
Phosphoproteomic Comparison of Four Eimeria tenella Life Cycle Stages


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Identification, Quantification, and Classification of Phosphopeptides
2.2. Identification of Differentially Phosphorylated Proteins
2.3. Functional Enrichment of DEPPs
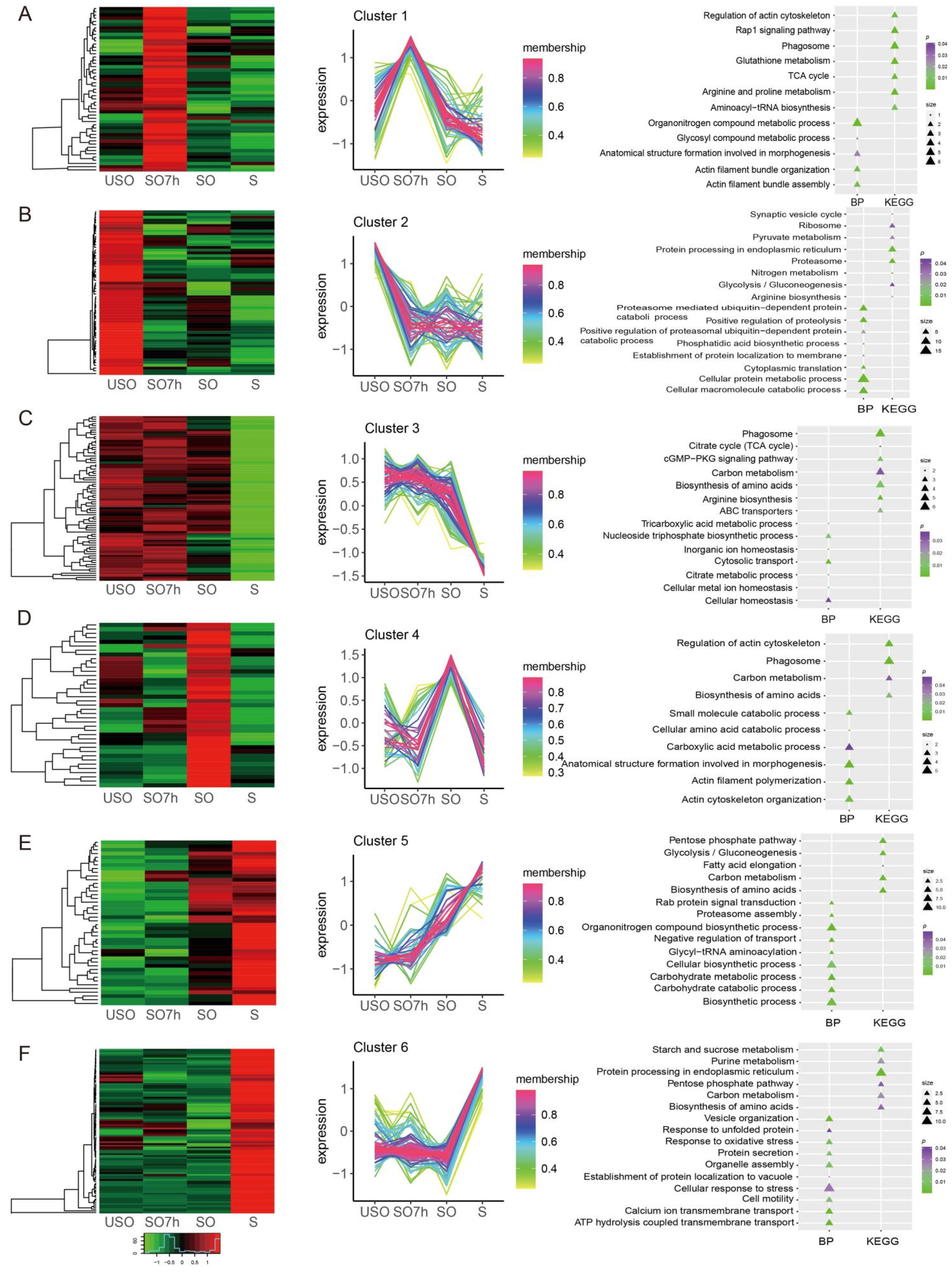
2.4. Protein Phosphorylation Patterns
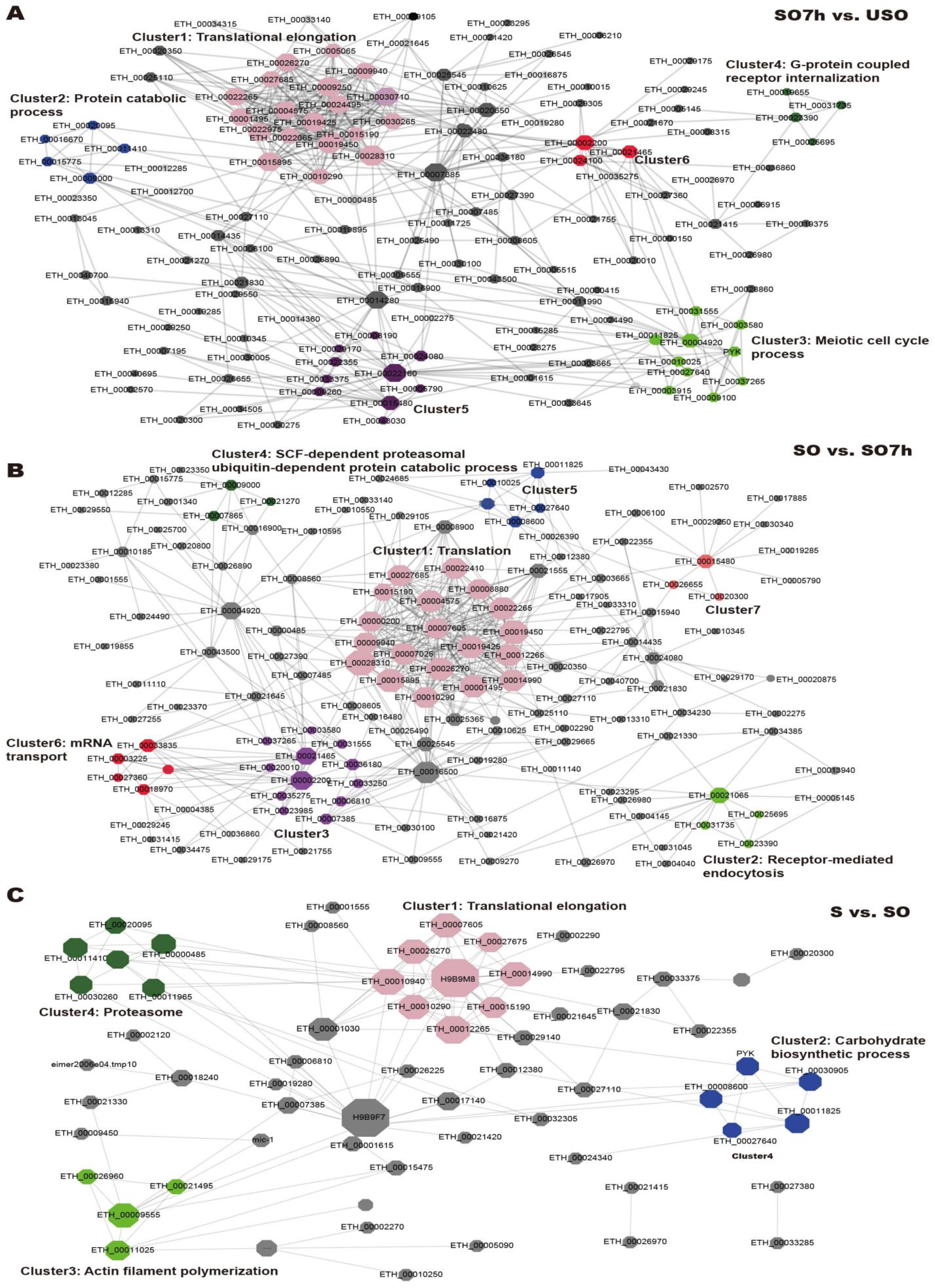
2.5. Protein–Protein Interaction (PPI) Network Analysis of DEPPs
3. Discussion
4. Materials and Methods
4.1. Chickens and Parasites
4.2. Protein Extraction
4.3. Trypsin Digestion
4.4. Affinity Enrichment
4.5. LC-MS/MS Analysis
4.6. Database Search
4.7. Bioinformatics Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shen, X.J.; Li, T.; Fu, J.J.; Zhang, K.Y.; Wang, X.Y.; Liu, Y.C.; Zhang, H.J.; Fan, C.; Fei, C.Z.; Xue, F.Q. Proteomic analysis of the effect of diclazuril on second-generation merozoites of Eimeria tenella. Parasitol. Res. 2014, 113, 903–909. [Google Scholar] [CrossRef]
- Walker, R.A.; Sharman, P.A.; Miller, C.M.; Lippuner, C.; Okoniewski, M.; Eichenberger, R.M.; Ramakrishnan, C.; Brossier, F.; Deplazes, P.; Hehl, A.B.; et al. RNA Seq analysis of the Eimeria tenella gametocyte transcriptome reveals clues about the molecular basis for sexual reproduction and oocyst biogenesis. BMC Genom. 2015, 16, 94. [Google Scholar] [CrossRef]
- Su, S.; Hou, Z.; Liu, D.; Jia, C.; Wang, L.; Xu, J.; Tao, J. Comparative transcriptome analysis of second- and third-generation merozoites of Eimeria necatrix. Parasites Vectors 2017, 10, 388. [Google Scholar] [CrossRef]
- Klotz, C.; Marhöfer, R.J.; Selzer, P.M.; Lucius, R.; Pogonka, T. Eimeria tenella: Identification of secretory and surface proteins from expressed sequence tags. Exp. Parasitol. 2005, 111, 14–23. [Google Scholar] [CrossRef]
- Miska, K.B.; Fetterer, R.H.; Barfield, R.C. Analysis of Transcripts Expressed by Eimeria Tenella Oocysts Using Subtractive Hybridization Methods. J. Parasitol. 2004, 90, 1245–1252. [Google Scholar] [CrossRef]
- Di Genova, B.M.; Knoll, L.J. Comparisons of the Sexual Cycles for the Coccidian Parasites Eimeria and Toxoplasma. Front. Cell Infect. Microbiol. 2020, 10, 604897. [Google Scholar] [CrossRef] [PubMed]
- Amiruddin, N.; Lee, X.-W.; Blake, D.P.; Suzuki, Y.; Tay, Y.-L.; Lim, L.-S.; Tomley, F.M.; Watanabe, J.; Sugimoto, C.; Wan, K.-L. Characterisation of full-length cDNA sequences provides insights into the Eimeria tenella transcriptome. BMC Genom. 2012, 13, 21. [Google Scholar] [CrossRef]
- Lal, K.; Bromley, E.; Oakes, R.; Prieto, J.H.; Sanderson, S.J.; Kurian, D.; Hunt, L.; Yates, J.R., III; Wastling, J.M.; Sinden, R.E.; et al. Proteomic comparison of four Eimeria tenella life-cycle stages: Unsporulated oocyst, sporulated oocyst, sporozoite and second-generation merozoite. Proteomics 2009, 9, 4566–4576. [Google Scholar] [CrossRef] [PubMed]
- De Venevelles, P.; Chich, J.F.; Faigle, W.; Loew, D.; Labbé, M.; Girard-Misguich, F.; Péry, P. Towards a reference map of Eimeria tenella sporozoite proteins by two-dimensional electrophoresis and mass spectrometry. Int. J. Parasitol. 2004, 34, 1321–1331. [Google Scholar] [CrossRef] [PubMed]
- Oakes, R.D.; Kurian, D.; Bromley, E.; Ward, C.; Lal, K.; Blake, D.P.; Reid, A.J.; Pain, A.; Sinden, R.E.; Wastling, J.M.; et al. The rhoptry proteome of Eimeria tenella sporozoites. Int. J. Parasitol. 2013, 43, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-Y.; Liu, L.-L.; Zhang, M.; Zhang, L.-F.; Wang, X.-Y.; Wang, M.; Zhang, K.-Y.; Liu, Y.-C.; Wang, C.-M.; Xue, F.-Q.; et al. Proteomic analysis of the second-generation merozoites of Eimeria tenella under nitromezuril and ethanamizuril stress. Parasites Vectors 2019, 12, 592. [Google Scholar] [CrossRef]
- Thabet, A.; Honscha, W.; Daugschies, A.; Bangoura, B. Quantitative proteomic studies in resistance mechanisms of Eimeria tenella against polyether ionophores. Parasitol. Res. 2017, 116, 1553–1559. [Google Scholar] [CrossRef]
- Yakubu, R.R.; Weiss, L.M.; Silmon de Monerri, N.C. Post-translational modifications as key regulators of apicomplexan biology: Insights from proteome-wide studies. Mol. Microbiol. 2018, 107, 1–23. [Google Scholar] [CrossRef]
- Shumyantseva, V.V.; Suprun, E.V.; Bulko, T.V.; Archakov, A.I. Electrochemical methods for detection of post-translational modifications of proteins. Biosens. Bioelectron. 2014, 61, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.V.; Blagoev, B.; Gnad, F.; Macek, B.; Kumar, C.; Mortensen, P.; Mann, M. Global, In Vivo, and Site-Specific Phosphorylation Dynamics in Signaling Networks. Cell 2006, 127, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Green, J.L.; Rees-Channer, R.R.; Howell, S.A.; Martin, S.R.; Knuepfer, E.; Taylor, H.M.; Grainger, M.; Holder, A.A. The Motor Complex of Plasmodium falciparum: Phosphorylation by a Calcium-Dependent Protein Kinase. J. Biol. Chem. 2008, 283, 30980–30989. [Google Scholar] [CrossRef] [PubMed]
- Joyce, B.R.; Queener, S.F.; Wek, R.C.; Sullivan, W.J. Phosphorylation of eukaryotic initiation factor-2α promotes the extracellular survival of obligate intracellular parasite Toxoplasma gondii. Proc. Natl. Acad. Sci. USA 2010, 107, 17200. [Google Scholar] [CrossRef]
- Leykauf, K.; Treeck, M.; Gilson, P.R.; Nebl, T.; Braulke, T.; Cowman, A.F.; Gilberger, T.W.; Crabb, B.S. Protein kinase a dependent phosphorylation of apical membrane antigen 1 plays an important role in erythrocyte invasion by the malaria parasite. PLoS Pathog. 2010, 6, e1000941. [Google Scholar] [CrossRef]
- Treeck, M.; Zacherl, S.; Herrmann, S.; Cabrera, A.; Kono, M.; Struck, N.S.; Engelberg, K.; Haase, S.; Frischknecht, F.; Miura, K.; et al. Functional analysis of the leading malaria vaccine candidate AMA-1 reveals an essential role for the cytoplasmic domain in the invasion process. PLoS Pathog. 2009, 5, e1000322. [Google Scholar] [CrossRef]
- Matsubayashi, M.; Hatta, T.; Miyoshi, T.; Sasai, K.; Shimura, K.; Isobe, T.; Kita, K.; Tsuji, N. High-throughput RNA sequencing profiles and transcriptional evidence of aerobic respiratory enzymes in sporulating oocysts and sporozoites of Eimeria tenella. Infect. Genet. Evol. 2013, 18, 269–276. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, Q.; Zhu, S.; Dong, H.; Yu, S.; Wang, Q.; Yu, Y.; Liang, S.; Zhao, H.; Huang, B.; et al. Molecular characterization and functional analysis of Eimeria tenella citrate synthase. Parasitol. Res. 2021, 120, 1025–1035. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, S.; Li, C.; Liu, L. Immunoproteomic analysis of the protein repertoire of unsporulated Eimeria tenella oocysts. Parasite 2017, 24, 48. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bozdech, Z.; Ginsburg, H. Data mining of the transcriptome of Plasmodium falciparum: The pentose phosphate pathway and ancillary processes. Malar. J. 2005, 4, 17. [Google Scholar] [CrossRef]
- MacRae, J.I.; Sheiner, L.; Nahid, A.; Tonkin, C.; Striepen, B.; McConville, M.J. Mitochondrial Metabolism of Glucose and Glutamine Is Required for Intracellular Growth of Toxoplasma gondii. Cell Host Microbe 2012, 12, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Olson, W.J.; Di Genova, B.M.; Gallego-Lopez, G.; Dawson, A.R.; Stevenson, D.; Amador-Noguez, D.; Knoll, L.J. Dual metabolomic profiling uncovers Toxoplasma manipulation of the host metabolome and the discovery of a novel parasite metabolic capability. PLoS Pathog. 2020, 16, e1008432. [Google Scholar] [CrossRef]
- Karkhanis, Y.D.; Allocco, J.J.; Schmatz, D.M. Amylopectin Synthase of Eimeria tenella: Identification and Kinetic Characterization. J. Eukaryot. Microbiol. 1993, 40, 594–598. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, A.N.; Kok, J.J.; van den Boogaart, P.; Dijkema, R.; Claessens, J.A.J. Eimeria refractile body proteins contain two potentially functional characteristics: Transhydrogenase and carbohydrate transport. FEMS Microbiol. Lett. 1993, 110, 223–229. [Google Scholar] [CrossRef]
- Michalski, W.P.; Edgar, J.A.; Prowse, S.J. Mannitol metabolism in Eimeria tenella. Int. J. Parasitol. 1992, 22, 1157–1163. [Google Scholar] [CrossRef]
- Nakai, Y.; Ogimoto, K. Relationship between amylopectin and infectivity of Eimeria tenella sporozoite. Jpn. J. Vet. Sci. 1987, 49, 447–452. [Google Scholar] [CrossRef]
- Nakai, Y.; Ogimoto, K. Relationship between amylopectin and viability of Eimeria tenella sporozoite. Jpn. J. Vet. Sci. 1983, 45, 127–129. [Google Scholar] [CrossRef]
- Labbé, M.; Péroval, M.; Bourdieu, C.; Girard-Misguich, F.; Péry, P. Eimeria tenella enolase and pyruvate kinase: A likely role in glycolysis and in others functions. Int. J. Parasitol. 2006, 36, 1443–1452. [Google Scholar] [CrossRef] [PubMed]
- Morrissette, N.S.; Sibley, L.D. Cytoskeleton of Apicomplexan Parasites. Microbiol. Mol. Biol. Rev. 2002, 66, 21–38. [Google Scholar] [CrossRef]
- Jacot, D.; Daher, W.; Soldati-Favre, D. Toxoplasma gondii myosin F, an essential motor for centrosomes positioning and apicoplast inheritance. EMBO J. 2013, 32, 1702–1716. [Google Scholar] [CrossRef]
- Das, S.; Stortz, J.F.; Meissner, M.; Periz, J. The multiple functions of actin in apicomplexan parasites. Cell. Microbiol. 2021, 23, e13345. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Lemgruber, L.; Tay, C.L.; Baum, J.; Meissner, M. Multiple essential functions of Plasmodium falciparum actin-1 during malaria blood-stage development. BMC Biol. 2017, 15, 70. [Google Scholar] [CrossRef] [PubMed]
- Andenmatten, N.; Egarter, S.; Jackson, A.J.; Jullien, N.; Herman, J.-P.; Meissner, M. Conditional genome engineering in Toxoplasma gondii uncovers alternative invasion mechanisms. Nat. Methods 2013, 10, 125–127. [Google Scholar] [CrossRef]
- Whitelaw, J.A.; Latorre-Barragan, F.; Gras, S.; Pall, G.S.; Leung, J.M.; Heaslip, A.; Egarter, S.; Andenmatten, N.; Nelson, S.R.; Warshaw, D.M.; et al. Surface attachment, promoted by the actomyosin system of Toxoplasma gondii is important for efficient gliding motility and invasion. BMC Biol. 2017, 15, 1. [Google Scholar] [CrossRef]
- Smythe, W.A.; Joiner, K.A.; Hoppe, H.C. Actin is required for endocytic trafficking in the malaria parasite Plasmodium falciparum. Cell. Microbiol. 2008, 10, 452–464. [Google Scholar] [CrossRef]
- Burrell, A.; Tomley, F.M.; Vaughan, S.; Marugan-Hernandez, V. Life cycle stages, specific organelles and invasion mechanisms of Eimeria species. Parasitology 2020, 147, 263–278. [Google Scholar] [CrossRef]
- Dubois, D.J.; Soldati-Favre, D. Biogenesis and secretion of micronemes in Toxoplasma gondii. Cell. Microbiol. 2019, 21, e13018. [Google Scholar] [CrossRef]
- Bullen, H.E.; Soldati-Favre, D. A central role for phosphatidic acid as a lipid mediator of regulated exocytosis in apicomplexa. FEBS Lett. 2016, 590, 2469–2481. [Google Scholar] [CrossRef]
- Tomley, F.M.; Bumstead, J.M.; Billington, K.J.; Dunn, P.P.J. Molecular cloning and characterization of a novel acidic microneme protein (Etmic-2) from the apicomplexan protozoan parasite, Eimeria tenella. Mol. Biochem. Parasitol. 1996, 79, 195–206. [Google Scholar] [CrossRef]
- Dubremetz, J.F.; Garcia-Réguet, N.; Conseil, V.; Fourmaux, M.N. Invited review Apical organelles and host-cell invasion by Apicomplexa. Int. J. Parasitol. 1998, 28, 1007–1013. [Google Scholar] [CrossRef]
- Zerial, M.; McBride, H. Rab proteins as membrane organizers. Nat. Rev. Mol. Cell Biol. 2001, 2, 107–117. [Google Scholar] [CrossRef]
- Morlon-Guyot, J.; Pastore, S.; Berry, L.; Lebrun, M.; Daher, W. Toxoplasma gondii Vps11, a subunit of HOPS and CORVET tethering complexes, is essential for the biogenesis of secretory organelles. Cell. Microbiol. 2015, 17, 1157–1178. [Google Scholar] [CrossRef]
- Kremer, K.; Kamin, D.; Rittweger, E.; Wilkes, J.; Flammer, H.; Mahler, S.; Heng, J.; Tonkin, C.J.; Langsley, G.; Hell, S.W.; et al. An overexpression screen of Toxoplasma gondii Rab-GTPases reveals distinct transport routes to the micronemes. PLoS Pathog. 2013, 9, e1003213. [Google Scholar] [CrossRef] [PubMed]
- Quevillon, E.; Spielmann, T.; Brahimi, K.; Chattopadhyay, D.; Yeramian, E.; Langsley, G. The Plasmodium falciparum family of Rab GTPases. Gene 2003, 306, 13–25. [Google Scholar] [CrossRef]
- Tisdale, E.J.; Balch, W.E. Rab2 Is Essential for the Maturation of Pre-Golgi Intermediates. J. Biol. Chem. 1996, 271, 29372–29379. [Google Scholar] [CrossRef]
- Daubenberger, C.A.; Tisdale, E.J.; Curcic, M.; Diaz, D.; Silvie, O.; Mazier, D.; Eling, W.; Bohrmann, B.; Matile, H.; Pluschke, G. The N’-Terminal Domain of Glyceraldehyde-3-Phosphate Dehydrogenase of the Apicomplexan Plasmodium falciparum Mediates GTPase Rab2-Dependent Recruitment to Membranes. Biol Chem. 2003, 384, 1227–1237. [Google Scholar] [CrossRef] [PubMed]
- Dhir, V.; Goulding, D.; Field, M.C. TbRAB1 and TbRAB2 mediate trafficking through the early secretory pathway of Trypanosoma brucei. Mol. Biochem. Parasitol. 2004, 137, 253–265. [Google Scholar] [CrossRef]
- Yan, X.; Tao, G.; Liu, X.; Ji, Y.; Suo, X. Calcium-dependent microneme protein discharge and in vitro egress of Eimeria tenella sporozoites. Exp. Parasitol. 2016, 170, 193–197. [Google Scholar] [CrossRef]
- Katrib, M.; Ikin, R.J.; Brossier, F.; Robinson, M.; Slapetova, I.; Sharman, P.A.; Walker, R.A.; Belli, S.I.; Tomley, F.M.; Smith, N.C. Stage-specific expression of protease genes in the apicomplexan parasite, Eimeria tenella. BMC Genom. 2012, 13, 685. [Google Scholar] [CrossRef]
- Tomley, F. Techniques for Isolation and Characterization of Apical Organelles from Eimeria tenella Sporozoites. Methods 1997, 13, 171–176. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Camon, E.; Magrane, M.; Barrell, D.; Lee, V.; Dimmer, E.; Maslen, J.; Binns, D.; Harte, N.; Lopez, R.; Apweiler, R. The Gene Ontology Annotation (GOA) Database: Sharing knowledge in Uniprot with Gene Ontology. Nucleic Acids Res. 2004, 32, D262–D266. [Google Scholar] [CrossRef] [PubMed]
- Apweiler, R.; Bairoch, A.; Wu, C.H.; Barker, W.C.; Boeckmann, B.; Ferro, S.; Gasteiger, E.; Huang, H.; Lopez, R.; Magrane, M.; et al. UniProt: The Universal Protein knowledgebase. Nucleic Acids Res. 2004, 32, D115–D119. [Google Scholar] [CrossRef]
- Apweiler, R.; Attwood, T.K.; Bairoch, A.; Bateman, A.; Birney, E.; Biswas, M.; Bucher, P.; Cerutti, L.; Corpet, F.; Croning, M.D.R.; et al. InterPro—an integrated documentation resource for protein families, domains and functional sites. Bioinformatics 2000, 16, 1145–1150. [Google Scholar] [CrossRef]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Bader, G.D.; Hogue, C.W.V. An automated method for finding molecular complexes in large protein interaction networks. BMC Bioinform. 2003, 4, 2. [Google Scholar] [CrossRef] [PubMed]
- Jun, M.; Gu, L. TCseq: Time Course Sequencing Data Analysis. 2021. Available online: http://www.bioconductor.org/packages/release/bioc/html/TCseq.html (accessed on 5 August 2021).





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, X.; Liu, B.; Gong, Z.; Qu, Z.; Cai, J. Phosphoproteomic Comparison of Four Eimeria tenella Life Cycle Stages. Int. J. Mol. Sci. 2021, 22, 12110. https://doi.org/10.3390/ijms222212110
Ma X, Liu B, Gong Z, Qu Z, Cai J. Phosphoproteomic Comparison of Four Eimeria tenella Life Cycle Stages. International Journal of Molecular Sciences. 2021; 22(22):12110. https://doi.org/10.3390/ijms222212110
Chicago/Turabian StyleMa, Xueting, Baohong Liu, Zhenxing Gong, Zigang Qu, and Jianping Cai. 2021. "Phosphoproteomic Comparison of Four Eimeria tenella Life Cycle Stages" International Journal of Molecular Sciences 22, no. 22: 12110. https://doi.org/10.3390/ijms222212110
APA StyleMa, X., Liu, B., Gong, Z., Qu, Z., & Cai, J. (2021). Phosphoproteomic Comparison of Four Eimeria tenella Life Cycle Stages. International Journal of Molecular Sciences, 22(22), 12110. https://doi.org/10.3390/ijms222212110