
Reduction of Oxidative Stress in Peripheral Blood Mononuclear Cells Attenuates the Inflammatory Response of Fibroblast-like Synoviocytes in Rheumatoid Arthritis


Abstract
1. Introduction
2. Results
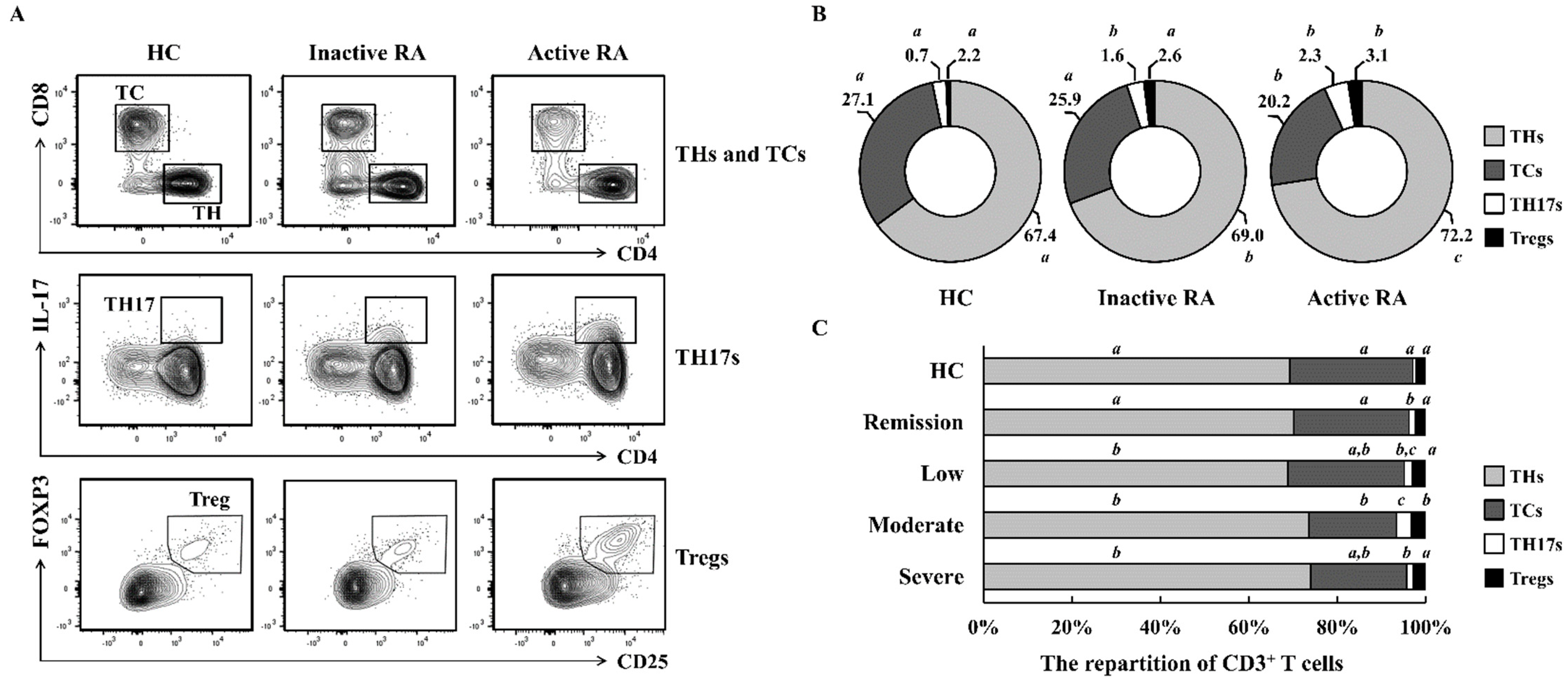
2.1. The Repartition of CD3+ T Cells Differs According to RA Severity
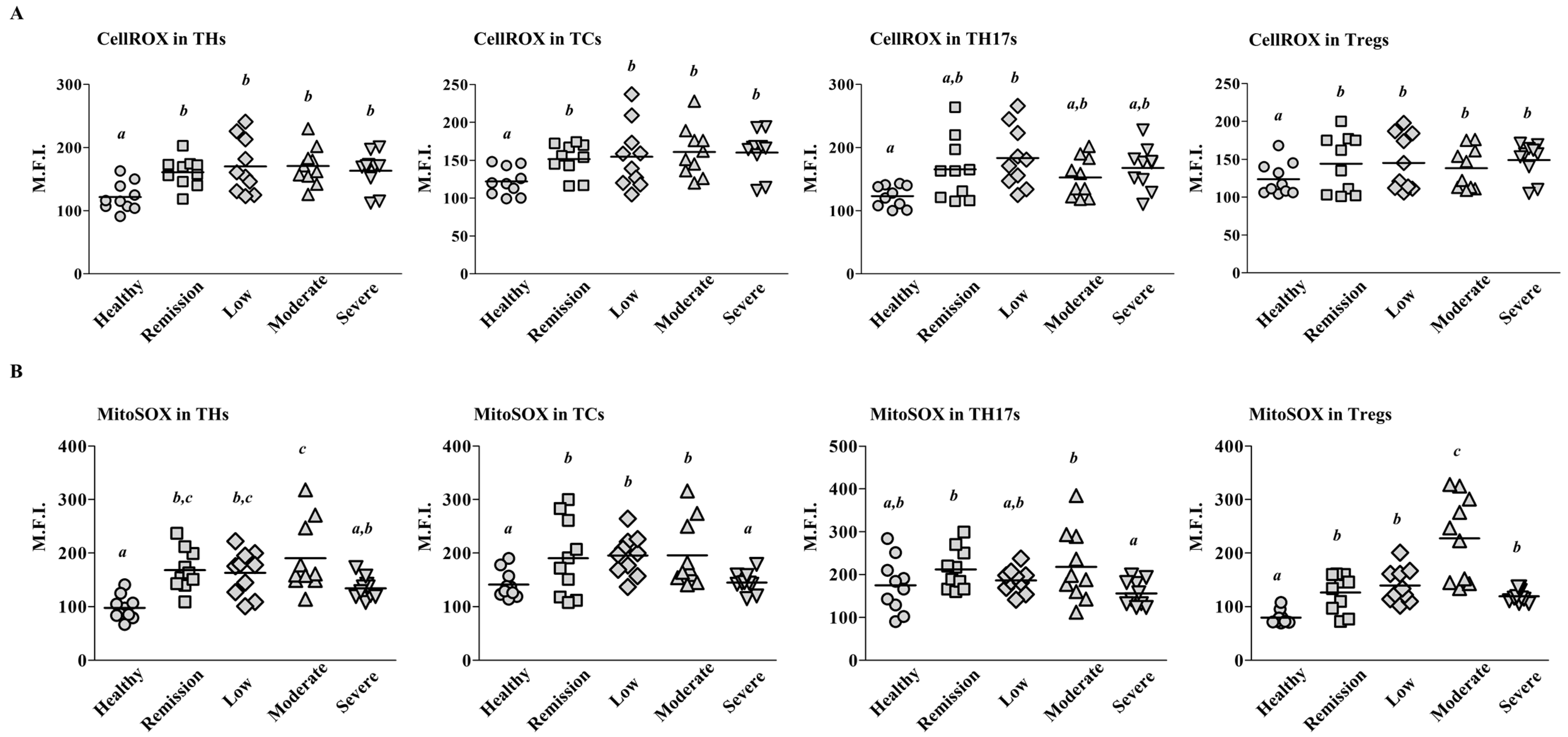
2.2. Mitochondrial ROS Levels Differ among T Cell Subsets According to RA Severity
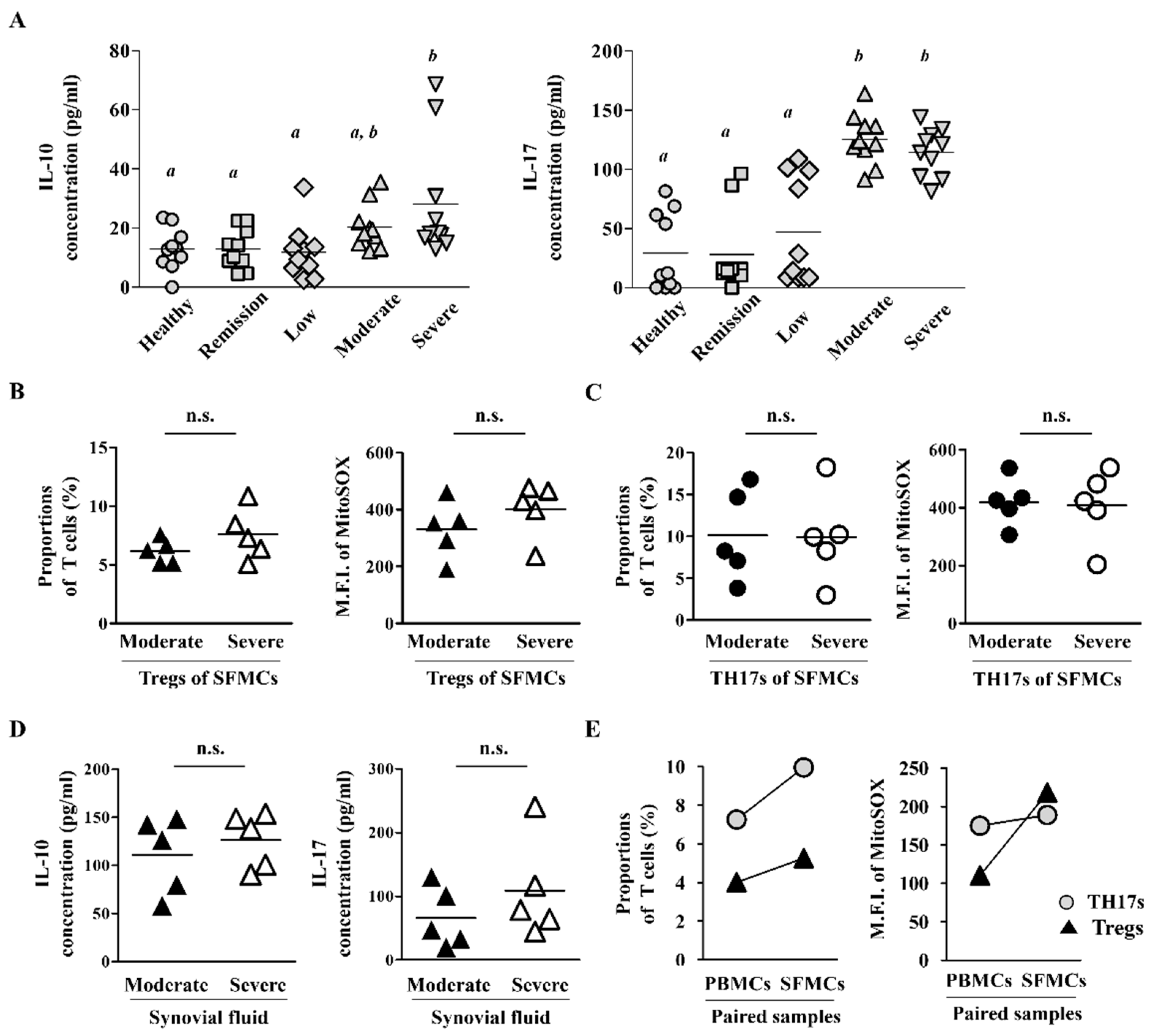
2.3. Synovial Fluid-Derived Mononuclear Cells (SFMCs) Have Higher ROS Levels and T Cell Population Than PBMCs
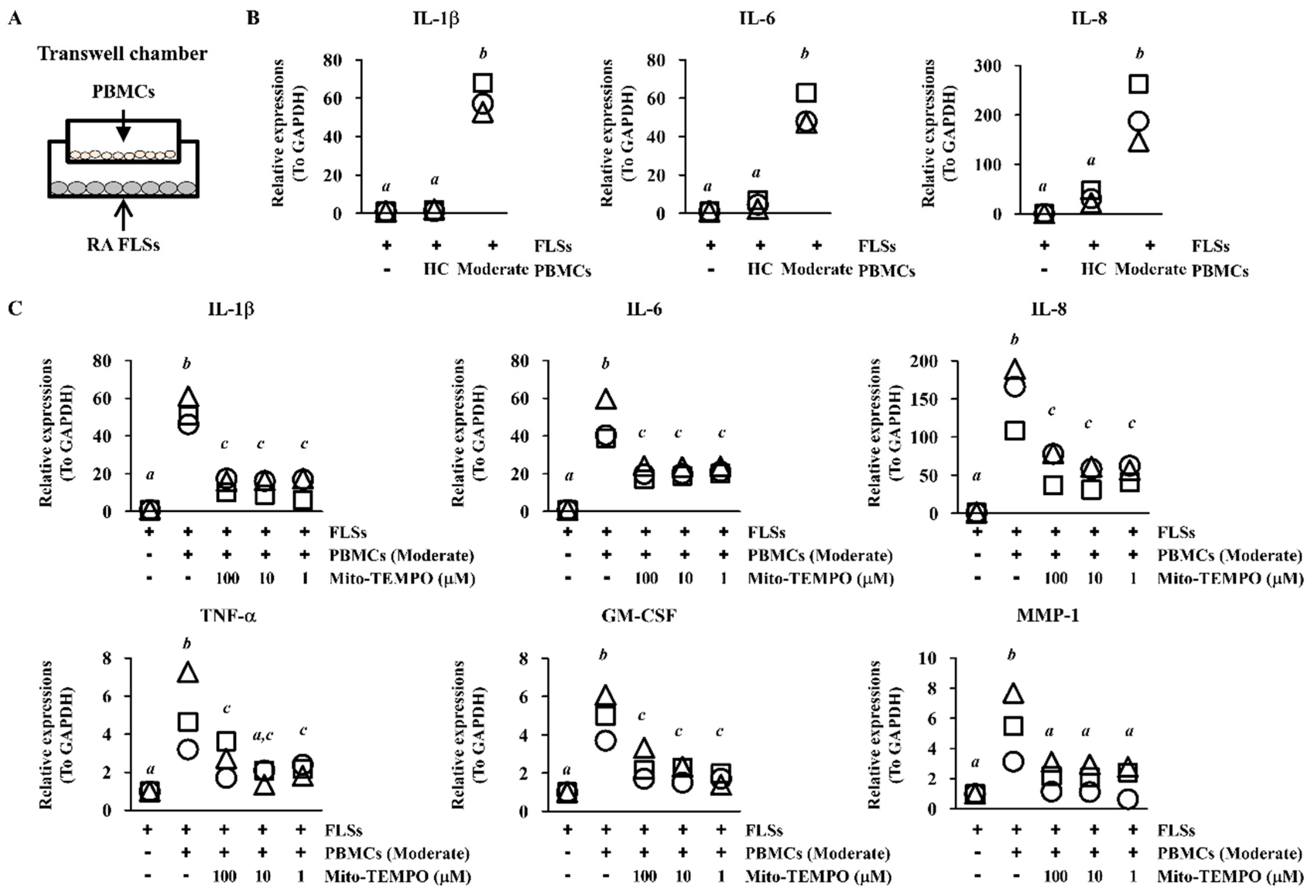
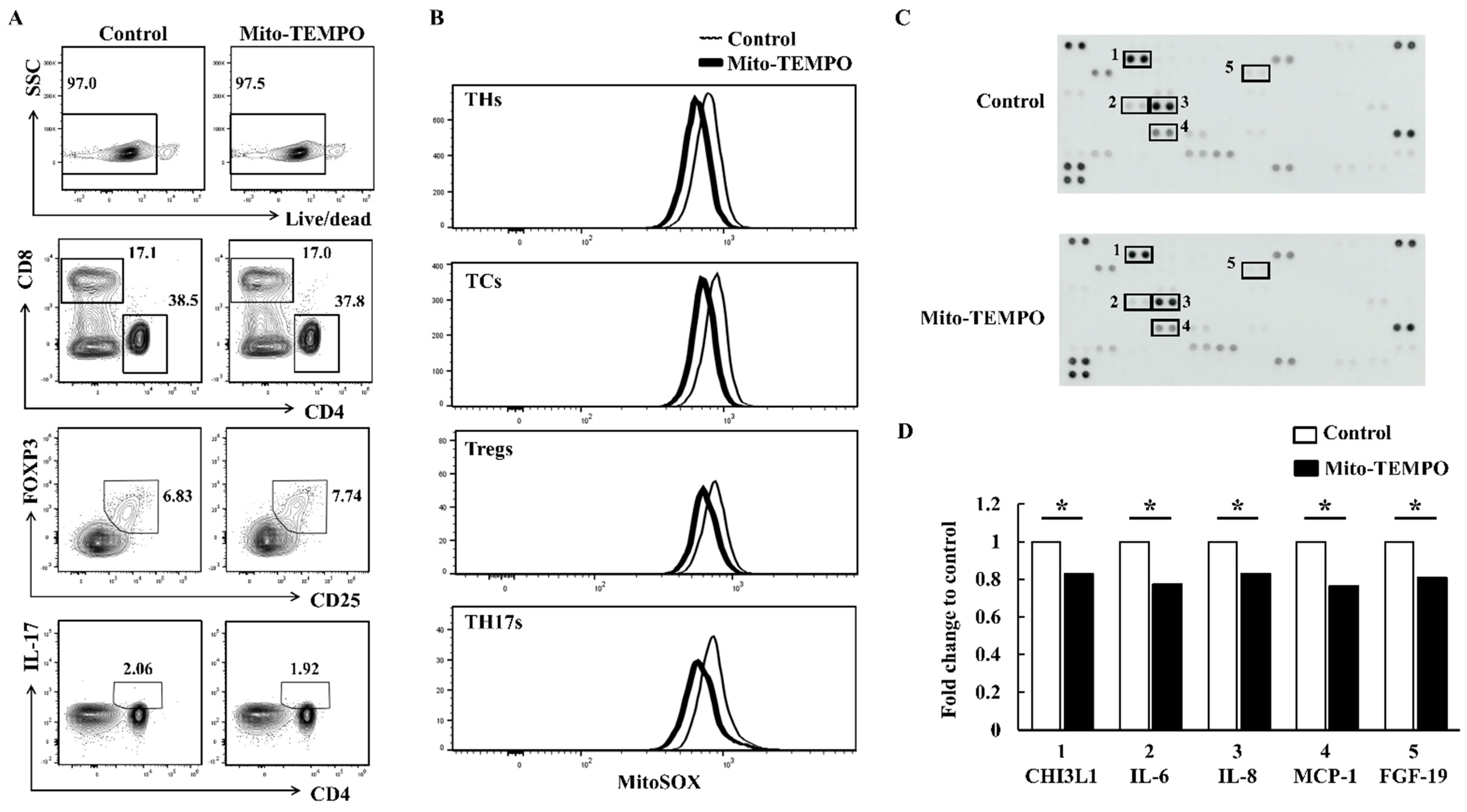
2.4. Inflammatory Response of FLS Is Suppressed via ROS Inhibition in PBMCs
3. Discussion
4. Materials and Methods
4.1. Human Subjects
4.2. Ethics Statement
4.3. Isolation of PBMCs and SFMCs
4.4. Flow Cytometry Analysis
4.5. Enzyme-Linked Immunosorbent Assay (ELISA)
4.6. Co-Culture Using Transwell Inserts and Cytokine Array
4.7. Real-Time PCR
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Firestein, G.S. Evolving concepts of rheumatoid arthritis. Nature 2003, 423, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Mellado, M.; Martinez-Munoz, L.; Cascio, G.; Lucas, P.; Pablos, J.L.; Rodriguez-Frade, J.M. T Cell Migration in Rheumatoid Arthritis. Front. Immunol. 2015, 6, 384. [Google Scholar] [CrossRef] [PubMed]
- McInnes, I.B.; Schett, G. Cytokines in the pathogenesis of rheumatoid arthritis. Nat. Rev. Immunol. 2007, 7, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Szekanecz, Z.; Besenyei, T.; Paragh, G.; Koch, A.E. New insights in synovial angiogenesis. Jt. Bone Spine 2010, 77, 13–19. [Google Scholar] [CrossRef][Green Version]
- Sakaguchi, S. Naturally arising Foxp3-expressing CD25+CD4+ regulatory T cells in immunological tolerance to self and non-self. Nat. Immunol. 2005, 6, 345–352. [Google Scholar] [CrossRef]
- Gol-Ara, M.; Jadidi-Niaragh, F.; Sadria, R.; Azizi, G.; Mirshafiey, A. The role of different subsets of regulatory T cells in immunopathogenesis of rheumatoid arthritis. Arthritis 2012, 2012, 805875. [Google Scholar] [CrossRef]
- Bayry, J.; Siberil, S.; Triebel, F.; Tough, D.F.; Kaveri, S.V. Rescuing CD4+CD25+ regulatory T-cell functions in rheumatoid arthritis by cytokine-targeted monoclonal antibody therapy. Drug Discov. Today 2007, 12, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Han, G.M.; O’Neil-Andersen, N.J.; Zurier, R.B.; Lawrence, D.A. CD4+CD25high T cell numbers are enriched in the peripheral blood of patients with rheumatoid arthritis. Cell Immunol. 2008, 253, 92–101. [Google Scholar] [CrossRef]
- van Amelsfort, J.M.; van Roon, J.A.; Noordegraaf, M.; Jacobs, K.M.; Bijlsma, J.W.; Lafeber, F.P.; Taams, L.S. Proinflammatory mediator-induced reversal of CD4+, CD25+ regulatory T cell-mediated suppression in rheumatoid arthritis. Arthritis Rheum. 2007, 56, 732–742. [Google Scholar] [CrossRef]
- Sakaguchi, S. Regulatory T cells in the past and for the future. Eur. J. Immunol. 2008, 38, 901–937. [Google Scholar] [CrossRef]
- Wehrens, E.J.; Prakken, B.J.; van Wijk, F. T cells out of control--impaired immune regulation in the inflamed joint. Nat. Rev. Rheumatol. 2013, 9, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Nie, H.; Zheng, Y.; Li, R.; Guo, T.B.; He, D.; Fang, L.; Liu, X.; Xiao, L.; Chen, X.; Wan, B.; et al. Phosphorylation of FOXP3 controls regulatory T cell function and is inhibited by TNF-alpha in rheumatoid arthritis. Nat. Med. 2013, 19, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Yoshitomi, H. Regulation of Immune Responses and Chronic Inflammation by Fibroblast-Like Synoviocytes. Front. Immunol. 2019, 10, 1395. [Google Scholar] [CrossRef]
- Veselinovic, M.; Barudzic, N.; Vuletic, M.; Zivkovic, V.; Tomic-Lucic, A.; Djuric, D.; Jakovljevic, V. Oxidative stress in rheumatoid arthritis patients: Relationship to diseases activity. Mol. Cell Biochem. 2014, 391, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Mateen, S.; Moin, S.; Khan, A.Q.; Zafar, A.; Fatima, N. Increased Reactive Oxygen Species Formation and Oxidative Stress in Rheumatoid Arthritis. PLoS ONE 2016, 11, e0152925. [Google Scholar] [CrossRef]
- Okamoto, T.; Sakurada, S.; Yang, J.P.; Merin, J.P. Regulation of NF-kappa B and disease control: Identification of a novel serine kinase and thioredoxin as effectors for signal transduction pathway for NF-kappa B activation. Curr. Top. Cell Regul. 1997, 35, 149–161. [Google Scholar]
- Moynagh, P.N. The NF-kappaB pathway. J. Cell Sci. 2005, 118, 4589–4592. [Google Scholar] [CrossRef]
- Mateen, S.; Moin, S.; Shahzad, S.; Khan, A.Q. Level of inflammatory cytokines in rheumatoid arthritis patients: Correlation with 25-hydroxy vitamin D and reactive oxygen species. PLoS ONE 2017, 12, e0178879. [Google Scholar] [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef]
- Devadas, S.; Zaritskaya, L.; Rhee, S.G.; Oberley, L.; Williams, M.S. Discrete generation of superoxide and hydrogen peroxide by T cell receptor stimulation: Selective regulation of mitogen-activated protein kinase activation and fas ligand expression. J. Exp. Med. 2002, 195, 59–70. [Google Scholar] [CrossRef]
- Weyand, C.M.; Shen, Y.; Goronzy, J.J. Redox-sensitive signaling in inflammatory T cells and in autoimmune disease. Free Radic. Biol. Med. 2018, 125, 36–43. [Google Scholar] [CrossRef]
- Okamoto, H.; Hoshi, D.; Kiire, A.; Yamanaka, H.; Kamatani, N. Molecular targets of rheumatoid arthritis. Inflamm. Allergy Drug Targets 2008, 7, 53–66. [Google Scholar] [CrossRef]
- Miyara, M.; Gorochov, G.; Ehrenstein, M.; Musset, L.; Sakaguchi, S.; Amoura, Z. Human FoxP3+ regulatory T cells in systemic autoimmune diseases. Autoimmun. Rev. 2011, 10, 744–755. [Google Scholar] [CrossRef] [PubMed]
- Kosmaczewska, A.; Swierkot, J.; Ciszak, L.; Szteblich, A.; Chrobak, A.; Karabon, L.; Partyka, A.; Szechinski, J.; Wiland, P.; Frydecka, I. Patients with the most advanced rheumatoid arthritis remain with Th1 systemic defects after TNF inhibitors treatment despite clinical improvement. Rheumatol. Int. 2014, 34, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, J.; Gao, H.; Wang, C.; Luo, J.; Lv, Z.; Li, X. Comprehensive evaluation of different T-helper cell subsets differentiation and function in rheumatoid arthritis. J. Biomed. Biotechnol. 2012, 2012, 535361. [Google Scholar] [CrossRef]
- Kao, J.K.; Hsue, Y.T.; Lin, C.Y. Role of new population of peripheral CD11c(+)CD8(+) T cells and CD4(+)CD25(+) regulatory T cells during acute and remission stages in rheumatoid arthritis patients. J. Microbiol. Immunol. Infect. 2007, 40, 419–427. [Google Scholar]
- van Amelsfort, J.M.; Jacobs, K.M.; Bijlsma, J.W.; Lafeber, F.P.; Taams, L.S. CD4(+)CD25(+) regulatory T cells in rheumatoid arthritis: Differences in the presence, phenotype, and function between peripheral blood and synovial fluid. Arthritis Rheum. 2004, 50, 2775–2785. [Google Scholar] [CrossRef]
- Miyara, M.; Yoshioka, Y.; Kitoh, A.; Shima, T.; Wing, K.; Niwa, A.; Parizot, C.; Taflin, C.; Heike, T.; Valeyre, D.; et al. Functional delineation and differentiation dynamics of human CD4+ T cells expressing the FoxP3 transcription factor. Immunity 2009, 30, 899–911. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Yang, G.; Liu, Q.; Wang, S.; Cui, D. Function and Role of Regulatory T Cells in Rheumatoid Arthritis. Front. Immunol 2021, 12, 626193. [Google Scholar] [CrossRef]
- Pan, X.; Yuan, X.; Zheng, Y.; Wang, W.; Shan, J.; Lin, F.; Jiang, G.; Yang, Y.H.; Wang, D.; Xu, D.; et al. Increased CD45RA+ FoxP3(low) regulatory T cells with impaired suppressive function in patients with systemic lupus erythematosus. PLoS ONE 2012, 7, e34662. [Google Scholar]
- Szalay, B.; Vasarhelyi, B.; Cseh, A.; Tulassay, T.; Deak, M.; Kovacs, L.; Balog, A. The impact of conventional DMARD and biological therapies on CD4+ cell subsets in rheumatoid arthritis: A follow-up study. Clin. Rheumatol. 2014, 33, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Fearon, U.; Canavan, M.; Biniecka, M.; Veale, D.J. Hypoxia, mitochondrial dysfunction and synovial invasiveness in rheumatoid arthritis. Nat. Rev. Rheumatol. 2016, 12, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Kaarniranta, K.; Pawlowska, E.; Szczepanska, J.; Jablkowska, A.; Blasiak, J. Role of Mitochondrial DNA Damage in ROS-Mediated Pathogenesis of Age-Related Macular Degeneration (AMD). Int. J. Mol. Sci. 2019, 20, 2374. [Google Scholar] [CrossRef]
- Guo, Z.; Wang, G.; Wu, B.; Chou, W.C.; Cheng, L.; Zhou, C.; Lou, J.; Wu, D.; Su, L.; Zheng, J.; et al. DCAF1 regulates Treg senescence via the ROS axis during immunological aging. J. Clin. Investig. 2020, 130, 5893–5908. [Google Scholar] [CrossRef]
- Fessler, J.; Husic, R.; Schwetz, V.; Lerchbaum, E.; Aberer, F.; Fasching, P.; Ficjan, A.; Obermayer-Pietsch, B.; Duftner, C.; Graninger, W.; et al. Senescent T-Cells Promote Bone Loss in Rheumatoid Arthritis. Front. Immunol. 2018, 9, 95. [Google Scholar] [CrossRef] [PubMed]
- Fessler, J.; Raicht, A.; Husic, R.; Ficjan, A.; Schwarz, C.; Duftner, C.; Schwinger, W.; Graninger, W.B.; Stradner, M.H.; Dejaco, C. Novel Senescent Regulatory T-Cell Subset with Impaired Suppressive Function in Rheumatoid Arthritis. Front. Immunol. 2017, 8, 300. [Google Scholar] [CrossRef]
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham, C.O., 3rd; Birnbaum, N.S.; Burmester, G.R.; Bykerk, V.P.; Cohen, M.D.; et al. 2010 Rheumatoid arthritis classification criteria: An American College of Rheumatology/European League Against Rheumatism collaborative initiative. Arthritis Rheum. 2010, 62, 2569–2581. [Google Scholar] [CrossRef]
- Prevoo, M.L.; van ’t Hof, M.A.; Kuper, H.H.; van Leeuwen, M.A.; van de Putte, L.B.; van Riel, P.L. Modified disease activity scores that include twenty-eight-joint counts. Development and validation in a prospective longitudinal study of patients with rheumatoid arthritis. Arthritis Rheum. 1995, 38, 44–48. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Active RA (DAS28 ≥ 3.2) | Inactive RA (DAS28 < 3.2) | Health (n = 10) | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| Characteristics | Severe (n = 10) | Moderate (n = 10) | Low (n = 10) | Remission (n = 10) | ||||
| Female, n (%) | 7 (70) | 7 (70) | 8 (80) | 7 (70) | 7 (70) | 0.983 | ||
| Age, years | 56.9 ± 7.5 (44–68) | 55.2 ± 14.0 (30–72) | 59.4 ± 12.8 (35–80) | 53.0 ± 12.8 (28–69) | 56.1 ± 12.0 (35–70) | 0.823 | ||
| Onset age, years | 55.8 ± 6.4 (44–64) | 51.2 ± 14.3 (21–68) | 53.5 ± 15.2 (24–67) | 47.0 ± 14.3 (28–69) | 0.484 | |||
| From onset time to treatment time, months | 14.4 ± 29.1 | 13.6 ± 18.86 | 71.2 ± 76.82 | 33.5 ± 46.89 | 2,3:0.023 | |||
| Duration of disease, months | 27.8 ± 42.9 (2–120) | 65.0 ± 56.5 (1.5–151) | 133.4 ± 87.8 (18–276) | 98.4 ± 70.0 (14–228) | 1,4:0.003 1,3:<0.001 2,3:0.035 | |||
| Rheumatoid factor–positive, n (%) | 9 (90) | 9 (90) | 8/9 (89) | 7/9 (78) | 0.603 | |||
| Rheumatoid factor titer (IU/mL) | 94.1 ± 69.4 | 106.2 ± 81.0 | 145.2 ± 119.6 | 92.6 ± 88.4 | 0.743 | |||
| Anti CCP antibody–positive, n (%) | 10 (100) | 6/7 (86) | 5/5 (100) | 5/6 (83) | 0.489 | |||
| Anti CCP antibody titer (U/mL) | 367.2 ± 174.8 | 172.6 ± 181.2 | 500 ± 0 | 189.3 ± 243.7 | 1,2:0.019 2,3:0.003 | |||
| DAS28 (ESR) | 6.3 ± 0.8 | 4.2 ± 0.5 | 2.8 ± 0.3 | 1.4 ± 0.6 | <0.001 | |||
| SJC | 11.2 ± 7.5 | 1.6 ± 1.6 | 0.1 ± 0.3 | 0.1 ± 0.3 | 1,2 <0.001 1,3 <0.001 1,4 <0.001 2,3:0.004 2,4:0.004 | |||
| TJC | 11.5 ± 7.1 | 1.8 ± 1.6 | 0.4 ± 0.7 | 0.1 ± 0.3 | 1,2 <0.001 1,3 <0.001 1,4 <0.001 2,4 <0.001 | |||
| VAS | 73 ± 15.0 | 41 ± 15.2 | 16.5 ± 9.4 | 9.8 ± 4.1 | 1,2:0.001 1,3 <0.001 1,4 <0.001 2,3 <0.001 2,4 <0.001 | |||
| ESR | 52.4 ± 38.3 | 49.5 ± 25.9 | 29.2 ± 17.5 | 8.6 ± 9.2 | 1,4 <0.001 2,3:0.011 2,4 <0.001 3,4:0.011 | |||
| CRP | 2.1 ± 2.3 | 1.9 ± 2.2 | 0.5 ± 0.8 | 0.06 ± 0.1 | 1,4:0.015 2,4 <0.001 | |||
| Duration of treatment, months | 2.2 ± 3.4 (0–8) | 44.5 ± 54.8 (0.5–139) | 79.2 ± 71.2 (6–204) | 65.5 ± 59.7 (1.5–187) | 1,4:0.001 1,3:<0.001 1,2:0.005 2,3:0.023 | |||
| Treatment | Naïve, n (%) | 5 (50) | 2 (20) | 0 | 0 | 0.280 | ||
| Steroid, n (%) | 5 (50) | 7 (70) | 9 (90) | 8 (80) | 0.539 | |||
| Prednisolone dose (mg/day) | 6.5 ± 1.3 | 5.5 ± 2.6 | 2.6 ± 1.9 | 4.1 ± 3.7 | ||||
| DMARD, biologic, n (%) | 0 | 0 | 0 | 2 (20) | ||||
| DMARD, conventional, n (%) | total | 5 (50) | 8 (80) | 9 (90) | 7 (70) | |||
| Methotrexate, n (%) | 5 (50) | 8 (80) | 9 (90) | 7 (70) | ||||
| Methotrexate dose (mg /week) | 11.3 ± 3.6 | 12.8 ± 2.8 | 10.8 ± 4.9 | 9.5 ± 6.7 | ||||
| Sulfasalazine, n (%) | 3 (30) | 2 (20) | 3 (30) | 3 (30) | ||||
| Hydroxychloroquine, n (%) | 2 (20) | 6 (60) | 4 (40) | 3 (30) | ||||
| Leflunomide, n (%) | 0 | 0 | 3 (30) | 4 (40) | ||||
| Sense Primer | Antisense Primer | |
|---|---|---|
| IL-1β | GGATATGGAGCAACAAGTGG | ATGTACCAGTTGGGGAACTG |
| IL-6 | AACCTGAACCTTCCAAAGATGG | TCTGGCTTGTTCCTCACTACT |
| IL-8 | CATACTCCAAACCTTTCCACCCC | TCAGCCCTCTTCAAAAACTTCTCCA |
| TNF-α | CCCGAGTGACAAGCCTGTAG | GATGGCAGAGAGGAGGTTGAC |
| GM-CSF | TCTCAGAAATGTTTGACCTCCA | GCCCTTGAGCTTGGTGAG |
| MMP-1 | GGCCCACAAACCCCAAAAG | ATCTCTGTCGGCAAATTCGTAAGC |
| GAPDH | CACATGGCCTCCAAGGAGTAA | TGAGGGTCTCTCTCTTCCTCTTGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-R.; Yoo, S.-J.; Kim, J.; Park, C.K.; Kang, S.W. Reduction of Oxidative Stress in Peripheral Blood Mononuclear Cells Attenuates the Inflammatory Response of Fibroblast-like Synoviocytes in Rheumatoid Arthritis. Int. J. Mol. Sci. 2021, 22, 12411. https://doi.org/10.3390/ijms222212411
Lee H-R, Yoo S-J, Kim J, Park CK, Kang SW. Reduction of Oxidative Stress in Peripheral Blood Mononuclear Cells Attenuates the Inflammatory Response of Fibroblast-like Synoviocytes in Rheumatoid Arthritis. International Journal of Molecular Sciences. 2021; 22(22):12411. https://doi.org/10.3390/ijms222212411
Chicago/Turabian StyleLee, Ha-Reum, Su-Jin Yoo, Jinhyun Kim, Chan Keol Park, and Seong Wook Kang. 2021. "Reduction of Oxidative Stress in Peripheral Blood Mononuclear Cells Attenuates the Inflammatory Response of Fibroblast-like Synoviocytes in Rheumatoid Arthritis" International Journal of Molecular Sciences 22, no. 22: 12411. https://doi.org/10.3390/ijms222212411
APA StyleLee, H.-R., Yoo, S.-J., Kim, J., Park, C. K., & Kang, S. W. (2021). Reduction of Oxidative Stress in Peripheral Blood Mononuclear Cells Attenuates the Inflammatory Response of Fibroblast-like Synoviocytes in Rheumatoid Arthritis. International Journal of Molecular Sciences, 22(22), 12411. https://doi.org/10.3390/ijms222212411