
Transcriptomic Signatures of End-Stage Human Dilated Cardiomyopathy Hearts with and without Left Ventricular Assist Device Support

 ,
,
Abstract
1. Introduction
2. Results
2.1. Patient Clinical Characteristics
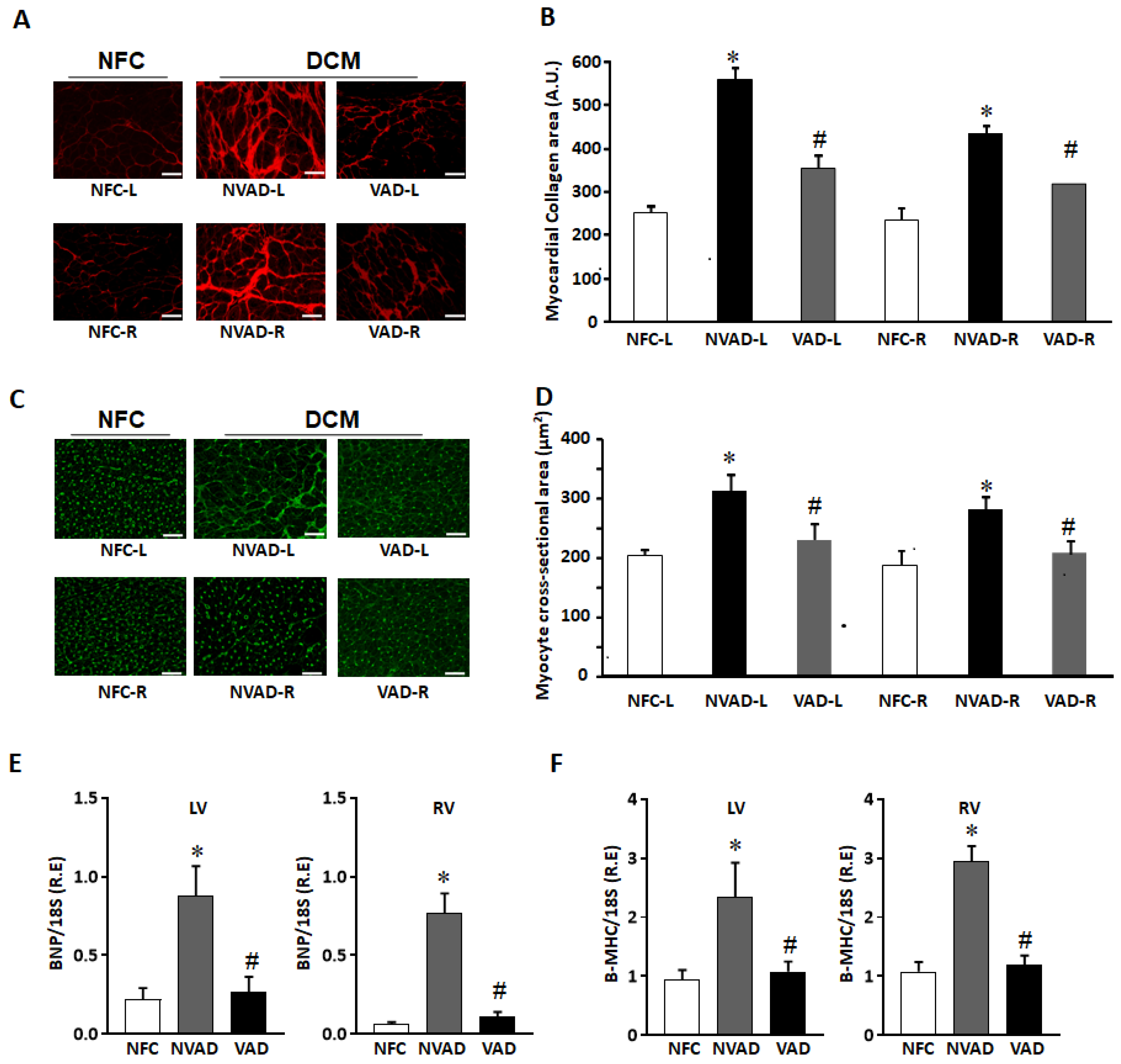
2.2. Histological Characteristics of Explanted DCM Hearts with and without LVAD
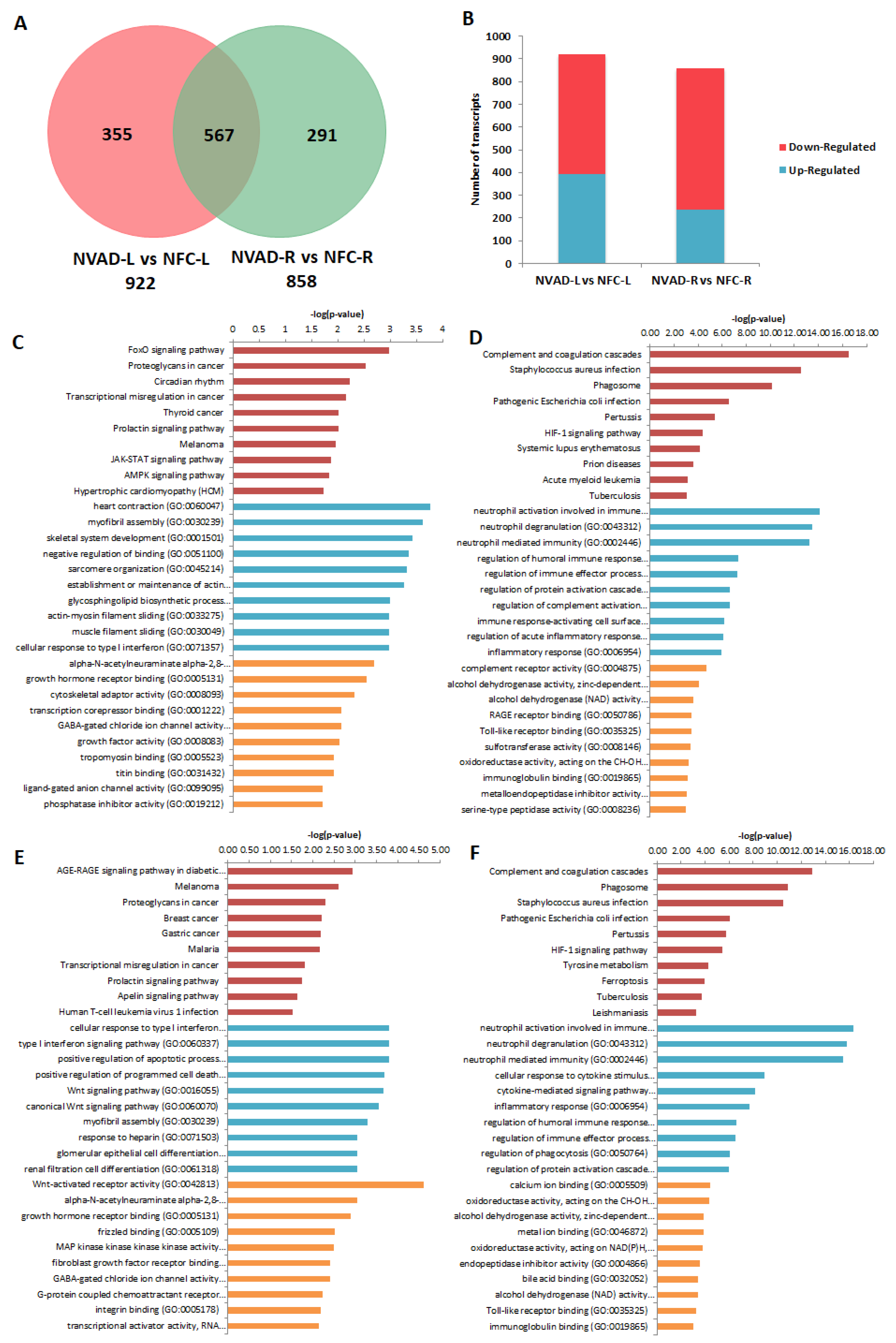
2.3. Pathological Genes of DCM Hearts
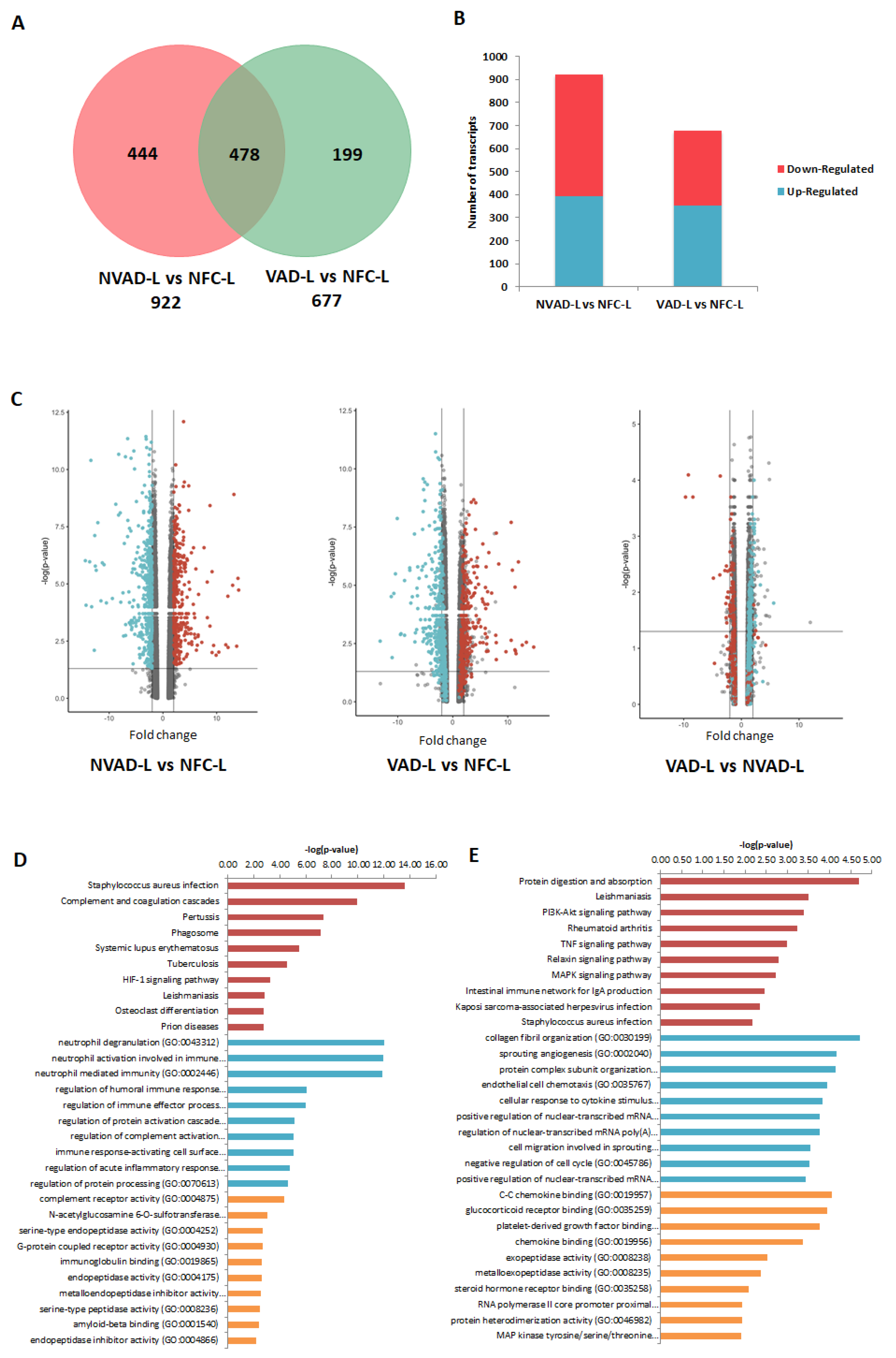
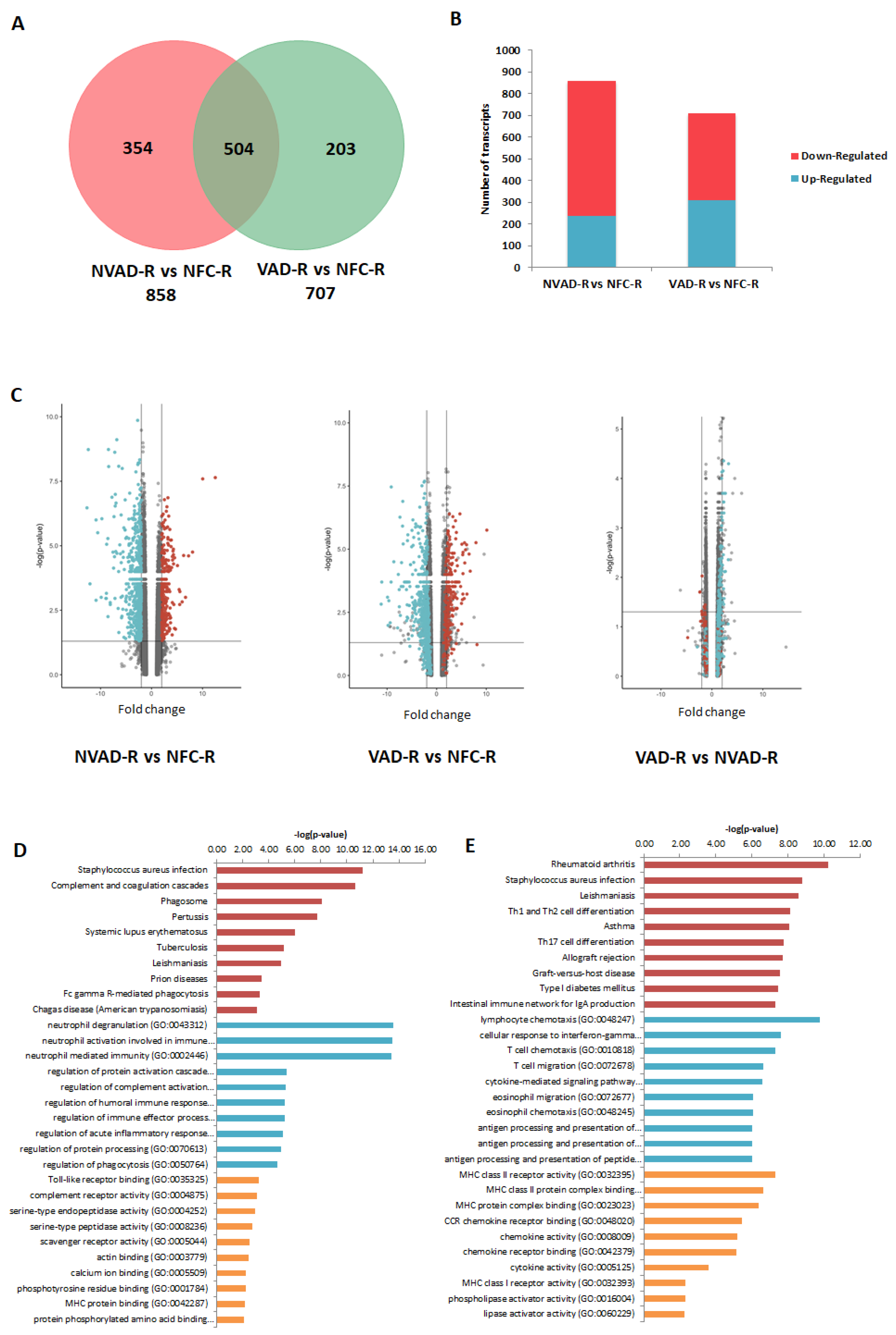
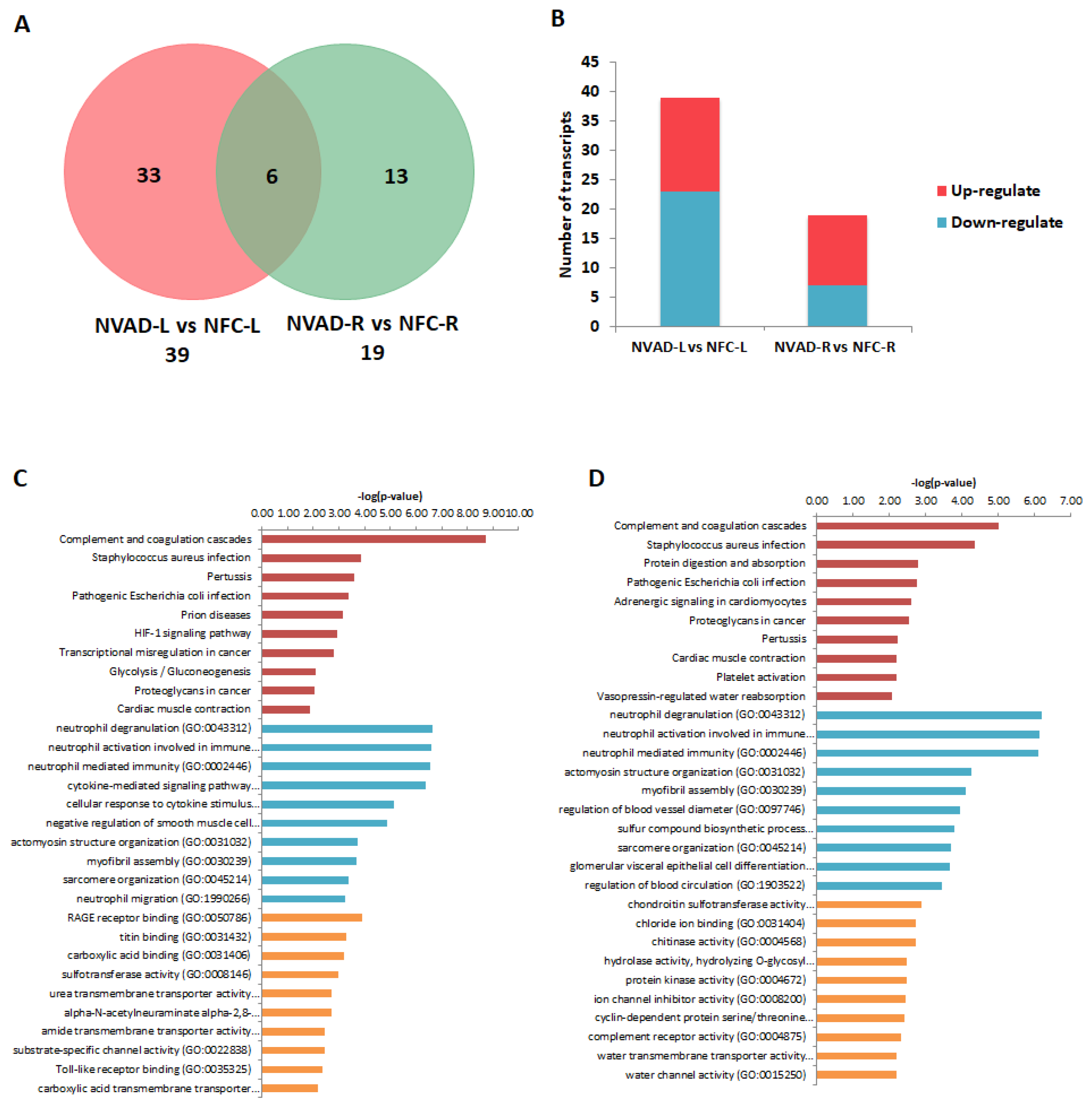
2.4. Gene Expression Changes in the LV and RV Post LVAD Implantation
2.5. Pathological miRNAs of DCM Hearts
2.6. miRNA Expression Changes in the LV and RV Post LVAD Implantation
3. Discussion
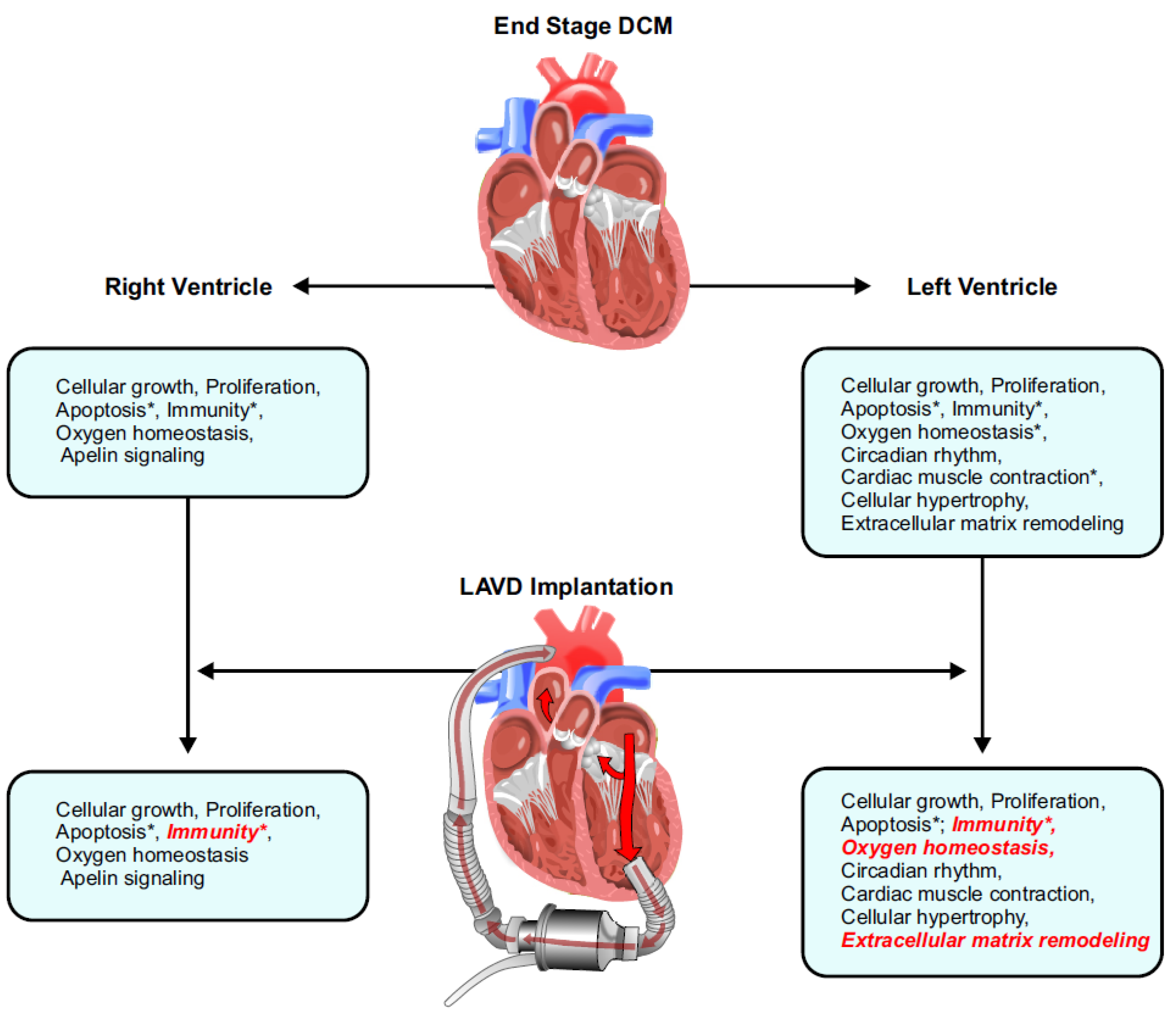
3.1. Pathological Genes Common in Both the LV and RV
3.2. LV- and RV-Specific Pathological Genes
3.3. Gene Expression Post-LVAD Use
3.4. Gene Regulation Post-LVAD Use
3.5. Strengths and Limitations
4. Materials and Methods
4.1. Study Patients and Protocol
4.2. Heart Tissue Procurement
4.3. Histology
4.3.1. Picrosirius Red Staining
4.3.2. Wheat Germ Agglutinin Staining
4.4. RNA Processing and Microarray
4.5. Data Generation and Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yogasundaram, H.; Alhumaid, W.; Dzwiniel, T.; Christian, S.; Oudit, G.Y. Cardiomyopathies and genetic testing in heart failure: Role in defining phenotype-targeted approaches and management. Can. J. Cardiol. 2021, 37, 547–559. [Google Scholar] [CrossRef]
- Dec, G.W.; Fuster, V. Idiopathic dilated cardiomyopathy. N. Engl. J. Med. 1994, 331, 1564–1575. [Google Scholar] [CrossRef]
- Caplan, A.L. Finding a solution to the organ shortage. Can. Med. Assoc. J. 2016, 188, 1182–1183. [Google Scholar] [CrossRef]
- Zhang, H.; Viveiros, A.; Nikhanj, A.; Nguyen, Q.; Wang, K.; Wang, W.; Freed, D.H.; Mullen, J.C.; MacArthur, R.; Kim, D.H.; et al. The human explanted heart program: A translational bridge for cardiovascular medicine. Biochim. Biophys. Acta -Mol. Basis Dis. 2021, 1867, 165995–166009. [Google Scholar] [CrossRef] [PubMed]
- Jacob, K.A.; Buijsrogge, M.P.; Ramjankhan, F.Z. Left ventricular assist devices for advanced heart failure. N. Engl. J. Med. 2017, 376, 1893–1894. [Google Scholar] [CrossRef] [PubMed]
- Heerdt, P.M.; Holmes, J.W.; Cai, B.; Barbone, A.; Madigan, J.D.; Reiken, S.; Lee, D.L.; Oz, M.C.; Marks, A.R.; Burkhoff, D. Chronic unloading by left ventricular assist device reverses contractile dysfunction and alters gene expression in end-stage heart failure. Circulation 2000, 102, 2713–2719. [Google Scholar] [CrossRef] [PubMed]
- Burkhoff, D.; Topkara, V.K.; Sayer, G.; Uriel, N. Reverse remodeling with left ventricular assist devices. Circ. Res. 2021, 128, 1594–1612. [Google Scholar] [CrossRef]
- Rose, E.A.; Gelijns, A.C.; Moskowitz, A.J.; Heitjan, D.F.; Stevenson, L.W.; Dembitsky, W.; Long, J.W.; Ascheim, D.D.; Tierney, A.R.; Levitan, R.G.; et al. Long-term use of a left ventricular assist device for end-stage heart failure. N. Engl. J. Med. 2001, 345, 1435–1443. [Google Scholar] [CrossRef]
- Rogers, J.G.; Butler, J.; Lansman, S.L.; Gass, A.; Portner, P.M.; Pasque, M.K.; Pierson, R.N., 3rd; Investigators, I.N. Chronic mechanical circulatory support for inotrope-dependent heart failure patients who are not transplant candidates: Results of the intrepid trial. J. Am. Coll. Cardiol. 2007, 50, 741–747. [Google Scholar] [CrossRef]
- Owens, A.T.; Jessup, M. Should left ventricular assist device be standard of care for patients with refractory heart failure who are not transplantation candidates? Left ventricular assist devices should not be standard of care for transplantation-ineligible patients. Circulation 2012, 126, 3088–3094. [Google Scholar] [CrossRef]
- Slaughter, M.S.; Rogers, J.G.; Milano, C.A.; Russell, S.D.; Conte, J.V.; Feldman, D.; Sun, B.; Tatooles, A.J.; Delgado, R.M., 3rd; Long, J.W.; et al. Advanced heart failure treated with continuous-flow left ventricular assist device. N. Engl. J. Med. 2009, 361, 2241–2251. [Google Scholar] [CrossRef] [PubMed]
- Kirklin, J.K.; Naftel, D.C.; Pagani, F.D.; Kormos, R.L.; Stevenson, L.W.; Blume, E.D.; Miller, M.A.; Timothy Baldwin, J.; Young, J.B. Sixth intermacs annual report: A 10,000-patient database. J. Heart Lung Transplant. 2014, 33, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Friedberg, M.K.; Redington, A.N. Right versus left ventricular failure: Differences, similarities, and interactions. Circulation 2014, 129, 1033–1044. [Google Scholar] [CrossRef] [PubMed]
- Frankfurter, C.; Molinero, M.; Vishram-Nielsen, J.K.K.; Foroutan, F.; Mak, S.; Rao, V.; Billia, F.; Orchanian-Cheff, A.; Alba, A.C. Predicting the risk of right ventricular failure in patients undergoing left ventricular assist device implantation: A systematic review. Circ. Heart Fail. 2020, 13, e006994. [Google Scholar] [CrossRef] [PubMed]
- Colak, D.; Alaiya, A.A.; Kaya, N.; Muiya, N.P.; AlHarazi, O.; Shinwari, Z.; Andres, E.; Dzimiri, N. Integrated left ventricular global transcriptome and proteome profiling in human end-stage dilated cardiomyopathy. PLoS ONE 2016, 11, e0162669. [Google Scholar] [CrossRef] [PubMed]
- Narula, J.; Haider, N.; Virmani, R.; DiSalvo, T.G.; Kolodgie, F.D.; Hajjar, R.J.; Schmidt, U.; Semigran, M.J.; Dec, G.W.; Khaw, B.A. Apoptosis in myocytes in end-stage heart failure. N. Engl. J. Med. 1996, 335, 1182–1189. [Google Scholar] [CrossRef] [PubMed]
- Zorc, M.; Vraspir-Porenta, O.; Zorc-Pleskovic, R.; Radovanovic, N.; Petrovic, D. Apoptosis of myocytes and proliferation markers as prognostic factors in end-stage dilated cardiomyopathy. Cardiovasc. Pathol. 2003, 12, 36–39. [Google Scholar] [CrossRef]
- Frantz, S.; Falcao-Pires, I.; Balligand, J.L.; Bauersachs, J.; Brutsaert, D.; Ciccarelli, M.; Dawson, D.; de Windt, L.J.; Giacca, M.; Hamdani, N.; et al. The innate immune system in chronic cardiomyopathy: A european society of cardiology (esc) scientific statement from the working group on myocardial function of the esc. Eur. J. Heart Fail. 2018, 20, 445–459. [Google Scholar] [CrossRef]
- Adamo, L.; Rocha-Resende, C.; Prabhu, S.D.; Mann, D.L. Reappraising the role of inflammation in heart failure. Nat. Rev. Cardiol. 2020, 17, 269–285. [Google Scholar] [CrossRef]
- Barth, A.S.; Kuner, R.; Buness, A.; Ruschhaupt, M.; Merk, S.; Zwermann, L.; Kaab, S.; Kreuzer, E.; Steinbeck, G.; Mansmann, U.; et al. Identification of a common gene expression signature in dilated cardiomyopathy across independent microarray studies. J. Am. Coll. Cardiol. 2006, 48, 1610–1617. [Google Scholar] [CrossRef]
- Semenza, G.L. Hypoxia-inducible factor 1 and cardiovascular disease. Annu. Rev. Physiol. 2014, 76, 39–56. [Google Scholar] [CrossRef] [PubMed]
- Kido, M.; Du, L.; Sullivan, C.C.; Li, X.; Deutsch, R.; Jamieson, S.W.; Thistlethwaite, P.A. Hypoxia-inducible factor 1-alpha reduces infarction and attenuates progression of cardiac dysfunction after myocardial infarction in the mouse. J. Am. Coll. Cardiol. 2005, 46, 2116–2124. [Google Scholar] [CrossRef] [PubMed]
- Holscher, M.; Schafer, K.; Krull, S.; Farhat, K.; Hesse, A.; Silter, M.; Lin, Y.; Pichler, B.J.; Thistlethwaite, P.; El-Armouche, A.; et al. Unfavourable consequences of chronic cardiac hif-1alpha stabilization. Cardiovasc. Res. 2012, 94, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Moslehi, J.; Minamishima, Y.A.; Shi, J.; Neuberg, D.; Charytan, D.M.; Padera, R.F.; Signoretti, S.; Liao, R.; Kaelin, W.G., Jr. Loss of hypoxia-inducible factor prolyl hydroxylase activity in cardiomyocytes phenocopies ischemic cardiomyopathy. Circulation 2010, 122, 1004–1016. [Google Scholar] [CrossRef] [PubMed]
- Crnko, S.; Du Pre, B.C.; Sluijter, J.P.G.; Van Laake, L.W. Circadian rhythms and the molecular clock in cardiovascular biology and disease. Nat. Rev. Cardiol. 2019, 16, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Alibhai, F.J.; LaMarre, J.; Reitz, C.J.; Tsimakouridze, E.V.; Kroetsch, J.T.; Bolz, S.S.; Shulman, A.; Steinberg, S.; Burris, T.P.; Oudit, G.Y.; et al. Disrupting the key circadian regulator clock leads to age-dependent cardiovascular disease. J. Mol. Cell. Cardiol. 2017, 105, 24–37. [Google Scholar] [CrossRef]
- Martino, T.A.; Oudit, G.Y.; Herzenberg, A.M.; Tata, N.; Koletar, M.M.; Kabir, G.M.; Belsham, D.D.; Backx, P.H.; Ralph, M.R.; Sole, M.J. Circadian rhythm disorganization produces profound cardiovascular and renal disease in hamsters. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R1675–R1683. [Google Scholar] [CrossRef]
- Sakamuri, S.S.; Takawale, A.; Basu, R.; Fedak, P.W.; Freed, D.; Sergi, C.; Oudit, G.Y.; Kassiri, Z. Differential impact of mechanical unloading on structural and nonstructural components of the extracellular matrix in advanced human heart failure. Transl. Res. 2016, 172, 30–44. [Google Scholar] [CrossRef]
- Takawale, A.; Sakamuri, S.S.; Kassiri, Z. Extracellular matrix communication and turnover in cardiac physiology and pathology. Compr. Physiol. 2015, 5, 687–719. [Google Scholar] [CrossRef]
- Jana, S.; Zhang, H.; Lopaschuk, G.D.; Freed, D.H.; Sergi, C.; Kantor, P.F.; Oudit, G.Y.; Kassiri, Z. Disparate remodeling of the extracellular matrix and proteoglycans in failing pediatric versus adult hearts. J. Am. Heart Assoc. 2018, 7, e010427. [Google Scholar] [CrossRef]
- Spinale, F.G. Myocardial matrix remodeling and the matrix metalloproteinases: Influence on cardiac form and function. Physiol. Rev. 2007, 87, 1285–1342. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.C.; Zhang, Z.Z.; Wang, W.; McKinnie, S.M.K.; Vederas, J.C.; Oudit, G.Y. Targeting the apelin pathway as a novel therapeutic approach for cardiovascular diseases. Biochim. Biophys. Acta 2017, 1863, 1942–1950. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, A.A.; Vergara, A.; Wang, X.; Vederas, J.C.; Oudit, G.Y. Apelin pathway in cardiovascular, kidney, and metabolic diseases: Therapeutic role of apelin analogs and apelin receptor agonists. Peptides 2021, 147, 170697. [Google Scholar] [CrossRef]
- Voelkel, N.F.; Quaife, R.A.; Leinwand, L.A.; Barst, R.J.; McGoon, M.D.; Meldrum, D.R.; Dupuis, J.; Long, C.S.; Rubin, L.J.; Smart, F.W.; et al. Right ventricular function and failure: Report of a national heart, lung, and blood institute working group on cellular and molecular mechanisms of right heart failure. Circulation 2006, 114, 1883–1891. [Google Scholar] [CrossRef] [PubMed]
- Shahinian, J.H.; Mayer, B.; Tholen, S.; Brehm, K.; Biniossek, M.L.; Fullgraf, H.; Kiefer, S.; Heizmann, U.; Heilmann, C.; Ruter, F.; et al. Proteomics highlights decrease of matricellular proteins in left ventricular assist device therapydagger. Eur. J. Cardiothorac. Surg. 2017, 51, 1063–1071. [Google Scholar] [CrossRef][Green Version]
- Maybaum, S.; Mancini, D.; Xydas, S.; Starling, R.C.; Aaronson, K.; Pagani, F.D.; Miller, L.W.; Margulies, K.; McRee, S.; Frazier, O.H.; et al. Cardiac improvement during mechanical circulatory support: A prospective multicenter study of the lvad working group. Circulation 2007, 115, 2497–2505. [Google Scholar] [CrossRef]
- Bruckner, B.A.; Stetson, S.J.; Farmer, J.A.; Radovancevic, B.; Frazier, O.H.; Noon, G.P.; Entman, M.L.; Torre-Amione, G.; Youker, K.A. The implications for cardiac recovery of left ventricular assist device support on myocardial collagen content. Am. J. Surg. 2000, 180, 498–501. [Google Scholar] [CrossRef]
- Bruckner, B.A.; Stetson, S.J.; Perez-Verdia, A.; Youker, K.A.; Radovancevic, B.; Connelly, J.H.; Koerner, M.M.; Entman, M.E.; Frazier, O.H.; Noon, G.P.; et al. Regression of fibrosis and hypertrophy in failing myocardium following mechanical circulatory support. J. Heart Lung Transplant. 2001, 20, 457–464. [Google Scholar] [CrossRef]
- Fan, D.; Takawale, A.; Lee, J.; Kassiri, Z. Cardiac fibroblasts, fibrosis and extracellular matrix remodeling in heart disease. Fibrogenes. Tissue Repair 2012, 5, 15. [Google Scholar] [CrossRef]
- Knutson, A.K.; Williams, A.L.; Boisvert, W.A.; Shohet, R.V. Hif in the heart: Development, metabolism, ischemia, and atherosclerosis. J. Clin. Investig. 2021, 131, e137557. [Google Scholar] [CrossRef]
- Oudit, G.Y.; Penninger, J.M. Cardiac regulation by phosphoinositide 3-kinases and pten. Cardiovasc. Res. 2009, 82, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.B.; Zhabyeyev, P.; Chen, X.; Wang, F.; Paul, M.; Fan, D.; McLean, B.A.; Basu, R.; Zhang, P.; Shah, S.; et al. Pi3kalpha-regulated gelsolin activity is a critical determinant of cardiac cytoskeletal remodeling and heart disease. Nat. Commun. 2018, 9, 5390. [Google Scholar] [CrossRef] [PubMed]
- Grabellus, F.; Schmid, C.; Levkau, B.; Breukelmann, D.; Halloran, P.F.; August, C.; Takeda, N.; Takeda, A.; Wilhelm, M.; Deng, M.C.; et al. Reduction of hypoxia-inducible heme oxygenase-1 in the myocardium after left ventricular mechanical support. J. Pathol. 2002, 197, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Park, S.; Li, Y.; Missov, E.; Hou, M.; Han, X.; Hall, J.L.; Miller, L.W.; Bache, R.J. Alterations of gene expression in failing myocardium following left ventricular assist device support. Physiol. Genom. 2003, 14, 251–260. [Google Scholar] [CrossRef]
- Heineke, J.; Molkentin, J.D. Regulation of cardiac hypertrophy by intracellular signalling pathways. Nat. Rev. Mol. Cell Biol. 2006, 7, 589–600. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, J.; Jin, X.; Li, C.; Zhou, G.; Feng, J. Nrf1-enhanced mir-4458 alleviates cardiac hypertrophy through releasing ttp-inhibited tfam. Vitr. Cell. Dev. Biol. Anim. 2020, 56, 120–128. [Google Scholar] [CrossRef]
- Liu, X.; Wang, J.; Zhang, G. Mir-4458 regulates cell proliferation and apoptosis through targeting socs1 in triple-negative breast cancer. J. Cell Biochem. 2019, 120, 12943–12948. [Google Scholar] [CrossRef]
- Wu, M.; Tang, Y.; Hu, G.; Yang, C.; Ye, K.; Liu, X. Mir-4458 directly targets igf1r to inhibit cell proliferation and promote apoptosis in hemangioma. Exp. Ther. Med. 2020, 19, 3017–3023. [Google Scholar] [CrossRef]
- Kura, B.; Kalocayova, B.; Devaux, Y.; Bartekova, M. Potential clinical implications of mir-1 and mir-21 in heart disease and cardioprotection. Int. J. Mol. Sci. 2020, 21, 700. [Google Scholar] [CrossRef]
- Duygu, B.; Da Costa Martins, P.A. Mir-21: A star player in cardiac hypertrophy. Cardiovasc. Res. 2015, 105, 235–237. [Google Scholar] [CrossRef]
- Bang, C.; Batkai, S.; Dangwal, S.; Gupta, S.K.; Foinquinos, A.; Holzmann, A.; Just, A.; Remke, J.; Zimmer, K.; Zeug, A.; et al. Cardiac fibroblast-derived microrna passenger strand-enriched exosomes mediate cardiomyocyte hypertrophy. J. Clin. Investig. 2014, 124, 2136–2146. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Chen, C.; Gong, W.; Yin, Z.; Zhou, L.; Chaugai, S.; Wang, D.W. Mir-21-3p regulates cardiac hypertrophic response by targeting histone deacetylase-8. Cardiovasc. Res. 2015, 105, 340–352. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.C.; Yamada, K.A.; Patel, A.Y.; Topkara, V.K.; George, I.; Cheema, F.H.; Ewald, G.A.; Mann, D.L.; Nerbonne, J.M. Deep rna sequencing reveals dynamic regulation of myocardial noncoding rnas in failing human heart and remodeling with mechanical circulatory support. Circulation 2014, 129, 1009–1021. [Google Scholar] [CrossRef] [PubMed]
- Litvinukova, M.; Talavera-Lopez, C.; Maatz, H.; Reichart, D.; Worth, C.L.; Lindberg, E.L.; Kanda, M.; Polanski, K.; Heinig, M.; Lee, M.; et al. Cells of the adult human heart. Nature 2020, 588, 466–472. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Criteria | No LVAD (n = 8) | LVAD (n = 8) | p Value |
|---|---|---|---|
| Age at transplant, years | 50 (43–54) | 57 (45–59) | 0.7814 |
| Female sex | 1 (12.5) | 1 (12.5) | >0.9999 |
| SBP, mmHg | 92 (89–97) | 91 (80–103) | 0.8131 |
| HR, bpm | 86 (69–106) | 80 (77–96) | 0.8935 |
| BMI, kg/m2 | 24 (23–27) | 25 (24–29) | 0.3949 |
| NYHA class | |||
| III | 6 (75) | 4 (50) | 0.5594 |
| IV | 1 (12.5) | 3 (37.5) | 0.5594 |
| Medical history | |||
| HF duration, mo | 39 (6–108) | 48 (30–120) | 0.5493 |
| Hypertension | 1 (12.5) | 1 (12.5) | >0.9999 |
| Dyslipidemia | 0 (0) | 2 (25) | 0.4667 |
| Kidney disease | 2 (25) | 2 (25) | >0.9999 |
| Liver disease | 4 (50) | 2 (25) | 0.3147 |
| Diabetes | 0 (0) | 1 (12.5) | >0.9999 |
| COPD | 1 (12.5) | 1 (12.5) | >0.9999 |
| Discharge medication | |||
| ACEi/ARB/sacubitril/valsartan | 7 (87.5) | 6 (75) | 0.3147 |
| Beta-blocker | 7 (87.5) | 6 (75) | >0.9999 |
| MRA | 2 (25) | 4 (50) | 0.6084 |
| Laboratory | |||
| Cr, umol/L | 108 (95–118) | 83 (78–112) | 0.9511 |
| eGFR, mL/m in/1.73 m2 | 61 (56–69) | 89 (65–112) | 0.2811 |
| Hb, g/L | 118 (107–131) | 98 (92–109) | 0.1229 |
| Echocardiography | |||
| EF ≤ 50% | 7 (87.5) | 4 (50) | >0.9999 |
| EF, % | 18 (15–26) | 13 (10–20) | 0.6223 |
| LVEDD, mm | 64 (59–69) | 57 (51–62) | 0.3556 |
| LVESD, mm | 60 (54–66) | 44 (38–59) | 0.1523 |
| ID | Fold Change | p-Value | Target Transcripts from Corresponding mRNA Comparisons | |
|---|---|---|---|---|
| Number of Transcripts | Target Genes | |||
| hsa-miR-451 | 16.22 | 4.30 × 10−8 | 43 | CCL18, FRZB, ACTC1, EIF1AY, C3AR1, RAMP1, AQP4, PLP2, F5, SLC35F1, KCNJ8, SLC25A27, LY96, STK38L, LDHA, TNFAIP8, EGLN3, ATP5F1, GPRASP1, UTY, SSPN, CTH, SORL1, GMNN, PTPRB |
| hsa-miR-182 | 15.33 | 1.03 × 10−6 | 41 | IGF1R, DNER, BCAT1, CREB3L1, RARRES1, RNASE6, TPST2, HIF3A, EMB, MGST1, FCN3, MARK3, FIGF, MLF1, SLITRK4, BDNF, HS3ST1, SAMSN1, GABRB1, RSAD2, AASS, MYH6, ZMYND12, PKD1L2, TM2D3, DCN |
| hsa-miR-495 | 2.87 | 1.14 × 10−5 | 30 | C3, CITED2, DNER, ALOX5AP, ACTC1, BCAT1, CRYM, AQP4, HSPA2, FAM46B, LARP6, CADM1, ATP5F1, GPR22, LRRTM4, RSAD2, AASS, DUSP27, AGTPBP1, PCDH20, HCLS1, TUBA1C, METTL7A |
| hsa-miR-135b | −3.95 | 0.0003 | 30 | SERPINA3, CD53, GADD45A, FCER1G, RNASE6, IFIT2, ADH1A, FGF1, TIMP4, ANP32E, LILRB2, EFCAB2, RNASE2, CYP4Z1, ITIH5, IL1RL1, GABRB1, MAEL, RASAL2, HAS2, SLCO4A1, AGTPBP1, PTPRB |
| hsa-miR-374b | 2.35 | 0.0005 | 25 | F13A1, GADD45A, EGR1, CHST2, ASPN, CCL18, BIRC3, STK38L, ANKRD1, CTH, GOLGA8A, TMED5, ATP2A2, CNKSR2, PTPRB |
| hsa-miR-218 | 4.42 | 0.0017 | 41 | PER3, FRZB, CRYM, PLCE1, IFI44L, DNAJB5, RABGAP1L, HAPLN1, SMTNL2, HEY2, ANKRD1, MLF1, CA2, ANK1, OGN, GABRB1, AASS, DUSP27, PHACTR1, GMNN, ST8SIA5, ASS1, UCHL1, RHOBTB1, DCN, EDIL3 |
| hsa-miR-208a | −2.58 | 0.0018 | 24 | SH3BGRL3, HMOX1, NPTX2, EMB, IFI44L, MGST1, TIMP4, LRRC1, MLF1, GPRASP1, C6, PDCD4, AGTPBP1, CPAMD8, HK2, BLM, TUBA1C |
| hsa-miR-373 | 5.76 | 0.0026 | 27 | CD24, MAP4, NINJ2, PLCE1, MYBL1, C2orf40, GPR34, CD68, HS3ST1, CSAD, CNN1, ITIH5, GPRASP1, CHRFAM7A, SLMAP, SCUBE2, PRRT2, PKD1L2, UCHL1, IFI44 |
| hsa-miR-628-5p | 2.59 | 0.0037 | 32 | CTSC, SMOC2, SLCO2A1, SLC1A2, BCL6, PLCE1, REPS2, OGDHL, PTN, DPY19L2, TYRP1, KCNIP2, C4A, MYOC, SCUBE2, GMNN, TSPAN13, BLM, PECAM1, TYROBP, ATP2A2 |
| hsa-miR-431 | −2.33 | 0.0055 | 20 | CD53, SLC7A8, NPR3, LARP6, ITGAM, MYBPC1, STAT4, ITIH5, WISP2, GSG1L, CLEC10A, PHACTR1, ARPC3, CPAMD8, DDAH1 |
| hsa-miR-224 | 2.82 | 0.0058 | 30 | HLA-C, PACSIN3, TPST2, CYP2J2, HIF3A, GRB14, OGDHL, TMEM45A, HSD17B11, ARL4C, CHST9, CA2, GSG1L, C6, CTH, ARPC3, FHL1, HMOX2, ASS1, PTPRB |
| hsa-miR-95 | 4.09 | 0.0069 | 36 | SERPINA3, C1QB, TUBB2A, PENK, TBXAS1, PLK2, CA14, TTC9, IL20RA, EDARADD, ST8SIA2, TFPI, CHST9, EGLN3, IL1RL1, GPRASP1, GSG1L, RSAD2, MGAT4C, ENO1, GMNN, PTPRB |
| hsa-miR-940 | −2.26 | 0.0078 | 17 | SH3BGRL3, SERPINA3, S100A9, NES, SERPINA1, HIF3A, GCHFR, CENPA, CYP4B1, CAPG, TNFRSF12A, IFI30 |
| hsa-miR-601 | −2.29 | 0.0134 | 23 | PIM1, CD109, PLA2G2A, C1QC, SLC6A6, CYP2J2, GCHFR, CENPA, C1orf162, LILRB2, NR4A3, GRK5, TPM3, CA2, HMOX2 |
| hsa-miR-329 | 2.22 | 0.0148 | 25 | CITED2, HIF3A, AHNAK, KCNJ8, DPY19L2, CADM1, CD68, LRRTM4, PHACTR1, PRDX6, CEBPD, CAMK1D, NNMT, SLMAP, PKD1L2, AQP3, OBSCN |
| hsa-miR-187 | −6.09 | 0.0186 | 33 | SH3BGRL3, CHGB, S100A4, IGF1R, ALOX5AP, CYP2J2, PLP2, FAM58A, PRIMA1, MGST1, DPY19L2, HFE2, LILRB2, STK38L, STAT4, WISP2, GSG1L, DUSP27, PHACTR1, MXRA5, S100A8, CPAMD8, UCHL1, OBSCN |
| hsa-miR-10b | 2.55 | 0.0238 | 36 | APOD, FCER1G, RAMP1, DIO2, TMEM97, CA14, MYBL1, FAM46B, SMTNL2, GRK5, CD68, TNFRSF11B, LCN6, CNN1, ANK1, ATP5F1, SGPP2, C10orf71, PRDX6, SLMAP, SCUBE2, AGTPBP1, PKD1L2, OBSCN |
| hsa-miR-223 | 2.99 | 0.0311 | 51 | GADD45A, PER3, AKR1C1, RRAS2, UBR1, PLCE1, RABGAP1L, MYO5A, TDRD9, GPR34, C1orf162, VIT, SLC14A1, EFCAB2, TFPI, ANK1, GPR22, OGN, MAEL, FKBP5, DUSP27, HVCN1, FHL1, CNKSR2 |
| hsa-miR-4269 | 2.38 | 2.70E-06 | - | - |
| hsa-miR-299-5p | 3.58 | 9.71 × 10−5 | - | - |
| hsa-miR-4270 | −2.12 | 0.0001 | - | - |
| hsa-miR-4539 | −2.06 | 0.0003 | - | - |
| hsa-miR-1825 | −2.04 | 0.0007 | - | - |
| hsa-miR-3187-3p | −2.4 | 0.001 | - | - |
| ENSG00000202498 | −2.28 | 0.0013 | - | - |
| ENSG00000202498_x | −2.31 | 0.0016 | - | - |
| hsa-miR-3910 | −2.03 | 0.0045 | - | - |
| hsa-miR-4687-3p | −2.22 | 0.0057 | - | - |
| hsa-miR-4741 | −2.08 | 0.0071 | - | - |
| hsa-miR-548x | −2 | 0.0077 | - | - |
| hsa-miR-4689 | −2.28 | 0.0107 | - | - |
| hsa-miR-3128 | −2.3 | 0.0109 | - | - |
| hsa-miR-4793-3p | −2.69 | 0.011 | - | - |
| hsa-miR-3201 | −2.11 | 0.0114 | - | - |
| hsa-miR-103b | −2.58 | 0.015 | - | - |
| hsa-miR-1226 | 4.09 | 0.0158 | - | - |
| hsa-miR-4458 | 2.21 | 0.0198 | - | - |
| hsa-miR-4521 | −3.93 | 0.0275 | - | - |
| hsa-miR-1226 | −2.12 | 0.0304 | - | - |
| ID | Fold Change | p-Value | Target Transcripts from Corresponding mRNA Comparisons | |
|---|---|---|---|---|
| Number of Transcripts | Target Genes | |||
| hsa-miR-182 | 19.89 | 6.64 × 10−6 | 48 | ANXA1, IL1R1, DNER, BCAT1, CREB3L1, RARRES1, RNASE6, PC, EMB, MGST1, APOB, FCN3, ACE2, MARK3, OLFM1, FIGF, CLEC4G, KCNN3, KCNMB2, PTGER3, SAMSN1, GABRB1, RSAD2, MYH6, ATP1B3, ZMYND12, MLF1, PKD1L2, CHI3L2, TM2D3, DCN |
| hsa-miR-124 | −2.16 | 0.0002 | 38 | CDKN1A, IQGAP1, IL1R1, CD59, RDH10, PENK, BCAT1, NAP1L3, GCHFR, AGTR1, KLF15, OLFM1, IRAK3, PAPSS2, CADM1, RNASE2, TRIM45, RGS4, RBM47, CYP4B1, NID1, SLCO4A1, ARPC1B, GOLGA8A, CPAMD8, TWIST2, C1S, TSC22D1 |
| hsa-miR-451 | 7.13 | 0.0004 | 38 | KLF6, HCK, CCL18, FRZB, ACTC1, C3AR1, RAMP1, AQP4, F5, CDKN2B, LY96, STK38L, B3GNT7, FYB, SCGB1D2, TNFAIP8, GPRASP1, MAN1A1, SSPN, CTH, SAT1, CTHRC1, ARPC1B, UTY |
| hsa-miR-181a-2 | 2.34 | 0.0006 | 20 | VAMP8, RBP4, SOX4, IER3, ACLY, DIO2, ABCG2, C1orf162, CD163, SUSD4, ANKRD1, TPO, PHACTR1, NNMT, AQP3 |
| hsa-miR-373 | 6.81 | 0.0009 | 25 | CD24, KLF6, ENSA, NINJ2, PLCE1, UAP1, MYBL1, GPR34, CD68, MYPN, CNN1, ITIH5, GPRASP1, RASSF2, CHRFAM7A, IFI44, SCUBE2, PKD1L2 |
| hsa-miR-138 | −2.39 | 0.0013 | 28 | LYZ, EIF4EBP1, FCER1G, CSTA, C3AR1, ADH1A, DOCK2, UAP1, APBB1IP, FIGF, VIT, TPM4, ITGAM, LCN6, SYNPO2L, TMEM74, STON1, PHACTR1, PKD1L2, C1S, DCN |
| hsa-miR-431 | −4.17 | 0.0016 | 24 | CD53, IGFBP6, HCK, SLC7A8, NPR3, LARP6, BMPR1B, HPR, ITGAM, MYBPC1, HP, ITIH5, TPO, CLEC10A, PHACTR1, CPAMD8, COL12A1 |
| hsa-miR-92a-1 | −2.77 | 0.0025 | 29 | TIMP1, SH3BGRL3, SERPINA3, SEC14L1, AIF1, PLA2G2A, FCER1G, RAMP1, UAP1, METTL7B, LY96, GPX3, DPYSL4, ITIH5, PHACTR1, SAT1, CTHRC1, DUSP4, TKT, TYROBP |
| hsa-miR-21 | −2.87 | 0.0034 | 22 | SMOC2, IGFBP6, ALOX5AP, PENK, BCAT1, NQO2, BIRC3, CENPA, TDRD9, LARP6, TNFSF12, RGS4, C5AR1, ITIH5, MAP3K8, PKD1L2, ASRGL1 |
| hsa-miR-10b | 2.64 | 0.0051 | 26 | ENSA, FCER1G, RAMP1, DIO2, CA14, HS6ST2, UAP1, MYBL1, SMTNL2, GRK5, CD68, SCN3A, LCN6, CNN1, C10orf71, SRGN, PRDX6, ARPC1B, SCUBE2, AGTPBP1, PKD1L2 |
| hsa-miR-95 | 2.86 | 0.0193 | 38 | SERPINA3, EMP1, C1QB, TUBB2A, PENK, TBXAS1, CA14, ACE2, EDARADD, ST8SIA2, CHST9, SHMT1, IL1RL1, GPRASP1, RSAD2, STON1, MGAT4C, SAT1, ENO1, CHI3L2, CHI3L1, GADD45B |
| hsa-miR-217 | 3.1 | 0.0487 | 38 | CHST7, ENSA, HMOX1, MFAP5, KCNS3, FNDC1, CENPA, DOCK2, COL21A1, GFRA1, FCGR2A, FIGF, STRBP, CD163, HSPA4L, MXI1, MYPN, FGF10, STON1, DUSP27, MGAT4C, SAT1, CAMK1D, CPAMD8, PKD1L2 |
| hsa-miR-216a | 3.02 | 0.0488 | 25 | KLF6, ENSA, FCER1G, NPR3, IFI44L, PITPNC1, STK17B, HFE2, VIT, CADM1, ITIH5, PTGER3, CTH, CYP4B1, ZMYND12, CCDC109B, HCLS1, CHI3L2 |
| HBII-52-32_x | 4.76 | 0.0028 | - | - |
| hsa-miR-1972 | 2.57 | 0.0046 | - | - |
| hsa-miR-3065-3p | −3.04 | 0.0068 | - | - |
| hsa-miR-4524 | 2.05 | 0.0101 | - | - |
| hsa-miR-1247 | −2.56 | 0.0187 | - | - |
| hsa-miR-4461 | 2.3 | 0.0321 | - | - |
| ID | FC in NVAD vs. NFC | FC in VAD vs. NFC | p-Value in VAD vs. NFC |
|---|---|---|---|
| Insignificant change compared to NFC | |||
| hsa-miR-4458_st | 2.21 | 1.73 | 0.5994 |
| hsa-miR-4793-3p_st | −2.69 | −1.96 | 0.3478 |
| hsa-miR-187_st | −6.09 | −1.68 | 0.3441 |
| hsa-miR-373_st | 5.76 | 1.48 | 0.1852 |
| hsa-miR-103b_st | −2.58 | −1.81 | 0.096 |
| hsa-miR-95_st | 4.09 | 2.58 | 0.0677 |
| hsa-miR-431_st | −2.33 | −1.76 | 0.0659 |
| hsa-miR-1226_st | 4.09 | 2.2 | 0.064 |
| Decreased in FC | |||
| hsa-miR-182_st | 15.33 | 9.68 | 3.58 × 10−6 |
| hsa-miR-451_st | 16.22 | 10.96 | 3.10 × 10−8 |
| hsa-miR-3187-3p_st | −2.4 | −1.37 | 0.0139 |
| hsa-miR-548x_st | −2 | −1.36 | 0.0076 |
| hsa-miR-299-5p_st | 3.58 | 2.95 | 0.0019 |
| hsa-miR-4539_st | −2.06 | −1.58 | 0.03 |
| hsa-miR-628-5p_st | 2.59 | 2.17 | 0.0047 |
| hsa-miR-1226-star_st | −2.12 | −1.76 | 0.0171 |
| hsa-miR-4687-3p_st | −2.22 | −1.91 | 0.0144 |
| ENSG00000202498_x_st | −2.31 | −2.08 | 0.0044 |
| hsa-miR-223_st | 2.99 | 2.8 | 0.0225 |
| hsa-miR-10b_st | 2.55 | 2.38 | 0.0145 |
| hsa-miR-374b_st | 2.35 | 2.21 | 0.0015 |
| ENSG00000202498_st | −2.28 | −2.16 | 0.0012 |
| hsa-miR-4689_st | −2.28 | −2.16 | 0.046 |
| hsa-miR-1825_st | −2.04 | −1.93 | 0.0033 |
| hsa-miR-601_st | −2.29 | −2.19 | 0.0158 |
| hsa-miR-940_st | −2.26 | −2.19 | 0.0282 |
| hsa-miR-4741_st | −2.08 | −2.02 | 0.005 |
| hsa-miR-3910_st | −2.03 | −1.99 | 0.0172 |
| hsa-miR-4270_st | −2.12 | −2.09 | 9.94 × 10−5 |
| ID | FC in NVAD vs. NFC | FC in VAD vs. NFC | p-Value in VAD vs. NFC |
|---|---|---|---|
| Insignificant change compared to NFC | |||
| hsa-miR-1972_st | 2.57 | 1.22 | 0.8742 |
| hsa-miR-4461_st | 2.3 | 1.14 | 0.7984 |
| hsa-miR-21-star_st | −2.87 | −1.27 | 0.647 |
| hsa-miR-4524-star_st | 2.05 | 1.32 | 0.3365 |
| hsa-miR-373_st | 6.81 | 1.66 | 0.0801 |
| hsa-miR-431_st | −4.17 | −3.62 | 0.0738 |
| hsa-miR-1247_st | −2.56 | −2.33 | 0.0583 |
| Decreased in FC | |||
| hsa-miR-182_st | 19.89 | 11.3 | 1.75 × 10−5 |
| hsa-miR-451_st | 7.13 | 5.16 | 0.0002 |
| HBII-52-32_x_st | 4.76 | 3.48 | 0.0029 |
| hsa-miR-181a-2-star_st | 2.34 | 1.91 | 0.0098 |
| hsa-miR-10b_st | 2.64 | 2.53 | 0.0068 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parikh, M.; Shah, S.; Basu, R.; Famulski, K.S.; Kim, D.; Mullen, J.C.; Halloran, P.F.; Oudit, G.Y. Transcriptomic Signatures of End-Stage Human Dilated Cardiomyopathy Hearts with and without Left Ventricular Assist Device Support. Int. J. Mol. Sci. 2022, 23, 2050. https://doi.org/10.3390/ijms23042050
Parikh M, Shah S, Basu R, Famulski KS, Kim D, Mullen JC, Halloran PF, Oudit GY. Transcriptomic Signatures of End-Stage Human Dilated Cardiomyopathy Hearts with and without Left Ventricular Assist Device Support. International Journal of Molecular Sciences. 2022; 23(4):2050. https://doi.org/10.3390/ijms23042050
Chicago/Turabian StyleParikh, Mihir, Saumya Shah, Ratnadeep Basu, Konrad S. Famulski, Daniel Kim, John C. Mullen, Philip F. Halloran, and Gavin Y. Oudit. 2022. "Transcriptomic Signatures of End-Stage Human Dilated Cardiomyopathy Hearts with and without Left Ventricular Assist Device Support" International Journal of Molecular Sciences 23, no. 4: 2050. https://doi.org/10.3390/ijms23042050
APA StyleParikh, M., Shah, S., Basu, R., Famulski, K. S., Kim, D., Mullen, J. C., Halloran, P. F., & Oudit, G. Y. (2022). Transcriptomic Signatures of End-Stage Human Dilated Cardiomyopathy Hearts with and without Left Ventricular Assist Device Support. International Journal of Molecular Sciences, 23(4), 2050. https://doi.org/10.3390/ijms23042050