
The Dopaminergic Control of Movement-Evolutionary Considerations



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
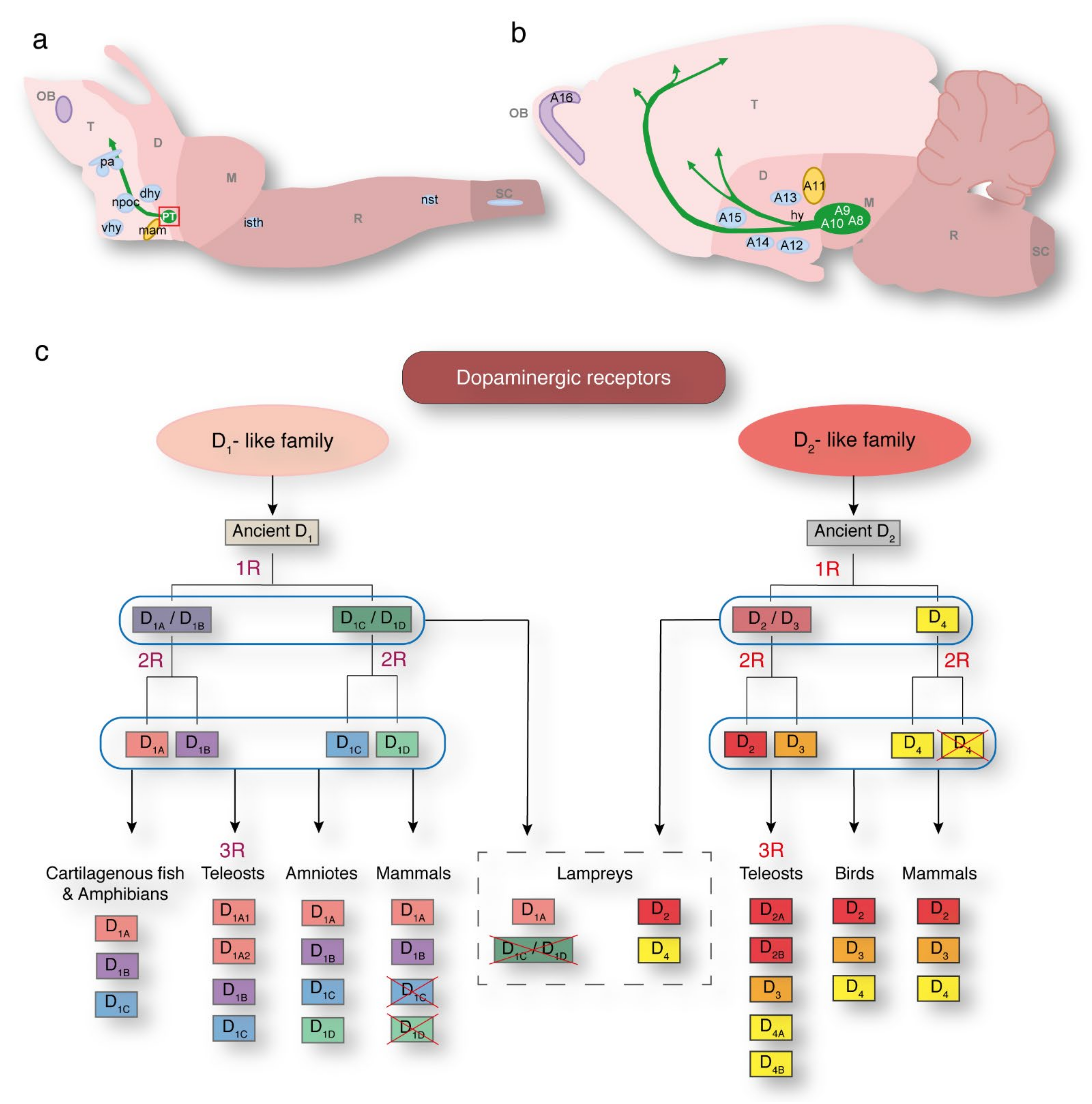
2. Dopaminergic Populations and Receptors in the Lamprey
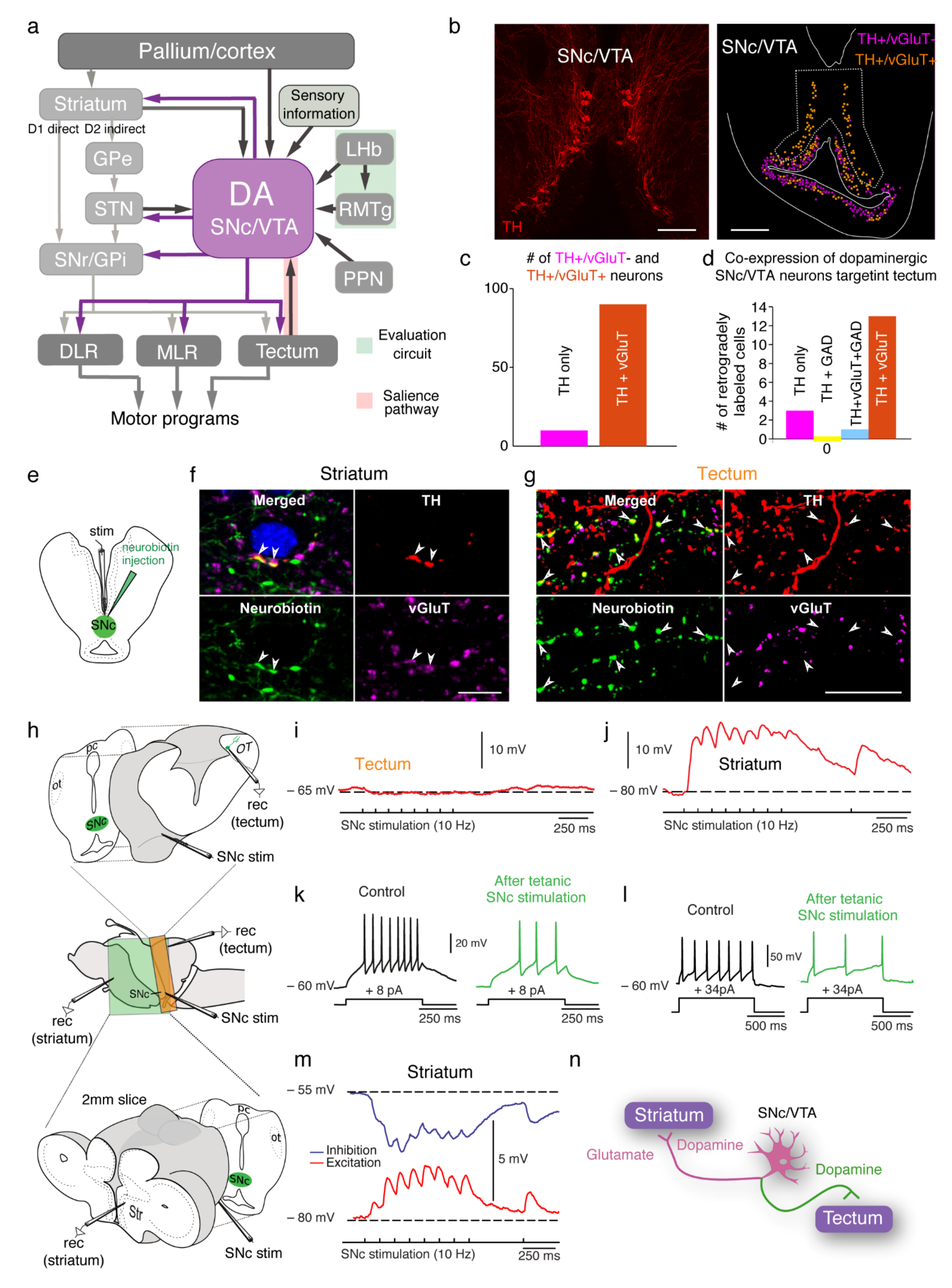
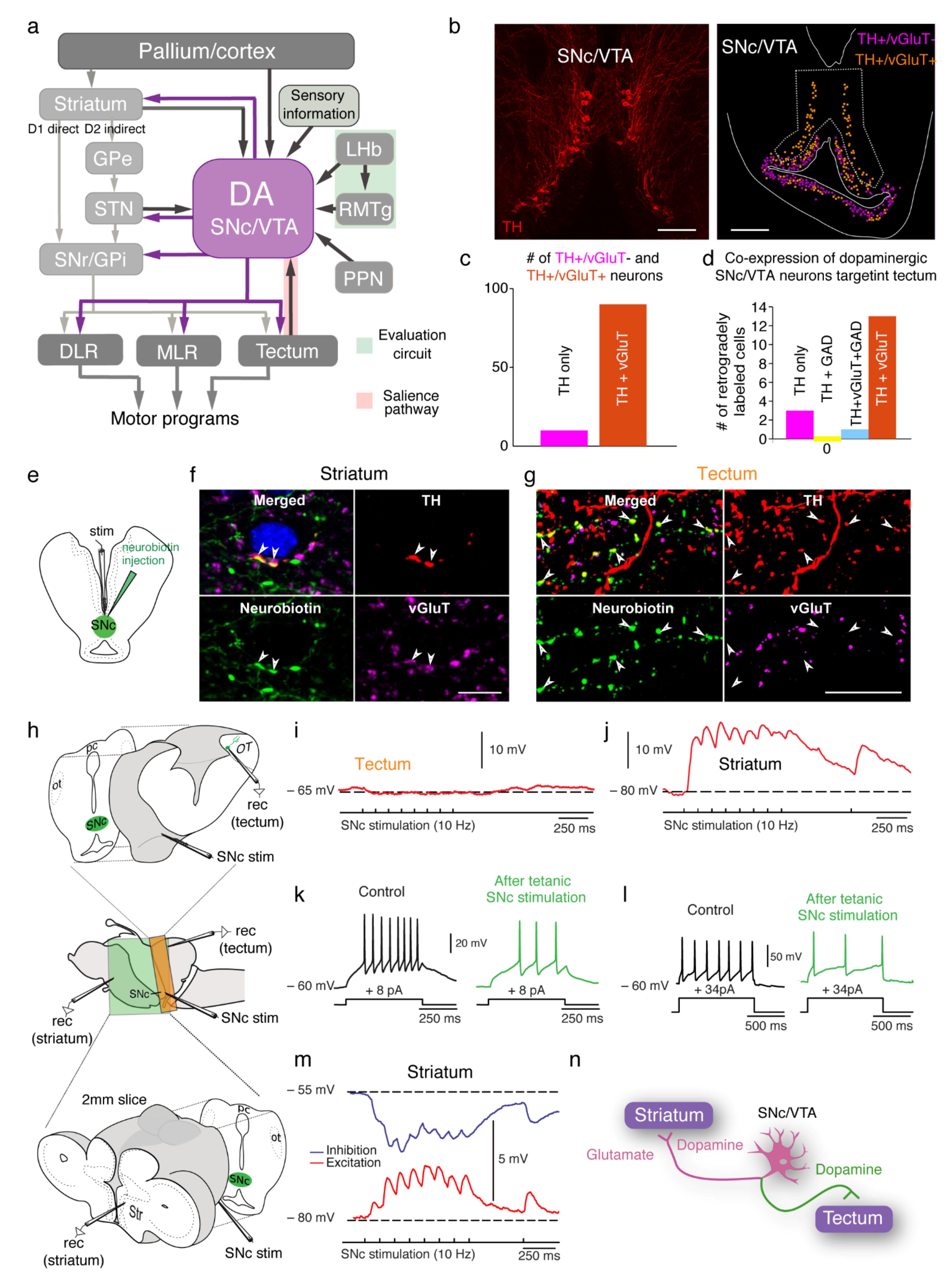
3. The Basal Ganglia Are Largely Conserved from Lampreys to Mammals
4. The SNc/VTA at the Base of Vertebrate Evolution
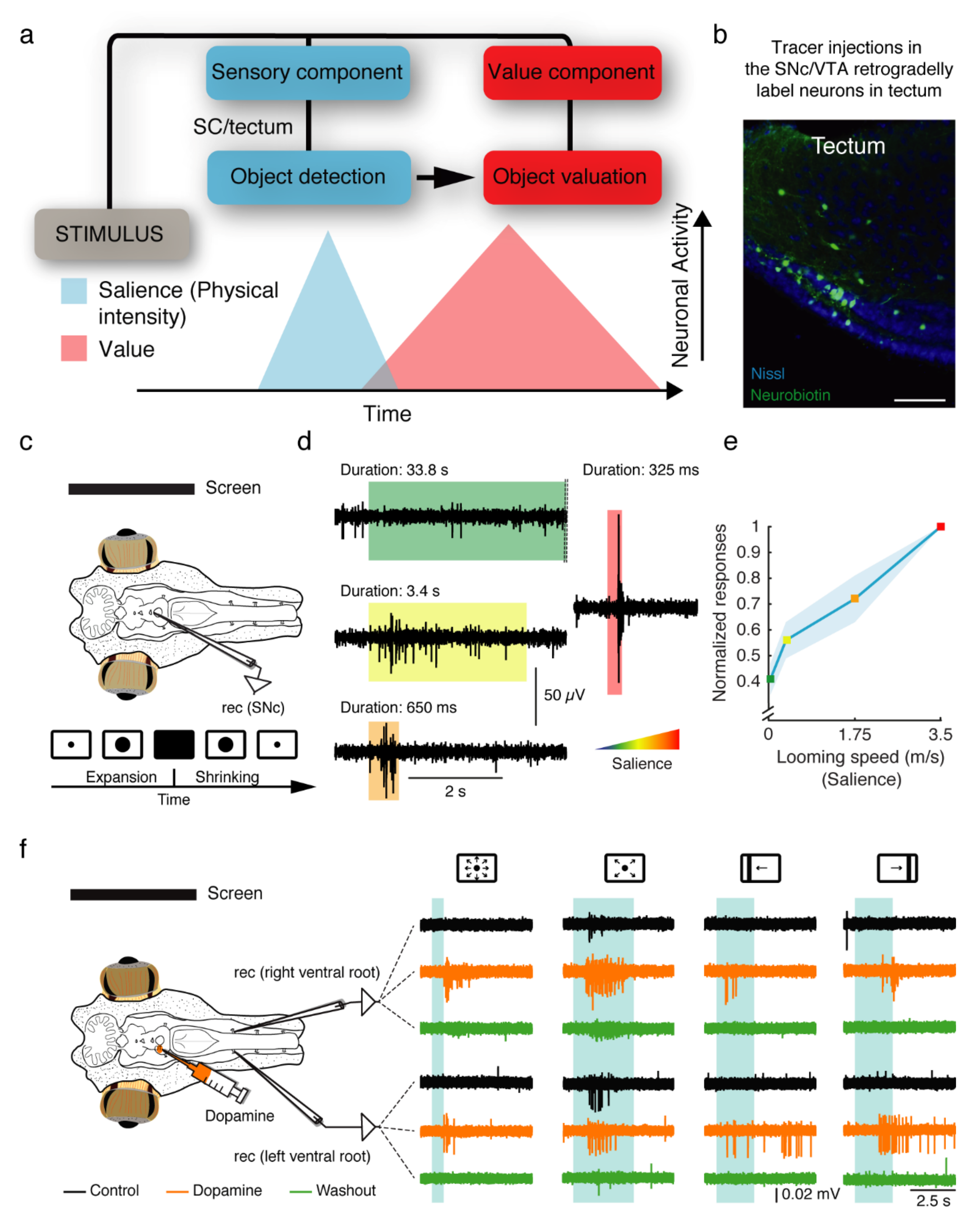
5. Early Origin of the Dopaminergic Coding of Salience and Reward
6. Direct Dopaminergic Control of Downstream Centers
7. Co-Release of Dopamine and Other Neurotransmitters
8. Evolution of Mesodiencephalic Dopaminergic Neurons
9. Evolution of the Mesolimbic and Mesocortical Pathways
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diederich, N.J.; Uchihara, T.; Grillner, S.; Goetz, C.G. The evolution-driven signature of Parkinson’s disease. Trends Neurosci. 2020, 43, 475–492. [Google Scholar] [CrossRef]
- Thompson, R.H.; Ménard, A.; Pombal, M.; Grillner, S. Forebrain dopamine depletion impairs motor behavior in lamprey. Eur. J. Neurosci. 2008, 27, 1452–1460. [Google Scholar] [CrossRef]
- Pérez-Fernández, J.; Stephenson-Jones, M.; Suryanarayana, S.M.; Robertson, B.; Grillner, S. Evolutionarily conserved organization of the dopaminergic system in lamprey: SNc/VTA afferent and efferent connectivity and D2 receptor expression. J. Comp. Neurol. 2014, 522, 3775–3794. [Google Scholar] [CrossRef] [Green Version]
- Suryanarayana, S.M.; Pérez-Fernández, J.; Robertson, B.; Grillner, S. The lamprey forebrain—Evolutionary implications. Brain. Behav. Evol. 2021, 1–16. [Google Scholar] [CrossRef]
- Grillner, S.; Robertson, B. The basal ganglia over 500 million years. Curr. Biol. 2016, 26, R1088–R1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumgarten, H.G. Biogenic monoamines in the cyclostome and lower vertebrate brain. Prog. Histochem. Cytochem. 1972, 4, III-8. [Google Scholar] [CrossRef]
- Pombal, M.A.; Manira, A.E.L.; Grillner, S. Afferents of the lamprey striatum with special reference to the dopaminergic system: A combined tracing and immunohistochemical study. J. Comp. Neurol. 1997, 386, 71–91. [Google Scholar] [CrossRef]
- Pérez-Fernández, J.; Kardamakis, A.A.; Suzuki, D.G.; Robertson, B.; Grillner, S. Direct dopaminergic projections from the SNc modulate visuomotor transformation in the lamprey tectum. Neuron 2017, 96, 910–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierre, J.; Mahouche, M.; Suderevskaya, E.I.; Repérant, J.; Ward, R. Immunocytochemical localization of dopamine and its synthetic enzymes in the central nervous system of the lamprey Lampetra fluviatilis. J. Comp. Neurol. 1997, 380, 119–135. [Google Scholar] [CrossRef]
- Abalo, X.M.; Villar-Cheda, B.; Anadón, R.; Rodicio, M.C. Development of the dopamine-immunoreactive system in the central nervous system of the sea lamprey. Brain Res. Bull. 2005, 66, 560–564. [Google Scholar] [CrossRef]
- Björklund, A.; Dunnett, S.B. Dopamine neuron systems in the brain: An update. Trends Neurosci. 2007, 30, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Barreiro-Iglesias, A.; Villar-Cerviño, V.; Anadón, R.; Rodicio, M.C. Descending brain-spinal cord projections in a primitive vertebrate, the lamprey: Cerebrospinal fluid-contacting and dopaminergic neurons. J. Comp. Neurol. 2008, 511, 711–723. [Google Scholar] [CrossRef]
- Schotland, J.; Shupliakov, O.; Wikström, M.; Brodin, L.; Srinivasan, M.; You, Z.B.; Herrera-Marschitz, M.; Zhang, W.; Hökfelt, T.; Grillner, S. Control of lamprey locomotor neurons by colocalized monoamine transmitters. Nature 1995, 374, 262–266. [Google Scholar] [CrossRef]
- Misganaw, D. Heteromerization of dopaminergic receptors in the brain: Pharmacological implications. Pharmacol. Res. 2021, 170, 105600. [Google Scholar] [CrossRef]
- Xin, J.; Fan, T.; Guo, P.; Wang, J. Identification of functional divergence sites in dopamine receptors of vertebrates. Comput. Biol. Chem. 2019, 83, 107140. [Google Scholar] [CrossRef]
- Yamamoto, K.; Fontaine, R.; Pasqualini, C.; Vernier, P. Classification of dopamine receptor genes in vertebrates: Nine subtypes in osteichthyes. Brain. Behav. Evol. 2015, 86, 164–175. [Google Scholar] [CrossRef]
- Missale, C.; Russel Nash, S.; Robinson, S.W.; Jaber, M.; Caron, M.G. Dopamine receptors: From structure to function. Physiol. Rev. 1998, 78, 189–225. [Google Scholar] [CrossRef] [Green Version]
- Callier, S.; Snapyan, M.; Le Crom, S.; Prou, D.; Vincent, J.D.; Vernier, P. Evolution and cell biology of dopamine receptors in vertebrates. Biol. Cell 2003, 95, 489–502. [Google Scholar] [CrossRef]
- Nakatani, Y.; Shingate, P.; Ravi, V.; Pillai, N.E.; Prasad, A.; McLysaght, A.; Venkatesh, B. Reconstruction of proto-vertebrate, proto-cyclostome and proto-gnathostome genomes provides new insights into early vertebrate evolution. Nat. Commun. 2021, 12, 4489. [Google Scholar] [CrossRef]
- Le Crom, S.; Sugamori, K.S.; Sidhu, A.; Niznik, H.B.; Vernier, P. Delineation of the conserved functional properties of D1A, D1B and D1C dopamine receptor subtypes in vertebrates. Biol. Cell 2004, 96, 383–394. [Google Scholar] [CrossRef]
- Yamamoto, K.; Vernier, P. The evolution of dopamine systems in chordates. Front. Neuroanat. 2011, 5, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Fernandez, J.; Megías, M.; Pombal, M.A. Expression of a novel D4 dopamine receptor in the lamprey brain. Evolutionary considerations about dopamine receptors. Front. Neuroanat. 2016, 9, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasqualini, C.; Weltzien, F.A.; Vidal, B.; Baloche, S.; Rouget, C.; Gilles, N.; Servent, D.; Vernier, P.; Dufour, S. Two distinct dopamine D2 receptor genes in the european eel: Molecular characterization, tissue-specific transcription, and regulation by sex steroids. Endocrinology 2009, 150, 1377–1392. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Fernández, J.; Megías, M.; Pombal, M.A. Distribution of a Y1 receptor mRNA in the brain of two lamprey species, the sea lamprey (Petromyzon marinus) and the river lamprey (Lampetra fluviatilis). J. Comp. Neurol. 2013, 521, 426–447. [Google Scholar] [CrossRef] [PubMed]
- Osório, J.; Rétaux, S. The lamprey in evolutionary studies. Dev. Genes Evol. 2008, 218, 221–235. [Google Scholar] [CrossRef]
- Smith, J.J.; Kuraku, S.; Holt, C.; Sauka-Spengler, T.; Jiang, N.; Campbell, M.S.; Yandell, M.D.; Manousaki, T.; Meyer, A.; Bloom, O.E.; et al. Sequencing of the sea lamprey (Petromyzon marinus) genome provides insights into vertebrate evolution. Nat. Genet. 2013, 45, 415–421. [Google Scholar] [CrossRef]
- Kasahara, M. The 2R hypothesis: An update. Curr. Opin. Immunol. 2007, 19, 547–552. [Google Scholar] [CrossRef]
- Ericsson, J.; Stephenson-Jones, M.; Pérez-Fernández, J.; Robertson, B.; Silberberg, G.; Grillner, S. Dopamine differentially modulates the excitability of striatal neurons of the direct and indirect pathways in lamprey. J. Neurosci. 2013, 33, 8045–8054. [Google Scholar] [CrossRef]
- Pérez-Fernández, J. Characterization of Y and Dopamine Receptors in Lampreys by Using in situ Hybridization: An Evolutionary Approach. PhD Thesis, University of Vigo, Pontevedra, Spain, 2013. [Google Scholar]
- Robertson, B.; Huerta-Ocampo, I.; Ericsson, J.; Stephenson-Jones, M.; Pérez-Fernández, J.; Bolam, J.P.; Diaz-Heijtz, R.; Grillner, S. The dopamine D2 receptor gene in lamprey, its expression in the striatum and cellular effects of D2 receptor activation. PLoS ONE 2012, 7, e35642. [Google Scholar] [CrossRef]
- Athalye, V.R.; Carmena, J.M.; Costa, R.M. Neural reinforcement: Re-entering and refining neural dynamics leading to desirable outcomes. Curr. Opin. Neurobiol. 2020, 60, 145–154. [Google Scholar] [CrossRef]
- Ericsson, J.; Stephenson-Jones, M.; Kardamakis, A.; Robertson, B.; Silberberg, G.; Grillner, S. Evolutionarily conserved differences in pallial and thalamic short-term synaptic plasticity in striatum. J. Physiol. 2013, 591, 859–874. [Google Scholar] [CrossRef] [Green Version]
- Stephenson-Jones, M.; Samuelsson, E.; Ericsson, J.; Robertson, B.; Grillner, S. Evolutionary conservation of the basal ganglia as a common vertebrate mechanism for action selection. Curr. Biol. 2011, 21, 1081–1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephenson-Jones, M.; Ericsson, J.; Robertson, B.; Grillner, S. Evolution of the basal ganglia: Dual-output pathways conserved throughout vertebrate phylogeny. J. Comp. Neurol. 2012, 520, 2957–2973. [Google Scholar] [CrossRef] [Green Version]
- Stephenson-Jones, M.; Floros, O.; Robertson, B.; Grillner, S. Evolutionary conservation of the habenular nuclei and their circuitry controlling the dopamine and 5-hydroxytryptophan (5-HT) systems. Proc. Natl. Acad. Sci. USA 2012, 109, E164–E173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephenson-Jones, M.; Kardamakis, A.A.; Robertson, B.; Grillner, S. Independent circuits in the basal ganglia for the evaluation and selection of actions. Proc. Natl. Acad. Sci. USA 2013, 110, E3670–E3679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ericsson, J.; Silberberg, G.; Robertson, B.; Wikström, M.A.; Grillner, S. Striatal cellular properties conserved from lampreys to mammals. J. Physiol. 2011, 589, 2979–2992. [Google Scholar] [CrossRef]
- Gerfen, C.R.; Surmeier, D.J. Modulation of striatal projection systems by dopamine. Annu. Rev. Neurosci. 2011, 34, 441–466. [Google Scholar] [CrossRef] [Green Version]
- Sugahara, F.; Pascual-Anaya, J.; Oisi, Y.; Kuraku, S.; Aota, S.I.; Adachi, N.; Takagi, W.; Hirai, T.; Sato, N.; Murakami, Y.; et al. Evidence from cyclostomes for complex regionalization of the ancestral vertebrate brain. Nature 2016, 531, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Sugahara, F.; Murakami, Y.; Pascual-Anaya, J.; Kuratani, S. Reconstructing the ancestral vertebrate brain. Dev. Growth Differ. 2017, 59, 163–174. [Google Scholar] [CrossRef] [Green Version]
- Von Twickel, A.; Kowatschew, D.; Saltürk, M.; Schauer, M.; Robertson, B.; Korsching, S.; Walkowiak, W.; Grillner, S.; Pérez-Fernández, J. Individual dopaminergic neurons of lamprey SNc/VTA project to both the striatum and optic tectum but restrict co-release of glutamate to striatum only. Curr. Biol. 2019, 29, 677–685.e6. [Google Scholar] [CrossRef] [Green Version]
- Ocaña, F.M.; Suryanarayana, S.M.; Saitoh, K.; Kardamakis, A.A.; Capantini, L.; Robertson, B.; Grillner, S. The lamprey pallium provides a blueprint of the mammalian motor projections from cortex. Curr. Biol. 2015, 25, 413–423. [Google Scholar] [CrossRef] [Green Version]
- Suryanarayana, S.M.; Robertson, B.; Wallén, P.; Grillner, S. The Lamprey Pallium Provides a Blueprint of the Mammalian Layered Cortex. Curr. Biol. 2017, 27, 3264–3277. [Google Scholar] [CrossRef] [Green Version]
- Suryanarayana, S.M.; Pérez-Fernández, J.; Robertson, B.; Grillner, S. The evolutionary origin of visual and somatosensory representation in the vertebrate pallium. Nat. Ecol. Evol. 2020, 4, 639–651. [Google Scholar] [CrossRef]
- Bevan, M.D.; Hallworth, N.E.; Baufreton, J. GABAergic control of the subthalamic nucleus. Prog. Brain Res. 2007, 160, 173–188. [Google Scholar] [CrossRef]
- Derjean, D.; Moussaddy, A.; Atallah, E.; St-Pierre, M.; Auclair, F.; Chang, S.; Ren, X.; Zielinski, B.; Dubuc, R. A novel neural substrate for the transformation of olfactory inputs into motor output. PLoS Biol. 2010, 8, e1000567. [Google Scholar] [CrossRef] [Green Version]
- Beauséjour, P.A.; Auclair, F.; Daghfous, G.; Ngovandan, C.; Veilleux, D.; Zielinski, B.; Dubuc, R. Dopaminergic modulation of olfactory-evoked motor output in sea lampreys (Petromyzon marinus L.). J. Comp. Neurol. 2020, 528, 114–134. [Google Scholar] [CrossRef] [Green Version]
- Daghfous, G.; Green, W.W.; Alford, S.T.; Zielinski, B.S.; Dubuc, R. Sensory activation of command cells for locomotion and modulatory mechanisms: Lessons from lampreys. Front. Neural Circuits 2016, 10, 18. [Google Scholar] [CrossRef] [Green Version]
- Suryanarayana, S.M.; Pérez-Fernández, J.; Robertson, B.; Grillner, S. Olfaction in lamprey pallium revisited—Dual projections of mitral and tufted cells. Cell Rep. 2021, 34, 108596. [Google Scholar] [CrossRef] [PubMed]
- Capantini, L.; von Twickel, A.; Robertson, B.; Grillner, S. The pretectal connectome in lamprey. J. Comp. Neurol. 2017, 525, 753–772. [Google Scholar] [CrossRef] [PubMed]
- Kardamakis, A.A.; Pérez-Fernández, J.; Grillner, S. Spatiotemporal interplay between multisensory excitation and recruited inhibition in the lamprey optic tectum. eLife 2016, 5, e16472. [Google Scholar] [CrossRef] [PubMed]
- Wibble, T.; Pansell, T.; Grillner, S.; Pérez-Fernández, J. Visuo-vestibular gaze control—conserved subcortical processing. bioRxiv 2021. [Google Scholar] [CrossRef]
- Pitts, E.G.; Stowe, T.A.; Christensen, B.A.; Ferris, M.J. Comparing dopamine release, uptake, and D2 autoreceptor function across the ventromedial to dorsolateral striatum in adolescent and adult male and female rats. Neuropharmacology 2020, 175, 108163. [Google Scholar] [CrossRef]
- Herculano-Houzel, S.; Mota, B.; Lent, R. Cellular scaling rules for rodent brains. Proc. Natl. Acad. Sci. USA 2006, 103, 12138–12143. [Google Scholar] [CrossRef] [Green Version]
- Bromberg-Martin, E.S.; Matsumoto, M.; Hikosaka, O. Dopamine in Motivational Control: Rewarding, Aversive, and Alerting. Neuron 2010, 68, 815–834. [Google Scholar] [CrossRef] [Green Version]
- Grillner, S.; von Twickel, A.; Robertson, B. The blueprint of the vertebrate forebrain—With special reference to the habenulae. Semin. Cell Dev. Biol. 2018, 78, 103–106. [Google Scholar] [CrossRef]
- Schultz, W.; Stauffer, W.R.; Lak, A. The phasic dopamine signal maturing: From reward via behavioural activation to formal economic utility. Curr. Opin. Neurobiol. 2017, 43, 139–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, W. Recent advances in understanding the role of phasic dopamine activity. F1000Research 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Schultz, W. Dopamine reward prediction-error signalling: A two-component response. Nat. Rev. Neurosci. 2016, 17, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.X.; Coddington, L.T.; Dudman, J.T. Dissociable contributions of phasic dopamine activity to reward and prediction. Cell Rep. 2021, 36, 109684. [Google Scholar] [CrossRef]
- Kutlu, M.G.; Zachry, J.E.; Melugin, P.R.; Cajigas, S.A.; Chevee, M.F.; Kelley, S.J.; Kutlu, B.; Tian, L.; Siciliano, C.A.; Calipari, E.S. Dopamine release in the nucleus accumbens core signals perceived saliency. Curr. Biol. 2021, 31, 1–14. [Google Scholar] [CrossRef]
- Overton, P.G.; Vautrelle, N.; Redgrave, P. Sensory regulation of dopaminergic cell activity: Phenomenology, circuitry and function. Neuroscience 2014, 282, 1–12. [Google Scholar] [CrossRef]
- Saitoh, K.; Ménard, A.; Grillner, S. Tectal control of locomotion, steering, and eye movements in lamprey. J. Neurophysiol. 2007, 97, 3093–3108. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, D.G.; Pérez-Fernández, J.; Wibble, T.; Kardamakis, A.A.; Grillner, S. The role of the optic tectum for visually evoked orienting and evasive movements. Proc. Natl. Acad. Sci. USA 2019, 116, 15272–15281. [Google Scholar] [CrossRef] [Green Version]
- Kardamakis, A.A.; Saitoh, K.; Grillner, S. Tectal microcircuit generating visual selection commands on gaze-controlling neurons. Proc. Natl. Acad. Sci. USA 2015, 112, E1956–E1965. [Google Scholar] [CrossRef] [Green Version]
- Isa, T.; Marquez-Legorreta, E.; Grillner, S.; Scott, E.K. The tectum/superior colliculus as the vertebrate solution for spatial sensory integration and action. Curr. Biol. 2021, 31, R741–R762. [Google Scholar] [CrossRef] [PubMed]
- Comoli, E.; Coizet, V.; Boyes, J.; Bolam, J.P.; Canteras, N.S.; Quirk, R.H.; Overton, P.G.; Redgrave, P. A direct projection from superior colliculus to substantia nigra for detecting salient visual events. Nat. Neurosci. 2003, 6, 974–980. [Google Scholar] [CrossRef] [PubMed]
- Dommett, E.; Coizet, V.; Blaha, C.D.; Martindale, J.; Lefebvre, V.; Walton, N.; Mayhew, J.E.W.; Overton, P.G.; Redgrave, P. How visual stimuli activate dopaminergic neurons at short latency. Science 2005, 307, 1476–1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redgrave, P.; Gurney, K. The short-latency dopamine signal: A role in discovering novel actions? Nat. Rev. Neurosci. 2006, 7, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Redgrave, P.; Coizet, V.; Comoli, E.; McHaffie, J.G.; Leriche, M.; Vautrelle, N.; Hayes, L.M.; Overton, P. Interactions between the midbrain superior colliculus and the basal ganglia. Front. Neuroanat. 2010, 4, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takakuwa, N.; Kato, R.; Redgrave, P.; Isa, T. Emergence of visually-evoked reward expectation signals in dopamine neurons via the superior colliculus in V1 lesioned monkeys. eLife 2017, 6, e24459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, M.; Li, D.; Cheng, X.; Pei, Q.; Xie, Z.; Gu, H.; Zhang, X.; Chen, Z.; Liu, A.; Wang, Y.; et al. The tectonigral pathway regulates appetitive locomotion in predatory hunting in mice. Nat. Commun. 2021, 12, 4409. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, W.Y.; Diao, Y.P.; Xu, W.; Zhong, Y.H.; Zhang, J.Y.; Lazarus, M.; Liu, Y.Y.; Qu, W.M.; Huang, Z.L. Superior colliculus GABAergic neurons are essential for acute dark induction of wakefulness in mice. Curr. Biol. 2019, 29, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Liu, X.; Chen, S.; Zhang, Z.; Liu, Y.; Montardy, Q.; Tang, Y.; Wei, P.; Liu, N.; Li, L.; et al. A VTA GABAergic neural circuit mediates visually evoked innate defensive responses. Neuron 2019, 103, 473–488. [Google Scholar] [CrossRef]
- Pérez-Fernández, J.; Robertson, B.; Grillner, S. Mechanisms for saliency coding and transmission from the lamprey tectum to the SNc/VTA. In Proceedings of the Society for Neuroscience, San Diego, CA, USA, 3–7 November 2018; pp. 700–701. [Google Scholar]
- Stephenson-Jones, M.; Yu, K.; Ahrens, S.; Tucciarone, J.M.; Van Huijstee, A.N.; Mejia, L.A.; Penzo, M.A.; Tai, L.H.; Wilbrecht, L.; Li, B. A basal ganglia circuit for evaluating action outcomes. Nature 2016, 539, 289–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazaridis, I.; Tzortzi, O.; Weglage, M.; Märtin, A.; Xuan, Y.; Parent, M.; Johansson, Y.; Fuzik, J.; Fürth, D.; Fenno, L.E.; et al. A hypothalamus-habenula circuit controls aversion. Mol. Psychiatry 2019, 24, 1351–1368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, P.M.; Jhou, T.; Li, B.; Matsumoto, M.; Mizumori, S.J.Y.; Stephenson-Jones, M.; Vicentic, A. The lateral habenula circuitry: Reward processing and cognitive control. J. Neurosci. 2016, 36, 11482–11488. [Google Scholar] [CrossRef] [Green Version]
- Duboué, E.R.; Hong, E.; Eldred, K.C.; Halpern, M.E. Left habenular activity attenuates fear responses in larval zebrafish. Curr. Biol. 2017, 27, 2154–2162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palumbo, F.; Serneels, B.; Pelgrims, R.; Yaksi, E. The zebrafish dorsolateral habenula is required for updating learned behaviors. Cell Rep. 2020, 32, 108054. [Google Scholar] [CrossRef]
- Amo, R.; Aizawa, H.; Takahoko, M.; Kobayashi, M.; Takahashi, R.; Aoki, T.; Okamoto, H. Identification of the zebrafish ventral habenula as a homolog of the mammalian lateral habenula. J. Neurosci. 2010, 30, 1566–1574. [Google Scholar] [CrossRef]
- Yáñez, J.; Anadón, R. Afferent and efferent connections of the habenula in the rainbow trout (Oncorhynchus mykiss): An indocarbocyanine dye (DiI) study. J. Comp. Neurol. 1996, 372, 529–543. [Google Scholar] [CrossRef]
- Ryczko, D.; Grätsch, S.; Auclair, F.; Dubé, C.; Bergeron, S.; Alpert, M.H.; Cone, J.J.; Roitman, M.F.; Alford, S.; Dubuc, R. Forebrain dopamine neurons project down to a brainstem region controlling locomotion. Proc. Natl. Acad. Sci. USA 2013, 110, E3235–E3242. [Google Scholar] [CrossRef] [Green Version]
- Ryczko, D.; Cone, J.J.; Alpert, M.H.; Goetz, L.; Auclair, F.; Dubé, C.; Parent, M.; Roitman, M.F.; Alford, S.; Dubuc, R. A descending dopamine pathway conserved from basal vertebrates to mammals. Proc. Natl. Acad. Sci. USA. 2016, 113, E2440–E2449. [Google Scholar] [CrossRef] [Green Version]
- Ryczko, D.; Dubuc, R. Dopamine and the brainstem locomotor networks: From lamprey to human. Front. Neurosci. 2017, 11, 295. [Google Scholar] [CrossRef] [Green Version]
- Ryczko, D.; Grätsch, S.; Alpert, M.H.; Cone, J.J.; Kasemir, J.; Ruthe, A.; Beauséjour, P.A.; Auclair, F.; Roitman, M.F.; Alford, S.; et al. Descending dopaminergic inputs to reticulospinal neurons promote locomotor movements. J. Neurosci. 2020, 40, 8478–8490. [Google Scholar] [CrossRef]
- Sánchez-Camacho, C.; Marín, O.; González, A. Distribution and origin of the catecholaminergic innervation in the amphibian mesencephalic tectum. Vis. Neurosci. 2002, 19, 321–333. [Google Scholar] [CrossRef]
- Tay, T.L.; Ronneberger, O.; Ryu, S.; Nitschke, R.; Driever, W. Comprehensive catecholaminergic projectome analysis reveals single-neuron integration of zebrafish ascending and descending dopaminergic systems. Nat. Commun. 2011, 2, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, K.J.; Takada, M. Bilateral tectal projection of single nigrostriatal dopamine cells in the rat. Neuroscience 1989, 33, 311–321. [Google Scholar] [CrossRef]
- Bolton, A.D.; Murata, Y.; Kirchner, R.; Kim, S.Y.; Young, A.; Dang, T.; Yanagawa, Y.; Constantine-Paton, M. A Diencephalic dopamine source provides input to the superior colliculus, where D1 and D2 receptors segregate to distinct functional zones. Cell Rep. 2015, 13, 1003–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montardy, Q.; Zhou, Z.; Li, L.; Yang, Q.; Lei, Z.; Feng, X.; Chen, S.; Shi, Q.; Zhang, H.; Chen, S.; et al. Dopamine modulates visual threat processing in the superior colliculus via D2 receptors. bioRxiv 2021. [Google Scholar] [CrossRef]
- Da Silva, J.A.; Tecuapetla, F.; Paixão, V.; Costa, R.M. Dopamine neuron activity before action initiation gates and invigorates future movements. Nature 2018, 554, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Tiklová, K.; Björklund, Å.K.; Lahti, L.; Fiorenzano, A.; Nolbrant, S.; Gillberg, L.; Volakakis, N.; Yokota, C.; Hilscher, M.M.; Hauling, T.; et al. Single-cell RNA sequencing reveals midbrain dopamine neuron diversity emerging during mouse brain development. Nat. Commun. 2019, 10, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, T.; Wang, H.L.; Li, X.; Ng, T.H.; Morales, M. Mesocorticolimbic glutamatergic pathway. J. Neurosci. 2011, 31, 8476–8490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, T.; Wang, H.L.; Morales, M. Glutamate neurons in the substantia nigra compacta and retrorubral field. Eur. J. Neurosci. 2013, 38, 3602–3610. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Qi, J.; Wang, H.L.; Zhang, S.; Morales, M. Glutamatergic and dopaminergic neurons in the mouse ventral tegmental area. Eur. J. Neurosci. 2015, 41, 760–772. [Google Scholar] [CrossRef] [Green Version]
- Morales, M.; Margolis, E.B. Ventral tegmental area: Cellular heterogeneity, connectivity and behaviour. Nat. Rev. Neurosci. 2017, 18, 73–85. [Google Scholar] [CrossRef]
- Root, D.H.; Wang, H.L.; Liu, B.; Barker, D.J.; Mód, L.; Szocsics, P.; Silva, A.C.; Maglóczky, Z.; Morales, M. Glutamate neurons are intermixed with midbrain dopamine neurons in nonhuman primates and humans. Sci. Rep. 2016, 6, 30615. [Google Scholar] [CrossRef] [Green Version]
- Dumas, S.; Wallén-Mackenzie, Å. Developmental co-expression of Vglut2 and Nurr1 in a mes-di-encephalic continuum preceeds dopamine and glutamate neuron specification. Front. Cell Dev. Biol. 2019, 7, 307. [Google Scholar] [CrossRef] [Green Version]
- Stuber, G.D.; Hnasko, T.S.; Britt, J.P.; Edwards, R.H.; Bonci, A. Dopaminergic terminals in the nucleus accumbens but not the dorsal striatum corelease glutamate. J. Neurosci. 2010, 30, 8229–8233. [Google Scholar] [CrossRef]
- Tecuapetla, F.; Patel, J.C.; Xenias, H.; English, D.; Tadros, I.; Shah, F.; Berlin, J.; Deisseroth, K.; Rice, M.E.; Tepper, J.M.; et al. Glutamatergic signaling by mesolimbic dopamine neurons in the nucleus accumbens. J. Neurosci. 2010, 30, 7105–7110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tritsch, N.X.; Ding, J.B.; Sabatini, B.L. Dopaminergic neurons inhibit striatal output through non-canonical release of GABA. Nature 2012, 490, 262–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Qi, J.; Li, X.; Wang, H.L.; Britt, J.P.; Hoffman, A.F.; Bonci, A.; Lupica, C.R.; Morales, M. Dopaminergic and glutamatergic microdomains in a subset of rodent mesoaccumbens axons. Nat. Neurosci. 2015, 18, 386–396. [Google Scholar] [CrossRef]
- Mongia, S.; Yamaguchi, T.; Liu, B.; Zhang, S.; Wang, H.; Morales, M. The Ventral Tegmental Area has calbindin neurons with the capability to co-release glutamate and dopamine into the nucleus accumbens. Eur. J. Neurosci. 2019, 50, 3968–3984. [Google Scholar] [CrossRef]
- Barbano, M.F.; Wang, H.L.; Zhang, S.; Miranda-Barrientos, J.; Estrin, D.J.; Figueroa-González, A.; Liu, B.; Barker, D.J.; Morales, M. VTA glutamatergic neurons mediate innate defensive behaviors. Neuron 2020, 107, 368–382. [Google Scholar] [CrossRef] [PubMed]
- Moriya, S.; Yamashita, A.; Kawashima, S.; Nishi, R.; Yamanaka, A.; Kuwaki, T. Acute aversive stimuli rapidly increase the activity of ventral tegmental area dopamine neurons in awake mice. Neuroscience 2018, 386, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Papathanou, M.; Creed, M.; Dorst, M.C.; Bimpisidis, Z.; Dumas, S.; Pettersson, H.; Bellone, C.; Silberberg, G.; Lüscher, C.; Wallén-Mackenzie, Å. Targeting VGLUT2 in mature dopamine neurons decreases mesoaccumbal glutamatergic transmission and identifies a role for glutamate co-release in synaptic plasticity by increasing baseline AMPA/NMDA ratio. Front. Neural Circuits 2018, 12, 64. [Google Scholar] [CrossRef]
- Bimpisidis, Z.; Wallén-Mackenzie, Å. Neurocircuitry of reward and addiction: Potential impact of dopamine–glutamate co-release as future target in substance use disorder. J. Clin. Med. 2019, 8, 1887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filippi, A.; Mueller, T.; Driever, W. Vglut2 and gad expression reveal distinct patterns of dual GABAergic versus glutamatergic cotransmitter phenotypes of dopaminergic and noradrenergic neurons in the zebrafish brain. J. Comp. Neurol. 2014, 522, 2019–2037. [Google Scholar] [CrossRef] [Green Version]
- Ryczko, D.; Grätsch, S.; Schläger, L.; Keuyalian, A.; Boukhatem, Z.; Garcia, C.; Auclair, F.; Buschges, A.; Dubuc, R. Nigral glutamatergic neurons control the speed of locomotion. J. Neurosci. 2017, 37, 9759–9770. [Google Scholar] [CrossRef]
- Verney, C.; Zecevic, N.; Puelles, L. Structure of longitudinal brain zones that provide the origin for the substantia nigra and ventral tegmental area in human embryos, as revealed by cytoarchitecture and tyrosine hydroxylase, calretinin, calbindin, and GABA immunoreactions. J. Comp. Neurol. 2001, 429, 22–44. [Google Scholar] [CrossRef]
- Puelles, E.; Martínez-de-la-Torre, M.; Watson, C.; Puelles, L. Midbrain. In The Mouse Nervous System; Watson, C., Paxinos, G., Puelles, L., Eds.; Elsevier Inc.: San Diego, CA, USA, 2012; pp. 337–359. [Google Scholar]
- Wullimann, M.F. Ancestry of basal ganglia circuits: New evidence in teleosts. J. Comp. Neurol. 2014, 522, 2013–2018. [Google Scholar] [CrossRef]
- Smeets, W.J.A.J.; Marin, O.; Gonzalez, A. Evolution of the basal ganglia: New perspectives through a comparative approach. J. Anat. 2000, 196, 501–517. [Google Scholar] [CrossRef] [Green Version]
- Marín, O.; González, A.; Smeets, W.J.A.J. Evidence for a mesolimbic pathway in anuran amphibians: A combined tract-tracing/immunohistochemical study. Neurosci. Lett. 1995, 190, 183–186. [Google Scholar] [CrossRef]
- Marín, O.; Smeets, W.J.A.J.; González, A. Basal ganglia organization in amphibians: Catecholaminergic innervation of the striatum and the nucleus accumbens. J. Comp. Neurol. 1997, 378, 50–69. [Google Scholar] [CrossRef]
- Freudenmacher, L.; Schauer, M.; Walkowiak, W.; von Twickel, A. Refinement of the dopaminergic system of anuran amphibians based on connectivity with habenula, basal ganglia, limbic system, pallium, and spinal cord. J. Comp. Neurol. 2020, 528, 972–988. [Google Scholar] [CrossRef]
- Carrera, I.; Sueiro, C.; Molist, P.; Ferreiro, S.; Adrio, F.; Rodríguez, M.A.; Anadón, R.; Rodríguez-Moldes, I. Temporal and spatial organization of tyrosine hydroxylase-immunoreactive cell groups in the embryonic brain of an elasmobranch, the lesser-spotted dogfish Scyliorhinus canicula. Brain Res. Bull. 2005, 66, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Quintana-Urzainqui, I.; Sueiro, C.; Carrera, I.; Ferreiro-Galve, S.; Santos-Durn, G.; Pose-Mndez, S.; Mazan, S.; Candal, E.; Rodrguez-Moldes, I. Contributions of developmental studies in the dogfish Scyliorhinus canicula to the brain anatomy of elasmobranchs: Insights on the basal ganglia. Brain. Behav. Evol. 2012, 80, 127–141. [Google Scholar] [CrossRef] [PubMed]
- Rink, E.; Wullimann, M.F. Connections of the ventral telencephalon and tyrosine hydroxylase distribution in the zebrafish brain (Danio rerio) lead to identification of an ascending dopaminergic system in a teleost. Brain Res. Bull. 2002, 57, 385–387. [Google Scholar] [CrossRef]
- Vernier, P.; Wullimann, M.F. Evolution of the Posterior Tuberculum and Preglomerular Nuclear Complex. In Encyclopedia of Neuroscience; Binder, M.D., Hirokawa, N., Windhorst, U., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1404–1413. [Google Scholar]
- Reinig, S.; Driever, W.; Arrenberg, A.B. The descending diencephalic dopamine system is tuned to sensory stimuli. Curr. Biol. 2017, 27, 318–333. [Google Scholar] [CrossRef] [Green Version]
- Wullimann, M.F.; Umeasalugo, K.E. Sonic hedgehog expression in zebrafish forebrain identifies the teleostean pallidal signaling center and shows preglomerular complex and posterior tubercular dopamine cells to arise from shh cells. J. Comp. Neurol. 2020, 528, 1321–1348. [Google Scholar] [CrossRef] [Green Version]
- Blaess, S.; Bodea, G.O.; Kabanova, A.; Chanet, S.; Mugniery, E.; Derouiche, A.; Stephen, D.; Joyner, A.L. Temporal-spatial changes in Sonic Hedgehog expression and signaling reveal different potentials of ventral mesencephalic progenitors to populate distinct ventral midbrain nuclei. Neural Dev. 2011, 6, 29. [Google Scholar] [CrossRef]
- Wullimann, M.F. Neural origins of basal diencephalon in teleost fishes: Radial versus tangential migration. J. Morphol. 2020, 281, 1133–1141. [Google Scholar] [CrossRef]
- Blin, M.; Norton, W.; Bally-Cuif, L.; Vernier, P. NR4A2 controls the differentiation of selective dopaminergic nuclei in the zebrafish brain. Mol. Cell. Neurosci. 2008, 39, 592–604. [Google Scholar] [CrossRef]
- Lozano, D.; Morona, R.; González, A.; López, J.M. Comparative analysis of the organization of the catecholaminergic systems in the brain of holostean fishes (Actinopterygii/Neopterygii). Brain. Behav. Evol. 2019, 93, 206–235. [Google Scholar] [CrossRef]
- López, J.M.; González, A. Organization of the catecholaminergic systems in the brain of lungfishes, the closest living relatives of terrestrial vertebrates. J. Comp. Neurol. 2017, 525, 3083–3109. [Google Scholar] [CrossRef]
- Strausfeld, N.J.; Hirth, F. Deep homology of arthropod central complex and vertebrate basal ganglia. Science 2013, 340, 157–161. [Google Scholar] [CrossRef] [Green Version]
- Moret, F.; Guilland, J.C.; Coudouel, S.; Rochette, L.; Vernier, P. Distribution of tyrosine hydroxylase, dopamine, and serotonin in the central nervous system of amphioxus (Branchiostoma lanceolatum): Implications for the evolution of catecholamine systems in vertebrates. J. Comp. Neurol. 2004, 468, 135–150. [Google Scholar] [CrossRef]
- Zieger, E.; Lacalli, T.C.; Pestarino, M.; Schubert, M.; Candiani, S. The origin of dopaminergic systems in chordate brains: Insights from amphioxus. Int. J. Dev. Biol. 2017, 61, 749–761. [Google Scholar] [CrossRef]
- Rodríguez-López, C.; Clascá, F.; Prensa, L. The mesoaccumbens pathway: A retrograde labeling and single-cell axon tracing analysis in the mouse. Front. Neuroanat. 2017, 11, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watabe-Uchida, M.; Zhu, L.; Ogawa, S.K.; Vamanrao, A.; Uchida, N. Whole-brain mapping of direct inputs to midbrain dopamine neurons. Neuron 2012, 74, 858–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, Y.; Moghimyfiroozabad, S.; Moghimyfiroozabad, A.; Tierney, T.S.; Alavian, K.N. The factors for the early and late development of midbrain dopaminergic neurons segregate into two distinct evolutionary clusters. Brain Disord. 2021, 1, 100002. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Fernández, J.; Barandela, M.; Jiménez-López, C. The Dopaminergic Control of Movement-Evolutionary Considerations. Int. J. Mol. Sci. 2021, 22, 11284. https://doi.org/10.3390/ijms222011284
Pérez-Fernández J, Barandela M, Jiménez-López C. The Dopaminergic Control of Movement-Evolutionary Considerations. International Journal of Molecular Sciences. 2021; 22(20):11284. https://doi.org/10.3390/ijms222011284
Chicago/Turabian StylePérez-Fernández, Juan, Marta Barandela, and Cecilia Jiménez-López. 2021. "The Dopaminergic Control of Movement-Evolutionary Considerations" International Journal of Molecular Sciences 22, no. 20: 11284. https://doi.org/10.3390/ijms222011284
APA StylePérez-Fernández, J., Barandela, M., & Jiménez-López, C. (2021). The Dopaminergic Control of Movement-Evolutionary Considerations. International Journal of Molecular Sciences, 22(20), 11284. https://doi.org/10.3390/ijms222011284