
IFN-Gamma and TNF-Alpha as a Priming Strategy to Enhance the Immunomodulatory Capacity of Secretomes from Menstrual Blood-Derived Stromal Cells

 ,
,  , , , , ,
, , , , ,  , and
, and 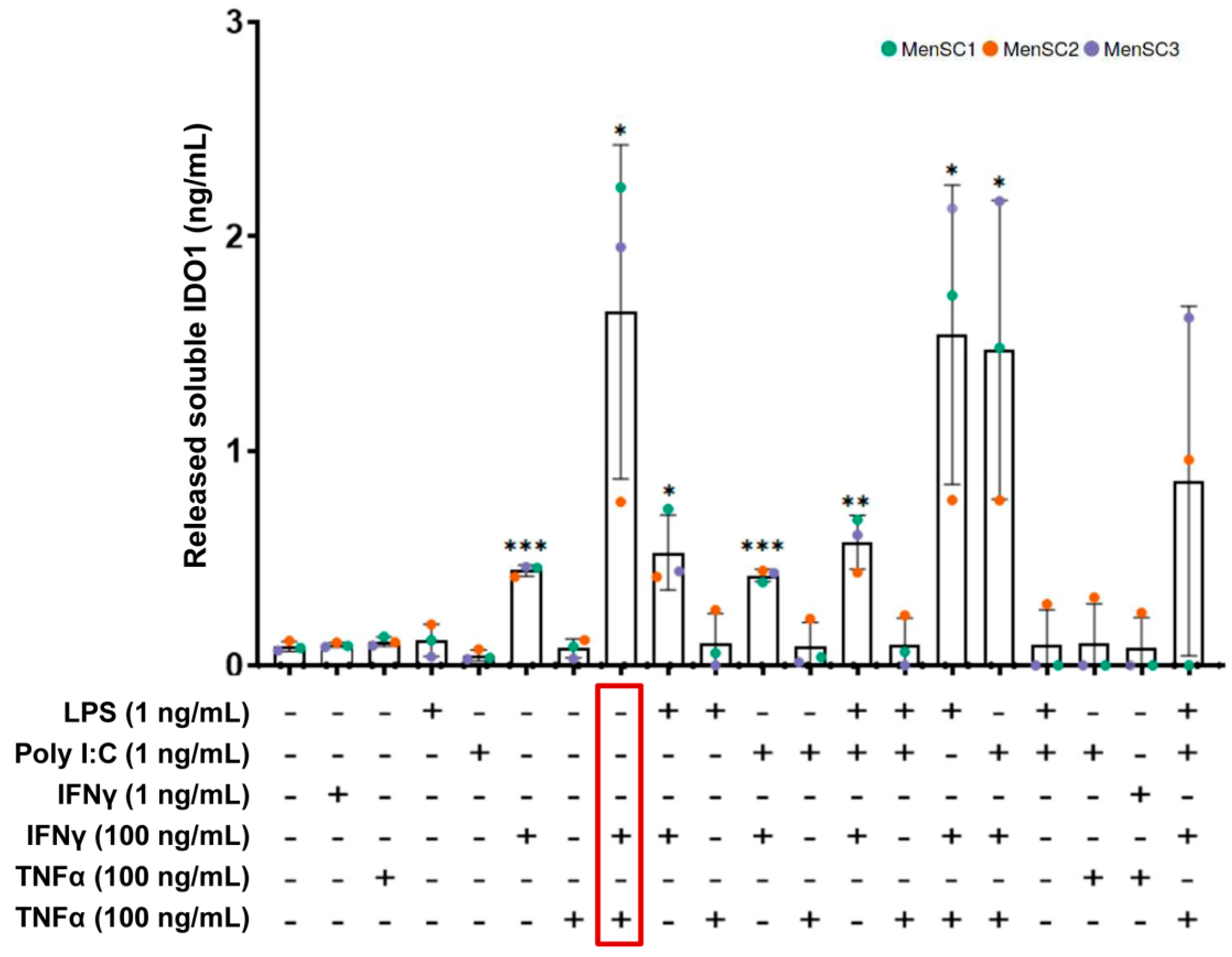{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Characterization of In Vitro Isolated and Expanded MenSCs
2.2. Effect of MenSC Priming on IDO1 Release
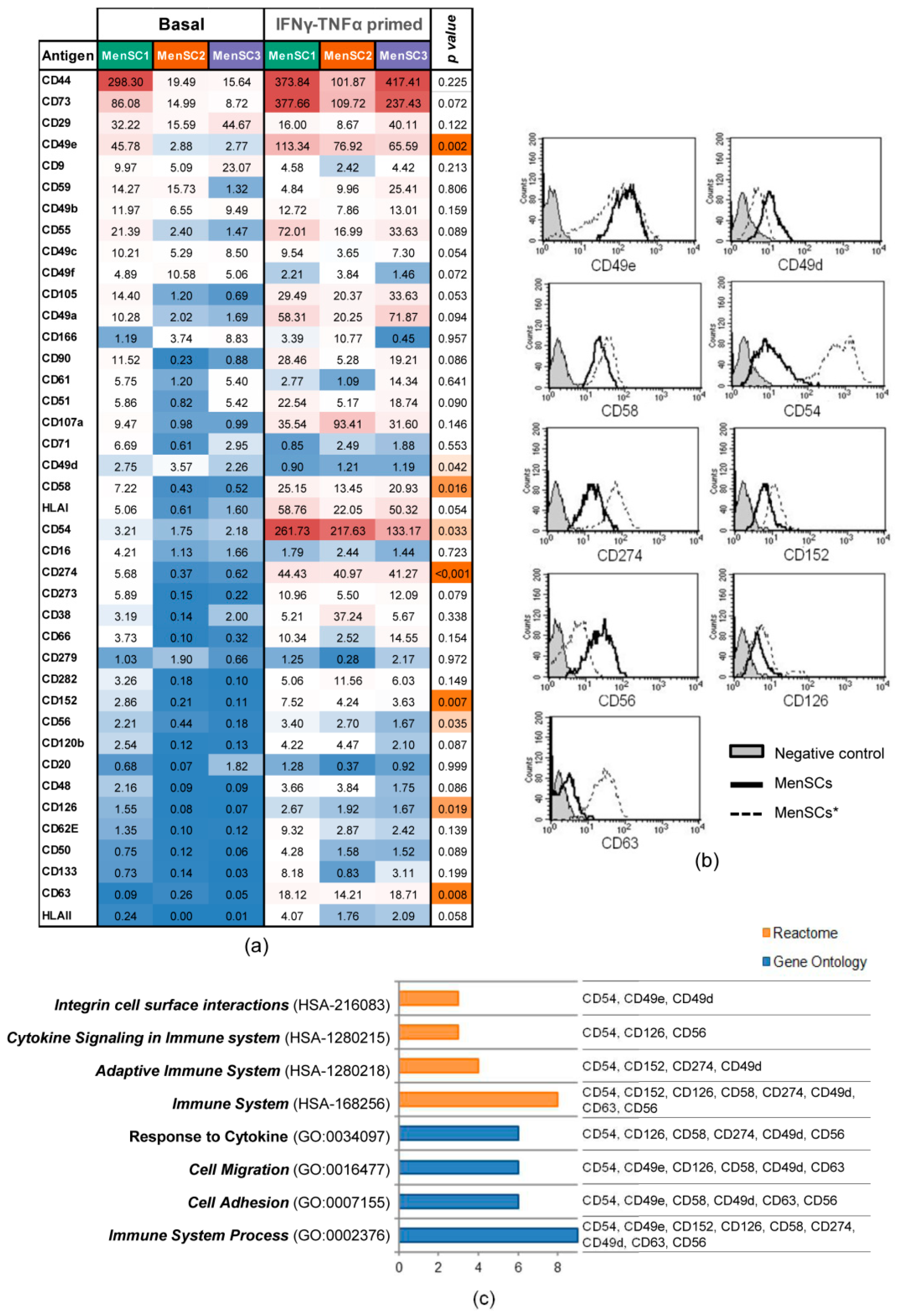
2.3. Phenotypic Profile of Surface Markers on MenSCs
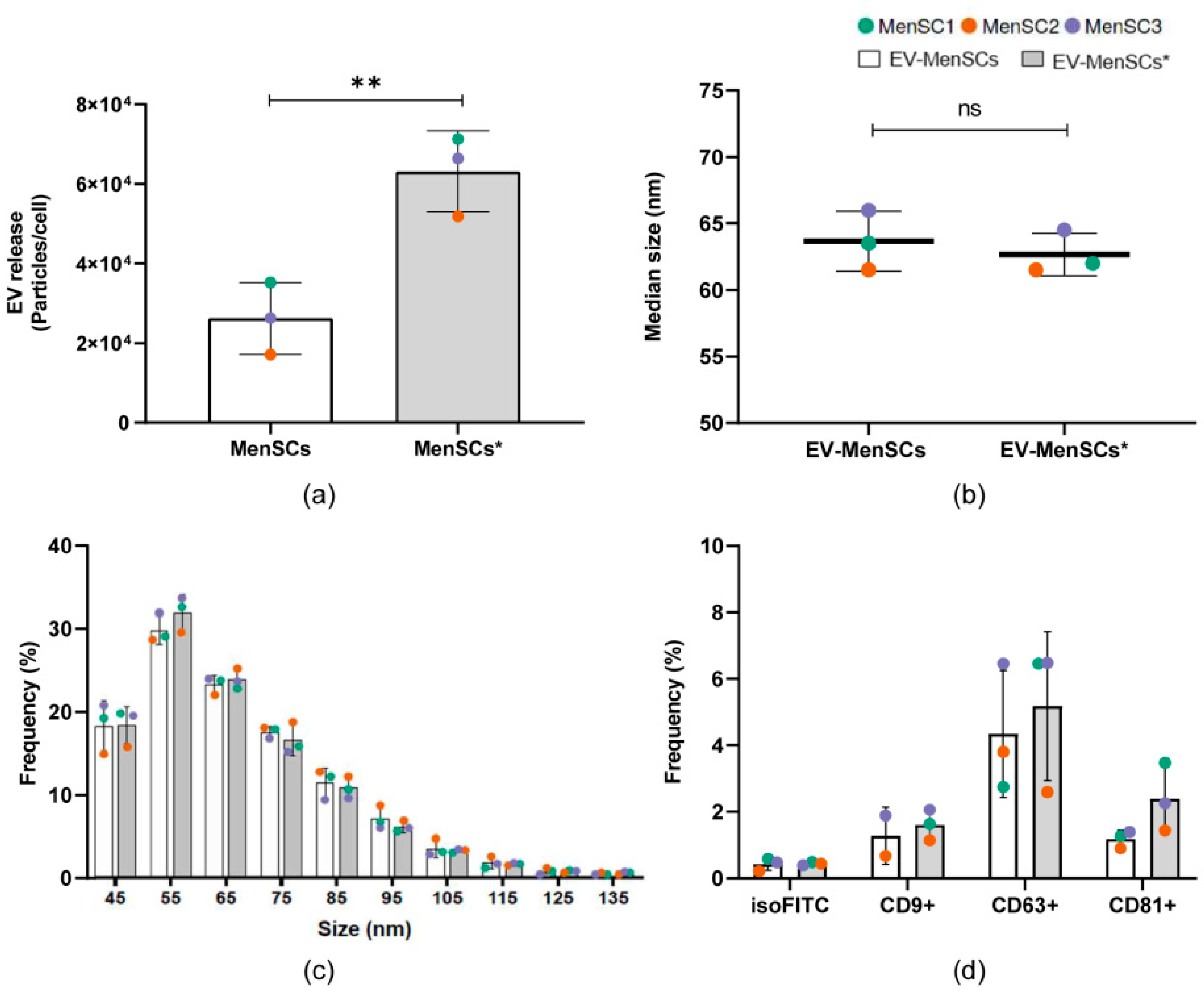
2.4. EV Secretion Profile of MenSCs and Phenotypic Analysis of Released Particles
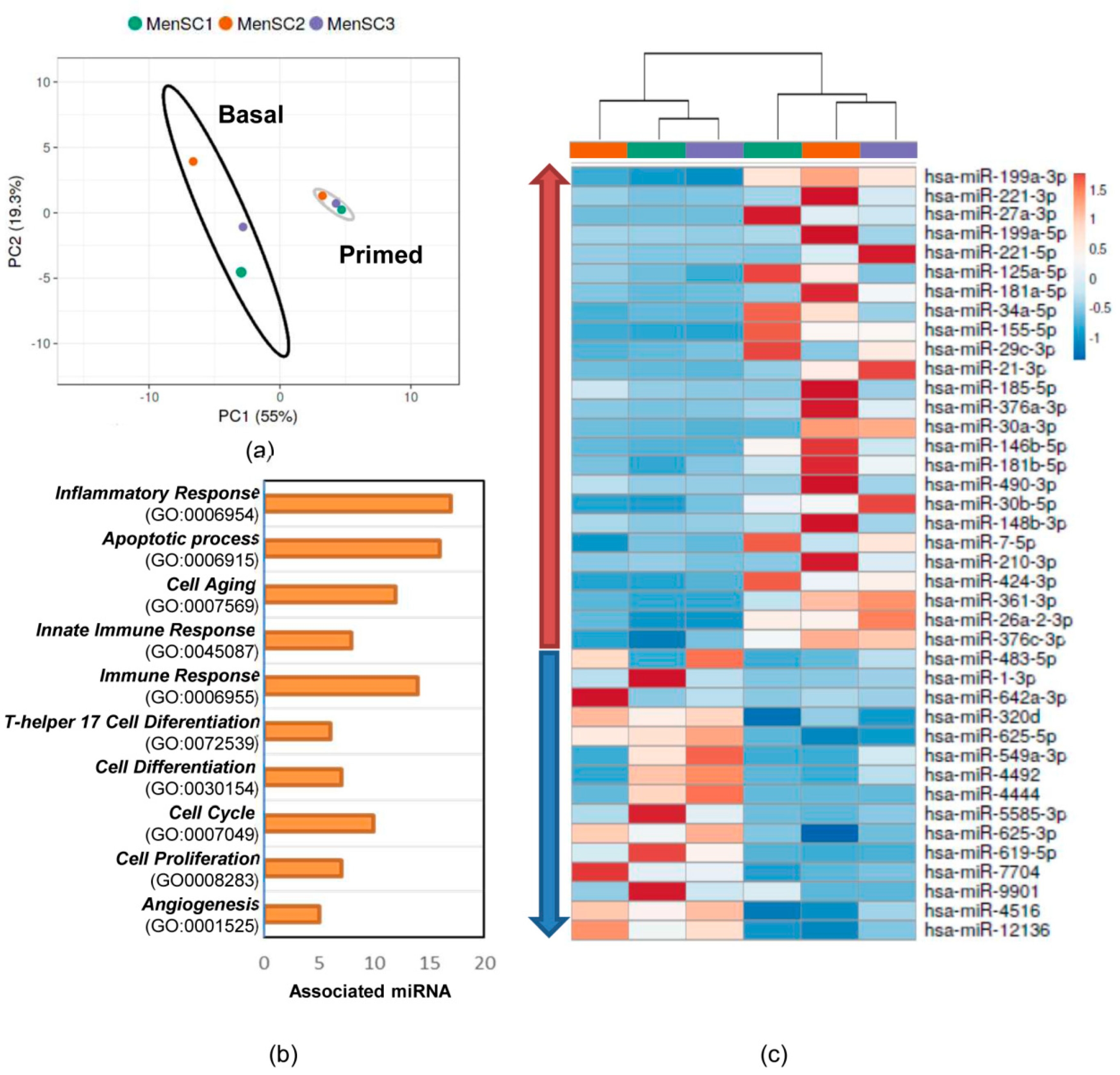
2.5. Effects of IFNγ and TNFα Priming in the miRNome
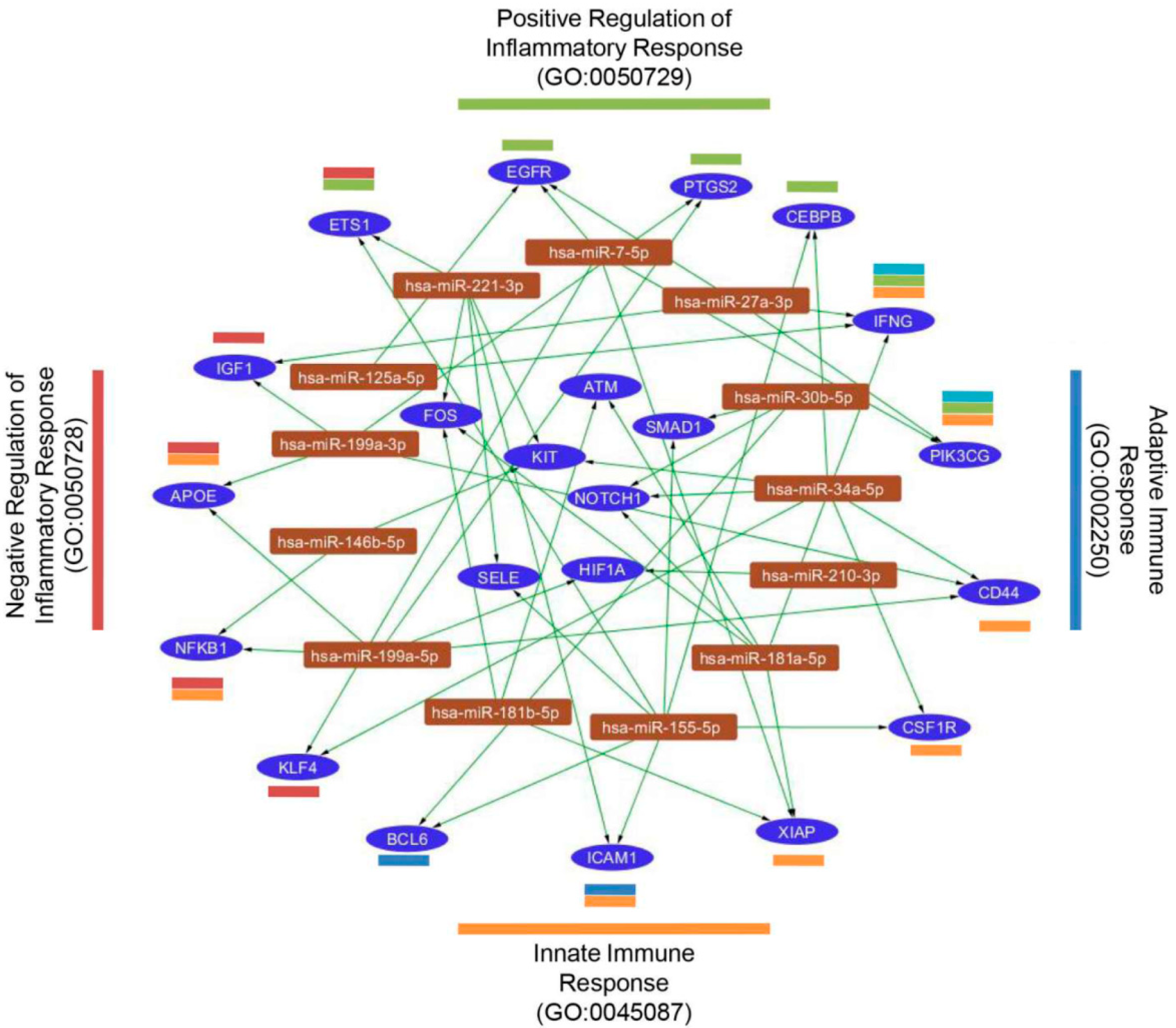
2.6. miRNA Target Prediction in Inflammatory Response
2.7. Validation of miRNA Analyses by qPCR
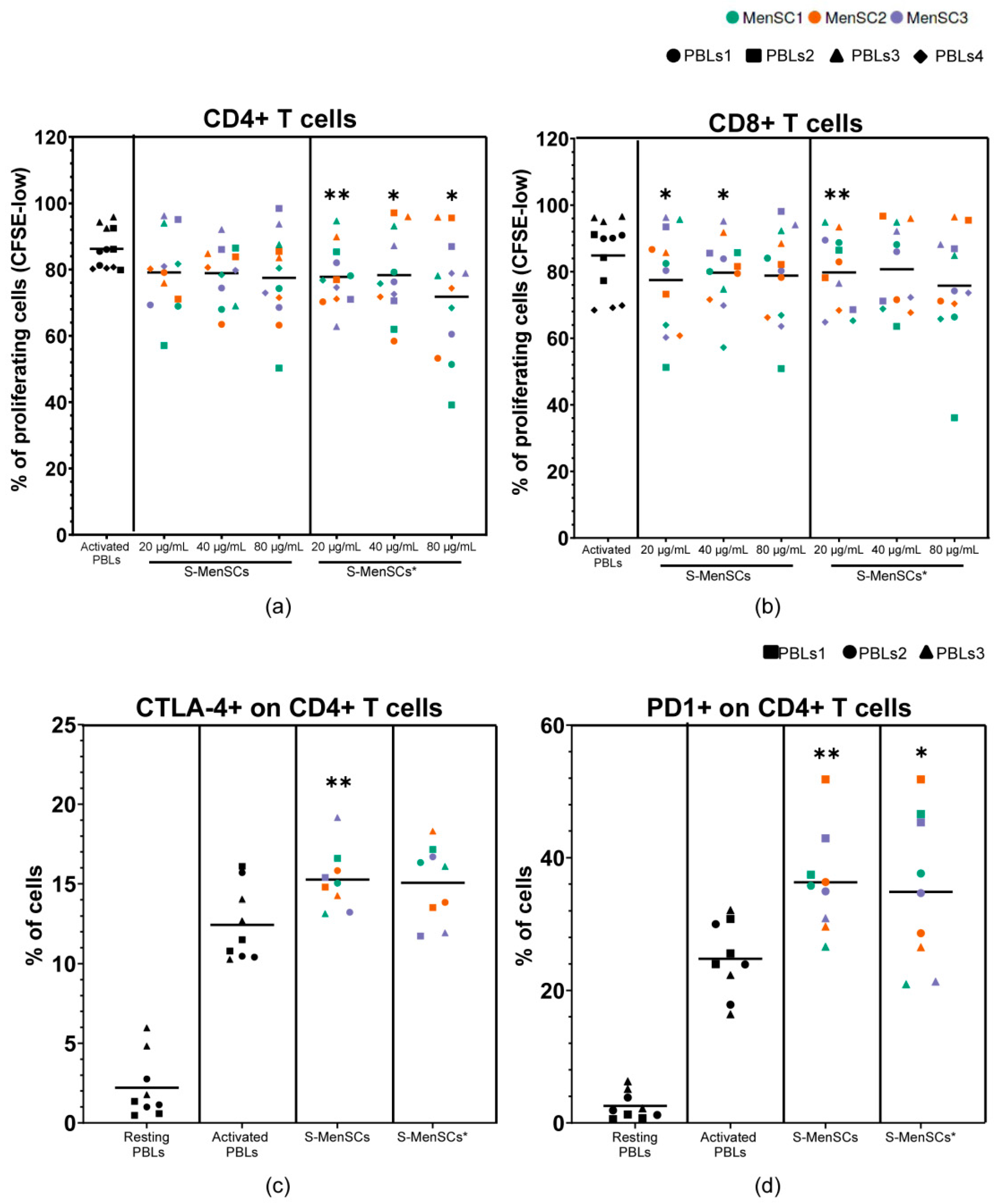
2.8. Immunomodulatory Effect of S-MenSCs and S-MenSCs* on In Vitro Activated T Cells
3. Discussion
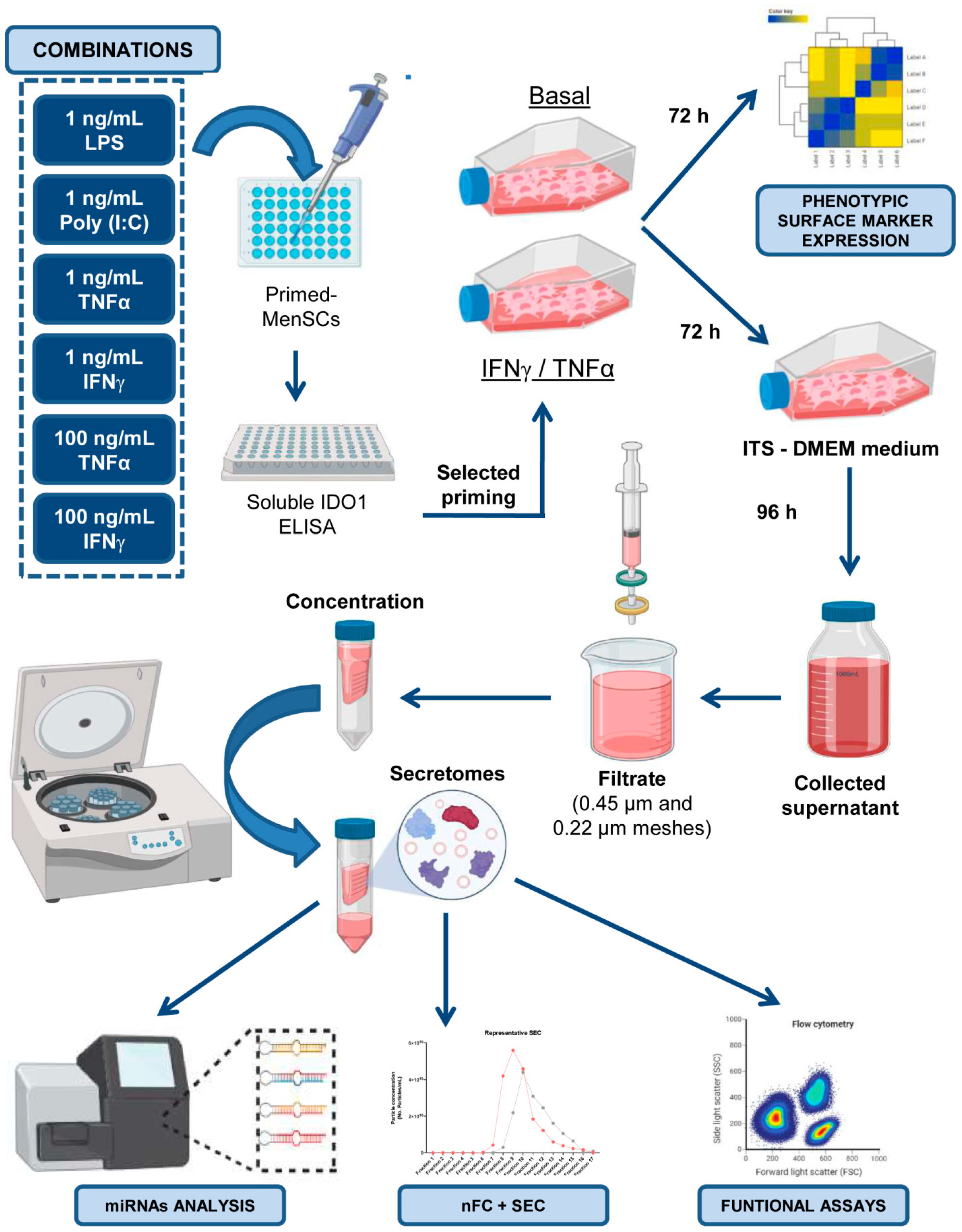
4. Materials and Methods
4.1. Isolation, Culture, and Characterization of Human MenSCs
4.2. Priming of MenSCs and IDO1 Quantification
4.3. Phenotypic Analysis and Comparison of MenSCs and MenSCs* Surface Markers
4.4. Collection, Concentration, and Quantification of MenSC-Derived Secretomes
4.5. Characterization of Extracellular Vesicles in MenSC-Derived Secretomes
4.6. miRNA Analysis by Next-Generation Sequencing (NGS)
4.7. Validation of Transcriptomics Results by qPCR
4.8. Immunomodulatory Assays on In Vitro Activated Peripheral Blood Lymphocytes
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, L.; Qu, J.; Cheng, T.; Chen, X.; Xiang, C. Menstrual Blood-Derived Stem Cells: Toward Therapeutic Mechanisms, Novel Strategies, and Future Perspectives in the Treatment of Diseases. Stem Cell Res. 2019, 10, 406. [Google Scholar] [CrossRef]
- Nikoo, S.; Ebtekar, M.; Jeddi-Tehrani, M.; Shervin, A.; Bozorgmehr, M.; Kazemnejad, S.; Zarnani, A.H. Effect of Menstrual Blood-Derived Stromal Stem Cells on Proliferative Capacity of Peripheral Blood Mononuclear Cells in Allogeneic Mixed Lymphocyte Reaction. J. Obstet. Gynaecol. Res. 2012, 38, 804–809. [Google Scholar] [CrossRef] [PubMed]
- Nikoo, S.; Ebtekar, M.; Jeddi-Tehrani, M.; Shervin, A.; Bozorgmehr, M.; Vafaei, S.; Kazemnejad, S.; Zarnani, A.-H. Menstrual Blood-Derived Stromal Stem Cells from Women with and without Endometriosis Reveal Different Phenotypic and Functional Characteristics. Mol. Hum. Reprod. 2014, 20, 905–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peron, J.P.S.; Jazedje, T.; Brandão, W.N.; Perin, P.M.; Maluf, M.; Evangelista, L.P.; Halpern, S.; Nisenbaum, M.G.; Czeresnia, C.E.; Zatz, M.; et al. Human Endometrial-Derived Mesenchymal Stem Cells Suppress Inflammation in the Central Nervous System of EAE Mice. Stem Cell Rev. 2012, 8, 940–952. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Yuan, Q.; Qu, Y.; Zhou, Y.; Bei, J. Endometrial Mesenchymal Stem Cells Isolated from Menstrual Blood by Adherence. Stem Cells Int. 2016, 2016, 3573846. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Niu, R.; Yang, F.; Yan, Y.; Liang, S.; Sun, Y.; Shen, P.; Lin, J. Biological Characteristics of Human Menstrual Blood-Derived Endometrial Stem Cells. J. Cell. Mol. Med. 2018, 22, 1627–1639. [Google Scholar] [CrossRef]
- Eleuteri, S.; Fierabracci, A. Insights into the Secretome of Mesenchymal Stem Cells and Its Potential Applications. Int. J. Mol. Sci. 2019, 20, 4597. [Google Scholar] [CrossRef] [Green Version]
- Harrell, C.; Fellabaum, C.; Jovicic, N.; Djonov, V.; Arsenijevic, N.; Volarevic, V. Molecular Mechanisms Responsible for Therapeutic Potential of Mesenchymal Stem Cell-Derived Secretome. Cells 2019, 8, 467. [Google Scholar] [CrossRef] [Green Version]
- Cheng, A.; Choi, D.; Lora, M.; Shum-Tim, D.; Rak, J.; Colmegna, I. Human Multipotent Mesenchymal Stromal Cells Cytokine Priming Promotes RAB27B-Regulated Secretion of Small Extracellular Vesicles with Immunomodulatory Cargo. Stem Cell Res. 2020, 11, 539. [Google Scholar] [CrossRef]
- Mitchell, R.; Mellows, B.; Sheard, J.; Antonioli, M.; Kretz, O.; Chambers, D.; Zeuner, M.-T.; Tomkins, J.E.; Denecke, B.; Musante, L.; et al. Secretome of Adipose-Derived Mesenchymal Stem Cells Promotes Skeletal Muscle Regeneration through Synergistic Action of Extracellular Vesicle Cargo and Soluble Proteins. Stem Cell Res. 2019, 10, 116. [Google Scholar] [CrossRef] [PubMed]
- Baldari, S.; Di Rocco, G.; Piccoli, M.; Pozzobon, M.; Muraca, M.; Toietta, G. Challenges and Strategies for Improving the Regenerative Effects of Mesenchymal Stromal Cell-Based Therapies. Int. J. Mol. Sci. 2017, 18, 2087. [Google Scholar] [CrossRef] [Green Version]
- Noronha, N.C.; Mizukami, A.; Caliári-Oliveira, C.; Cominal, J.G.; Rocha, J.L.M.; Covas, D.T.; Swiech, K.; Malmegrim, K.C.R. Priming Approaches to Improve the Efficacy of Mesenchymal Stromal Cell-Based Therapies. Stem Cell Res. 2019, 10, 131. [Google Scholar] [CrossRef] [Green Version]
- Álvarez, V.; Sánchez-Margallo, F.M.; Macías-García, B.; Gómez-Serrano, M.; Jorge, I.; Vázquez, J.; Blázquez, R.; Casado, J.G. The Immunomodulatory Activity of Extracellular Vesicles Derived from Endometrial Mesenchymal Stem Cells on CD4+ T Cells Is Partially Mediated by TGFbeta. J. Tissue Eng. Regen. Med. 2018, 12, 2088–2098. [Google Scholar] [CrossRef] [PubMed]
- Marinaro, F.; Gómez-Serrano, M.; Jorge, I.; Silla-Castro, J.C.; Vázquez, J.; Sánchez-Margallo, F.M.; Blázquez, R.; López, E.; Álvarez, V.; Casado, J.G. Unraveling the Molecular Signature of Extracellular Vesicles From Endometrial-Derived Mesenchymal Stem Cells: Potential Modulatory Effects and Therapeutic Applications. Front. Bioeng. Biotechnol. 2019, 7, 431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varkouhi, A.K.; Jerkic, M.; Ormesher, L.; Gagnon, S.; Goyal, S.; Rabani, R.; Masterson, C.; Spring, C.; Chen, P.Z.; Gu, F.X.; et al. Extracellular Vesicles from Interferon-γ-Primed Human Umbilical Cord Mesenchymal Stromal Cells Reduce Escherichia Coli-Induced Acute Lung Injury in Rats. Anesthesiology 2019, 130, 778–790. [Google Scholar] [CrossRef]
- Gonçalves, F.C.; Luk, F.; Korevaar, S.S.; Bouzid, R.; Paz, A.H.; López-Iglesias, C.; Baan, C.C.; Merino, A.; Hoogduijn, M.J. Membrane Particles Generated from Mesenchymal Stromal Cells Modulate Immune Responses by Selective Targeting of Pro-Inflammatory Monocytes. Sci. Rep. 2017, 7, 12100. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.-C.; Wu, Y.-P.; Li, X.-D.; Chen, S.-H.; Ye, X.-J.; Xue, X.-Y.; Xu, N. TNF-α-Induced Exosomal MiR-146a Mediates Mesenchymal Stem Cell-Dependent Suppression of Urethral Stricture. J. Cell. Physiol. 2019, 234, 23243–23255. [Google Scholar] [CrossRef]
- Lu, Z.; Chen, Y.; Dunstan, C.; Roohani-Esfahani, S.; Zreiqat, H. Priming Adipose Stem Cells with Tumor Necrosis Factor-Alpha Preconditioning Potentiates Their Exosome Efficacy for Bone Regeneration. Tissue Eng. Part A 2017, 23, 1212–1220. [Google Scholar] [CrossRef]
- Mead, B.; Chamling, X.; Zack, D.J.; Ahmed, Z.; Tomarev, S. TNFα-Mediated Priming of Mesenchymal Stem Cells Enhances Their Neuroprotective Effect on Retinal Ganglion Cells. Investig. Ophthalmol. Vis. Sci. 2020, 61, 6. [Google Scholar] [CrossRef] [Green Version]
- Harting, M.T.; Srivastava, A.K.; Zhaorigetu, S.; Bair, H.; Prabhakara, K.S.; Furman, N.E.T.; Vykoukal, J.V.; Ruppert, K.A.; Cox, C.S.; Olson, S.D. Inflammation-Stimulated Mesenchymal Stromal Cell-Derived Extracellular Vesicles Attenuate Inflammation. Stem Cells 2018, 36, 79–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domenis, R.; Cifù, A.; Quaglia, S.; Pistis, C.; Moretti, M.; Vicario, A.; Parodi, P.C.; Fabris, M.; Niazi, K.R.; Soon-Shiong, P.; et al. Pro Inflammatory Stimuli Enhance the Immunosuppressive Functions of Adipose Mesenchymal Stem Cells-Derived Exosomes. Sci. Rep. 2018, 8, 13325. [Google Scholar] [CrossRef] [PubMed]
- An, J.-H.; Li, Q.; Bhang, D.-H.; Song, W.-J.; Youn, H.-Y. TNF-α and INF-γ Primed Canine Stem Cell-Derived Extracellular Vesicles Alleviate Experimental Murine Colitis. Sci. Rep. 2020, 10, 2115. [Google Scholar] [CrossRef]
- François, M.; Romieu-Mourez, R.; Li, M.; Galipeau, J. Human MSC Suppression Correlates with Cytokine Induction of Indoleamine 2,3-Dioxygenase and Bystander M2 Macrophage Differentiation. Mol. Ther. 2012, 20, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Haghighitalab, A.; Matin, M.M.; Amin, A.; Minaee, S.; Bidkhori, H.R.; Doeppner, T.R.; Bahrami, A.R. Investigating the Effects of IDO1, PTGS2, and TGF-Β1 Overexpression on Immunomodulatory Properties of HTERT-MSCs and Their Extracellular Vesicles. Sci. Rep. 2021, 11, 7825. [Google Scholar] [CrossRef]
- Marinaro, F.; Macías-García, B.; Sánchez-Margallo, F.M.; Blázquez, R.; Álvarez, V.; Matilla, E.; Hernández, N.; Gómez-Serrano, M.; Jorge, I.; Vázquez, J.; et al. Extracellular Vesicles Derived from Endometrial Human Mesenchymal Stem Cells Enhance Embryo Yield and Quality in an Aged Murine Model. Biol. Reprod. 2019, 100, 1180–1192. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Blanc, K.L.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Guerrouahen, B.S.; Sidahmed, H.; Al Sulaiti, A.; Al Khulaifi, M.; Cugno, C. Enhancing Mesenchymal Stromal Cell Immunomodulation for Treating Conditions Influenced by the Immune System. Stem Cells Int. 2019, 2019, 7219297. [Google Scholar] [CrossRef] [Green Version]
- Barkholt, L.; Flory, E.; Jekerle, V.; Lucas-Samuel, S.; Ahnert, P.; Bisset, L.; Büscher, D.; Fibbe, W.; Foussat, A.; Kwa, M.; et al. Risk of Tumorigenicity in Mesenchymal Stromal Cell-Based Therapies—Bridging Scientific Observations and Regulatory Viewpoints. Cytotherapy 2013, 15, 753–759. [Google Scholar] [CrossRef]
- Yin, J.Q.; Zhu, J.; Ankrum, J.A. Manufacturing of Primed Mesenchymal Stromal Cells for Therapy. Nat. Biomed. Eng. 2019, 3, 90–104. [Google Scholar] [CrossRef]
- Guess, A.J.; Daneault, B.; Wang, R.; Bradbury, H.; La Perle, K.M.D.; Fitch, J.; Hedrick, S.L.; Hamelberg, E.; Astbury, C.; White, P.; et al. Safety Profile of Good Manufacturing Practice Manufactured Interferon γ-Primed Mesenchymal Stem/Stromal Cells for Clinical Trials. Stem Cells Transl. Med. 2017, 6, 1868–1879. [Google Scholar] [CrossRef]
- Guan, Q.; Li, Y.; Shpiruk, T.; Bhagwat, S.; Wall, D.A. Inducible Indoleamine 2,3-Dioxygenase 1 and Programmed Death Ligand 1 Expression as the Potency Marker for Mesenchymal Stromal Cells. Cytotherapy 2018, 20, 639–649. [Google Scholar] [CrossRef]
- Rovira Gonzalez, Y.I.; Lynch, P.J.; Thompson, E.E.; Stultz, B.G.; Hursh, D.A. In Vitro Cytokine Licensing Induces Persistent Permissive Chromatin at the Indoleamine 2,3-Dioxygenase Promoter. Cytotherapy 2016, 18, 1114–1128. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.S.; Jang, I.K.; Lee, M.W.; Ko, Y.J.; Lee, D.-H.; Lee, J.W.; Sung, K.W.; Koo, H.H.; Yoo, K.H. Enhanced Immunosuppressive Properties of Human Mesenchymal Stem Cells Primed by Interferon-γ. EBioMedicine 2018, 28, 261–273. [Google Scholar] [CrossRef] [Green Version]
- López-García, L.; Castro-Manrreza, M.E. TNF-α and IFN-γ Participate in Improving the Immunoregulatory Capacity of Mesenchymal Stem/Stromal Cells: Importance of Cell–Cell Contact and Extracellular Vesicles. Int. J. Mol. Sci. 2021, 22, 9531. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Xia, Y.-Y.; Wang, L.; Liu, R.; Khoo, K.-S.; Feng, Z.-W. Neural Cell Adhesion Molecule Modulates Mesenchymal Stromal Cell Migration via Activation of MAPK/ERK Signaling. Exp. Cell Res. 2012, 318, 2257–2267. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, Q.; Ding, L.; Wang, Y.-X.; Zhao, Z.-D.; Mao, N.; Wu, C.-T.; Wang, H.; Zhu, H.; Ning, S.-B. Intercellular Adhesion Molecule-1 Enhances the Therapeutic Effects of MSCs in a Dextran Sulfate Sodium-Induced Colitis Models by Promoting MSCs Homing to Murine Colons and Spleens. Stem Cell Res. 2019, 10, 267. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Li, X.; Liu, Y.; Chen, X.; Li, X.; Chu, Y.; Zhu, H.; Liu, W.; Xu, F.; Zhou, F.; et al. The Therapeutic Effect of ICAM-1-Overexpressing Mesenchymal Stem Cells on Acute Graft-Versus-Host Disease. Cell. Physiol. Biochem. 2018, 46, 2624–2635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasef, A.; Chapel, A.; Mazurier, C.; Bouchet, S.; Lopez, M.; Mathieu, N.; Sensebé, L.; Zhang, Y.; Gorin, N.-C.; Thierry, D.; et al. Identification of IL-10 and TGF-β Transcripts Involved in the Inhibition of T-Lymphocyte Proliferation During Cell Contact with Human Mesenchymal Stem Cells. Gene Expr. 2018, 13, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Roson-Burgo, B.; Sanchez-Guijo, F.; Del Cañizo, C.; De Las Rivas, J. Insights into the Human Mesenchymal Stromal/Stem Cell Identity through Integrative Transcriptomic Profiling. BMC Genom. 2016, 17, 944. [Google Scholar] [CrossRef] [Green Version]
- Devine, S.M.; Cobbs, C.; Jennings, M.; Bartholomew, A.; Hoffman, R. Mesenchymal Stem Cells Distribute to a Wide Range of Tissues Following Systemic Infusion into Nonhuman Primates. Blood 2003, 101, 2999–3001. [Google Scholar] [CrossRef]
- Wu, Y.; Hoogduijn, M.J.; Baan, C.C.; Korevaar, S.S.; de Kuiper, R.; Yan, L.; Wang, L.; van Besouw, N.M. Adipose Tissue-Derived Mesenchymal Stem Cells Have a Heterogenic Cytokine Secretion Profile. Stem Cells Int. 2017, 2017, 4960831. [Google Scholar] [CrossRef]
- Xie, Z.; Tang, S.; Ye, G.; Wang, P.; Li, J.; Liu, W.; Li, M.; Wang, S.; Wu, X.; Cen, S.; et al. Interleukin-6/Interleukin-6 Receptor Complex Promotes Osteogenic Differentiation of Bone Marrow-Derived Mesenchymal Stem Cells. Stem Cell Res. 2018, 9, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, W.; Chen, H.; Su, H.; Wu, X.; Xie, Z.; Wu, Y.; Shen, H. IL6 Receptor Facilitates Adipogenesis Differentiation of Human Mesenchymal Stem Cells through Activating P38 Pathway. Int. J. Stem Cells 2020, 13, 142–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaber, T.; Schönbeck, K.; Hoff, H.; Tran, C.L.; Strehl, C.; Lang, A.; Ohrndorf, S.; Pfeiffenberger, M.; Röhner, E.; Matziolis, G.; et al. CTLA-4 Mediates Inhibitory Function of Mesenchymal Stem/Stromal Cells. Int. J. Mol. Sci. 2018, 19, 2312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, L.C.; Heldring, N.; Kadri, N.; Le Blanc, K. Mesenchymal Stromal Cell Secretion of Programmed Death-1 Ligands Regulates T Cell Mediated Immunosuppression. Stem Cells 2017, 35, 766–776. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, Q.; Zhang, J.; Li, C.; Miao, Y.-R.; Lei, Q.; Li, Q.; Guo, A.-Y. EVmiRNA: A Database of MiRNA Profiling in Extracellular Vesicles. Nucleic Acids Res. 2019, 47, D89–D93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohel, M.H. Extracellular/Circulating MicroRNAs: Release Mechanisms, Functions and Challenges. Achiev. Life Sci. 2016, 10, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, D.K.; Fenix, A.M.; Franklin, J.L.; Higginbotham, J.N.; Zhang, Q.; Zimmerman, L.J.; Liebler, D.C.; Ping, J.; Liu, Q.; Evans, R.; et al. Reassessment of Exosome Composition. Cell 2019, 177, 428–445.e18. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Huang, R.; Xu, Q.; Zheng, G.; Qiu, G.; Ge, M.; Shu, Q.; Xu, J. Mesenchymal Stem Cell–Derived Extracellular Vesicles Alleviate Acute Lung Injury Via Transfer of MiR-27a-3p*. Crit. Care Med. 2020, 48, e599. [Google Scholar] [CrossRef]
- Wang, J.; Jia, Z.; Wei, B.; Zhou, Y.; Niu, C.; Bai, S.; Tong, C.; Song, J.; Li, Y. MicroRNA-27a Restrains the Immune Response to Mycobacterium Tuberculosis Infection by Targeting IRAK4, a Promoter of the NF-ΚB Pathway. Int. J. Clin. Exp. Pathol. 2017, 10, 9894–9901. [Google Scholar]
- Wang, L.; Yin, P.; Wang, J.; Wang, Y.; Sun, Z.; Zhou, Y.; Guan, X. Delivery of Mesenchymal Stem Cells-Derived Extracellular Vesicles with Enriched MiR-185 Inhibits Progression of OPMD. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2481–2491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ragni, E.; Orfei, C.P.; De Luca, P.; Mondadori, C.; Viganò, M.; Colombini, A.; de Girolamo, L. Inflammatory Priming Enhances Mesenchymal Stromal Cell Secretome Potential as a Clinical Product for Regenerative Medicine Approaches through Secreted Factors and EV-MiRNAs: The Example of Joint Disease. Stem Cell Res. 2020, 11, 165. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Ren, G.; Cao, G.; Chen, Q.; Shou, P.; Zheng, C.; Du, L.; Han, X.; Jiang, M.; Yang, Q.; et al. MiR-155 Regulates Immune Modulatory Properties of Mesenchymal Stem Cells by Targeting TAK1-Binding Protein 2. J. Biol. Chem. 2013, 288, 11074–11079. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Hua, F.; Yang, Z.; Chen, Y.; Teng, X.; Huang, H.; Zhao, Y.; Shen, Z. Enhancement of Immunoregulatory Function of Modified Bone Marrow Mesenchymal Stem Cells by Targeting SOCS1. Available online: https://www.hindawi.com/journals/bmri/2018/3530647/ (accessed on 10 July 2020).
- BioRender. Available online: https://biorender.com/ (accessed on 9 November 2021).
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Gene Ontology Resource. Available online: http://geneontology.org/ (accessed on 17 May 2021).
- Home - Reactome Pathway Database. Available online: https://reactome.org/ (accessed on 17 May 2021).
- Babraham Bioinformatics—FastQC A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 20 October 2021).
- Metsalu, T.; Vilo, J. ClustVis: A Web Tool for Visualizing Clustering of Multivariate Data Using Principal Component Analysis and Heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- MiRNet. Available online: https://www.mirnet.ca/upload/MirUploadView.xhtml (accessed on 7 September 2021).
- TAM 2.0. Available online: http://www.lirmed.com/tam2/ (accessed on 7 September 2021).
- Hamberg, M.; Backes, C.; Fehlmann, T.; Hart, M.; Meder, B.; Meese, E.; Keller, A. MiRTargetLink--MiRNAs, Genes and Interaction Networks. Int. J. Mol. Sci. 2016, 17, 564. [Google Scholar] [CrossRef]
- FunRich: Functional Enrichment Analysis Tool: Home. Available online: http://www.funrich.org/ (accessed on 9 September 2021).
- Drobna, M.; Szarzyńska-Zawadzka, B.; Daca-Roszak, P.; Kosmalska, M.; Jaksik, R.; Witt, M.; Dawidowska, M. Identification of Endogenous Control MiRNAs for RT-QPCR in T-Cell Acute Lymphoblastic Leukemia. Int. J. Mol. Sci. 2018, 19, 2858. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Pedro, M.Á.; Gómez-Serrano, M.; Marinaro, F.; López, E.; Pulido, M.; Preußer, C.; Pogge von Strandmann, E.; Sánchez-Margallo, F.M.; Álvarez, V.; Casado, J.G. IFN-Gamma and TNF-Alpha as a Priming Strategy to Enhance the Immunomodulatory Capacity of Secretomes from Menstrual Blood-Derived Stromal Cells. Int. J. Mol. Sci. 2021, 22, 12177. https://doi.org/10.3390/ijms222212177
de Pedro MÁ, Gómez-Serrano M, Marinaro F, López E, Pulido M, Preußer C, Pogge von Strandmann E, Sánchez-Margallo FM, Álvarez V, Casado JG. IFN-Gamma and TNF-Alpha as a Priming Strategy to Enhance the Immunomodulatory Capacity of Secretomes from Menstrual Blood-Derived Stromal Cells. International Journal of Molecular Sciences. 2021; 22(22):12177. https://doi.org/10.3390/ijms222212177
Chicago/Turabian Stylede Pedro, María Ángeles, María Gómez-Serrano, Federica Marinaro, Esther López, María Pulido, Christian Preußer, Elke Pogge von Strandmann, Francisco Miguel Sánchez-Margallo, Verónica Álvarez, and Javier G. Casado. 2021. "IFN-Gamma and TNF-Alpha as a Priming Strategy to Enhance the Immunomodulatory Capacity of Secretomes from Menstrual Blood-Derived Stromal Cells" International Journal of Molecular Sciences 22, no. 22: 12177. https://doi.org/10.3390/ijms222212177
APA Stylede Pedro, M. Á., Gómez-Serrano, M., Marinaro, F., López, E., Pulido, M., Preußer, C., Pogge von Strandmann, E., Sánchez-Margallo, F. M., Álvarez, V., & Casado, J. G. (2021). IFN-Gamma and TNF-Alpha as a Priming Strategy to Enhance the Immunomodulatory Capacity of Secretomes from Menstrual Blood-Derived Stromal Cells. International Journal of Molecular Sciences, 22(22), 12177. https://doi.org/10.3390/ijms222212177