
Dietary Nano-ZnO Is Absorbed via Endocytosis and ZIP Pathways, Upregulates Lipogenesis, and Induces Lipotoxicity in the Intestine of Yellow Catfish

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. In Vivo Assessment
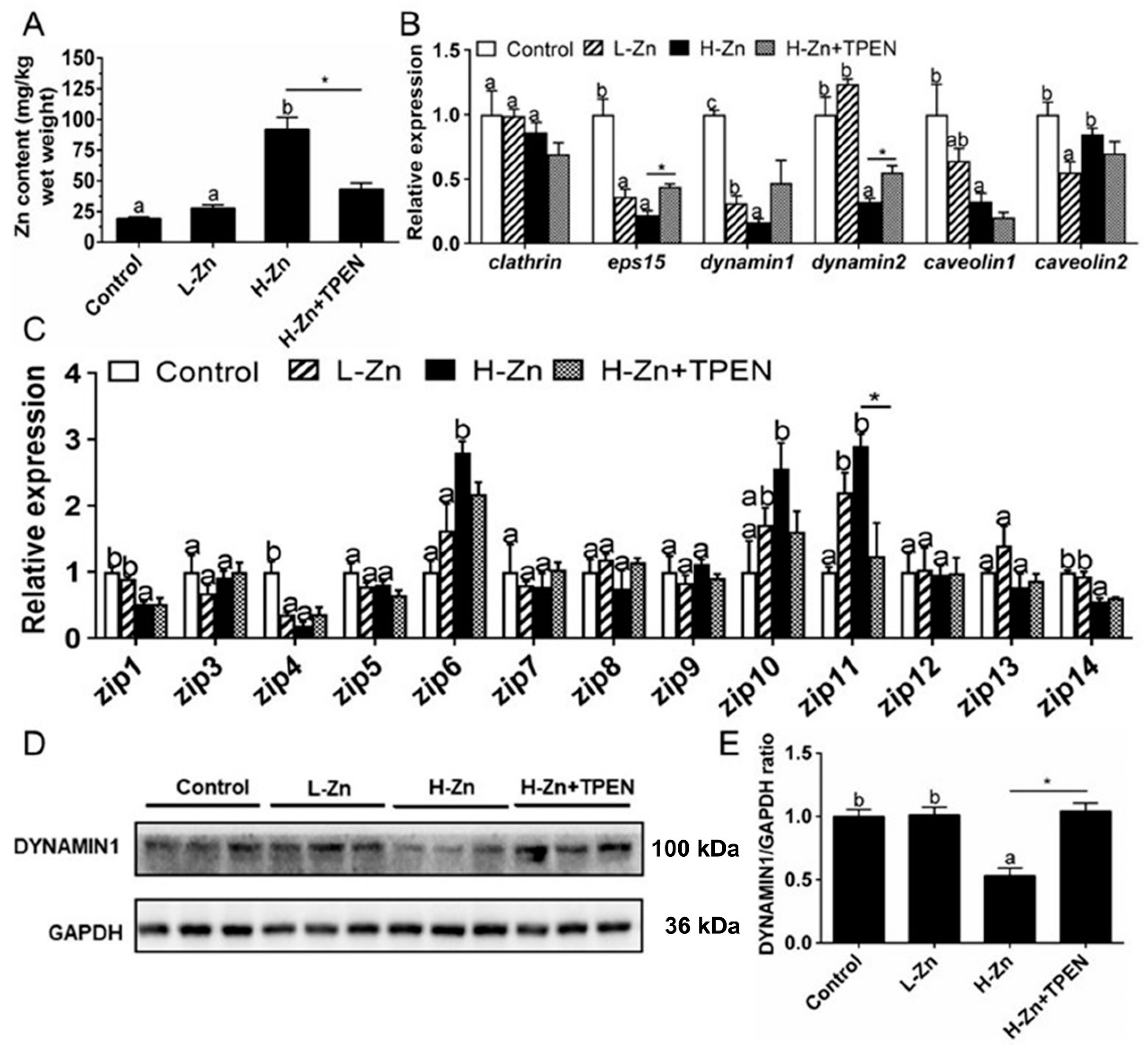
2.1.1. Zn Accumulation and Absorption in the Intestinal Tissues
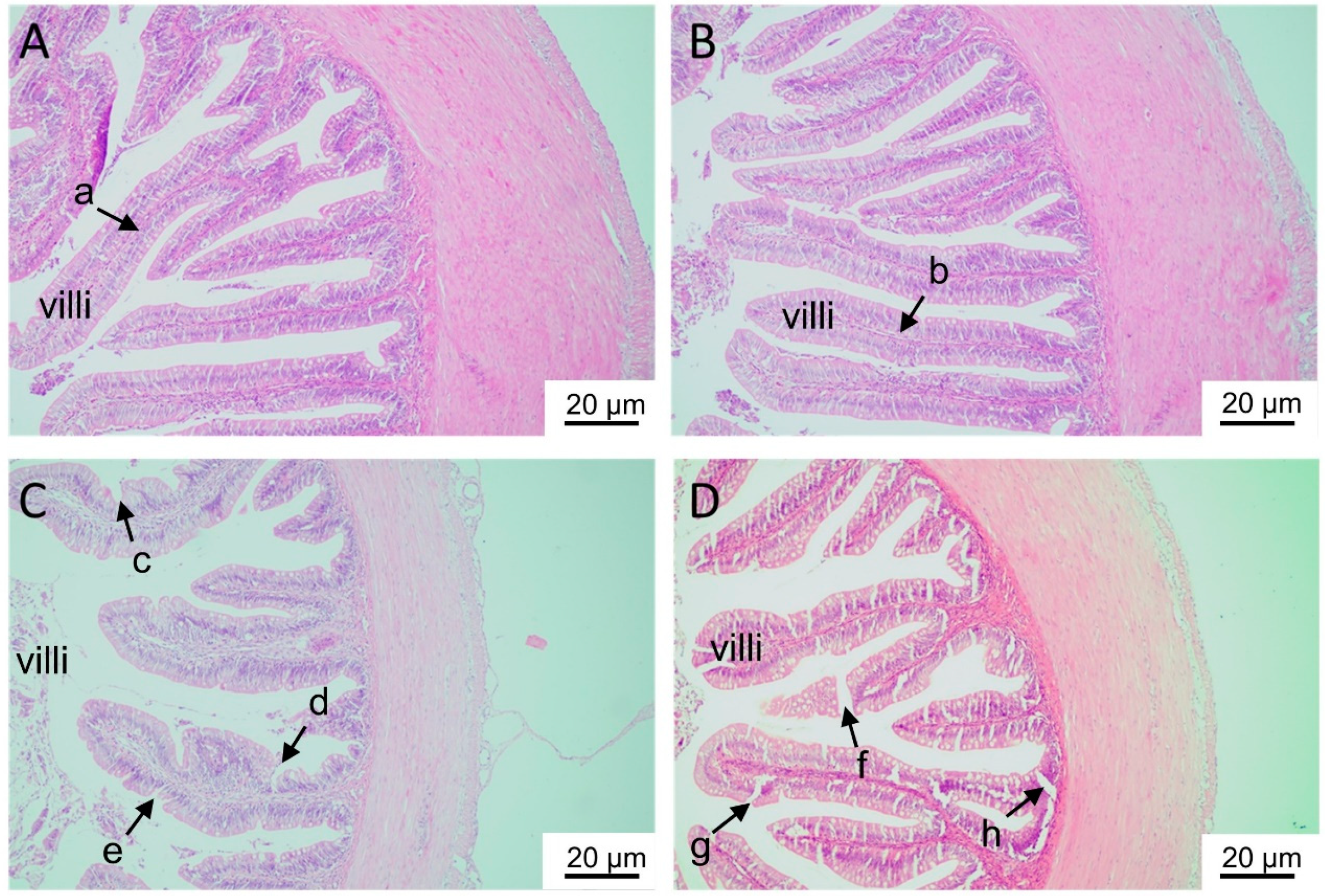
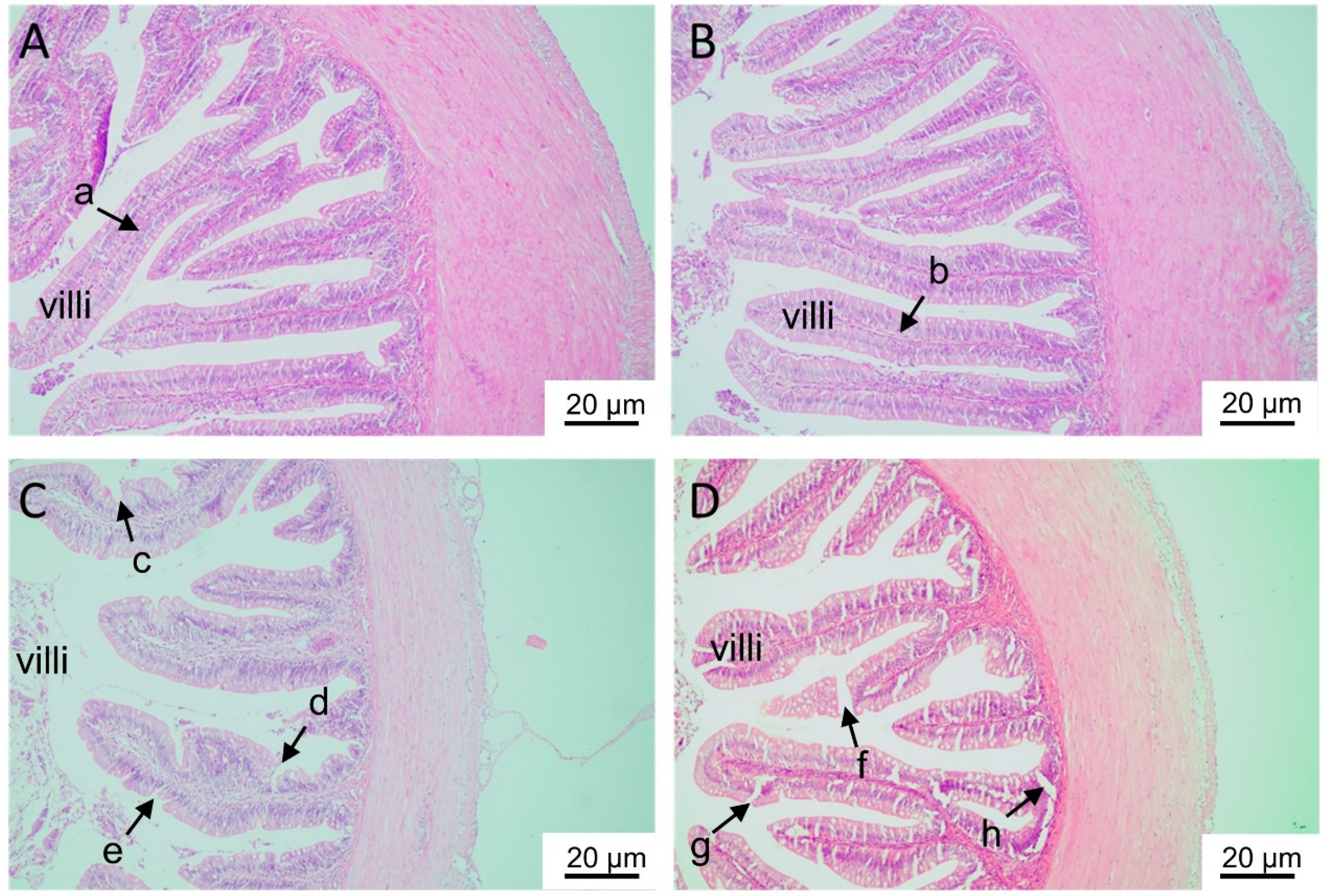
2.1.2. Histological Analyses
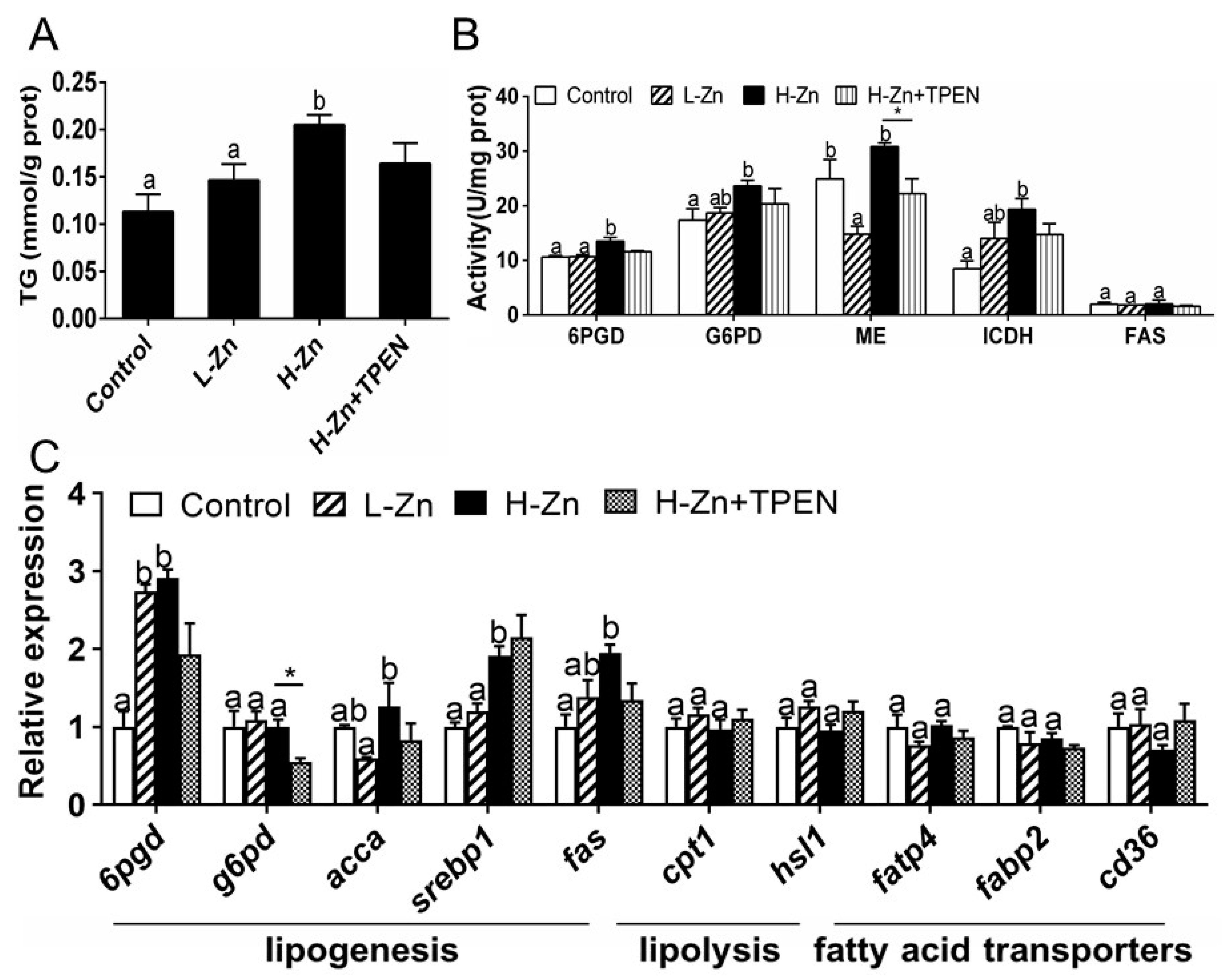
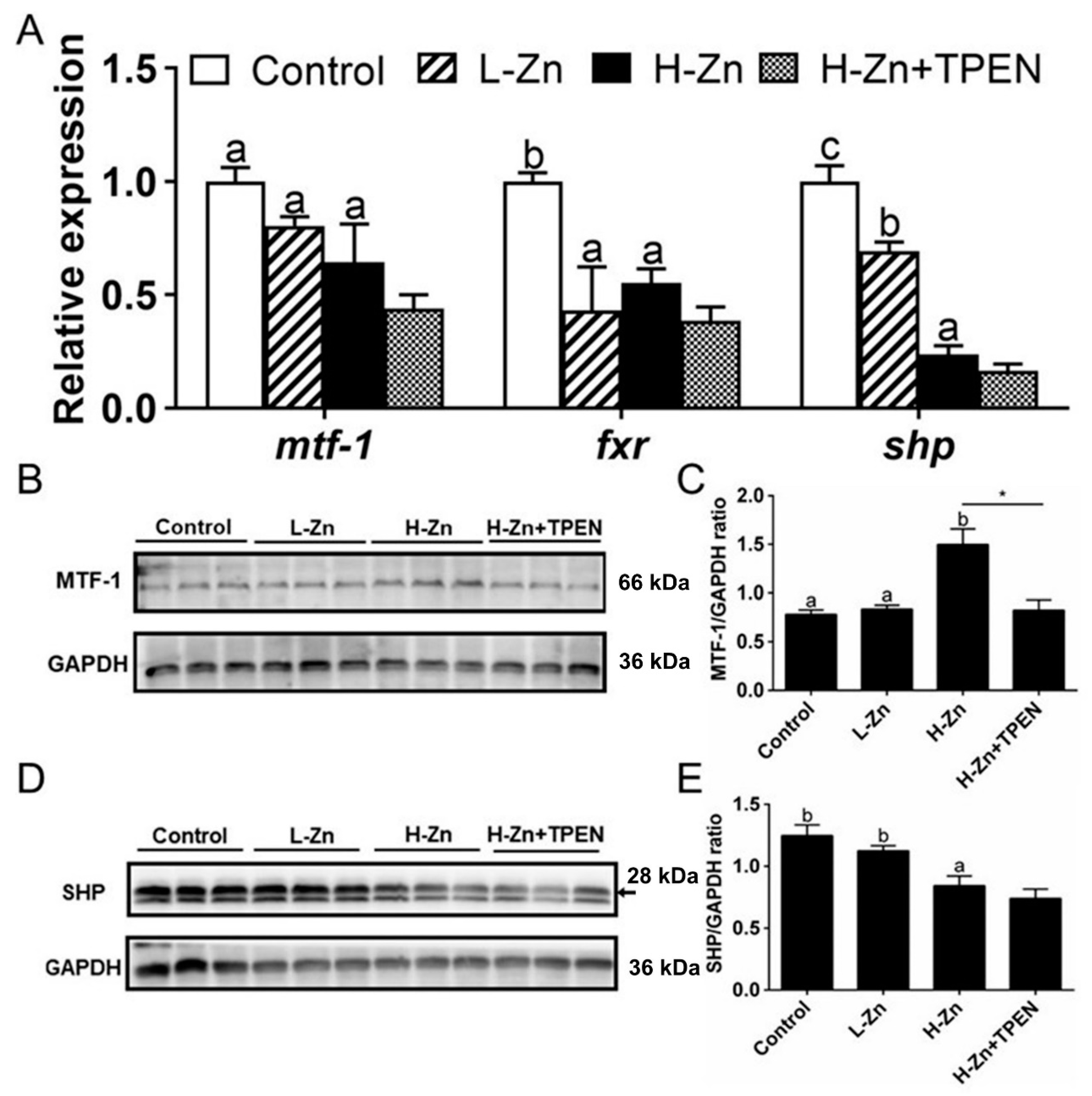
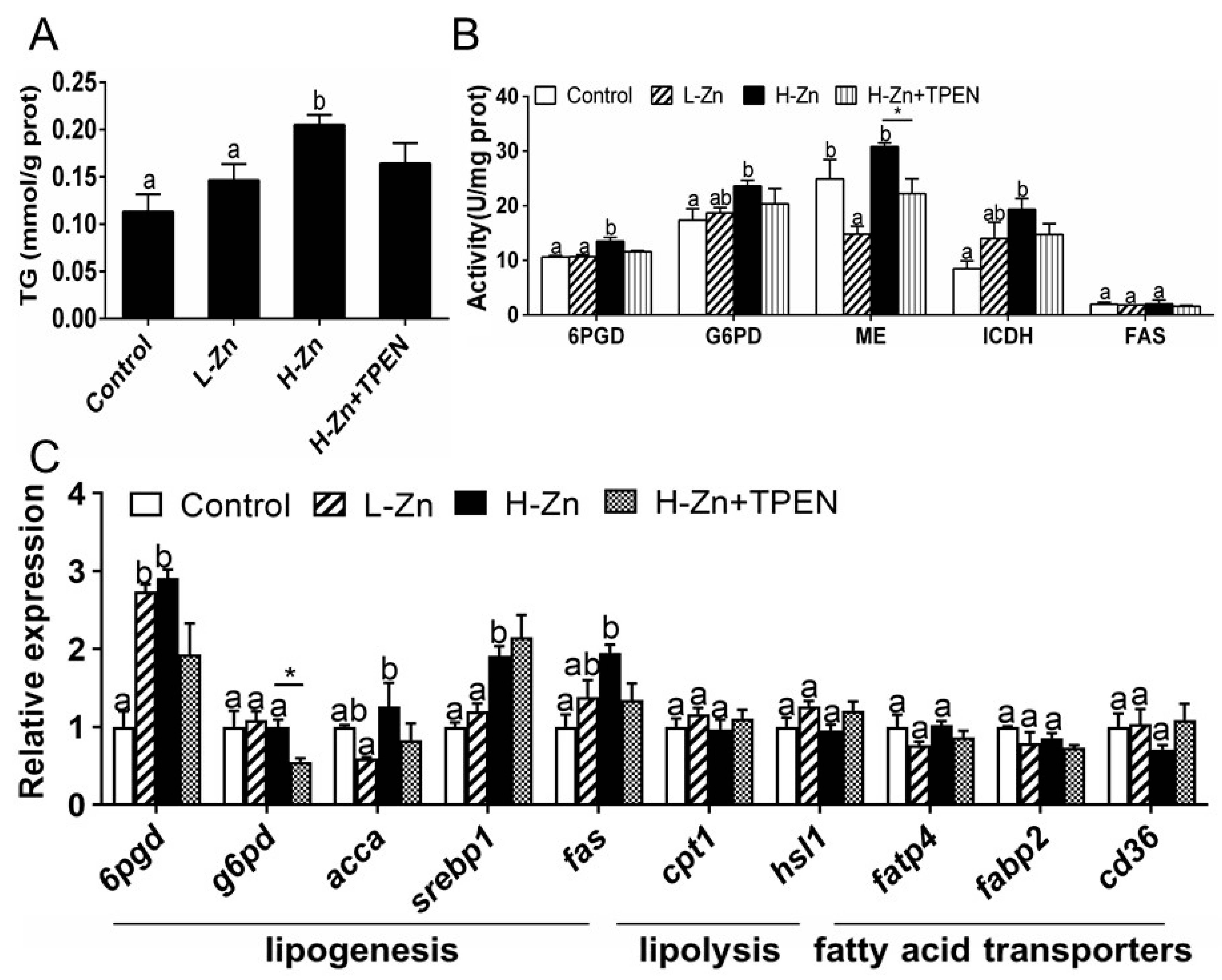
2.1.3. TG Content, Activities, and Expression of Enzymes and Genes Involved in Lipid Metabolism
2.2. In Vitro Assessment
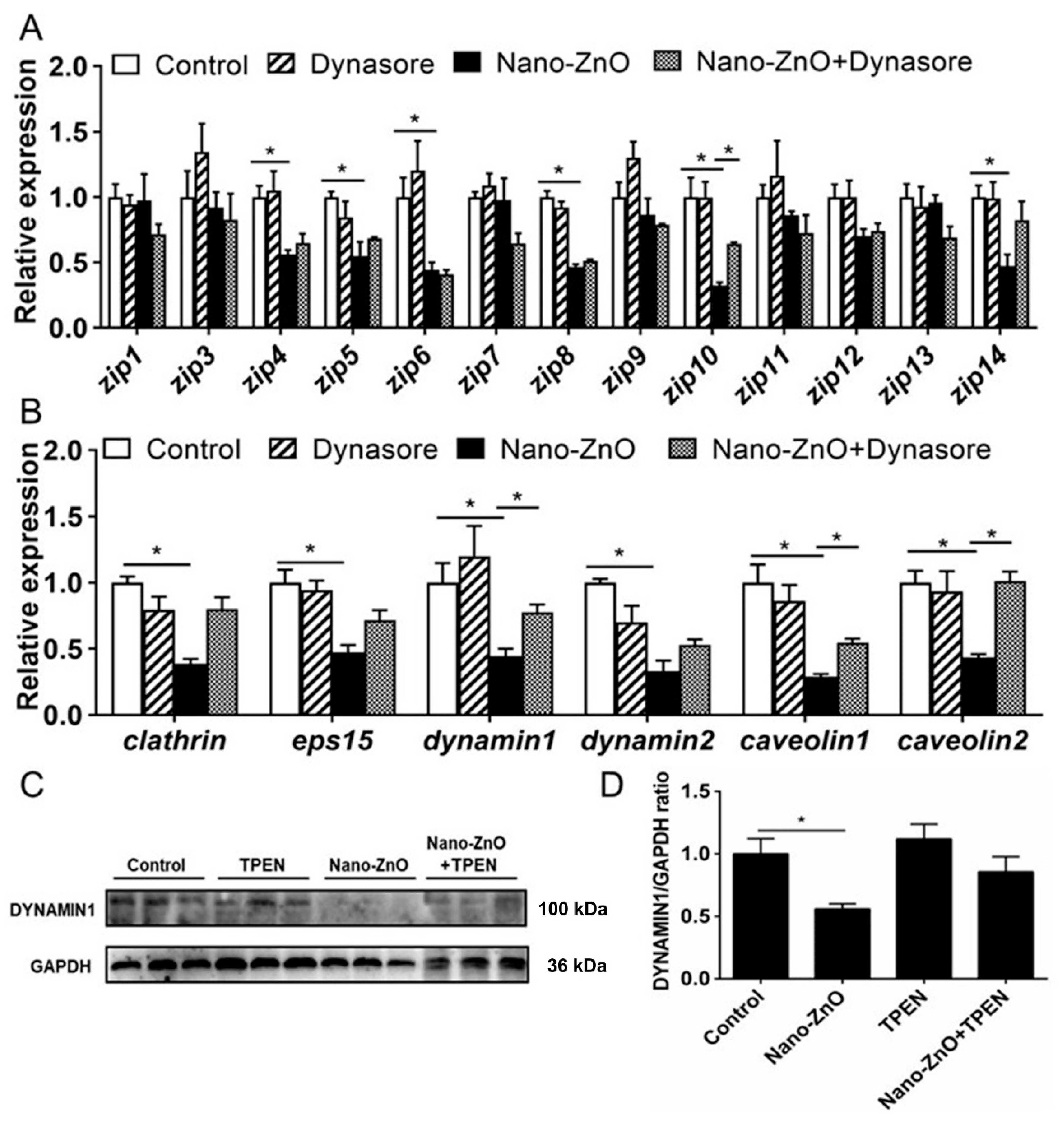
2.2.1. IECs Absorbed Nano-ZnO via Endocytosis Pathways
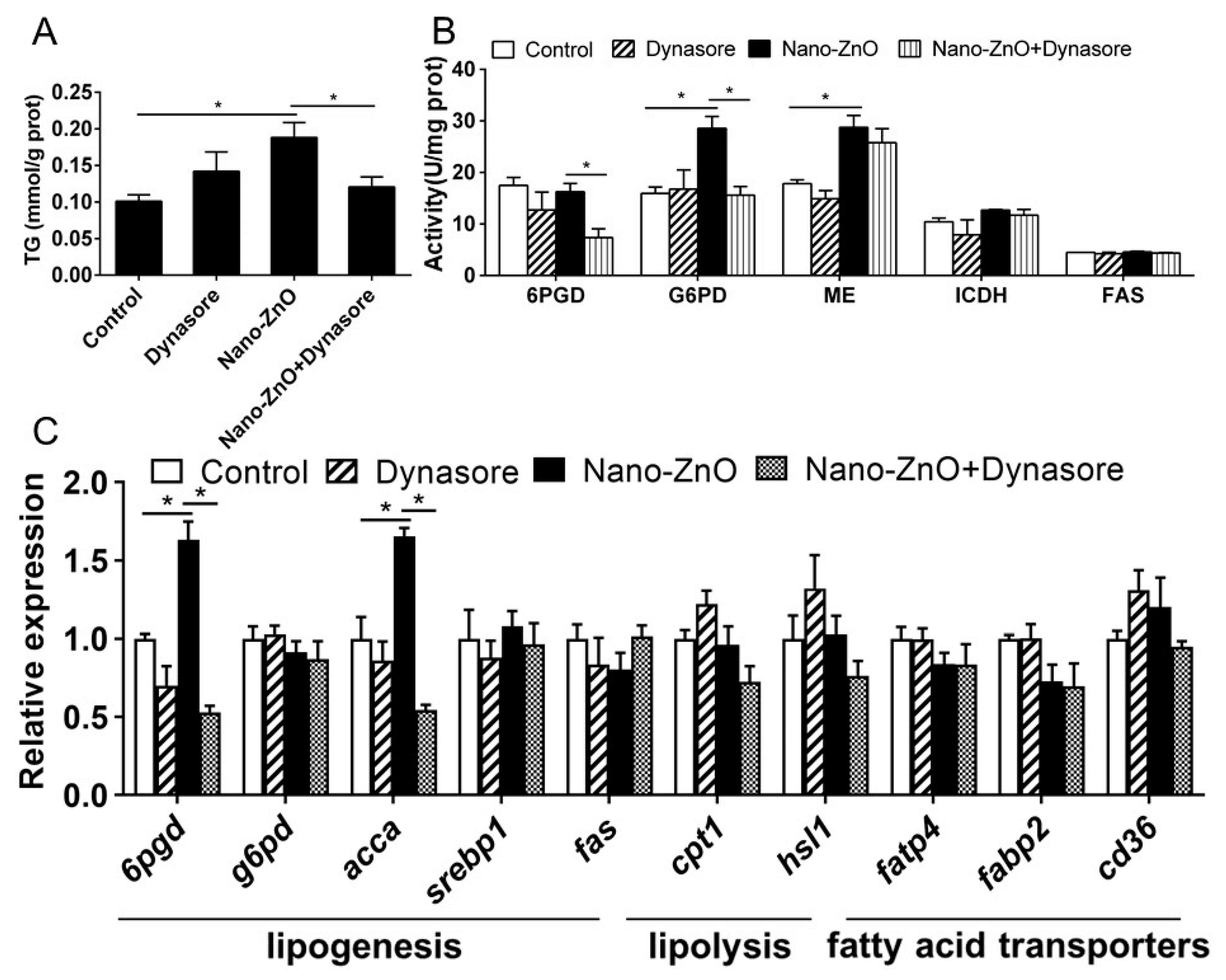
2.2.2. Nano-ZnO-Induced TG Deposition and Lipogenesis Was Partially Attributable to the Endocytosis of Nano-ZnO in IECs
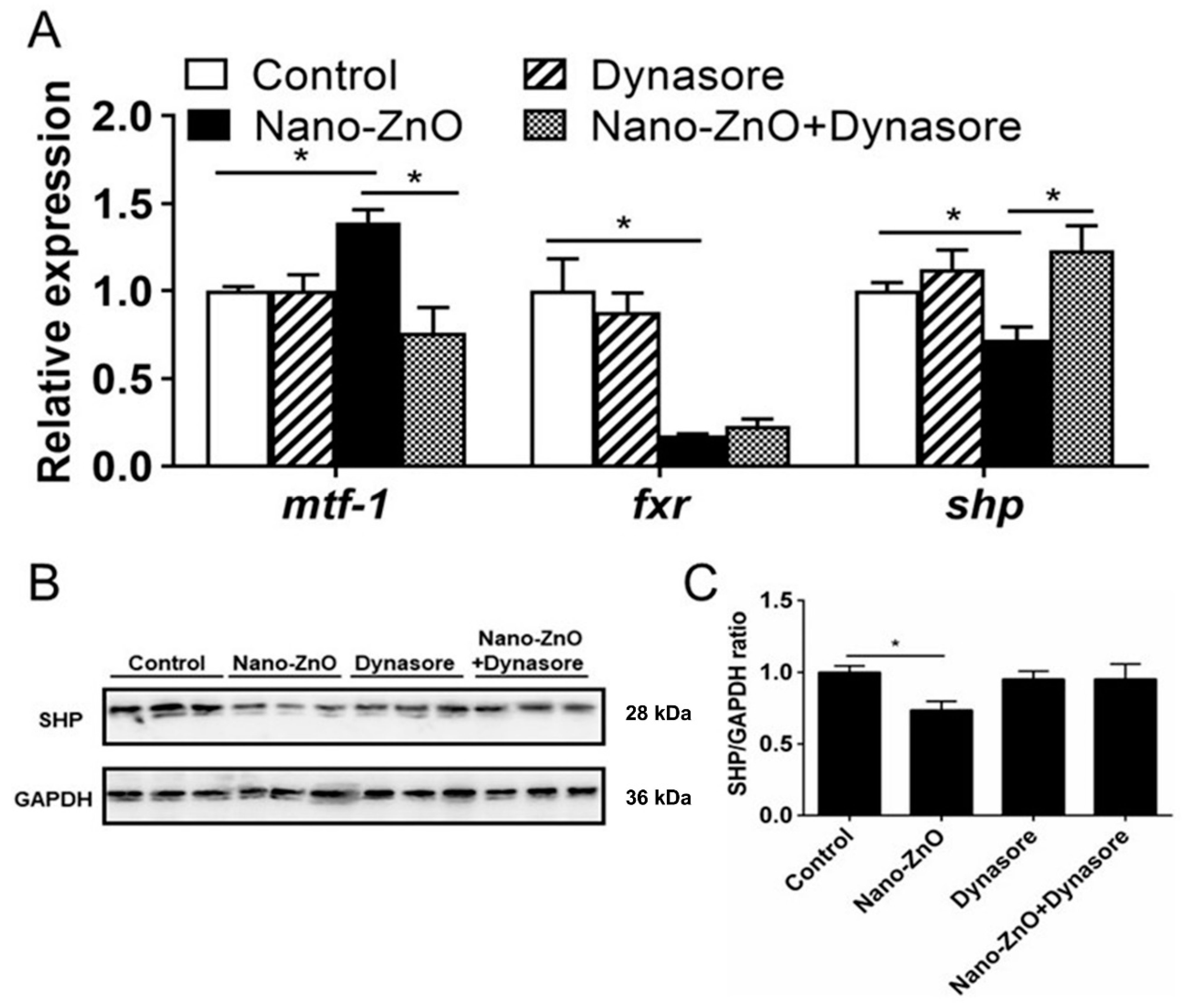
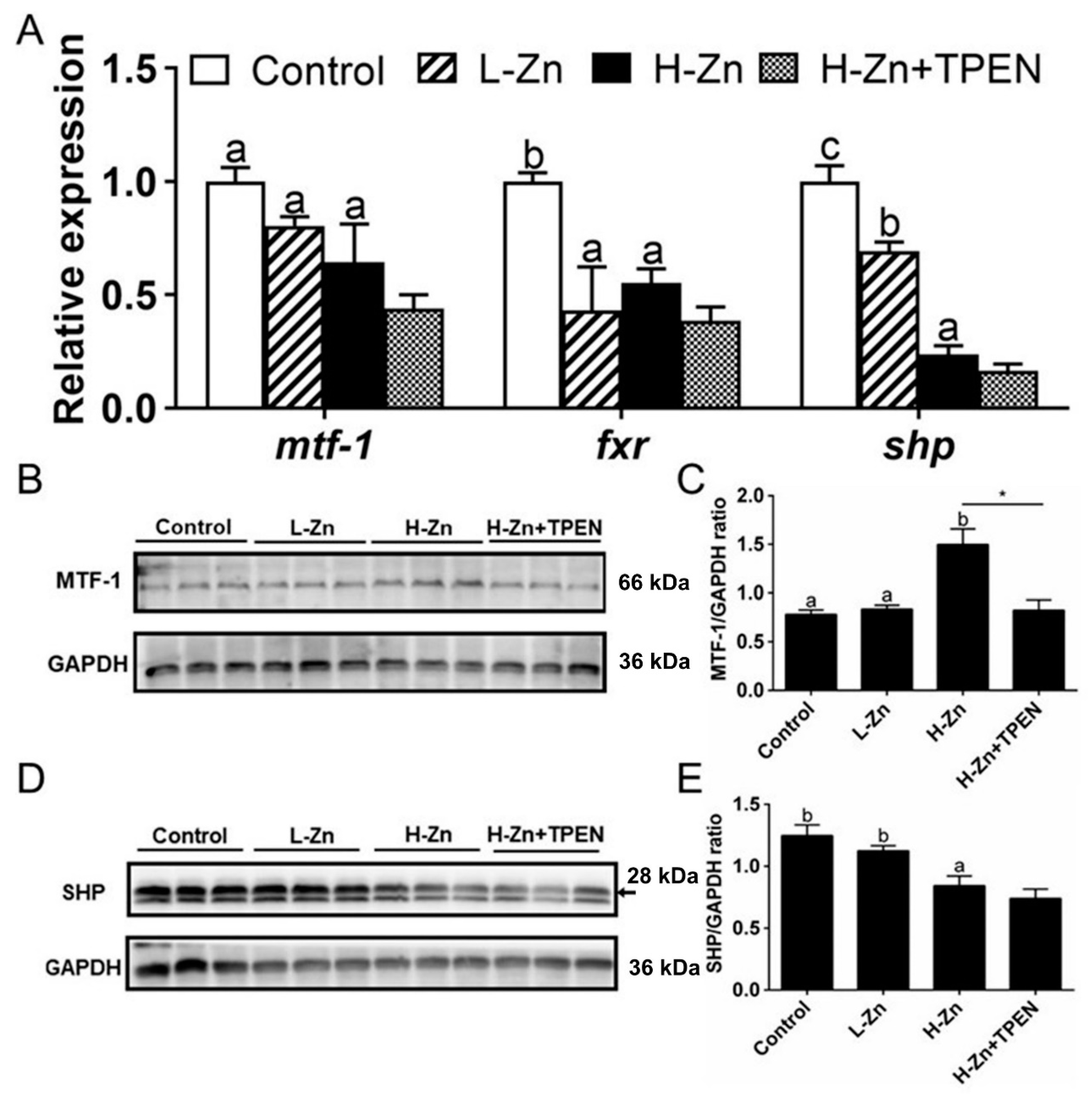
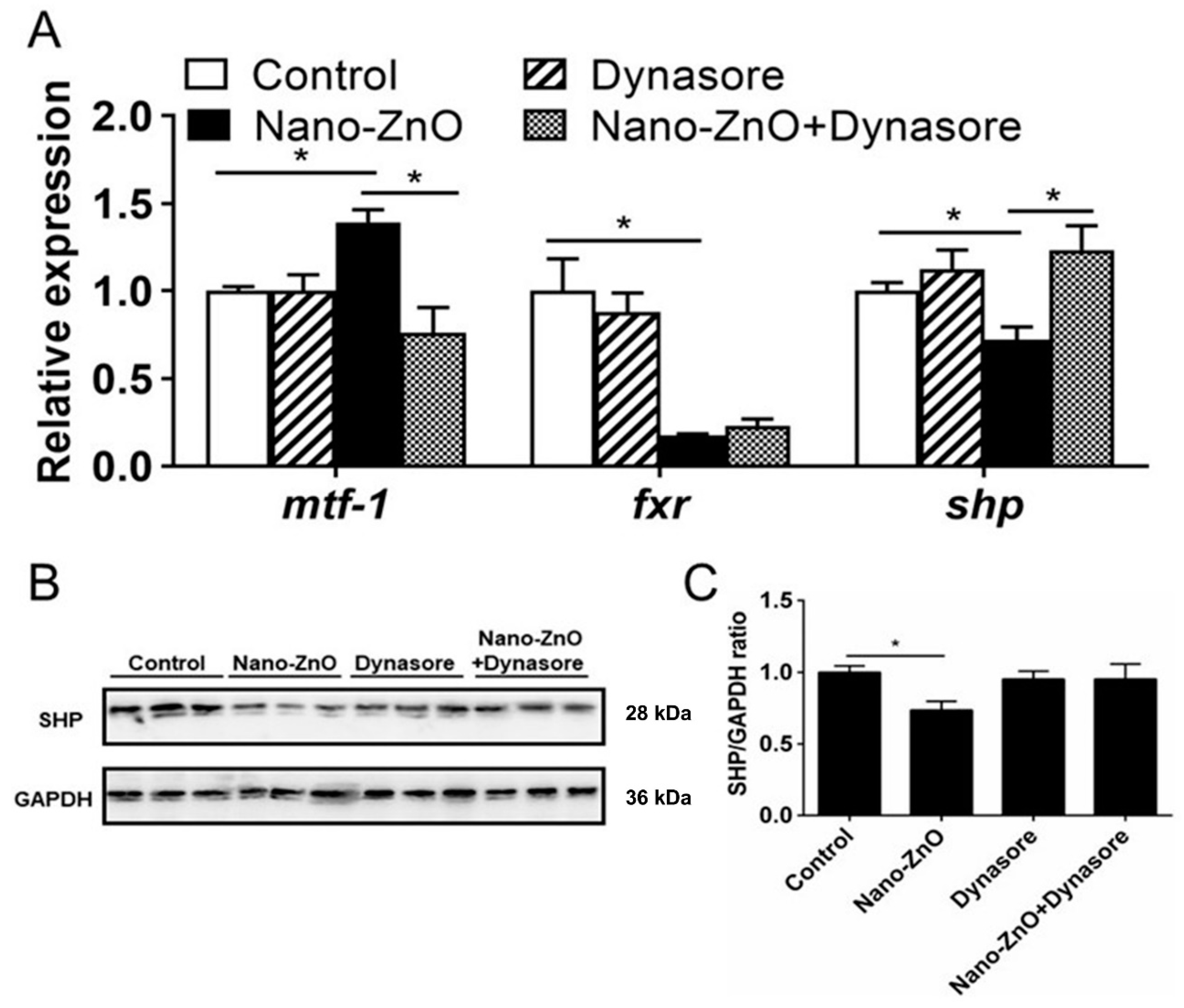
2.2.3. Nano-ZnO-Induced TG Deposition was Closely Related to the MTF-1-SHP Pathway
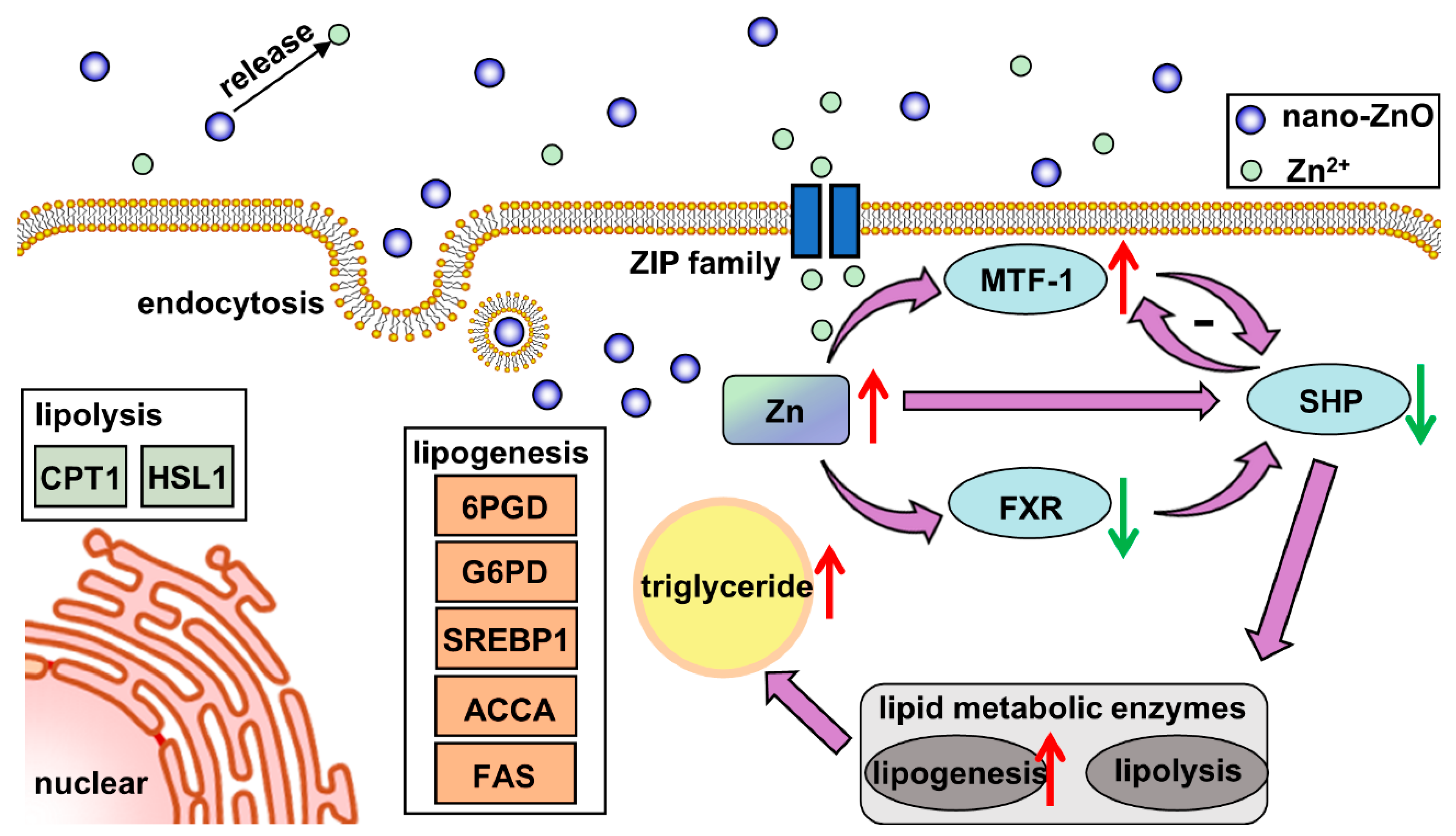
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. In Vivo Assessment
5.1.1. Fish Culture and Sampling
5.1.2. Zn and TG Contents
5.1.3. Histological Observation
5.1.4. Enzymatic Activity and Realtime Quantitative PCR (Q-PCR)
5.1.5. Western Blot (WB) Analysis
5.2. In Vitro Assessment
5.2.1. Enterocytes Culture and Treatments
5.2.2. TG Content, and Enzymatic Activity
5.2.3. Q-PCR and Western Blot Analysis
5.3. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, J.; Wang, A.; Wang, W.X. Evaluation of nano-ZnOs as a novel Zn source for marine fish: Importance of digestive physiology. Nanotoxicology 2017, 11, 1026–1039. [Google Scholar] [CrossRef]
- Boxall, A.; Chaudhry, Q.; Sinclair, C.; Jones, A.; Aitken, R.; Jefferson, B.; Watts, C. Current and Future Predicted Environmental Exposure to Engineered Nanoparticles; Central Science Laboratory: York, UK, 2007. [Google Scholar]
- Zhu, X.; Zhu, L.; Duan, Z.; Qi, R.; Li, Y.; Lang, Y. Comparative toxicity of several metal oxide nanoparticle aqueous suspensions to zebrafish (Danio rerio) early developmental stage. J. Environ. Sci. Health A Tox. Hazard. Subst. Environ. Eng. 2008, 43, 278–284. [Google Scholar] [CrossRef]
- Wong, S.W.; Leung, P.T.; Djurisic, A.B.; Leung, K.M. Toxicities of nano zinc oxide to five marine organisms: Influences of aggregate size and ion solubility. Anal. Bioanal. Chem. 2010, 396, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Taherian, S.M.R.; Hosseini, S.A.; Jafari, A.; Etminan, A.; Birjandi, M. Acute toxicity of zinc oxide nanoparticles from Satureja hortensis on rainbow trout (Oncorhynchus mykiss). Turk. J. Fish. Aquat. Sci. 2020, 20, 481–489. [Google Scholar] [CrossRef]
- Chupani, L.; Niksirat, H.; Lünsmann, V.; Haange, S.B.; von Bergen, M.; Jehmlich, N.; Zuskova, E. Insight into the modulation of intestinal proteome of juvenile common carp (Cyprinus carpio L.) after dietary exposure to ZnO nanoparticles. Sci. Total Environ. 2018, 613–614, 62–71. [Google Scholar] [CrossRef]
- Hogstrand, C. 3-Zinc. In Homeostasis and Toxicology of Essential Metals; Wood, C.M., Farrell, A.P., Brauner, C.J., Eds.; Academic Press: Waltham, MA, USA, 2011; Volume 31A, pp. 135–200. [Google Scholar]
- Tuomela, S.; Autio, R.; Buerki-Thurnherr, T.; Arslan, O.; Kunzmann, A.; Andersson-Willman, B.; Lahesmaa, R. Gene expression profiling of immune-competent human cells exposed to engineered zinc oxide or titanium dioxide nanoparticles. PLoS ONE 2013, 8, e68415. [Google Scholar] [CrossRef]
- Kambe, T.; Tsuji, T.; Hashimoto, A.; Itsumura, N. The physiological, biochemical, and molecular roles of zinc transporters in zinc homeostasis and metabolism. Physiol. Rev. 2015, 95, 749–784. [Google Scholar] [CrossRef]
- Oh, N.; Park, J.H. Endocytosis and exocytosis of nanoparticles in mammalian cells. Int. J. Nanomed. 2014, 9, 51. [Google Scholar]
- Tocher, D.R.; Bell, J.G.; Dick, J.R.; Crampton, V.O. Effects of dietary vegetable oil on Atlantic salmon hepatocyte fatty acid desaturase and liver fatty acid compositions. Lipids 2003, 38, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Ling, S.C.; Zhuo, M.Q.; Zhang, D.G.; Cui, H.Y.; Luo, Z. Nano-Zn increased Zn accumulation and triglyceride content by up-regulating lipogenesis in freshwater teleost, yellow catfish Pelteobagrus fulvidraco. Int. J. Mol. Sci. 2020, 21, 1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horton, J.D.; Goldstein, J.L.; Brown, M.S. SREBPs: Activators of the complete program of cholesterol and fatty acid synthesis in the liver. J. Clin. Investig. 2002, 109, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.C.; Xu, Y.H.; Zhao, T.; Wu, L.X.; Luo, Z. Waterborne Cu exposure increased lipid deposition and lipogenesis by affecting Wnt/β-catenin pathway and the β-catenin acetylation levels of grass carp Ctenopharyngodon idella. Environ. Pollut. 2020, 263, 114420. [Google Scholar] [CrossRef]
- Chen, N.; Wang, H.; Huang, Q.; Li, J.; Yan, J.; He, D.N.; Fan, C.H.; Song, H.Y. Long-term effects of nanoparticles on nutrition and metabolism. Small 2014, 10, 3603–3611. [Google Scholar] [CrossRef]
- Stahl, A.; Gimeno, R.E.; Tartaglia, L.A.; Lodish, H.F. Fatty acid transport proteins: A current view of a growing family. Trends Endocrinol. Metab. 2001, 12, 269–273. [Google Scholar] [CrossRef]
- Pohl, J.; Ring, A.; Hermaann, T.; Stremmel, W. Role of FATP in parenchymal cell fatty acid uptake. Biochim. Biophys. Acta. 2004, 1686, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bechmann, L.P.; Kocabayoglu, P.; Sowa, J.P.; Sydor, S.; Best, J.; Schlattjan, M.; Beilfuss, A.; Schmitt, J.; Hannivoort, R.A.; Kilicarslan, A.; et al. Free fatty acids repress small heterodimer partner (SHP) activation and adiponectin counteracts bile acid-induced liver injury in superobese patients with nonalcoholic steatohepatitis. Hepatology 2013, 57, 1394–1406. [Google Scholar] [CrossRef]
- Zou, A.; Magee, N.; Deng, F.; Lehn, S.; Zhong, C.; Zhang, Y. Hepatocyte nuclear receptor SHP suppresses inflammation and fibrosis in a mouse model of nonalcoholic steatohepatitis. J. Biol. Chem. 2018, 293, 8656–8671. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.; Zhao, T.; Hogstrand, C.; Xu, Y.C.; Ling, S.C.; Chen, G.H.; Luo, Z. FXR-mediated inhibition of autophagy contributes to FA-induced TG accumulation and accordingly reduces FA-induced lipotoxicity. Cell Commun. Signal. 2020, 18, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Andrews, G.K.; Wang, L. Zinc-induced Dnmt1 expression involves antagonism between MTF-1 and nuclear receptor SHP. Nucleic Acids Res. 2012, 40, 4850–4860. [Google Scholar] [CrossRef] [Green Version]
- Wei, C.C.; Luo, Z.; Hogstrand, C.; Xu, Y.H.; Wu, L.X.; Chen, G.H.; Song, Y.F. Zinc reduces hepatic lipid deposition and activates lipophagy via Zn2+/MTF-1/PPARα and Ca2+/CaMKKβ/AMPK pathways. FASEB J. 2018, 32, 6666–6680. [Google Scholar] [CrossRef] [PubMed]
- Gong, G.; Dan, C.; Xiao, S.; Guo, W.; Huang, P.; Xiong, Y.; Wu, J.; He, Y.; Zhang, J.; Li, X.; et al. Chromosomal-level assembly of yellow catfish genome using third-generation DNA sequencing and Hi-C analysis. Gigascience 2018, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.W.; Wu, K.; Lv, W.H.; Song, C.C.; Luo, Z. Molecular characterization of ten zinc (Zn) transporter genes and their regulation to Zn metabolism in freshwater teleost yellow catfish Pelteobagrus fulvidraco. J. Trace Elem. Med. Biol. 2020, 59, 126433. [Google Scholar] [CrossRef] [PubMed]
- Macia, E.; Ehrlich, M.; Massol, R.; Boucrot, E.; Brunner, C.; Kirchhausen, T. Dynasore, a cell-permeable inhibitor of dynamin. Dev. Cell 2006, 10, 839–850. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Jadhav, H.R.; Bhatt, T. Dynamin functions and ligands: Classical mechanisms behind. Mol. Pharmacol. 2017, 91, 123–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, S.; Cao, H.; Chen, J.; McNiven, M. Eps15 mediates vesicle trafficking from the trans-Golgi-network via an interaction with the clatherin adaptor Ap-1. Mol. Biol. Cell 2008, 19, 3564–3575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doherty, G.J.; McMahon, H.T. Mechanisms of endocytosis. Ann. Rev. Biochem. 2009, 78, 857–902. [Google Scholar] [CrossRef] [Green Version]
- Jenkitkasemwong, S.; Wang, C.Y.; Mackenzie, B.; Knutson, M.D. Physiologic implications of metal-ion transport by ZIP14 and ZIP8. Biometals 2012, 25, 643–655. [Google Scholar] [CrossRef] [Green Version]
- Ho, E.; Dukovic, S.; Hobson, B.; Wong, C.P.; Miller, G.; Hardin, K.; Traberab, M.G.; Tanguay, R.L. Zinc transporter expression in zebrafish (Danio rerio) during development. Comp. Biochem. Physiol. Part C 2012, 155, 26–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, T.; Zhang, S.H.; Zhang, C.; Zhang, J.L.; Yang, F.; Wang, G.Y.; Yang, Z.J.; Bai, D.Y.; Shi, Y.Y.; Liu, T.Q.; et al. The effects of 50 nm unmodified nano-ZnO on lipid metabolism and semen quality in male mice. Biol. Trace Elem. Res. 2020, 194, 432–442. [Google Scholar] [CrossRef]
- Zheng, J.L.; Luo, Z.; Hu, W.; Liu, C.X.; Chen, Q.L.; Zhu, Q.L.; Gong, Y. Different effects of dietary Zn deficiency and excess on lipid metabolism in yellow catfish Pelteobagrus fulvidraco. Aquaculture 2015, 435, 10–17. [Google Scholar] [CrossRef]
- Esmaeillou, M.; Moharamnejad, M.; Hsankhani, R.; Tehrani, A.A.; Maadi, H. Toxicity of ZnO nanoparticles in healthy adult mice. Environ. Toxicol. Pharmacol. 2013, 35, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Anakk, S.; Watanabe, M.; Ochsner, S.A.; McKenna, N.J.; Finegold, M.J.; Moore, D.D. Combined deletion of Fxr and Shp in mice induces Cyp17a1 and results in juvenile onset cholestasis. J. Clin. Investig. 2011, 121, 86–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, M.; Houten, S.M.; Wang, L.; Moschetta, A.; Mangelsdorf, D.J.; Heyman, R.A.; Auwerx, J. Bile acids lower triglyceride levels via a pathway involving FXR, SHP, and SREBP-1c. J. Clin. Investig. 2004, 113, 1408–1418. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Tan, X.Y.; Zheng, J.L.; Chen, Q.L.; Liu, C.X. Quantitative dietary zinc requirement of juvenile yellow catfish Pelteobagrus fulvidraco, and effects on hepatic intermediary metabolism and antioxidant responses. Aquaculture 2011, 319, 150–155. [Google Scholar] [CrossRef]
- Wu, K.; Luo, Z.; Hogstrand, C.; Chen, G.H.; Wei, C.C.; Li, D.D. Zn stimulates the phospholipids biosynthesis via the pathways of oxidative and endoplasmic reticulum stress in the intestine of freshwater teleost yellow catfish. Environ. Sci. Technol. 2018, 52, 9206–9214. [Google Scholar] [CrossRef] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by genometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.H.; Luo, Z.; Chen, F.; Shi, X.; Song, Y.F.; You, W.J.; Liu, X. PPARα, PPARγ and SREBP-1 pathways mediated waterborne iron (Fe)-induced reduction in hepatic lipid deposition of javelin goby Synechogobius hasta. Comp. Biochem. Physiol. Part C 2017, 197, 8–18. [Google Scholar] [CrossRef]
- Song, Y.F.; Hogstrand, C.; Wei, C.C.; Wu, K.; Pan, Y.X.; Luo, Z. Endoplasmic reticulum (ER) stress and cAMP/PKA pathway mediated Zn-induced hepatic lipolysis. Environ. Pollut. 2017, 228, 256–264. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.-W.; Lv, W.-H.; Wu, K.; Chen, G.-H.; Chen, F.; Song, C.-C.; Luo, Z. Dietary Nano-ZnO Is Absorbed via Endocytosis and ZIP Pathways, Upregulates Lipogenesis, and Induces Lipotoxicity in the Intestine of Yellow Catfish. Int. J. Mol. Sci. 2021, 22, 12047. https://doi.org/10.3390/ijms222112047
Chen S-W, Lv W-H, Wu K, Chen G-H, Chen F, Song C-C, Luo Z. Dietary Nano-ZnO Is Absorbed via Endocytosis and ZIP Pathways, Upregulates Lipogenesis, and Induces Lipotoxicity in the Intestine of Yellow Catfish. International Journal of Molecular Sciences. 2021; 22(21):12047. https://doi.org/10.3390/ijms222112047
Chicago/Turabian StyleChen, Shu-Wei, Wu-Hong Lv, Kun Wu, Guang-Hui Chen, Fang Chen, Chang-Chun Song, and Zhi Luo. 2021. "Dietary Nano-ZnO Is Absorbed via Endocytosis and ZIP Pathways, Upregulates Lipogenesis, and Induces Lipotoxicity in the Intestine of Yellow Catfish" International Journal of Molecular Sciences 22, no. 21: 12047. https://doi.org/10.3390/ijms222112047
APA StyleChen, S.-W., Lv, W.-H., Wu, K., Chen, G.-H., Chen, F., Song, C.-C., & Luo, Z. (2021). Dietary Nano-ZnO Is Absorbed via Endocytosis and ZIP Pathways, Upregulates Lipogenesis, and Induces Lipotoxicity in the Intestine of Yellow Catfish. International Journal of Molecular Sciences, 22(21), 12047. https://doi.org/10.3390/ijms222112047