
Effects of Itxasol© Components on Gene Expression in Bacteria Related to Infections of the Urinary Tract and to the Inflammation Process


Abstract
:1. Introduction
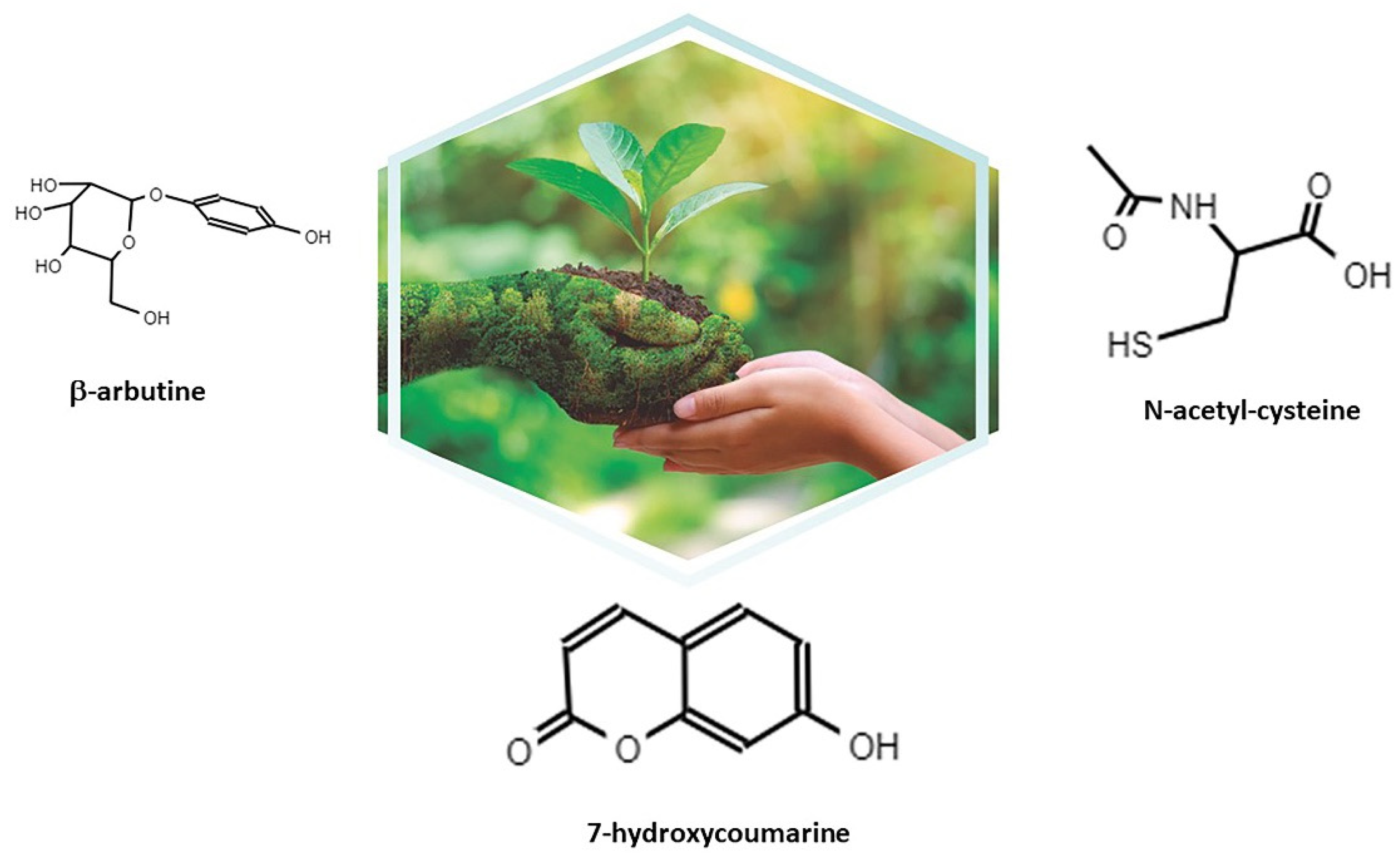
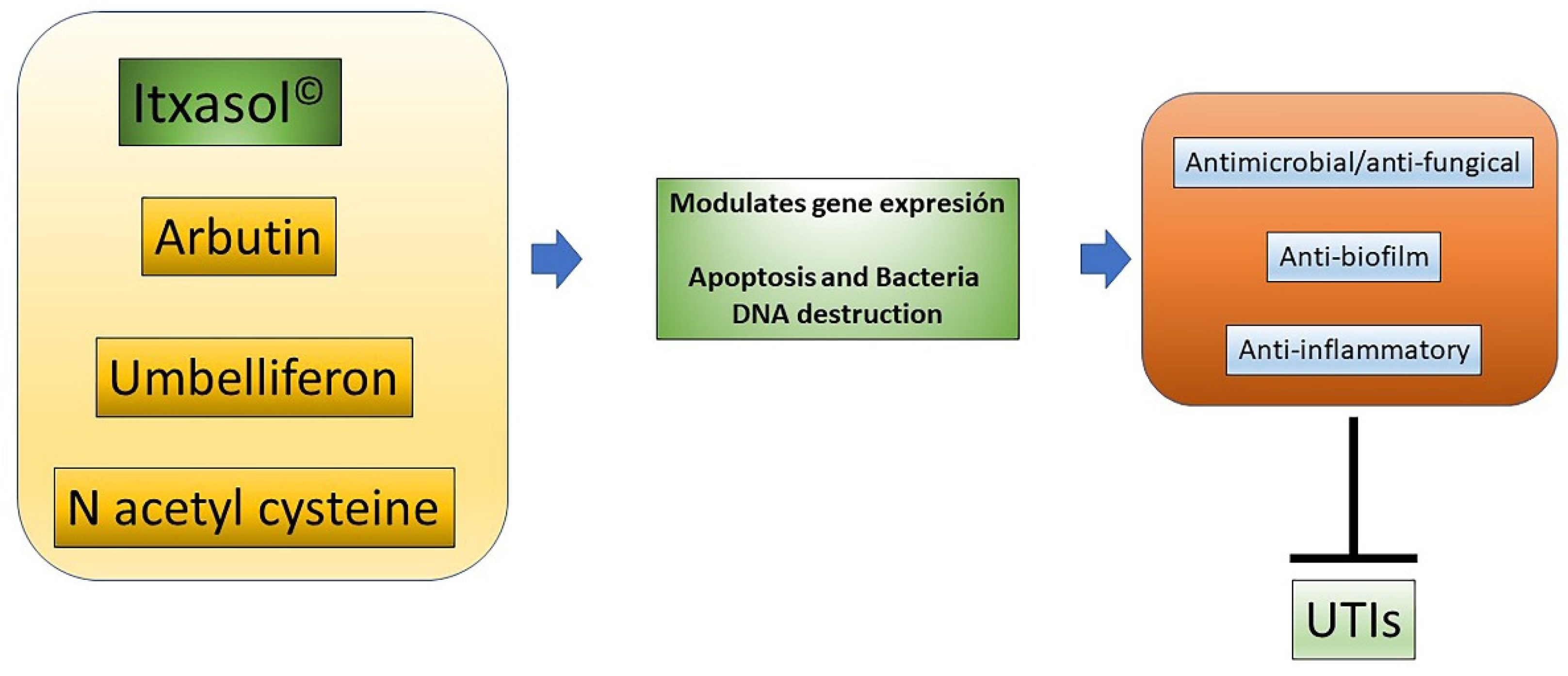
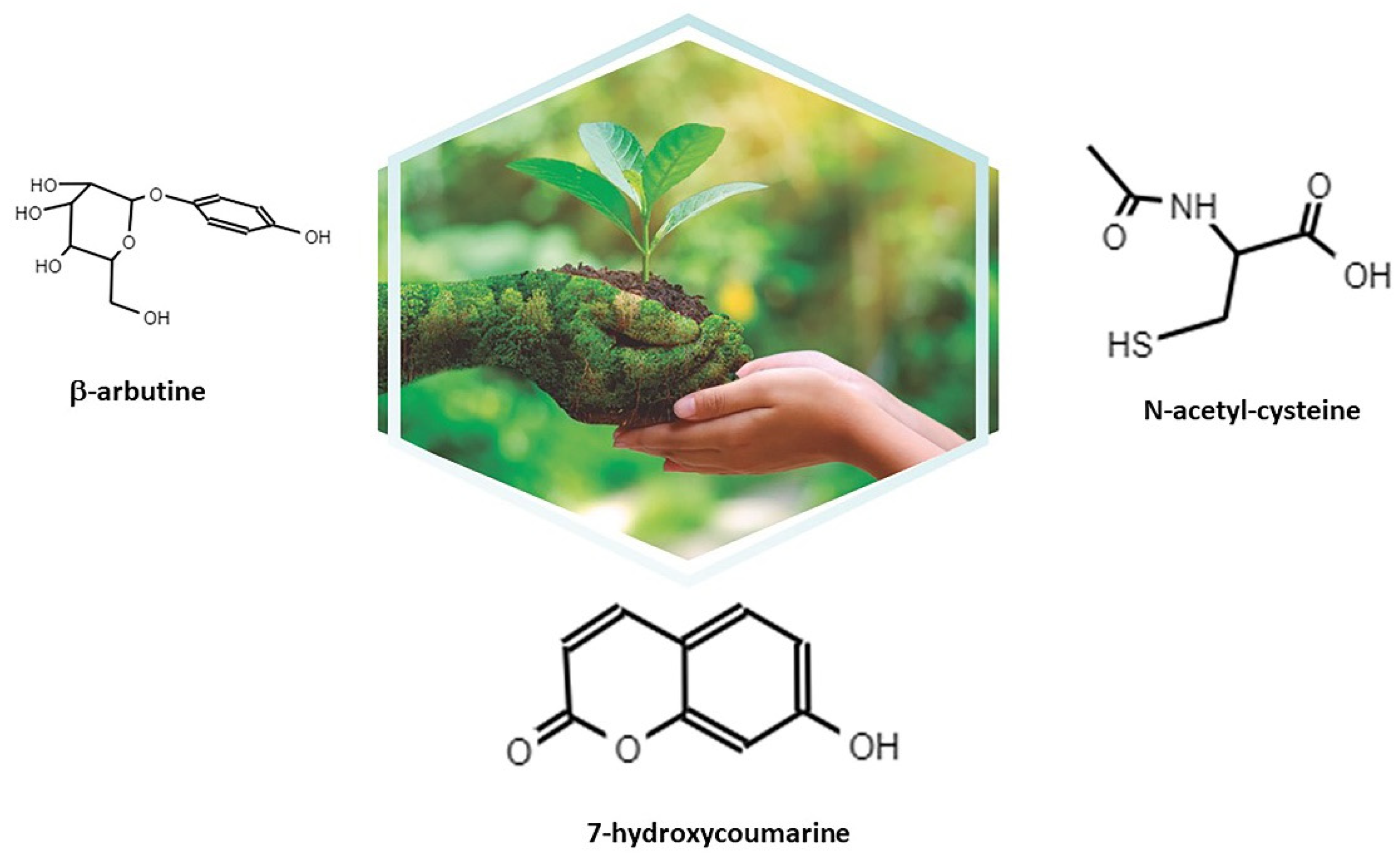
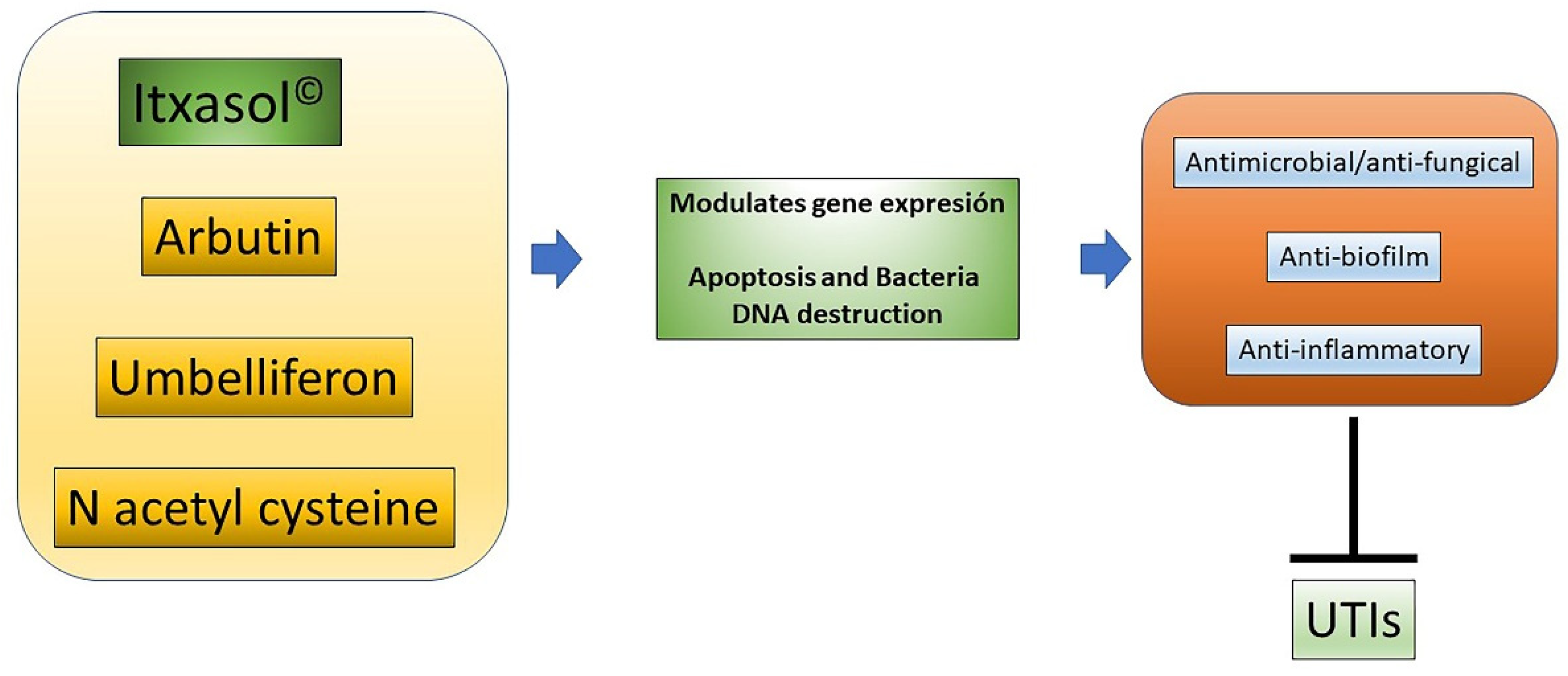
2. Itxasol© and Its Formulation
2.1. β-Arbutine
2.2. Umbelliferon (Umb)
2.3. NAC
3. Influence of Itxasol© in the Expression of Genes Related to Inflammation and Changes in Bacterial Genomics
3.1. β-Arbutin
3.2. Umbelliferon (Umb)

{kind=link}
{kind=link}
| Action/Finding | Reference |
|---|---|
| Downregulation of genes involved in biofilm production and adhesion | [60] |
| Downregulation of genes related to production of extracellular matrix and motility | [61] |
| Attenuation of DNA damage for oxidative stress | [62] |
| Reduction of inflammasome | [63] |
| Reduction of apoptosis of kidney cells | [64] |
| Produces DNA fragmentation in oral carcinoma cells | [65] |
| Cell cycle arrest in G1 apoptosis of adenocarcinoma cells | [66] |
3.3. N-acetyl-L-cysteine (NAC)
4. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Foxman, B. The Epidemiology of Urinary Tract Infection. Nat. Rev. Urol. 2010, 7, 653–660. [Google Scholar] [CrossRef]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary Tract Infections: Epidemiology, Mechanisms of Infection and Treatment Options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef]
- Krzysztof, C.; Broś-Konopielko, M.; Teliga-Czajkowska, J. Urinary Tract Infection in Women. Prz. Menopauzalny 2021, 20, 40–47. [Google Scholar] [CrossRef]
- Anger, J.T.; Saigal, C.S.; Wang, M.M.; Yano, E.M. Urologic Disease Burden in the United States: Veteran Users of Department of Veterans Affairs Healthcare. Urology 2008, 72, 37–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jennifer, A.; Lee, U.; Ackerman, A.L.; Chou, R.; Chughtai, B.; Clemens, J.Q.; Hickling, D.; Kapoor, A.; Kenton, K.S.; Kaufman, M.R.; et al. Recurrent Uncomplicated Urinary Tract Infections in Women: AUA/CUA/SUFU Guideline. J. Urol. 2019, 202, 282–289. [Google Scholar] [CrossRef]
- Mathiyalagen, P.; Peramasamy, B.; Vasudevan, K.; Basu, M.; Cherian, J.; Sundar, B. A Descriptive Cross-Sectional Study on Menstrual Hygiene and Perceived Reproductive Morbidity among Adolescent Girls in a Union Territory, India. J. Fam. Med. Prim. Care 2017, 6, 360. [Google Scholar] [CrossRef]
- Nicolle, L.E.G.; Harding, K.M.; Preiksaitis, J.; Ronald, A.R. The Association of Urinary Tract Infection with Sexual Intercourse. J. Infect. Dis. 1982, 146, 579–583. [Google Scholar] [CrossRef] [PubMed]
- Ingela, L.; Othman, J.; Hansson, M.; Ekelund, A.C.; Svanberg, T.; Strandell, A. New Types of Diaphragms and Cervical Caps versus Older Types of Diaphragms and Different Gels for Contraception: A Systematic Review. BMJ Sex. Reprod. Health 2020, 47, e12. [Google Scholar] [CrossRef]
- Jung, C.; Brubaker, L. The Etiology and Management of Recurrent Urinary Tract Infections in Postmenopausal Women. Climacteric 2019, 22, 242–249. [Google Scholar] [CrossRef]
- Manuel, E.; Galperine, T.; Caron, F. Urinary Tract Infections in Older Men. N. Engl. J. Med. 2016, 374, 2191–2192. [Google Scholar] [CrossRef]
- Linsenmeyer, T.A. Catheter-Associated Urinary Tract Infections in Persons with Neurogenic Bladders. J. Spinal Cord Med. 2018, 41, 132–141. [Google Scholar] [CrossRef]
- Betsy, F.; Gillespie, B.; Koopman, J.; Zhang, L.; Palin, K.; Tallman, P.; Marsh, J.V.; Spear, S.; Sobel, J.D.; Marty, M.J.; et al. Risk Factors for Second Urinary Tract Infection among College Women. Am. J. Epidemiol. 2000, 151, 1194–1205. [Google Scholar] [CrossRef] [Green Version]
- Griebling, T.L. Urologic Diseases in America Project: Trends in Resource Use for Urinary Tract Infections in Women. J. Urol. 2005, 173, 1281–1287. [Google Scholar] [CrossRef] [PubMed]
- Ventola, C.L. The Antibiotic Resistance Crisis: Causes and Threats. Pharm. Ther. J. 2015, 40, 277–283. [Google Scholar]
- Loukas, K.; Papachristodoulou, E.; Chra, P.; Panos, G. Mechanisms of Antibiotic Resistance in Important Gram-Positive and Gram-Negative Pathogens and Novel Antibiotic Solutions. Antibiotics 2021, 10, 415. [Google Scholar] [CrossRef]
- Morales-Alvarez, M.C. Nephrotoxicity of Antimicrobials and Antibiotics. Adv. Chronic Kidney Dis. 2020, 27, 31–37. [Google Scholar] [CrossRef]
- Kambiz, A.; Fleischmann, N.; Schmiemann, G.; Bleidorn, J.; Hummers-Pradier, E.; Friede, T.; Wegscheider, K.; Moore, M.; Gágyor, I. Reducing Antibiotic Use for Uncomplicated Urinary Tract Infection in General Practice by Treatment with Uva-Ursi (REGATTA)—A Double-Blind, Randomized, Controlled Comparative Effectiveness Trial. BMC Complement. Altern. Med. 2018, 18, 203. [Google Scholar] [CrossRef] [Green Version]
- Medina-Polo, J.; Naber, K.G.; Johansen, T.E.B. Healthcare-Associated Urinary Tract Infections in Urology. GMS Infect. Dis. 2021, 9, Doc05. [Google Scholar] [CrossRef] [PubMed]
- Tanu, A.; Azeem, K.; Husain, F.M.; Hussain, A.; Khan, M.N.; Alajmi, M.F.; Abid, M. Mechanistic Understanding of Candida albicans Biofilm Formation and Approaches for Its Inhibition. Front. Microbiol. 2021, 12, 932. [Google Scholar] [CrossRef]
- Nikky, G.; Fatima, S.W.; Kumar, S.; Sinha, R.; Khare, S.K. Antimicrobial Resistance in Biofilms: Exploring Marine Actinobacteria as a Potential Source of Antibiotics and Biofilm Inhibitors. Biotechnol. Rep. 2021, 30, e00613. [Google Scholar] [CrossRef]
- Rabin, N.; Zheng, Y.; Opoku-Temeng, C.; Du, Y.; Bonsu, E.; Sintim, H.O. Biofilm Formation Mechanisms and Targets for Developing Antibiofilm Agents. Future Med. Chem. 2015, 7, 493–512. [Google Scholar] [CrossRef]
- Goebel, M.C.; Trautner, B.W.; Grigoryan, L. The Five Ds of Outpatient Antibiotic Stewardship for Urinary Tract Infections. Clin. Microbiol. Rev. 2021, 34, e00003-20. [Google Scholar] [CrossRef]
- Heng, F.; Le, J.W.; Zhu, J.H. Protective Effect of N-Acetylcysteine Pretreatment on Acute Kidney Injury in Septic Rats. J. Surg. Res. 2020, 254, 125–134. [Google Scholar] [CrossRef]
- Yong, C.K.; Wong, S.K.; Ekeuku, S.O.; Pang, K.L. Relationship between Metabolic Syndrome and Bone Health—An Evaluation of Epidemiological Studies and Mechanisms Involved. Diabetes Metab. Syndr. Obes. Targets Ther. 2019. [Google Scholar] [CrossRef]
- Hooper, D.C. Levofloxacin. In Kucers the Use of Antibiotics: A Clinical Review of Antibacterial Antifungal, Antiparasitic, and Antiviral Drugs, 7th ed.; CRC Press: Boca Raton, FL, USA, 2017; pp. 2055–2084. [Google Scholar] [CrossRef]
- Nguyen, H.M.; Graber, C.J. A Critical Review of Cephalexin and Cefadroxil for the Treatment of Acute Uncomplicated Lower Urinary Tract Infection in the Era of ‘Bad Bugs, Few Drugs’. Int. J. Antimicrob. Agents 2020, 56, 106085. [Google Scholar] [CrossRef] [PubMed]
- Davide, C.; Siracusa, C.; Sulejmani, A.; Leoni, V.; Intra, J. Old and New Beta-Lactamase Inhibitors: Molecular Structure, Mechanism of Action and Clinical Use. Antibiotics 2021, 10, 995. [Google Scholar] [CrossRef]
- Gabriel, C.; Forestier, C.; Mathias, J.D. Antibiotic Resilience: A Necessary Concept to Complement Antibiotic Resistance? Proc. R. Soc. B Biol. Sci. 2019, 286, 20192408. [Google Scholar] [CrossRef] [Green Version]
- Beerepoot, M.A.J.; Riet, G.T.; Nys, S.; Wal, W.M.V.D.; Corianne, A.J.M.D.B.; Reijke, T.M.D.; Prins, J.M.; Koeijers, J.; Verbon, A.; Stobberingh, E.; et al. Lactobacilli vs Antibiotics to Prevent Urinary Tract Infections: A Randomized, Double-Blind, Noninferiority Trial in Postmenopausal Women. Arch. Intern. Med. 2012, 172, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhao, G.; Chao, X.; Xie, L.; Wang, H. The Characteristic of Virulence, Biofilm and Antibiotic Resistance of Klebsiella Pneumoniae. Int. J. Environ. Res. Public Health 2020, 17, 6278. [Google Scholar] [CrossRef]
- Vigil, P.D.; Stapleton, A.E.; Johnson, J.R.; Hooton, T.M.; Hodges, A.P.; He, Y.; Mobley, H.L.T. Presence of Putative Repeat-in-Toxin Gene TosA in Escherichia coli Predicts Successful Colonization of the Urinary Tract. MBio 2011, 2, e00066-11. [Google Scholar] [CrossRef] [Green Version]
- Lüthje, P.; Brauner, A. Virulence Factors of Uropathogenic E. Coli and Their Interaction with the Host. Adv. Microb. Physiol. 2014, 65, 337–372. [Google Scholar] [CrossRef]
- Sokurenko, E. Pathoadaptive Mutations in Uropathogenic Escherichia Coli. Microbiol. Spectr. 2016, 4, e01085-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupo, F.; Ingersoll, M.A.; Pineda, M.A. The Glycobiology of Uropathogenic E. Coli Infection: The Sweet and Bitter Role of Sugars in Urinary Tract Immunity. Immunology 2021, 164, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Meyrier, A. Acute Pyelonephritis. Rev. Prat. 2003, 53, 1777–1784. Available online: https://pubmed.ncbi.nlm.nih.gov/14702820/ (accessed on 1 November 2021).
- Kranz, J.; Wagenlehner, F.M.E.; Schneidewind, L. Complicated Urinary Tract Infections. Urologe 2020, 59, 1480–1485. [Google Scholar] [CrossRef] [PubMed]
- Vincent, J.F.V.; Bogatyreva, O.A.; Bogatyrev, N.R.; Bowyer, A.; Pahl, A.K. Biomimetics: Its Practice and Theory. J. R. Soc. Interface 2006, 3, 471. [Google Scholar] [CrossRef] [Green Version]
- Scribano, D.; Sarshar, M.; Prezioso, C.; Lucarelli, M.; Angeloni, A.; Zagaglia, C.; Palamara, A.T.; Ambrosi, C. D-Mannose Treatment Neither Affects Uropathogenic Escherichia coli Properties nor Induces Stable Fimh Modifications. Molecules 2020, 25, 316. [Google Scholar] [CrossRef] [Green Version]
- Bojana, K.; Papeš, D.; Altarac, S. D-Mannose Powder for Prophylaxis of Recurrent Urinary Tract Infections in Women: A Randomized Clinical Trial. World J. Urol. 2014, 32, 79–84. [Google Scholar] [CrossRef]
- Terlizzi, M.E.; Gribaudo, G.; Maffei, M.E. UroPathogenic Escherichia coli (UPEC) Infections: Virulence Factors, Bladder Responses, Antibiotic, and Non-Antibiotic Antimicrobial Strategies. Front. Microbiol. 2017, 8, 1566. [Google Scholar] [CrossRef]
- Ma, C.; He, N.; Zhao, Y.; Xia, D.; Wei, J.; Kang, W. Antimicrobial Mechanism of Hydroquinone. Appl. Biochem. Biotechnol. 2019, 189, 1291–1303. [Google Scholar] [CrossRef]
- Schindler, G.; Patzak, U.; Brinkhaus, B.; von Nieciecki, A.; Wittig, J.; Krähmer, N.; Glöckl, I.; Veit, M. Urinary Excretion and Metabolism of Arbutin after Oral Administration of Arctostaphylos Uvae Ursi Extract as Film-Coated Tablets and Aqueous Solution in Healthy Humans. J. Clin. Pharmacol. 2002, 42, 920–927. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, Y.; Song, Q.; Wang, F.; Sun, L.; Liu, L.; Yang, X.; Yi, J.; Bao, Y.; Ma, H.; et al. Anti-Biofilm Activities From Bergenia Crassifolia Leaves against Streptococcus Mutans. Front. Microbiol. 2017, 8, 1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germoush, M.O.; Othman, S.I.; Al-Qaraawi, M.A.; Al-Harbi, H.M.; Hussein, O.E.; Al-Basher, G.; Alotaibi, M.F.; Elgebaly, H.A.; Sandhu, M.A.; Allamg, A.A.; et al. Umbelliferone Prevents Oxidative Stress, Inflammation and Hematological Alterations, and Modulates Glutamate-Nitric Oxide-CGMP Signaling in Hyperammonemic Rats. Biomed. Pharmacother. 2018, 102, 392–402. [Google Scholar] [CrossRef]
- Hoult, J.R.S.; Payá, M. Pharmacological and Biochemical Actions of Simple Coumarins: Natural Products with Therapeutic Potential. Gen. Pharmacol. 1996, 27, 713–722. [Google Scholar] [CrossRef]
- Vikas, K.; Ahmed, D.; Anwar, F.; Ali, M.; Mujeeb, M. Enhanced Glycemic Control, Pancreas Protective, Antioxidant and Hepatoprotective Effects by Umbelliferon-α-D-Glucopyranosyl-(2I→1II)-α-Dglucopyranoside in Streptozotocin Induced Diabetic Rats. SpringerPlus 2013, 2, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Yao, Y.; Li, L. Coumarins as Potential Antidiabetic Agents. J. Pharm. Pharmacol. 2017, 69, 1253–1264. [Google Scholar] [CrossRef] [Green Version]
- Cal, P.M.S.D.; Matos, M.J.; Bernardes, G.J.L. Trends in Therapeutic Drug Conjugates for Bacterial Diseases: A Patent Review. Expert Opin. Ther. Pat. 2017, 27, 179–189. [Google Scholar] [CrossRef]
- Hu, X.-L.; Xu, Z.; Liu, M.-L.; Feng, L.-S.; Zhang, G.-D. Recent Developments of Coumarin Hybrids as Anti-Fungal Agents. Curr. Top. Med. Chem. 2018, 18, 114–123. [Google Scholar] [CrossRef]
- Schwalfenberg, G.K. N-Acetylcysteine: A Review of Clinical Usefulness (an Old Drug with New Tricks). J. Nutr. Metab. 2021, 2021, 9949453. [Google Scholar] [CrossRef]
- Arif, A.; Sunay, M.M.; Karakan, T.; Özcan, S.; Hasçiçek, A.M.; Yardimci, İ.; Surer, H.; Korkmaz, M.; Hücümenoğlu, S.; Huri, E. The Examination of the Nephroprotective Effect of Montelukast Sodium and N-Acetylcysteine In Renal Ischemia with Dimercaptosuccinic Acid Imaging in a Placebo-Controlled Rat Model. Acta Cir. Bras. 2020, 35, 1–9. [Google Scholar] [CrossRef]
- Kelly, A.M.; Dwamena, B.; Cronin, P.; Bernstein, S.J.; Carlos, R.C. Meta-Analysis: Effectiveness of Drugs for Preventing Contrast-Induced Nephropathy. Ann. Intern. Med. 2008, 148, 284–294. [Google Scholar] [CrossRef] [Green Version]
- Jong, L.H.; Kim, K.W. Anti-Inflammatory Effects of Arbutin in Lipopolysaccharide-Stimulated BV2 Microglial Cells. Inflamm. Res. 2012, 61, 817–825. [Google Scholar] [CrossRef]
- Shima, E.; Zabihi, E.; Golpour, M.; Aghajanpour-Mir, M. The Effect of Arbutin on the Expression of Tumor Suppressor P53, BAX/BCL-2 Ratio and Oxidative Stress Induced by Tert-Butyl Hydroperoxide in Fibroblast and LNcap Cell Lines. Cell J. 2021, 22, 532–541. [Google Scholar] [CrossRef]
- Man, X.; Yang, L.; Liu, S.; Yang, L.; Li, M.; Fu, Q. Arbutin Promotes MC3T3-E1 Mouse Osteoblast Precursor Cell Proliferation and Differentiation via the Wnt/Β-catenin Signaling Pathway. Mol. Med. Rep. 2019, 19, 4637–4644. [Google Scholar] [CrossRef] [Green Version]
- Akina, O.; Yoshimura, Y.; Deyama, Y.; Suzuki, K. Rosmarinic Acid and Arbutin Suppress Osteoclast Differentiation by Inhibiting Superoxide and NFATc1 Downregulation in RAW 264.7 Cells. Biomed. Rep. 2015, 3, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Jurica, K.; Karačonji, I.B.; Mikolić, A.; Milojković-Opsenica, D.; Benković, V.; Kopjar, N. In Vitro Safety Assessment of the Strawberry Tree (Arbutus unedo L.) Water Leaf Extract and Arbutin in Human Peripheral Blood Lymphocytes. Cytotechnology 2018, 70, 1261–1278. [Google Scholar] [CrossRef] [PubMed]
- Jurica, K.; Benković, V.; Sikirić, S.; Karačonji, I.B.; Kopjar, N. The Effects of Strawberry Tree (Arbutus unedo L.) Water Leaf Extract and Arbutin upon Kidney Function and Primary DNA Damage in Renal Cells of Rats. Nat. Prod. Res. 2020, 34, 2354–2357. [Google Scholar] [CrossRef]
- Swetha, T.K.; Pooranachithra, M.; Subramenium, G.A.; Divya, V.; Balamurugan, K.; Pandian, S.K. Umbelliferone Impedes Biofilm Formation and Virulence of Methicillin-Resistant Staphylococcus epidermidis via Impairment of Initial Attachment and Intercellular Adhesion. Front. Cell. Infect. Microbiol. 2019, 9, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.; Zhang, J.; Yu, L.; Wang, C.; Yang, Y.; Rong, X.; Xu, K.; Chu, M. Antifungal Activity of Coumarin against Candida albicans Is Related to Apoptosis. Front. Cell. Infect. Microbiol. 2019, 9, 445. [Google Scholar] [CrossRef] [Green Version]
- Hyung, L.J.; Kim, Y.G.; Cho, H.S.; Ryu, S.Y.; Cho, M.H.; Lee, J. Coumarins Reduce Biofilm Formation and the Virulence of Escherichia coli O157:H7. Phytomedicine 2014, 21, 1037–1042. [Google Scholar] [CrossRef]
- Alotaibi, M.F.; Al-Joufi, F.; Seif, H.S.A.; Alzoghaibi, M.A.; Djouhri, L.; Ahmeda, A.F.; Mahmoud, A.M. Umbelliferone Inhibits Spermatogenic Defects and Testicular Injury in Lead-Intoxicated Rats by Suppressing Oxidative Stress and Inflammation, and Improving Nrf2/HO-1 Signaling. Drug Des. Dev. Ther. 2020, 14, 4003–4019. [Google Scholar] [CrossRef]
- Hassanein, E.H.M.; Fares, E.M.A.; Kozman, M.R.; El-Ghafar, O.A.M.A. Umbelliferone Attenuates Gentamicin-Induced Renal Toxicity by Suppression of TLR-4/NF-ΚB-P65/NLRP-3 and JAK1/STAT-3 Signaling Pathways. Environ. Sci. Pollut. Res. 2021, 28, 11558–11571. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; Jin, B.-R.; An, H.-J. Umbelliferone Ameliorates Benign Prostatic Hyperplasia by Inhibiting Cell Proliferation and G1/S Phase Cell Cycle Progression through Regulation of STAT3/E2F1 Axis. Int. J. Mol. Sci. 2021, 22, 9019. [Google Scholar] [CrossRef]
- Vijayalakshmi, A.; Sindhu, G. Umbelliferone Arrest Cell Cycle at G0/G1 Phase and Induces Apoptosis in Human Oral Carcinoma (KB) Cells Possibly via Oxidative DNA Damage. Biomed. Pharmacother. 2017, 92, 661–671. [Google Scholar] [CrossRef]
- Lopez-Gonzalez, J.S.; Prado-Garcia, H.; Aguilar-Cazares, D.; Molina-Guarneros, J.A.; Morales-Fuentes, J.; Mandoki, J.J. Apoptosis and Cell Cycle Disturbances Induced by Coumarin and 7-Hydroxycoumarin on Human Lung Carcinoma Cell Lines. Lung Cancer 2004, 43, 275–283. [Google Scholar] [CrossRef]
- Manoharan, A.; Ognenovska, S.; Paino, D.; Whiteley, G.; Glasbey, T.; Kriel, F.H.; Farrell, J.; Moore, K.H.; Manos, J.; Das, T. N-Acetylcysteine Protects Bladder Epithelial Cells from Bacterial Invasion and Displays Antibiofilm Activity against Urinary Tract Bacterial Pathogens. Antibiotics 2021, 10, 900. [Google Scholar] [CrossRef] [PubMed]
- Irina, G.; Efremova, T.; Kirpichnikova, K.; Kever, L.; Komissarchik, Y.; Polozov, Y.; Khaitlina, S. N-Acetylcysteine-Induced Changes in Susceptibility of Transformed Eukaryotic Cells to Bacterial Invasion. Cell Biol. Int. 2006, 30, 319–325. [Google Scholar] [CrossRef]
- Makipour, K.; Friedenberg, F.K. The Potential Role of N-Acetylcysteine for the Treatment of Helicobacter pylori. J. Clin. Gastroenterol. 2011, 45, 841–843. [Google Scholar] [CrossRef] [Green Version]
- Zhao, T.; Liu, Y. N-Acetylcysteine Inhibit Biofilms Produced by Pseudomonas aeruginosa. BMC Microbiol. 2010, 10, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arthika, M.; Das, T.; Whiteley, G.S.; Glasbey, T.; Kriel, F.H.; Manos, J. The Effect of N-Acetylcysteine in a Combined Antibiofilm Treatment against Antibiotic-Resistant Staphylococcus aureus. J. Antimicrob. Chemother. 2020, 75, 1787–1798. [Google Scholar] [CrossRef]
- Kundukad, B.; Schussman, M.; Yang, K.; Seviour, T.; Yang, L.; Rice, S.A.; Kjelleberg, S.; Doyle, P.S. Mechanistic Action of Weak Acid Drugs on Biofilms. Sci. Rep. 2017, 7, 4783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theerthankar, D.; Kutty, S.K.; Tavallaie, R.; Ibugo, A.I.; Panchompoo, J.; Sehar, S.; Aldous, L.; Yeung, A.W.S.; Thomas, S.R.; Kumar, N.; et al. Phenazine Virulence Factor Binding to Extracellular DNA Is Important for Pseudomonas aeruginosa Biofilm Formation. Sci. Rep. 2015, 5, 8398. [Google Scholar] [CrossRef]
- Ryu, C.M.; Shin, J.H.; Yu, H.Y.; Ju, H.; Kim, S.; Lim, J.; Heo, J.; Lee, S.; Shin, D.M.; Choo, M.S. N-Acetylcysteine Prevents Bladder Tissue Fibrosis in a Lipopolysaccharide-Induced Cystitis Rat Model. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Binu, K.; Udayakumar, G.; Grela, E.; Kaur, D.; Rice, S.A.; Kjelleberg, S.; Doyle, P.S. Weak Acids as an Alternative Anti-Microbial Therapy. Biofilm 2020, 2, 100019. [Google Scholar] [CrossRef]
| Antibiotic | Mechanism of Action | Reference |
|---|---|---|
| Nitrofurantoin | Destroys bacterial RNA and DNA | [23] |
| Fosfomycin | Inhibits Gram positive and negative cell wall synthesis | [23] |
| Ciprofloxacin | A fluoroquinolone used against Gram negative bacteria that impairs DNA’s bacterial synthesis and inhibits topoisomorases’ actions | [24] |
| Trimethoprim | Inhibits bacterial folic acid synthesis | [23] |
| Levofloxacin | Inhibits topoisomerase IV and bacterial gyrase | [25] |
| Cephalexin | Beta lactam that inhibits cell wall synthesis | [26] |
| Cefpodoxime | Cephalosporin that inhibits cell wall synthesis | [27] |
| Ceftibuten | Beta lactam that inhibits cell wall synthesis | [27] |
| Piperacillin | Beta lactam that inhibits cell wall synthesis | [27] |
| Action/Finding | References |
|---|---|
| Reduces iNOS expression in B2 microglia cells and IL-1β, TNF-α, MCP-1, and IL-6 | [53] |
| Reduces oxidative stress levels in fibroblast and increases apoptosis of tumor cell line | [54] |
| Increases expression of collagen I | [55] |
| Decreases osteoclast activity | [56] |
| No DNA damage in lymphocytes | [57,58] |
| Action/Finding | References |
|---|---|
| Impairs adhesion of bacteria | [68] |
| Inhibits biofilm formation | [69,70,71] |
| DNA bacteria and biofilm components are derived from DNA destruction | [72,75] |
| Inhibits expression of genes related to fibrotic process | [74] |
| Reduces expression of genes related to apoptosis | [23] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cela-López, J.M.; Camacho Roldán, C.J.; Gómez-Lizarraga, G.; Martínez, V. Effects of Itxasol© Components on Gene Expression in Bacteria Related to Infections of the Urinary Tract and to the Inflammation Process. Int. J. Mol. Sci. 2021, 22, 12655. https://doi.org/10.3390/ijms222312655
Cela-López JM, Camacho Roldán CJ, Gómez-Lizarraga G, Martínez V. Effects of Itxasol© Components on Gene Expression in Bacteria Related to Infections of the Urinary Tract and to the Inflammation Process. International Journal of Molecular Sciences. 2021; 22(23):12655. https://doi.org/10.3390/ijms222312655
Chicago/Turabian StyleCela-López, José M., Claudio J. Camacho Roldán, Gorka Gómez-Lizarraga, and Vicente Martínez. 2021. "Effects of Itxasol© Components on Gene Expression in Bacteria Related to Infections of the Urinary Tract and to the Inflammation Process" International Journal of Molecular Sciences 22, no. 23: 12655. https://doi.org/10.3390/ijms222312655
APA StyleCela-López, J. M., Camacho Roldán, C. J., Gómez-Lizarraga, G., & Martínez, V. (2021). Effects of Itxasol© Components on Gene Expression in Bacteria Related to Infections of the Urinary Tract and to the Inflammation Process. International Journal of Molecular Sciences, 22(23), 12655. https://doi.org/10.3390/ijms222312655