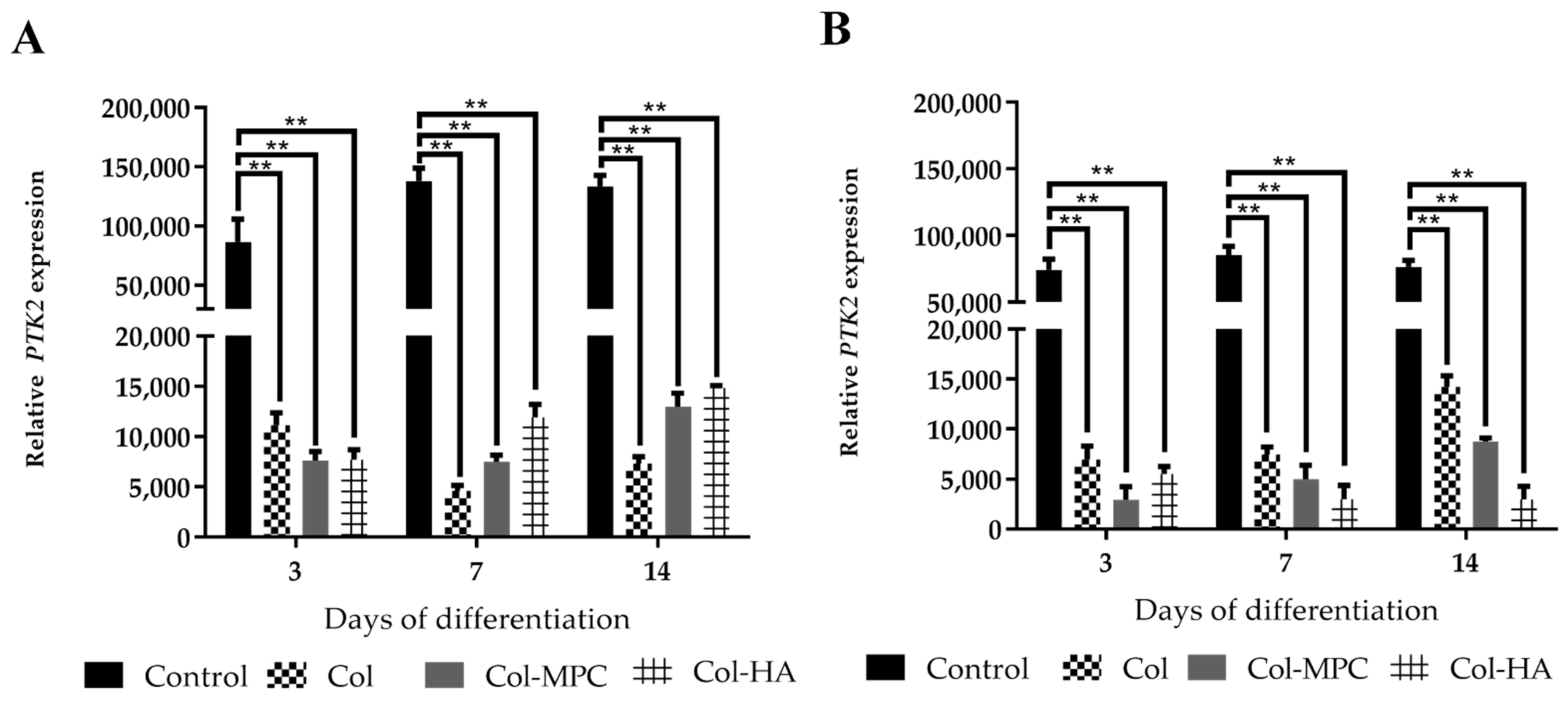
2.6. The Expression of Focal Adhesion Kinase (FAK) in Healthy and Pathological hmMSC Grown on Hybrid Collagen I Hydrogels
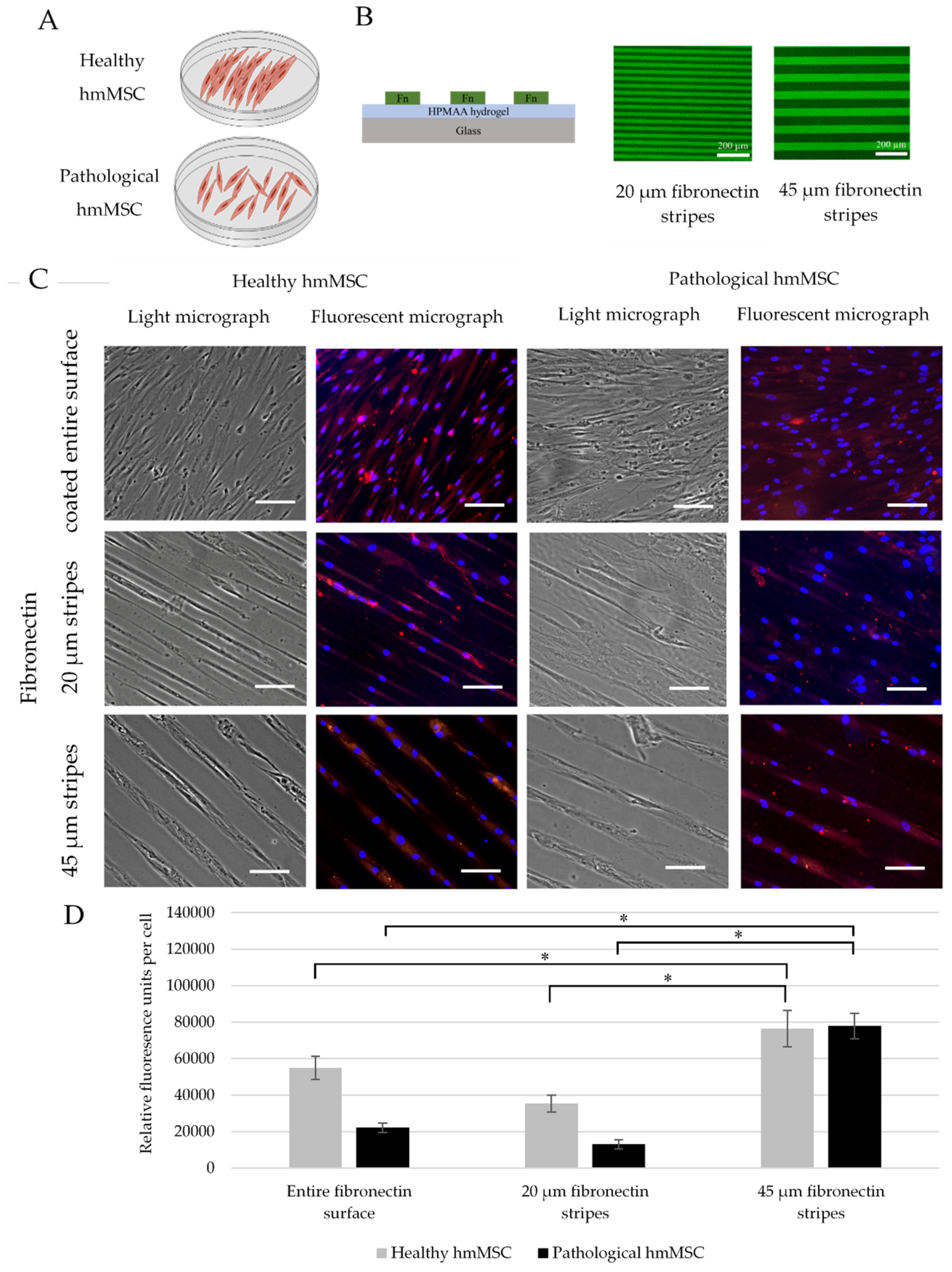
2.7. The Impact of Longitudinal hmMSC Culturing on the Cardiomyogenic Differentiation-Related Proteins
3. Discussion
4. Materials and Methods
4.1. Isolation and Cultivation of hmMSC
4.2. Evaluation of hmMSC Morphology
4.3. Identification of hmMSC Surface Markers
4.4. Differentiation of hmMSC to Adipo-, Osteo-, and Chondrogenic Directions
4.5. Proliferation of Healthy and DCM Myocardium-Derived hmMSC
4.6. Evaluation of Calcium Concentration with Flow Cytometry
4.7. Detection of HDAC Activity
4.8. Preparation of Hydrogels
4.9. Preparation of Fibronectine Lines on Glass Chips
4.10. Cultivation of hmMSC on Hydrogels and Biochips
4.11. Cardiomyogenic Differentiation of Healthy and Dilated Myocardium-Derived hmMSC
4.12. Gene Expression Levels
4.13. Immunofluorescence Assay
4.14. Statistical Analysis
4.15. Ethical Approval
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACTB | Actin beta |
| ACTC1 | Alpha cardiac actin 1 |
| CCK-8 | Cell counting kit 8 |
| APS | Ammonium persulfate |
| ECM | Extracellular matrix |
| DAPI | 40,6-diamidino-2-phenylindole |
| DCM | Dilated cardiomyopathy |
| DMTMM | 4-(4,6-dimethoxy-1,3,5-triazin-2-yl)-4-methylmorpholinium chloride |
| HA | Hyaluronic acid |
| HDAC | Histone deacetylases |
| hmMSC | Human myocardium-derived mesenchymal stem cell |
| MEF-2 | Myocyte enhancer factor-2 |
| MES | 2-(N-morpholino)ethanesulfonic acid |
| MPC | 2-methacryloyloxyethyl phosphorylcholine |
| MSC | Mesenchymal stem cells |
| PEGDA | poly(ethylene glycol) diacrylate |
| SAHA | Suberoylanilide hydroxamic acid |
| TEMED | N,N,N′,N′-tetramethylethane-1,2-diamine |
| TNNT2 | Cardiac Muscle Troponin T |
References
- Richardson, P.; McKenna, W.; Bristow, M.; Maisch, B.; Mautner, B.; O’Connell, J.; Olsen, E.; Thiene, G.; Goodwin, J.; Gyarfas, I.; et al. Report of the 1995 World Health Organization/International Society and Federation of Cardiology Task Force on the Definition and Classification of Cardiomyopathies. Circulation 1996, 93, 841–842. [Google Scholar] [CrossRef] [PubMed]
- Lakdawala, N.K.; Winterfield, J.R.; Funke, B.H. Dilated Cardiomyopathy. Circ. Arrhythmia Electrophysiol. 2013, 6, 228–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambrosy, A.P.; Fonarow, G.C.; Butler, J.; Chioncel, O.; Greene, S.J.; Vaduganathan, M.; Nodari, S.; Lam, C.S.P.; Sato, N.; Shah, A.N.; et al. The global health and economic burden of hospitalizations for heart failure: Les-sons learned from hospitalized heart failure registries. J. Am. Coll. Cardiol. 2014, 63, 1123–1133. [Google Scholar] [CrossRef]
- Cahill, T.J.; Choudhury, R.P.; Riley, P.R. Heart regeneration and repair after myocardial infarction: Transla-tional opportunities for novel therapeutics. Nat. Rev. Drug Discov. 2017, 16, 699–717. [Google Scholar] [CrossRef]
- Telukuntla, K.S.; Suncion, V.Y.; Schulman, I.H.; Hare, J.M. The Advancing Field of Cell-Based Therapy: Insights and Lessons from Clinical Trials. J. Am. Hear. Assoc. 2013, 2, e000338. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.-N.; Cores, J.; Huang, K.; Cui, X.; Luo, L.; Zhang, J.-Y.; Li, T.-S.; Qian, L.; Cheng, K. Concise Review: Is Cardiac Cell Therapy Dead? Embarrassing Trial Outcomes and New Directions for the Future. STEM CELLS Transl. Med. 2018, 7, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Segers, V.F.; Lee, R.T. Biomaterials to Enhance Stem Cell Function in the Heart. Circ. Res. 2011, 109, 910–922. [Google Scholar] [CrossRef]
- Heallen, T.R.; Martin, J.F. Heart repair via cardiomyocyte-secreted vesicles. Nat. Biomed. Eng. 2018, 2, 271–272. [Google Scholar] [CrossRef]
- Liu, B.; Lee, B.W.; Nakanishi, K.; Villasante, A.; Williamson, R.; Metz, J.; Kim, J.; Kanai, M.; Bi, L.; Brown, K.; et al. Cardiac recovery via extended cell-free delivery of extracellular vesicles secreted by cardiomyocytes derived from induced pluripotent stem cells. Nat. Biomed. Eng. 2018, 2, 293–303. [Google Scholar] [CrossRef]
- Taunton, J.; Hassig, C.A.; Schreiber, S.L. A Mammalian Histone Deacetylase Related to the Yeast Transcriptional Regulator Rpd3p. Science 1996, 272, 408–411. [Google Scholar] [CrossRef] [PubMed]
- Glozak, M.A.; Sengupta, N.; Zhang, X.; Seto, E. Acetylation and deacetylation of non-histone proteins. Gene 2005, 363, 15–23. [Google Scholar] [CrossRef]
- Yoon, S.; Eom, G.H. HDAC and HDAC Inhibitor: From Cancer to Cardiovascular Diseases. Chonnam Med. J. 2016, 52, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faller, Y.D.A.D.V. Transcription Regulation by Class III Histone Deacetylases (HDACs)—Sirtuins. Transl. Oncogenom. 2008, 1, 53–65. [Google Scholar] [CrossRef]
- Liu, S.-S.; Wu, F.; Jin, Y.-M.; Chang, W.Q.; Xu, T.-M. HDAC11: A rising star in epigenetics. Biomed. Pharmacother. 2020, 131, 110607. [Google Scholar] [CrossRef]
- Glauben, R.; Batra, A.; Stroh, T.; Erben, U.; Fedke, I.; Lehr, H.A.; Leoni, F.; Mascagni, P.; Dinarello, C.A.; Zeitz, M.; et al. Histone deacetylases: Novel targets for prevention of colitis-associated cancer in mice. Gut 2008, 57, 613–622. [Google Scholar] [CrossRef]
- Simões-Pires, C.; Zwick, V.; Nurisso, A.; Schenker, E.; Carrupt, P.A.; Cuendet, M. HDAC6 as a target for neu-rodegenerative diseases: What makes it different from the other HDACs? Mol. Neurodegener. 2013, 8, 7. [Google Scholar] [CrossRef] [Green Version]
- Adcock, I.M. HDAC inhibitors as anti-inflammatory agents. Br. J. Pharmacol. 2007, 150, 829–831. [Google Scholar] [CrossRef] [PubMed]
- Caliari, S.; Burdick, J.A. A practical guide to hydrogels for cell culture. Nat. Methods 2016, 13, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Izadpanah, R.; Kaushal, D.; Kriedt, C.; Tsien, F.; Patel, B.; Dufour, J.; Bunnell, B.A. Long-Term In Vitro Expansion Alters the Biology of Adult Mesenchymal Stem Cells. Cancer Res. 2008, 68, 4229–4238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valencik, M.L.; McDonald, J.A. Cardiac expression of a gain-of-function α5-integrin results in perinatal lethality. Am. J. Physiol. Circ. Physiol. 2001, 280, H361–H367. [Google Scholar] [CrossRef]
- Shai, S.-Y.; Harpf, A.E.; Babbitt, C.J.; Jordan, M.C.; Fishbein, M.C.; Chen, J.; Omura, M.; Leil, T.; Becker, K.D.; Jiang, M.; et al. Cardiac Myocyte-Specific Excision of the β1 Integrin Gene Results in Myocardial Fibrosis and Cardiac Failure. Circ. Res. 2002, 90, 458–464. [Google Scholar] [CrossRef] [Green Version]
- Valencik, M.L.; Keller, R.S.; Loftus, J.C.; McDonald, J.A. A lethal perinatal cardiac phenotype resulting from altered integrin function in cardiomyocytes. J. Card. Fail. 2002, 8, 262–272. [Google Scholar] [CrossRef]
- Parsons, J.T. Focal adhesion kinase: The first ten years. J. Cell Sci. 2003, 116, 1409–1416. [Google Scholar] [CrossRef] [Green Version]
- Dorn, G.W.; Force, T. Protein kinase cascades in the regulation of cardiac hypertrophy. J. Clin. Investig. 2005, 115, 527–537. [Google Scholar] [CrossRef] [Green Version]
- Torsoni, A.S.; Constancio, S.S.; NadruzJr, W.; Hanks, S.K.; Franchini, K.G. Focal Adhesion Kinase Is Activated and Mediates the Early Hypertrophic Response to Stretch in Cardiac Myocytes. Circ. Res. 2003, 93, 140–147. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.; Kraus, M.S.; Wei, H.; Shen, T.-L.; Pariaut, R.; Alcaraz, A.; Ji, G.; Cheng, L.; Yang, Q.; Kotlikoff, M.I.; et al. Inactivation of focal adhesion kinase in cardiomyocytes promotes eccentric cardiac hypertrophy and fibrosis in mice. J. Clin. Investig. 2005, 116, 217–227. [Google Scholar] [CrossRef] [Green Version]
- DiMichele, L.A.; Doherty, J.T.; Rojas, M.; Beggs, H.E.; Reichardt, L.F.; Mack, C.P.; Taylor, J.M. Myocyte-Restricted Focal Adhesion Kinase Deletion Attenuates Pressure Overload–Induced Hypertrophy. Circ. Res. 2006, 99, 636–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenger, M.P.; Bozec, L.; Horton, M.A.; Mesquida, P. Mechanical Properties of Collagen Fibrils. Biophys. J. 2007, 93, 1255–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazir, R.; Bruyneel, A.; Carr, C.; Czernuszka, J. Collagen type I and hyaluronic acid based hybrid scaffolds for heart valve tissue engineering. Biopolymers 2019, 110, e23278. [Google Scholar] [CrossRef]
- Simpson, F.C.; McTiernan, C.D.; Islam, M.M.; Buznyk, O.; Lewis, P.N.; Meek, K.M.; Haagdorens, M.; Audiger, C.; Lesage, S.; Gueriot, F.-X.; et al. Collagen analogs with phosphorylcholine are inflammation-suppressing scaffolds for corneal regeneration from alkali burns in mini-pigs. Commun. Biol. 2021, 4, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Skardal, A.; Mack, D.; Atala, A.; Soker, S. Substrate elasticity controls cell proliferation, surface marker expression and motile phenotype in amniotic fluid-derived stem cells. J. Mech. Behav. Biomed. Mater. 2013, 17, 307–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, P.; Galenano-Niño, J.L.; Graney, P.; Razal, J.; Minett, A.I.; Ribas, J.; Ovalle-Robles, R.; Biro, M.; Zreiqat, H. Relationship between nanotopographical alignment and stem cell fate with live imaging and shape analysis. Sci. Rep. 2016, 6, 37909. [Google Scholar] [CrossRef]
- Almonacid Suarez, A.M.; Zhou, Q.; Van Rijn, P.; Harmsen, M.C. Directional topography gradients drive optimum alignment and differentiation of human myoblasts. J. Tissue Eng. Regen. Med. 2019, 13, 2234–2245. [Google Scholar] [CrossRef] [Green Version]
- Mitcheson, J.S.; Hancox, J.C.; Levi, A.J. Cultured adult cardiac myocytes: Future applications, culture meth-ods, morphological and electrophysiological properties. Cardiovasc. Res. 1998, 39, 280–300. [Google Scholar] [CrossRef]
- Bergmann, O.; Bhardwaj, R.D.; Bernard, S.; Zdunek, S.; Barnabé-Heider, F.; Walsh, S.; Zupicich, J.; Alkass, K.; Buchholz, B.A.; Druid, H.; et al. Evidence for Cardiomyocyte Renewal in Humans. Science 2009, 324, 98–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, O.; Zdunek, S.; Felker, A.; Salehpour, M.; Alkass, K.; Bernard, S.; Sjostrom, S.L.; Szewczykowska, M.; Jackowska, T.; Dos Remedios, C.; et al. Dynamics of Cell Generation and Turnover in the Human Heart. Cell 2015, 161, 1566–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beltrami, A.P.; Urbanek, K.; Kajstura, J.; Yan, S.; Finato, N.; Bussani, R.; Nadal-Ginard, B.; Silvestri, F.; Leri, A.; Beltrami, C.A.; et al. Evidence That Human Cardiac Myocytes Divide after Myocardial Infarction. N. Engl. J. Med. 2001, 344, 1750–1757. [Google Scholar] [CrossRef]
- Senyo, S.; Steinhauser, M.L.; Pizzimenti, C.L.; Yang, V.K.; Cai, L.; Wang, M.; Wu, T.-D.; Guerquin-Kern, J.-L.; Lechene, C.P.; Lee, R.T. Mammalian heart renewal by pre-existing cardiomyocytes. Nature 2012, 493, 433–436. [Google Scholar] [CrossRef] [Green Version]
- Davis, D.R.; Smith, R.R.; Marbán, E. Human Cardiospheres are a Source of Stem Cells with Cardiomyogenic Potential. Stem Cells 2010, 28, 903–904. [Google Scholar] [CrossRef] [Green Version]
- Bearzi, C.; Rota, M.; Hosoda, T.; Tillmanns, J.; Nascimbene, A.; De Angelis, A.; Yasuzawa-Amano, S.; Trofimova, I.; Siggins, R.W.; LeCapitaine, N.; et al. Human cardiac stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 14068–14073. [Google Scholar] [CrossRef] [Green Version]
- Kulvinskiene, I.; Aldonyte, R.; Miksiunas, R.; Mobasheri, A.; Bironaite, D. Biomatrices for Heart Regeneration and Cardiac Tissue Modelling In Vitro. In Cell Biology and Translational Medicine; Springer: Cham, Switzerland, 2020; Volume 10, pp. 43–77. [Google Scholar] [CrossRef]
- Orbay, H.; Tobita, M.; Mizuno, H. Mesenchymal Stem Cells Isolated from Adipose and Other Tissues: Basic Biological Properties and Clinical Applications. Stem Cells Int. 2012, 2012, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, I.; Kwon, B.-S.; Park, H.-K.; Kim, K.S. Differentiation Potential of Mesenchymal Stem Cells Is Related to Their Intrinsic Mechanical Properties. Int. Neurourol. J. 2017, 21, S24–S31. [Google Scholar] [CrossRef] [PubMed]
- Garikipati, V.N.S.; Singh, S.P.; Mohanram, Y.; Gupta, A.K.; Kapoor, D.; Nityanand, S. Isolation and characterization of mesenchymal stem cells from human fetus heart. PLoS ONE 2018, 13, e0192244. [Google Scholar] [CrossRef] [Green Version]
- Zabirnyk, A.; Perez, M.D.M.; Blasco, M.; Stensløkken, K.-O.; Ferrer, M.D.; Salcedo, C.; Vaage, J. A Novel Ex Vivo Model of Aortic Valve Calcification. A Preliminary Report. Front. Pharmacol. 2020, 11, 568764. [Google Scholar] [CrossRef]
- Miksiunas, R.; Rucinskas, K.; Janusauskas, V.; Labeit, S.; Bironaite, D. Histone Deacetylase Inhibitor Suberoylanilide Hydroxamic Acid Improves Energetic Status and Cardiomyogenic Differentiation of Human Dilated Myocardium-Derived Primary Mesenchymal Cells. Int. J. Mol. Sci. 2020, 21, 4845. [Google Scholar] [CrossRef]
- Rossini, A.; Frati, C.; Lagrasta, C.; Graiani, G.; Scopece, A.; Cavalli, S.; Musso, E.; Baccarin, M.; Di Segni, M.; Fagnoni, F.; et al. Human cardiac and bone marrow stromal cells exhibit distinctive properties related to their origin. Cardiovasc. Res. 2010, 89, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Horrobin, D.F. Modern biomedical research: An internally self-consistent universe with little contact with medical reality? Nat. Rev. Drug Discov. 2003, 2, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Kim, H.J.; Bae, S.C. Histone deacetylase inhibitors: Molecular mechanisms of action and clinical trials as anti-cancer drugs. Am. J. Transl. Res. 2011, 3, 166–179. [Google Scholar] [PubMed]
- McKinsey, T.A. Isoform-selective HDAC inhibitors: Closing in on translational medicine for the heart. J. Mol. Cell. Cardiol. 2011, 51, 491–496. [Google Scholar] [CrossRef]
- Lu, J.; McKinsey, T.A.; Nicol, R.L.; Olson, E.N. Signal-dependent activation of the MEF2 transcription factor by dissociation from histone deacetylases. Proc. Natl. Acad. Sci. USA 2000, 97, 4070–4075. [Google Scholar] [CrossRef] [Green Version]
- Eom, G.H.; Kook, H. Role of histone deacetylase 2 and its posttranslational modifications in cardiac hyper-trophy. BMB Rep. 2015, 48, 131–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteforte, N.; Napolitano, C.; Priori, S.G. Genética y arritmias: Aplicaciones diagnósticas y pronósticas. Rev. Esp. Cardiol. 2012, 65, 278–286. [Google Scholar] [CrossRef]
- Bush, E.W.; McKinsey, T.A. Targeting histone deacetylases for heart failure. Expert Opin. Ther. Targets 2009, 13, 767–784. [Google Scholar] [CrossRef]
- da Silva Meirelles, L.; Chagastelles, P.C.; Nardi, N.B. Mesenchymal stem cells reside in virtually all post-natal organs and tissues. J. Cell Sci. 2006, 119, 2204–2213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.D.; Weiss, R.M.; Heistad, D.D. Calcific Aortic Valve Stenosis: Methods, Models, and Mechanisms. Circ. Res. 2011, 108, 1392–1412. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Yu, D.; Wu, H.; Liu, H.; Zhou, H.; Gu, R.; Zhang, R.; Zhang, S.; Wu, G. Anticancer activity of SAHA, a potent histone deacetylase inhibitor, in NCI-H460 human large-cell lung carcinoma cells in vitro and in vivo. Int. J. Oncol. 2013, 44, 451–458. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, R.L.; Davis, C.A.; Potthoff, M.J.; Haberland, M.; Fielitz, J.; Qi, X.; Hill, J.A.; Richardson, J.A.; Olson, E.N. Histone deacetylases 1 and 2 redundantly regulate cardiac morphogenesis, growth, and contractility. Genes Dev. 2007, 21, 1790–1802. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wang, H.; Zhao, Y.; Wang, J.; Dubielecka, P.M.; Zhuang, S.; Qin, G.; Chin, Y.E.; Kao, R.L.; Zhao, T.C. Myocyte-specific overexpressing HDAC4 promotes myocardial ischemia/reperfusion injury. Mol. Med. 2018, 24, 1–10. [Google Scholar] [CrossRef]
- Leng, Y.; Wu, Y.; Lei, S.; Zhou, B.; Qiu, Z.; Wang, K.; Xia, Z. Inhibition of HDAC6 Activity Alleviates Myocardial Ischemia/Reperfusion Injury in Diabetic Rats: Potential Role of Peroxiredoxin 1 Acetylation and Redox Regulation. Oxid. Med. Cell. Longev. 2018, 2018, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nural-Guvener, H.F.; Zakharova, L.; Nimlos, J.; Popovic, S.; Mastroeni, D.; Gaballa, M.A. HDAC class I inhibitor, Mocetinostat, reverses cardiac fibrosis in heart failure and diminishes CD90+ cardiac myofibroblast activation. Fibrogenes. Tissue Repair 2014, 7, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyer, A.; Fenning, A.; Lim, J.; Le, G.T.; Reid, R.C.; Halili, M.A.; Fairlie, D.P.; Brown, L. Antifibrotic activity of an inhibitor of histone deacetylases in DOCA-salt hypertensive rats. Br. J. Pharmacol. 2010, 159, 1408–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadler, K.E.; Baldock, C.; Bella, J.; Boot-Handford, R. Collagens at a glance. J. Cell Sci. 2007, 120, 1955–1958. [Google Scholar] [CrossRef] [Green Version]
- Horn, M.; Trafford, A.W. Aging and the cardiac collagen matrix: Novel mediators of fibrotic remodelling. J. Mol. Cell. Cardiol. 2015, 93, 175–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Souza, R. Aging of myocardial collagen. Biogerontology 2002, 3, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Janicki, J.S. Myocardial collagen remodeling and left ventricular diastolic function. Braz. J. Med Biol. Res. 1992, 25, 975–982. [Google Scholar]
- McLaughlin, S.; McNeill, B.; Podrebarac, J.; Hosoyama, K.; Sedlakova, V.; Cron, G.; Smyth, D.; Seymour, R.; Goel, K.; Liang, W.; et al. Injectable human recombinant collagen matrices limit adverse remodeling and improve cardiac function after myocardial infarction. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bai, R.; Tian, L.; Li, Y.; Zhang, J.; Wei, Y.; Jin, Z.; Liu, Z.; Liu, H. Combining ECM Hydrogels of Cardiac Bioactivity with Stem Cells of High Cardiomyogenic Potential for Myocardial Repair. Stem Cells Int. 2019, 2019, 6708435. [Google Scholar] [CrossRef]
- Lane, B.A.; Harmon, K.A.; Goodwin, R.L.; Yost, M.J.; Shazly, T.; Eberth, J.F. Constitutive modeling of compressible type-I collagen hydrogels. Med Eng. Phys. 2018, 53, 39–48. [Google Scholar] [CrossRef]
- Bonafè, F.; Govoni, M.; Giordano, E.; Caldarera, C.M.; Guarnieri, C.; Muscari, C. Hyaluronan and cardiac regeneration. J. Biomed. Sci. 2014, 21, 1–13. [Google Scholar] [CrossRef] [Green Version]
- John, H.E.; Price, R.D. Perspectives in the selection of hyaluronic acid fillers for facial wrinkles and aging skin. Patient Prefer. Adherence 2009, 3, 225–230. [Google Scholar]
- Iwasaki, Y.; Ishihara, K. Cell membrane-inspired phospholipid polymers for developing medical devices with excellent biointerfaces. Sci. Technol. Adv. Mater. 2012, 13, 064101. [Google Scholar] [CrossRef]
- Koike, M.; Kurosawa, H.; Amano, Y. A Round-bottom 96-well Polystyrene Plate Coated with 2-methacryloyloxyethyl Phosphorylcholine as an Effective Tool for Embryoid Body Formation. Cytotechnology 2005, 47, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.M.; Cėpla, V.; He, C.; Edin, J.; Rakickas, T.; Kobuch, K.; Ruželė, Ž.; Jackson, W.B.; Rafat, M.; Lohmann, C.P.; et al. Functional fabrication of recombinant human collagen–phosphorylcholine hydrogels for regenerative medicine applications. Acta Biomater. 2015, 12, 70–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedron, S.; van Lierop, S.; Horstman, P.; Penterman, R.; Broer, D.J.; Peeters, E. Stimuli Responsive Delivery Vehicles for Cardiac Microtissue Transplantation. Adv. Funct. Mater. 2011, 21, 1624–1630. [Google Scholar] [CrossRef]
- Birla, R.K.; Borschel, G.H.; Dennis, R.G.; Brown, D.L. Myocardial Engineeringin Vivo: Formation and Characterization of Contractile, Vascularized Three-Dimensional Cardiac Tissue. Tissue Eng. 2005, 11, 803–813. [Google Scholar] [CrossRef] [Green Version]
- Zustiak, S.P.; Wei, Y.; Leach, J.B. Protein-hydrogel interactions in tissue engineering: Mechanisms and applications. Tissue Eng. Part B Rev. 2013, 19, 160–171. [Google Scholar] [CrossRef] [Green Version]
- DeForest, C.A.; Anseth, K.S. Advances in Bioactive Hydrogels to Probe and Direct Cell Fate. Annu. Rev. Chem. Biomol. Eng. 2012, 3, 421–444. [Google Scholar] [CrossRef] [Green Version]
- Mitra, S.K.; Hanson, D.A.; Schlaepfer, D.D. Focal adhesion kinase: In command and control of cell motility. Nat. Rev. Mol. Cell Biol. 2005, 6, 56–68. [Google Scholar] [CrossRef]
- Lopes, M.M.; Ribeiro, G.C.A.; Tornatore, T.F.; Clemente, C.F.M.Z.; Teixeira, V.P.A.; Franchini, K.G. Increased expression and phosphorylation of focal adhesion kinase correlates with dysfunction in the volume-overloaded human heart. Clin. Sci. 2007, 113, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Clemente, C.F.; Tornatore, T.F.; Theizen, T.H.; Deckmann, A.C.; Pereira, T.C.; Lopes-Cendes, I.; Souza, J.R.M.; Franchini, K.G. Targeting Focal Adhesion Kinase with Small Interfering RNA Prevents and Reverses Load-Induced Cardiac Hypertrophy in Mice. Circ. Res. 2007, 101, 1339–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, G.-P.; Wang, W.; Zhao, H.; Cai, L.; Zhang, P.-D.; Yang, Z.-H.; Zhang, J.; Wang, X. Pharmacological Inhibition of Focal Adhesion Kinase Attenuates Cardiac Fibrosis in Mice Cardiac Fibroblast and Post-Myocardial-Infarction Models. Cell. Physiol. Biochem. 2015, 37, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Geckil, H.; Xu, F.; Zhang, X.; Moon, S.; Demirci, U. Engineering hydrogels as extracellular matrix mimics. Nanomedicine 2010, 5, 469–484. [Google Scholar] [CrossRef] [Green Version]
- Altomare, L.; Riehle, M.; Gadegaard, N.; Tanzi, M.C.; Farè, S. Microcontact Printing of Fibronectin on a Biodegradable Polymeric Surface for Skeletal Muscle Cell Orientation. Int. J. Artif. Organs 2010, 33, 535–543. [Google Scholar] [CrossRef]
- Bajaj, P.; Reddy, B.; Millet, L.; Wei, C.; Zorlutuna, P.; Bao, G.; Bashir, R. Patterning the differentiation of C2C12 skeletal myoblasts. Integr. Biol. 2011, 3, 897–909. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Andersson, H.; Martinsson, S.; Sabirsh, A.; Jonebring, A.; Wang, Q.-D.; Plowright, A.T.; Drowley, L. Aligned nanofiber scaffolds improve functionality of cardiomyocytes differentiated from human induced pluripotent stem cell-derived cardiac progenitor cells. Sci. Rep. 2020, 10, 13575. [Google Scholar] [CrossRef]
- Erratum to: Mesenchymal Progenitors Able to Differentiate into Osteogenic, Chondrogenic, and/or Adipogenic Cells In Vitro Are Present in Most Primary Fibroblast-Like Cell Populations. Stem Cells 2014, 32, 1995–1996. [CrossRef]
- Liu, Y.; Gan, L.; Carlsson, D.J.; Fagerholm, P.; Lagali, N.; Watsky, M.; Munger, R.; Hodge, W.G.; Priest, D.; Griffith, M. A Simple, Cross-linked Collagen Tissue Substitute for Corneal Implantation. Investig. Opthalmol. Vis. Sci. 2006, 47, 1869–1875. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Deng, C.; McLaughlin, C.R.; Fagerholm, P.; Lagali, N.S.; Heyne, B.; Scaiano, J.C.; Watsky, M.; Kato, Y.; Munger, R.; et al. Collagen–phosphorylcholine interpenetrating network hydrogels as corneal substitutes. Biomaterials 2009, 30, 1551–1559. [Google Scholar] [CrossRef]
- Cėpla, V.; Rakickas, T.; Stankevičienė, G.; Mazėtytė-Godienė, A.; Baradoke, A.; Ruželė, Ž.; Valiokas, R. Photografting and Patterning of Poly(ethylene glycol) Methacrylate Hydrogel on Glass for Biochip Applications. ACS Appl. Mater. Interfaces 2020, 12, 32233–32246. [Google Scholar] [CrossRef]
- Bironaite, D.; Brunk, U.; Venalis, A. Protective induction of Hsp70 in heat-stressed primary myoblasts: Involvement of MAPKs. J. Cell. Biochem. 2013, 114, 2024–2031. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miksiunas, R.; Aldonyte, R.; Vailionyte, A.; Jelinskas, T.; Eimont, R.; Stankeviciene, G.; Cepla, V.; Valiokas, R.; Rucinskas, K.; Janusauskas, V.; et al. Cardiomyogenic Differentiation Potential of Human Dilated Myocardium-Derived Mesenchymal Stem/Stromal Cells: The Impact of HDAC Inhibitor SAHA and Biomimetic Matrices. Int. J. Mol. Sci. 2021, 22, 12702. https://doi.org/10.3390/ijms222312702
Miksiunas R, Aldonyte R, Vailionyte A, Jelinskas T, Eimont R, Stankeviciene G, Cepla V, Valiokas R, Rucinskas K, Janusauskas V, et al. Cardiomyogenic Differentiation Potential of Human Dilated Myocardium-Derived Mesenchymal Stem/Stromal Cells: The Impact of HDAC Inhibitor SAHA and Biomimetic Matrices. International Journal of Molecular Sciences. 2021; 22(23):12702. https://doi.org/10.3390/ijms222312702
Chicago/Turabian StyleMiksiunas, Rokas, Ruta Aldonyte, Agne Vailionyte, Tadas Jelinskas, Romuald Eimont, Gintare Stankeviciene, Vytautas Cepla, Ramunas Valiokas, Kestutis Rucinskas, Vilius Janusauskas, and et al. 2021. "Cardiomyogenic Differentiation Potential of Human Dilated Myocardium-Derived Mesenchymal Stem/Stromal Cells: The Impact of HDAC Inhibitor SAHA and Biomimetic Matrices" International Journal of Molecular Sciences 22, no. 23: 12702. https://doi.org/10.3390/ijms222312702
APA StyleMiksiunas, R., Aldonyte, R., Vailionyte, A., Jelinskas, T., Eimont, R., Stankeviciene, G., Cepla, V., Valiokas, R., Rucinskas, K., Janusauskas, V., Labeit, S., & Bironaite, D. (2021). Cardiomyogenic Differentiation Potential of Human Dilated Myocardium-Derived Mesenchymal Stem/Stromal Cells: The Impact of HDAC Inhibitor SAHA and Biomimetic Matrices. International Journal of Molecular Sciences, 22(23), 12702. https://doi.org/10.3390/ijms222312702