
Not All Lectins Are Equally Suitable for Labeling Rodent Vasculature

 , , ,
, , ,
Abstract
1. Introduction
2. Results
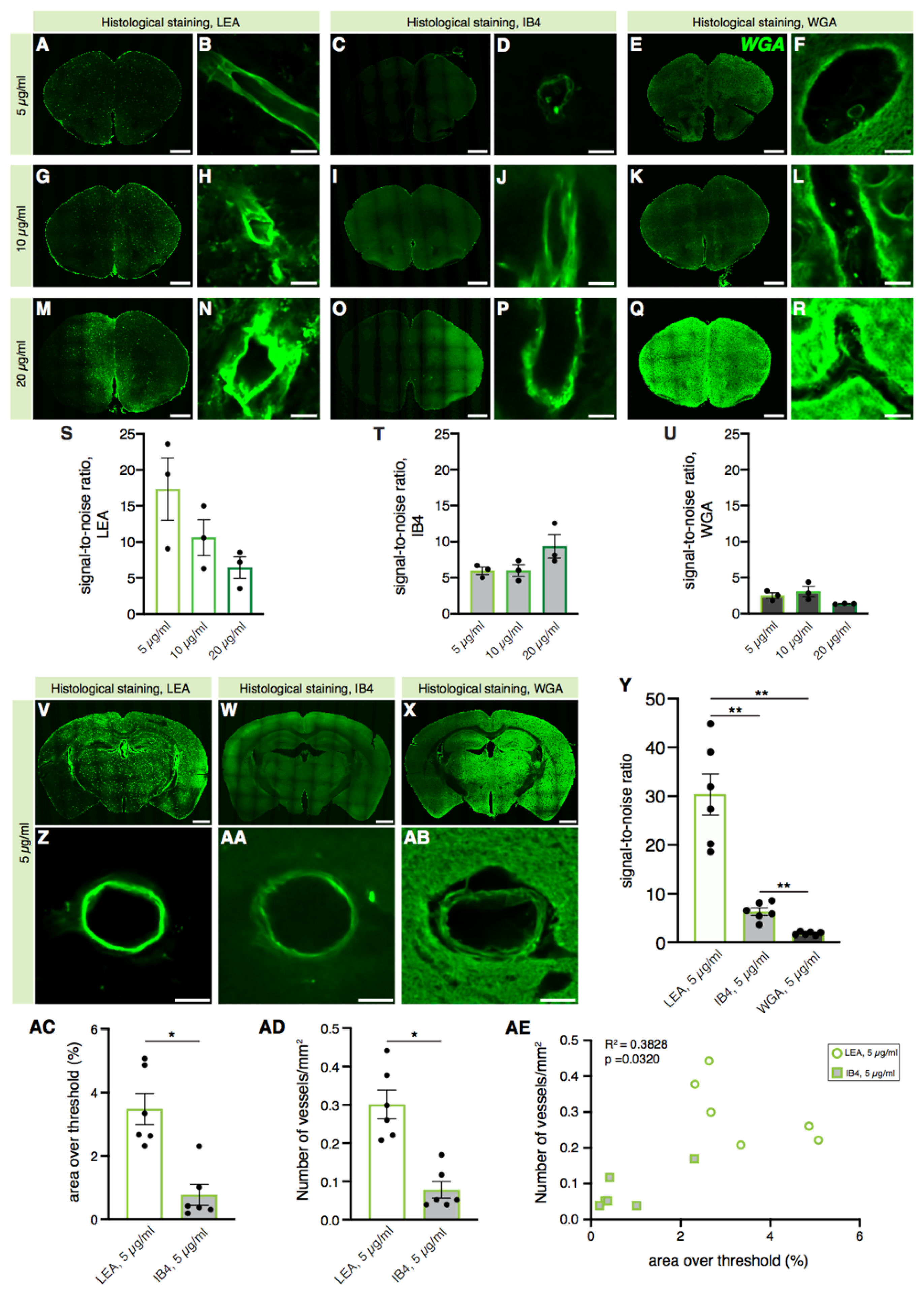
2.1. Histological Staining of Cerebral Blood Vessels with the LEA Lectin Gives a Better Signal-to-Noise Ratio than WGA and IB4 Lectins
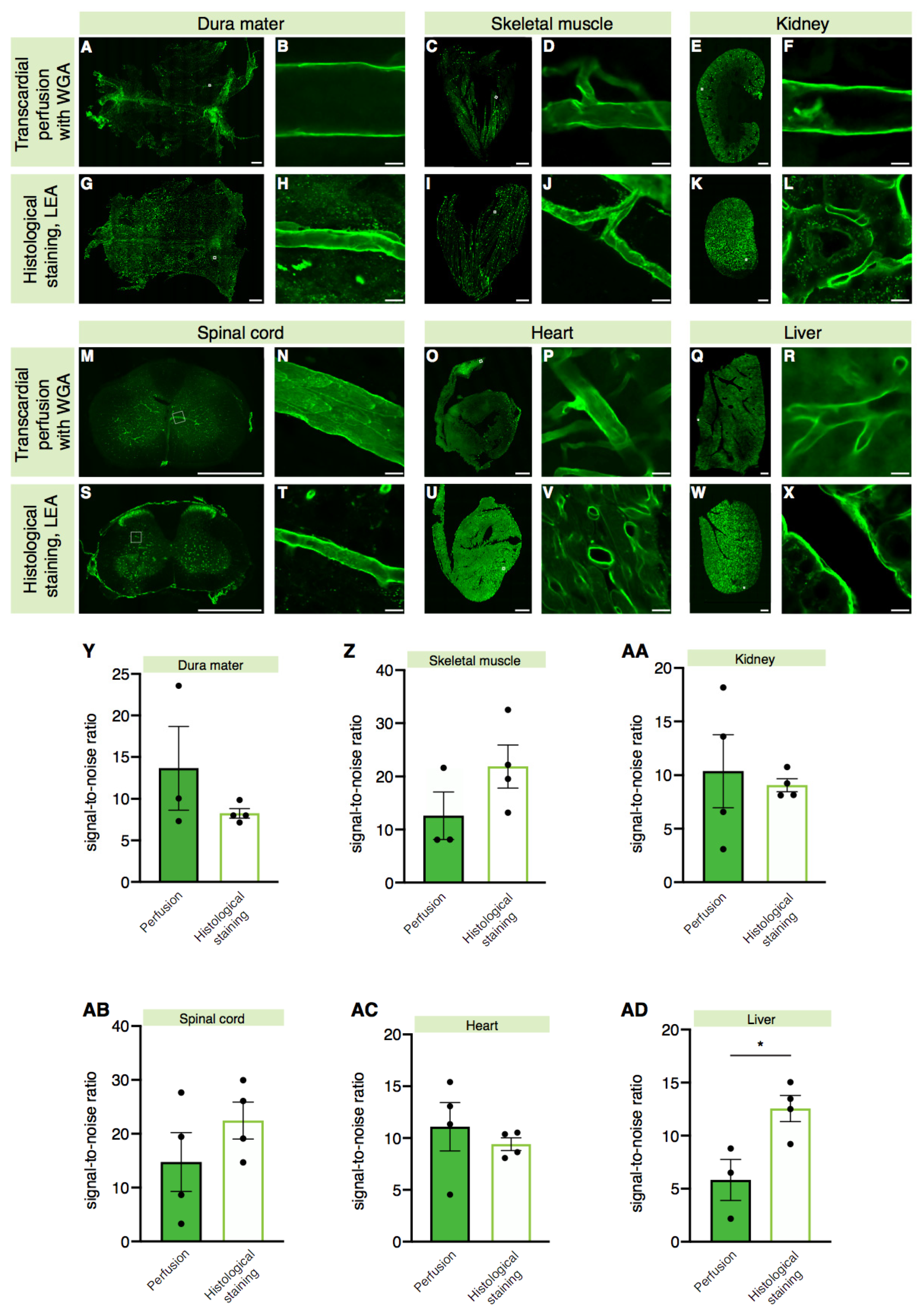
2.2. A Higher Signal-to-Noise Ratio Can Be Achieved by Intracardiac Perfusion of Lectins
2.3. Lectin Is Widely Applicable for Labeling Blood Vessels in Various Tissues
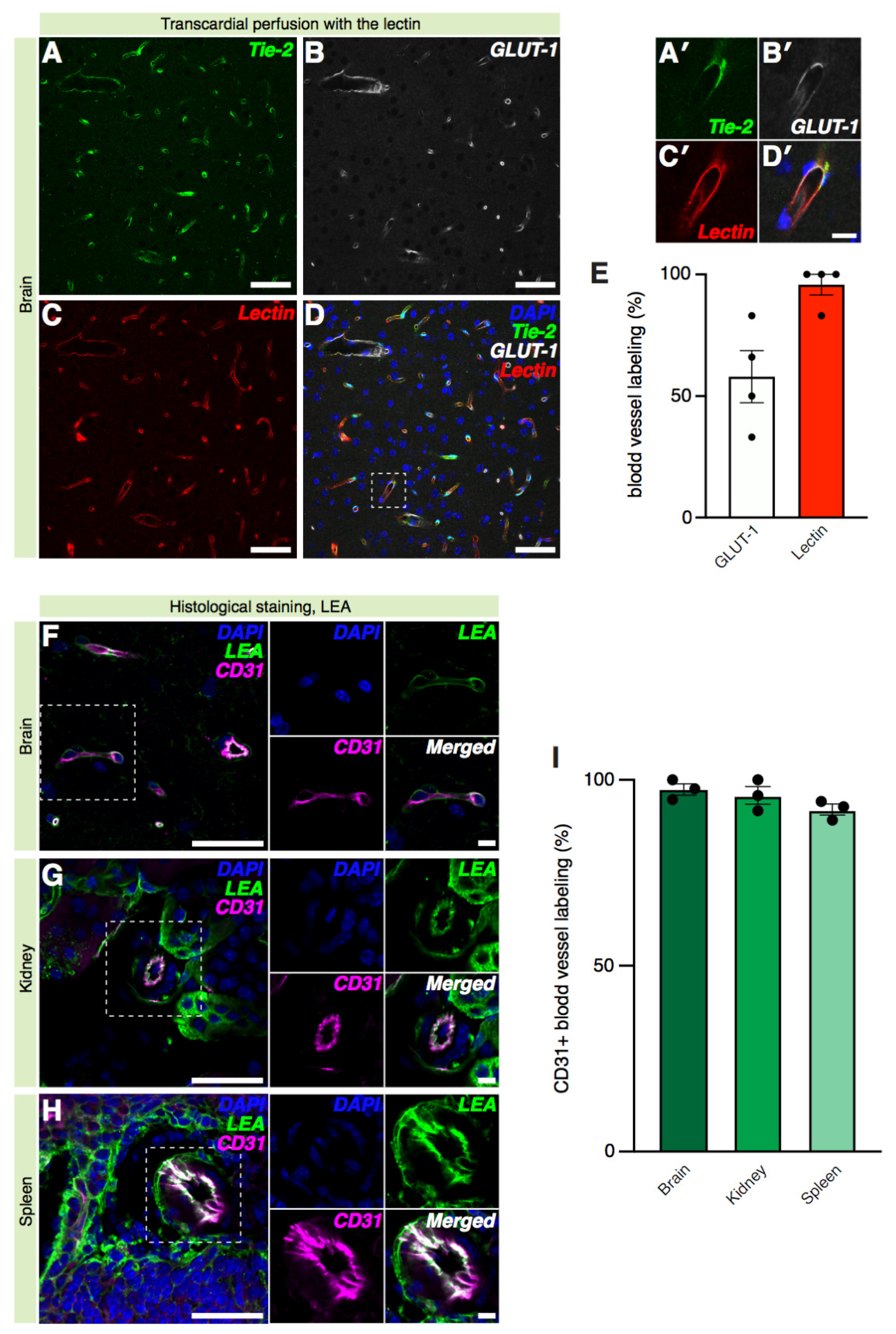
2.4. Lectin Labeling of Cerebral Blood Vessels Is Equivalent to Reporter Mice and Immunostaining
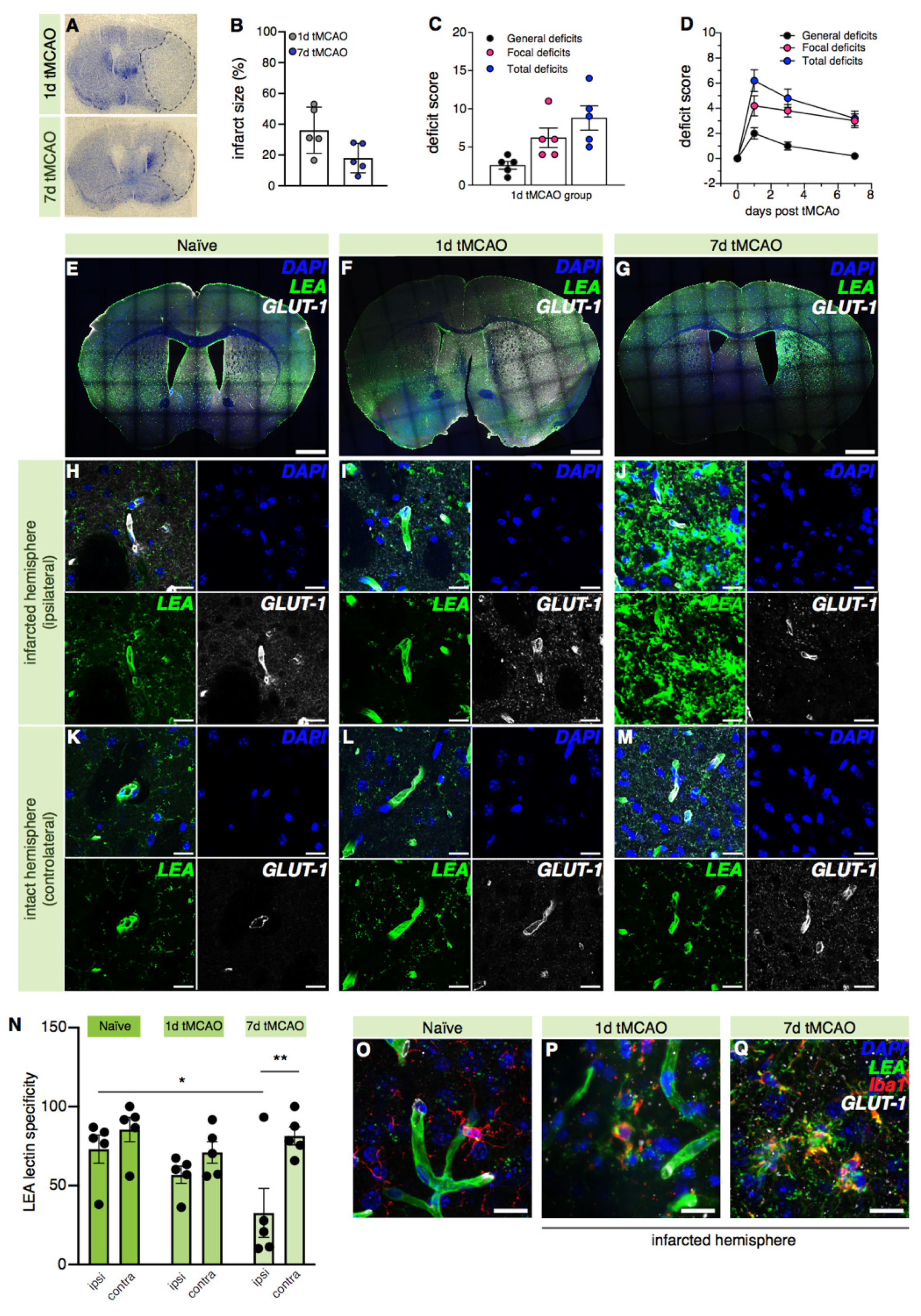
2.5. LEA Lectin’s Specificity for Blood Vessel Labeling Decreases in the Ischemic Brain
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Supplies
4.3. Transient Middle Cerebral Artery Occlusion (tMCAo)
4.4. Neuroscore Determination in Mice after tMCAo
4.5. Nissl Staining for Infarct Size Determination in Mice after tMCAo
4.6. Histological Labeling of Coronal Tissue Sections of C57BL/6 Mice with the IB4, LEA, and WGA Lectins
4.7. WGA Lectin Perfusion of C57BL/6 Mice
4.8. GLUT-1, Iba1, CD68, GFAP, CD31, α-SMA Immunostaining, and Biotin Revelation
4.9. Image Acquisition and Analysis
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aird, W.C. Discovery of the cardiovascular system: From Galen to William Harvey. J. Thromb. Haemost. 2011, 9, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Risau, W. Mechanisms of angiogenesis. Nature 1997, 386, 671–674. [Google Scholar] [CrossRef]
- Brown, W.R.; Thore, C.R. Review: Cerebral microvascular pathology in ageing and neurodegeneration. Neuropathol. Appl. Neurobiol. 2011, 37, 56–74. [Google Scholar] [CrossRef]
- Boero, J.A.; Ascher, J.; Arregui, A.; Rovainen, C.; Woolsey, T.A. Increased brain capillaries in chronic hypoxia. J. Appl. Physiol. 1999, 86, 1211–1219. [Google Scholar] [CrossRef]
- He, Q.W.; Li, Q.; Jin, H.J.; Zhi, F.; Suraj, B.; Zhu, Y.Y.; Xia, Y.P.; Mao, L.; Chen, X.L.; Hu, B. MiR-150 Regulates Poststroke Cerebral Angiogenesis via Vascular Endothelial Growth Factor in Rats. CNS Neurosci. Ther. 2016, 22, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Hayward, N.M.; Immonen, R.; Tuunanen, P.I.; Ndode-Ekane, X.E.; Grohn, O.; Pitkanen, A. Association of chronic vascular changes with functional outcome after traumatic brain injury in rats. J. Neurotrauma 2010, 27, 2203–2219. [Google Scholar] [CrossRef] [PubMed]
- Barcia, C.; Bautista, V.; Sanchez-Bahillo, A.; Fernandez-Villalba, E.; Faucheux, B.; Poza y Poza, M.; Barreiro, A.F.; Hirsch, E.C.; Herrero, M.T. Changes in vascularization in substantia nigra pars compacta of monkeys rendered parkinsonian. J. Neural Transm. 2005, 112, 1237–1248. [Google Scholar] [CrossRef] [PubMed]
- Ohlin, K.E.; Francardo, V.; Lindgren, H.S.; Sillivan, S.E.; O’Sullivan, S.S.; Luksik, A.S.; Vassoler, F.M.; Lees, A.J.; Konradi, C.; Cenci, M.A. Vascular endothelial growth factor is upregulated by L-dopa in the parkinsonian brain: Implications for the development of dyskinesia. Brain 2011, 134, 2339–2357. [Google Scholar] [CrossRef] [PubMed]
- Black, J.E.; Polinsky, M.; Greenough, W.T. Progressive failure of cerebral angiogenesis supporting neural plasticity in aging rats. Neurobiol. Aging 1989, 10, 353–358. [Google Scholar] [CrossRef]
- Hase, Y.; Ding, R.; Harrison, G.; Hawthorne, E.; King, A.; Gettings, S.; Platten, C.; Stevenson, W.; Craggs, L.J.L.; Kalaria, R.N. White matter capillaries in vascular and neurodegenerative dementias. Acta Neuropathol. Commun. 2019, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Muller, A.M.; Hermanns, M.I.; Skrzynski, C.; Nesslinger, M.; Muller, K.M.; Kirkpatrick, C.J. Expression of the endothelial markers PECAM-1, vWf, and CD34 in vivo and in vitro. Exp. Mol. Pathol. 2002, 72, 221–229. [Google Scholar] [CrossRef]
- Bell, R.D.; Winkler, E.A.; Sagare, A.P.; Singh, I.; LaRue, B.; Deane, R.; Zlokovic, B.V. Pericytes control key neurovascular functions and neuronal phenotype in the adult brain and during brain aging. Neuron 2010, 68, 409–427. [Google Scholar] [CrossRef] [PubMed]
- Koonce, N.A.; Griffin, R.J.; Dings, R.P.M. Galectin-1 Inhibitor OTX008 Induces Tumor Vessel Normalization and Tumor Growth Inhibition in Human Head and Neck Squamous Cell Carcinoma Models. Int. J. Mol. Sci. 2017, 18, 2671. [Google Scholar] [CrossRef] [PubMed]
- Eriksdotter-Nilsson, M.; Bjorklund, H.; Olson, L. Laminin immunohistochemistry: A simple method to visualize and quantitate vascular structures in the mammalian brain. J. Neurosci. Methods 1986, 17, 275–286. [Google Scholar] [CrossRef]
- Walchli, T.; Mateos, J.M.; Weinman, O.; Babic, D.; Regli, L.; Hoerstrup, S.P.; Gerhardt, H.; Schwab, M.E.; Vogel, J. Quantitative assessment of angiogenesis, perfused blood vessels and endothelial tip cells in the postnatal mouse brain. Nat. Protoc. 2015, 10, 53–74. [Google Scholar] [CrossRef] [PubMed]
- Honkura, N.; Richards, M.; Lavina, B.; Sainz-Jaspeado, M.; Betsholtz, C.; Claesson-Welsh, L. Intravital imaging-based analysis tools for vessel identification and assessment of concurrent dynamic vascular events. Nat. Commun. 2018, 9, 2746. [Google Scholar] [CrossRef] [PubMed]
- Iliff, J.J.; Wang, M.; Zeppenfeld, D.M.; Venkataraman, A.; Plog, B.A.; Liao, Y.; Deane, R.; Nedergaard, M. Cerebral arterial pulsation drives paravascular CSF-interstitial fluid exchange in the murine brain. J. Neurosci. 2013, 33, 18190–18199. [Google Scholar] [CrossRef] [PubMed]
- Mestre, H.; Tithof, J.; Du, T.; Song, W.; Peng, W.; Sweeney, A.M.; Olveda, G.; Thomas, J.H.; Nedergaard, M.; Kelley, D.H. Flow of cerebrospinal fluid is driven by arterial pulsations and is reduced in hypertension. Nat. Commun. 2018, 9, 4878. [Google Scholar] [CrossRef]
- Steinman, J.; Koletar, M.M.; Stefanovic, B.; Sled, J.G. 3D morphological analysis of the mouse cerebral vasculature: Comparison of in vivo and ex vivo methods. PLoS ONE 2017, 12, e0186676. [Google Scholar] [CrossRef] [PubMed]
- Schaad, L.; Hlushchuk, R.; Barre, S.; Gianni-Barrera, R.; Haberthur, D.; Banfi, A.; Djonov, V. Correlative Imaging of the Murine Hind Limb Vasculature and Muscle Tissue by MicroCT and Light Microscopy. Sci. Rep. 2017, 7, 41842. [Google Scholar] [CrossRef] [PubMed]
- Mazzetti, S.; Frigerio, S.; Gelati, M.; Salmaggi, A.; Vitellaro-Zuccarello, L. Lycopersicon esculentum lectin: An effective and versatile endothelial marker of normal and tumoral blood vessels in the central nervous system. Eur. J. Histochem. 2004, 48, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Sorriento, D.; Campanile, A.; Santulli, G.; Leggiero, E.; Pastore, L.; Trimarco, B.; Iaccarino, G. A new synthetic protein, TAT-RH, inhibits tumor growth through the regulation of NFkappaB activity. Mol. Cancer 2009, 8, 97. [Google Scholar] [CrossRef] [PubMed]
- Santulli, G.; Basilicata, M.F.; De Simone, M.; Del Giudice, C.; Anastasio, A.; Sorriento, D.; Saviano, M.; Del Gatto, A.; Trimarco, B.; Pedone, C.; et al. Evaluation of the anti-angiogenic properties of the new selective alphaVbeta3 integrin antagonist RGDechiHCit. J. Transl. Med. 2011, 9, 7. [Google Scholar] [CrossRef] [PubMed]
- Robertson, R.T.; Levine, S.T.; Haynes, S.M.; Gutierrez, P.; Baratta, J.L.; Tan, Z.; Longmuir, K.J. Use of labeled tomato lectin for imaging vasculature structures. Histochem. Cell Biol. 2015, 143, 225–234. [Google Scholar] [CrossRef]
- Chen, H.; Fu, X.; Jiang, J.; Han, S. C16 Peptide Promotes Vascular Growth and Reduces Inflammation in a Neuromyelitis Optica Model. Front. Pharmacol. 2019, 10, 1373. [Google Scholar] [CrossRef] [PubMed]
- Bryson, J.L.; Coles, M.C.; Manley, N.R. A method for labeling vasculature in embryonic mice. J. Vis. Exp. 2011, 56, 3267. [Google Scholar] [CrossRef] [PubMed]
- Simionescu, M.; Simionescu, N.; Palade, G.E. Differentiated microdomains on the luminal surface of capillary endothelium: Distribution of lectin receptors. J. Cell Biol. 1982, 94, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Minamikawa, T.; Miyake, T.; Takamatsu, T.; Fujita, S. A new method of lectin histochemistry for the study of brain angiogenesis. Lectin angiography. Histochemistry 1987, 87, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Todorov, M.I.; Paetzold, J.C.; Schoppe, O.; Tetteh, G.; Shit, S.; Efremov, V.; Todorov-Volgyi, K.; During, M.; Dichgans, M.; Piraud, M.; et al. Machine learning analysis of whole mouse brain vasculature. Nat. Methods 2020, 17, 442–449. [Google Scholar] [CrossRef]
- Jilani, S.M.; Murphy, T.J.; Thai, S.N.; Eichmann, A.; Alva, J.A.; Iruela-Arispe, M.L. Selective binding of lectins to embryonic chicken vasculature. J. Histochem. Cytochem. 2003, 51, 597–604. [Google Scholar] [CrossRef]
- Dullmann, J.; Van Damme, E.J.; Peumans, W.J.; Ziesenitz, M.; Schumacher, U. Lectin histochemistry of the rat lymph node: Visualisation of stroma, blood vessels, sinuses, and macrophages. A contribution to the concept of an immune accessory role of sinus-lining endothelia. Acta Histochem. 2002, 104, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.Y.; Peng, S.J.; Lin, H.Y.; Pasricha, P.J.; Tang, S.C. 3-D imaging and illustration of mouse intestinal neurovascular complex. Am. J. Physiol.-Gastrointest. Liver Physiol. 2013, 304, G1–G11. [Google Scholar] [CrossRef] [PubMed]
- Nag, S. Ultrastructural localization of lectin receptors on cerebral endothelium. Acta Neuropathol. 1985, 66, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Braet, F.; Wisse, E. Structural and functional aspects of liver sinusoidal endothelial cell fenestrae: A review. Comp. Hepatol. 2002, 1, 1. [Google Scholar] [CrossRef] [PubMed]
- Quaggin, S.E.; Kreidberg, J.A. Development of the renal glomerulus: Good neighbors and good fences. Development 2008, 135, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.; Chung, H.K.; Ramu, S.; Lee, H.N.; Kim, K.E.; Lee, S.; Yoo, J.; Choi, D.; Lee, Y.S.; Aguilar, B.; et al. Visualization of lymphatic vessels by Prox1-promoter directed GFP reporter in a bacterial artificial chromosome-based transgenic mouse. Blood 2011, 117, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Leino, R.L.; Gerhart, D.Z.; van Bueren, A.M.; McCall, A.L.; Drewes, L.R. Ultrastructural localization of GLUT 1 and GLUT 3 glucose transporters in rat brain. J. Neurosci. Res. 1997, 49, 617–626. [Google Scholar] [CrossRef]
- Dumont, D.J.; Yamaguchi, T.P.; Conlon, R.A.; Rossant, J.; Breitman, M.L. Tek, a novel tyrosine kinase gene located on mouse chromosome 4, is expressed in endothelial cells and their presumptive precursors. Oncogene 1992, 7, 1471–1480. [Google Scholar] [PubMed]
- Motoike, T.; Loughna, S.; Perens, E.; Roman, B.L.; Liao, W.; Chau, T.C.; Richardson, C.D.; Kawate, T.; Kuno, J.; Weinstein, B.M.; et al. Universal GFP reporter for the study of vascular development. Genesis 2000, 28, 75–81. [Google Scholar] [CrossRef]
- Dullmann, J.; Feldhaus, S.; Van Damme, E.J.; Peumans, W.J.; Schumacher, U. Lectin histochemistry of the spleen: A new lectin visualizes the stromal architecture of white pulp and the sinuses of red pulp. J. Histochem. Cytochem. 2000, 48, 923–931. [Google Scholar] [CrossRef]
- Nitzsche, A.; Poittevin, M.; Benarab, A.; Bonnin, P.; Faraco, G.; Uchida, H.; Favre, J.; Garcia-Bonilla, L.; Garcia, M.C.L.; Leger, P.L.; et al. Endothelial S1P1 Signaling Counteracts Infarct Expansion in Ischemic Stroke. Circ. Res. 2021, 128, 363–382. [Google Scholar] [CrossRef] [PubMed]
- Swendeman, S.L.; Xiong, Y.; Cantalupo, A.; Yuan, H.; Burg, N.; Hisano, Y.; Cartier, A.; Liu, C.H.; Engelbrecht, E.; Blaho, V.; et al. An engineered S1P chaperone attenuates hypertension and ischemic injury. Sci. Signal. 2017, 10. [Google Scholar] [CrossRef]
- Iadecola, C.; Anrather, J. The immunology of stroke: From mechanisms to translation. Nat. Med. 2011, 17, 796–808. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Lu, J.; Shao, A.; Zhang, J.H.; Zhang, J. Glial Cells: Role of the Immune Response in Ischemic Stroke. Front. Immunol. 2020, 11, 294. [Google Scholar] [CrossRef] [PubMed]
- Manalo, T.; May, A.; Quinn, J.; Lafontant, D.S.; Shifatu, O.; He, W.; Gonzalez-Rosa, J.M.; Burns, G.C.; Burns, C.E.; Burns, A.R.; et al. Differential Lectin Binding Patterns Identify Distinct Heart Regions in Giant Danio ( Devario aequipinnatus) and Zebrafish (Danio rerio) Hearts. J. Histochem. Cytochem. 2016, 64, 687–714. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Liu, W.A.; Zhang, X.J.; Shi, W.; Ren, S.Q.; Li, Z.; Brown, K.N.; Shi, S.H. Vascular Influence on Ventral Telencephalic Progenitors and Neocortical Interneuron Production. Dev. Cell 2016, 36, 624–638. [Google Scholar] [CrossRef] [PubMed]
- Birdsey, G.M.; Shah, A.V.; Dufton, N.; Reynolds, L.E.; Osuna Almagro, L.; Yang, Y.; Aspalter, I.M.; Khan, S.T.; Mason, J.C.; Dejana, E.; et al. The endothelial transcription factor ERG promotes vascular stability and growth through Wnt/beta-catenin signaling. Dev. Cell 2015, 32, 82–96. [Google Scholar] [CrossRef]
- Benedito, R.; Rocha, S.F.; Woeste, M.; Zamykal, M.; Radtke, F.; Casanovas, O.; Duarte, A.; Pytowski, B.; Adams, R.H. Notch-dependent VEGFR3 upregulation allows angiogenesis without VEGF-VEGFR2 signalling. Nature 2012, 484, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Poulet, M.; Sirois, J.; Boye, K.; Uetani, N.; Hardy, S.; Daubon, T.; Dubrac, A.; Tremblay, M.L.; Bikfalvi, A. PRL-2 phosphatase is required for vascular morphogenesis and angiogenic signaling. Commun. Biol. 2020, 3, 603. [Google Scholar] [CrossRef]
- Quan, W.; Luo, Q.; Tang, Q.; Furihata, T.; Li, D.; Fassbender, K.; Liu, Y. NLRP3 Is Involved in the Maintenance of Cerebral Pericytes. Front. Cell Neurosci. 2020, 14, 276. [Google Scholar] [CrossRef] [PubMed]
- Coucha, M.; Barrett, A.C.; Elgebaly, M.; Ergul, A.; Abdelsaid, M. Inhibition of Ephrin-B2 in brain pericytes decreases cerebral pathological neovascularization in diabetic rats. PLoS ONE 2019, 14, e0210523. [Google Scholar] [CrossRef] [PubMed]
- Jiao, C.; Adler, K.; Liu, X.; Sun, W.; Mullins, R.F.; Sohn, E.H. Visualization of Mouse Choroidal and Retinal Vasculature Using Fluorescent Tomato Lectin Perfusion. Transl. Vis. Sci. Technol. 2020, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Nakamura-Ishizu, A.; Morikawa, S.; Shimizu, K.; Ezaki, T. Characterization of sinusoidal endothelial cells of the liver and bone marrow using an intravital lectin injection method. J. Mol. Histol. 2008, 39, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, I.; Poretz, R.D. Isolation, physicochemical characterization, and carbohydrate—binding specificity of Iectins. In The Lectins. Properties, Functions, and Applications in Biology and Medicine; Academic Press: Cambridge, MA, USA, 2012; pp. 233–247. [Google Scholar]
- Wearne, K.A.; Winter, H.C.; O’Shea, K.; Goldstein, I.J. Use of lectins for probing differentiated human embryonic stem cells for carbohydrates. Glycobiology 2006, 16, 981–990. [Google Scholar] [CrossRef] [PubMed]
- Kostrominova, T.Y. Application of WGA lectin staining for visualization of the connective tissue in skeletal muscle, bone, and ligament/tendon studies. Microsc. Res. Tech. 2011, 74, 18–22. [Google Scholar] [CrossRef]
- Schwab, M.E.; Javoy-Agid, F.; Agid, Y. Labeled wheat germ agglutinin (WGA) as a new, highly sensitive retrograde tracer in the rat brain hippocampal system. Brain Res. 1978, 152, 145–150. [Google Scholar] [CrossRef]
- Gurd, J.W. Synaptic plasma membrane glycoproteins: Molecular identification of lectin receptors. Biochemistry 1977, 16, 369–374. [Google Scholar] [CrossRef]
- Melo-Junior, M.R.; Cavalcanti, C.L.; Pontes-Filho, N.T.; Carvalho, L.B., Jr.; Beltrao, E.I. Lectin staining patterns in human gastric mucosae with and without exposure to Helicobacter pylori. Braz. J. Microbiol. 2008, 39, 238–240. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cuzzocrea, S.; Mazzon, E.; Costantino, G.; Serraino, I.; De Sarro, A.; Caputi, A.P. Effects of n-acetylcysteine in a rat model of ischemia and reperfusion injury. Cardiovasc. Res. 2000, 47, 537–548. [Google Scholar] [CrossRef]
- Dai, W.; Wang, Y.; Yang, T.; Wang, J.; Wu, W.; Gu, J. Downregulation of exosomal CLEC3B in hepatocellular carcinoma promotes metastasis and angiogenesis via AMPK and VEGF signals. Cell Commun. Signal. 2019, 17, 113. [Google Scholar] [CrossRef] [PubMed]
- Hauke, C.; Korr, H. RCA-I lectin histochemistry after trypsinisation enables the identification of microglial cells in thin paraffin sections of the mouse brain. J. Neurosci. Methods 1993, 50, 273–277. [Google Scholar] [CrossRef]
- Andjelkovic, A.V.; Nikolic, B.; Pachter, J.S.; Zecevic, N. Macrophages/microglial cells in human central nervous system during development: An immunohistochemical study. Brain Res. 1998, 814, 13–25. [Google Scholar] [CrossRef]
- Hirko, A.C.; Dallasen, R.; Jomura, S.; Xu, Y. Modulation of inflammatory responses after global ischemia by transplanted umbilical cord matrix stem cells. Stem Cells 2008, 26, 2893–2901. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Li, P.; Guo, Y.; Wang, H.; Leak, R.K.; Chen, S.; Gao, Y.; Chen, J. Microglia/macrophage polarization dynamics reveal novel mechanism of injury expansion after focal cerebral ischemia. Stroke 2012, 43, 3063–3070. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Munch, A.E.; Chung, W.S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef]
- Choudhury, G.R.; Ding, S. Reactive astrocytes and therapeutic potential in focal ischemic stroke. Neurobiol. Dis. 2016, 85, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Rakers, C.; Schleif, M.; Blank, N.; Matuskova, H.; Ulas, T.; Handler, K.; Torres, S.V.; Schumacher, T.; Tai, K.; Schultze, J.L.; et al. Stroke target identification guided by astrocyte transcriptome analysis. Glia 2019, 67, 619–633. [Google Scholar] [CrossRef] [PubMed]
- Gleave, J.A.; Wong, M.D.; Dazai, J.; Altaf, M.; Henkelman, R.M.; Lerch, J.P.; Nieman, B.J. Neuroanatomical phenotyping of the mouse brain with three-dimensional autofluorescence imaging. Physiol. Genomics 2012, 44, 778–785. [Google Scholar] [CrossRef] [PubMed]
- Orsini, F.; Villa, P.; Parrella, S.; Zangari, R.; Zanier, E.R.; Gesuete, R.; Stravalaci, M.; Fumagalli, S.; Ottria, R.; Reina, J.J.; et al. Targeting mannose-binding lectin confers long-lasting protection with a surprisingly wide therapeutic window in cerebral ischemia. Circulation 2012, 126, 1484–1494. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lectin Abbreviation | Source | Carbohydrate Group | Nominal Carbohydrate Specificity |
|---|---|---|---|
| WGA | Triticum vulgaris | N-acetylglucosammine | GlcNAc(β1,4GlcNAc)1-2 > βGlcNAc > Neu5Ac |
| LEA | Lycopersicon esculentum (Tomato) | N-acetylglucosammine | GlcNAcβ1,4GlcNAcβ1,4GlcNAcβ1,4GlcNAc > GlcNAcβ1,4GlcNAcβ1,4GlcNAc > GlcNAcβ1,4GlcNAc |
| IB4 | Griffonia (Bandeiraea) simplicifolia | N-acetylglucosammine/ galactose | αGlcNAc = βGlcNAc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Battistella, R.; Kritsilis, M.; Matuskova, H.; Haswell, D.; Cheng, A.X.; Meissner, A.; Nedergaard, M.; Lundgaard, I. Not All Lectins Are Equally Suitable for Labeling Rodent Vasculature. Int. J. Mol. Sci. 2021, 22, 11554. https://doi.org/10.3390/ijms222111554
Battistella R, Kritsilis M, Matuskova H, Haswell D, Cheng AX, Meissner A, Nedergaard M, Lundgaard I. Not All Lectins Are Equally Suitable for Labeling Rodent Vasculature. International Journal of Molecular Sciences. 2021; 22(21):11554. https://doi.org/10.3390/ijms222111554
Chicago/Turabian StyleBattistella, Roberta, Marios Kritsilis, Hana Matuskova, Douglas Haswell, Anne Xiaoan Cheng, Anja Meissner, Maiken Nedergaard, and Iben Lundgaard. 2021. "Not All Lectins Are Equally Suitable for Labeling Rodent Vasculature" International Journal of Molecular Sciences 22, no. 21: 11554. https://doi.org/10.3390/ijms222111554
APA StyleBattistella, R., Kritsilis, M., Matuskova, H., Haswell, D., Cheng, A. X., Meissner, A., Nedergaard, M., & Lundgaard, I. (2021). Not All Lectins Are Equally Suitable for Labeling Rodent Vasculature. International Journal of Molecular Sciences, 22(21), 11554. https://doi.org/10.3390/ijms222111554