
Inhibition of Spinal TRPV1 Reduces NMDA Receptor 2B Phosphorylation and Produces Anti-Nociceptive Effects in Mice with Inflammatory Pain


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Development of Nociceptive Behavior Using Carrageenan
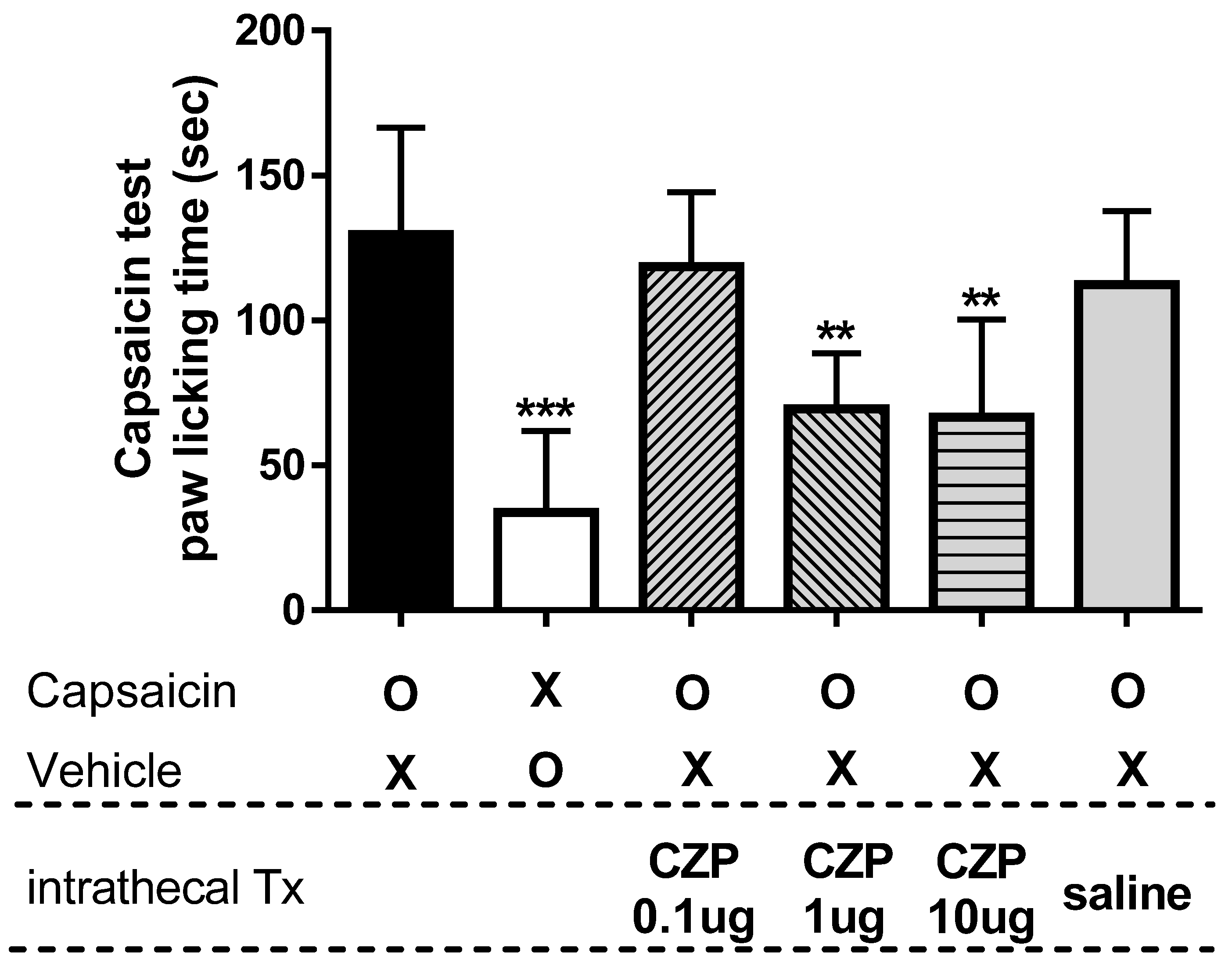
2.2. Dose-Dependency of Capsazepine in Capsaicin-Induced Spontaneous Pain Test
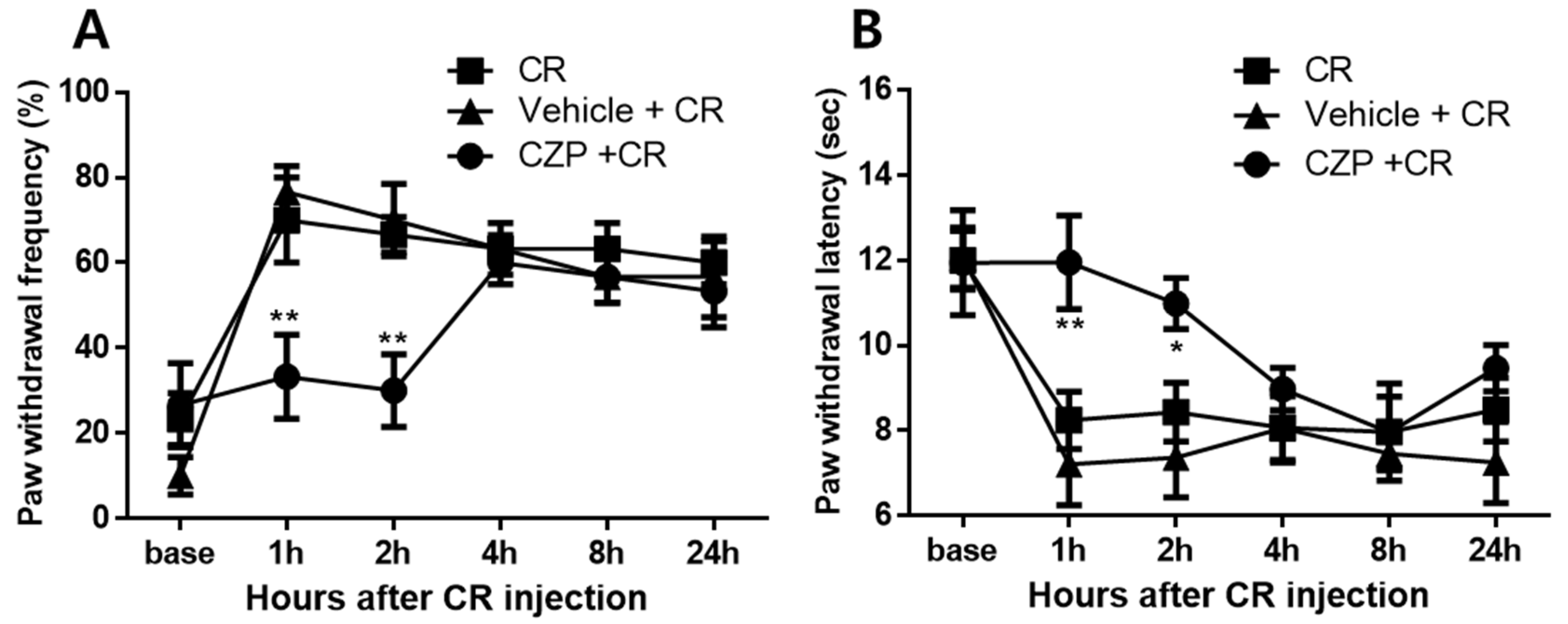
2.3. Anti-Nociceptive Effects of Intrathecal Capsazepine Administration
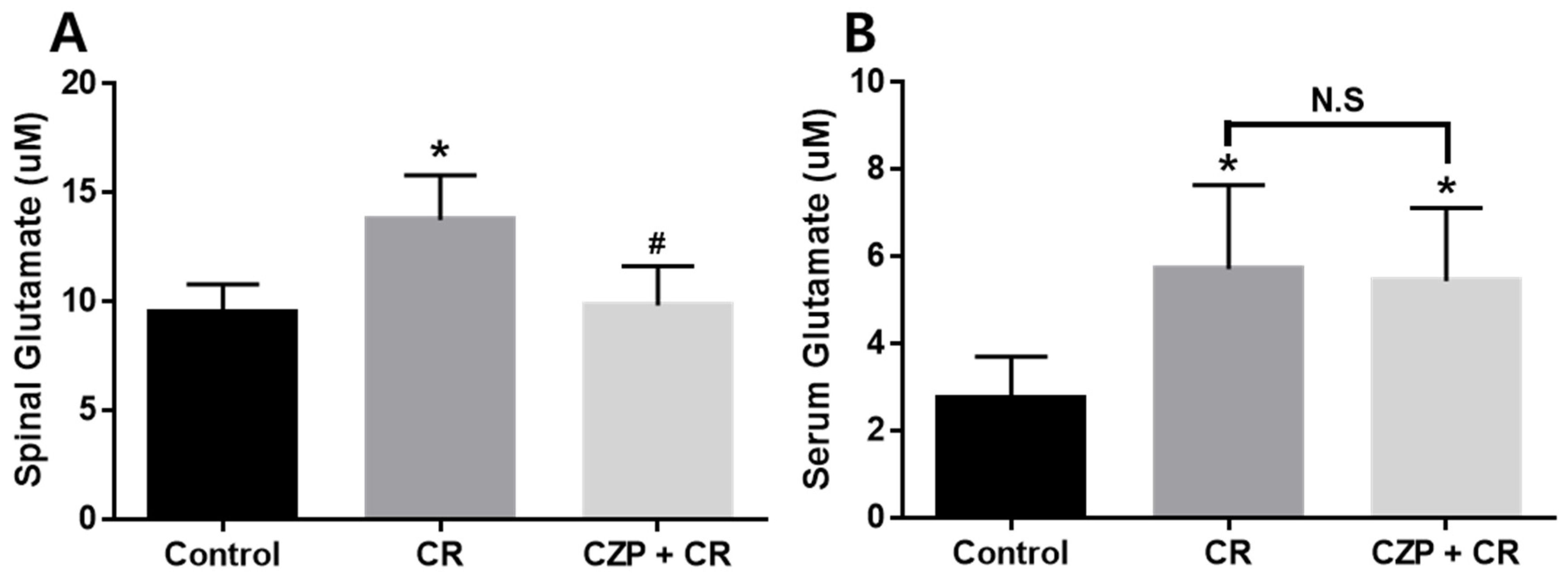
2.4. Concentration of Glutamate in the Spinal Cord and Serum
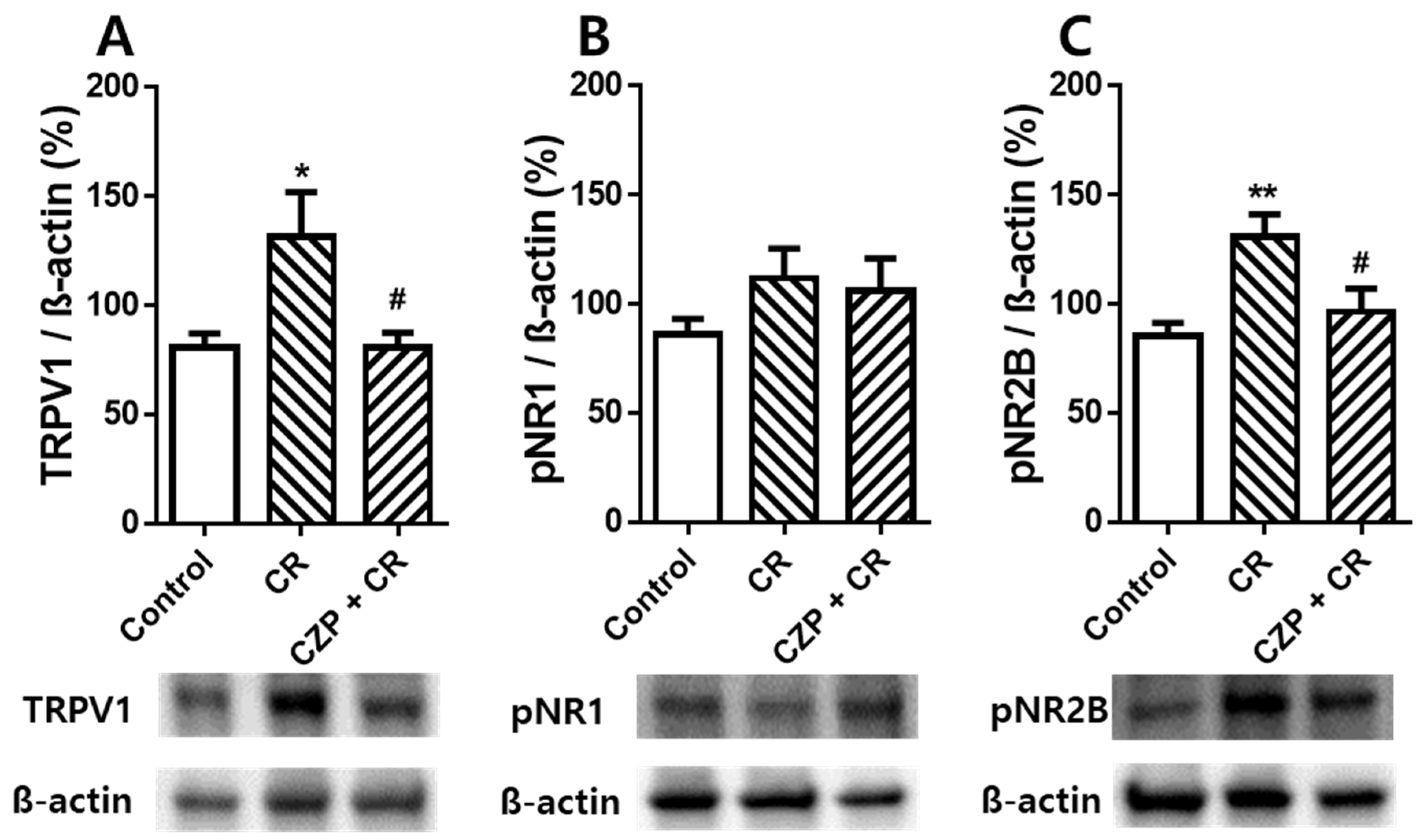
2.5. Expression of TRPV1, pNR1, and pNR2B in the Spinal Dorsal Horn
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Carrageenan-Induced Inflammatory Pain
4.3. Evaluation of Nociceptive Behaviors
4.4. Capsaicin-Induced Spontaneous Pain Test and Drug Injection
4.5. Glutamate Concentration Measurements
4.6. Western Blot and Image Analysis
4.7. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferrandiz-Huertas, C.; Mathivanan, S.; Wolf, C.J.; Devesa, I.; Ferrer-Montiel, A. Trafficking of ThermoTRP Channels. Membranes 2014, 4, 525–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandewauw, I.; De Clercq, K.; Mulier, M.; Held, K.; Pinto, S.; Van Ranst, N.; Segal, A.; Voet, T.; Vennekens, R.; Zimmermann, K.; et al. A TRP channel trio mediates acute noxious heat sensing. Nature 2018, 555, 662–666. [Google Scholar] [CrossRef] [PubMed]
- Silverman, H.A.; Chen, A.; Kravatz, N.L.; Chavan, S.S.; Chang, E.H. Involvement of Neural Transient Receptor Potential Channels in Peripheral Inflammation. Front. Immunol. 2020, 11, 590261. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, E.; Luongo, L.; de Novellis, V.; Rossi, F.; Marabese, I.; Maione, S. Transient receptor potential vanilloid type 1 and pain development. Curr. Opin. Pharmacol. 2012, 12, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Chen, S.; Chen, H.; Pan, H. Endogenous transient receptor potential ankyrin 1 and vanilloid 1 activity potentiates glutamatergic input to spinal lamina I neurons in inflammatory pain. J. Neurochem. 2019, 149, 381–398. [Google Scholar] [CrossRef] [PubMed]
- Fajrin, F.A.; Nugroho, A.E.; Nurrochmad, A.; Susilowati, R. Ginger extract and its compound, 6-shogaol, attenuates painful diabetic neuropathy in mice via reducing TRPV1 and NMDAR2B expressions in the spinal cord. J. Ethnopharmacol. 2020, 249, 112396. [Google Scholar] [CrossRef]
- Vyklický, L.; Nováková-Toušová, K.; Benedikt, J.; Samad, A.; Touška, F.; Vlachova, V. Calcium-dependent desensitization of vanilloid receptor TRPV1: A mechanism possibly involved in analgesia induced by topical application of capsaicin. Physiol. Res. 2008, 57, S59–S68. [Google Scholar] [CrossRef]
- Keeble, J.; Russell, F.; Curtis, B.; Starr, A.; Pinter, E.; Brain, S.D. Involvement of transient receptor potential vanilloid 1 in the vascular and hyperalgesic components of joint inflammation. Arthritis Rheum. 2005, 52, 3248–3256. [Google Scholar] [CrossRef]
- Jay, G.W.; Barkin, R.L. Neuropathic pain: Etiology, pathophysiology, mechanisms, and evaluations. Disease-a-Month 2014, 60, 6–47. [Google Scholar] [CrossRef]
- Zhou, H.-Y.; Chen, S.-R.; Pan, H.-L. TargetingN-methyl-D-aspartate receptors for treatment of neuropathic pain. Expert Rev. Clin. Pharmacol. 2011, 4, 379–388. [Google Scholar] [CrossRef] [Green Version]
- Vranken, J.H. Mechanisms and Treatment of Neuropathic Pain. Cent. Nerv. Syst. Agents Med. Chem. 2009, 9, 71–78. [Google Scholar] [CrossRef]
- Paoletti, P.; Neyton, J. NMDA receptor subunits: Function and pharmacology. Curr. Opin. Pharmacol. 2007, 7, 39–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benarroch, E.E. NMDA receptors: Recent insights and clinical correlations. Neurology 2011, 76, 1750–1757. [Google Scholar] [CrossRef] [PubMed]
- Tan, P.H.; Chia, Y.Y.; Chow, L.H.; Chen, J.J.; Yang, L.C.; Hung, K.C.; Chen, H.S.; Kuo, C.H. Gene knockdown of the N -Methyl-d-Aspartate receptor NR1 subunit with subcutaneous small interfering RNA reduces inflammation-induced nociception in rats. Anesthesiology 2010, 112, 1482–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.W.; Kang, S.Y.; Choi, J.G.; Kang, N.W.; Kim, S.J.; Lee, S.D.; Park, J.B.; Ryu, Y.-H.; Kim, H.W. Analgesic effect of electroacupuncture on paclitaxel-induced neuropathic pain via spinal opioidergic and adrenergic mechanisms in mice. Am. J. Chin. Med. 2015, 43, 57–70. [Google Scholar] [CrossRef]
- Kang, S.Y.; Bang, S.K.; Kwon, O.S.; Seo, S.Y.; Choi, K.H.; Cho, S.J.; Yoo, H.S.; Lee, J.S.; Kim, H.W.; Ryu, Y. Treatment of electrical wrist stimulation reduces chemotherapy-induced neuropathy and ultrasound vocalization via modulation of spinal NR2B phosphorylation. Brain Res. Bull. 2020, 162, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Salter, M.W.; Dong, Y.; Kalia, L.V.; Liu, X.J.; Pitcher, G. Regulation of NMDA receptors by kinases and phosphatases. In Advances in Vagal Afferent Neurobiology; Biology of the NMDA Receptor; Undem, B.J., Weinreich, D., Eds.; CRC Press; Taylor & Francis: Boca Raton, FL, USA, 2009; Chapter 7. [Google Scholar]
- Mellström, B.; Savignac, M.; Gomez-Villafuertes, R.; Naranjo, J.R. Ca2+-Operated transcriptional networks: Molecular mechanisms and in vivo models. Physiol. Rev. 2008, 88, 421–449. [Google Scholar] [CrossRef] [Green Version]
- Sandkühler, J. Models and mechanisms of hyperalgesia and allodynia. Physiol. Rev. 2009, 89, 707–758. [Google Scholar] [CrossRef]
- Alawi, K.; Keeble, J. The paradoxical role of the transient receptor potential vanilloid 1 receptor in inflammation. Pharmacol. Ther. 2010, 125, 181–195. [Google Scholar] [CrossRef]
- Watanabe, M.; Ueda, T.; Shibata, Y.; Kumamoto, N.; Ugawa, S. The role of TRPV1 channels in carrageenan-induced mechanical hyperalgesia in mice. NeuroReport 2015, 26, 173–178. [Google Scholar] [CrossRef]
- Walker, K.M.; Urban, L.; Medhurst, S.J.; Patel, S.; Panesar, M.; Fox, A.J.; McIntyre, P. The VR1 antagonist capsazepine reverses mechanical hyperalgesia in models of inflammatory and neuropathic pain. J. Pharmacol. Exp. Ther. 2003, 304, 56–62. [Google Scholar] [CrossRef]
- Antoniazzi, C.; Nassini, R.; Rigo, F.K.; Milioli, A.M.; Bellinaso, F.; Camponogara, C.; Silva, C.R.; De Almeida, A.S.; Rossato, M.F.; De Logu, F.; et al. Transient receptor potential ankyrin 1 (TRPA1) plays a critical role in a mouse model of cancer pain. Int. J. Cancer 2018, 144, 355–365. [Google Scholar] [CrossRef]
- Cun-Jin, S.; Jian-Hao, X.; Xu, L.; Feng-Lun, Z.; Jie, P.; Ai-Ming, S.; Duan-Min, H.; Yun-Li, Y.; Tong, L.; Yu-Song, Z. X-ray induces mechanical and heat allodynia in mouse via TRPA1 and TRPV1 activation. Mol. Pain 2019, 15, 1744806919849201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, C.; Liu, P.; Zhao, Q.; Guo, S.; Wang, G. TRPV1 channel contributes to remifentanil-induced postoperative hyperalgesia via regulation of NMDA receptor trafficking in dorsal root ganglion. J. Pain Res. 2019, 12, 667–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, G.; Kekesi, G.; Nagy, E.; Benedek, G. The role of TRPV1 receptors in the antinociceptive effect of anandamide at spinal level. Pain 2008, 134, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Bujak, J.; Kosmala, D.; Szopa, I.M.; Majchrzak, K.; Bednarczyk, P. Inflammation, cancer and immunity—Implication of TRPV1 channel. Front. Oncol. 2019, 9, 1087. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Kang, J.; Lin, B.; Li, J.; Zhu, Y.; Du, J.; Yang, X.; Xi, Z.; Li, R. Mediating role of TRPV1 Ion channels in the Co-exposure to PM2.5 and formaldehyde of Balb/c mice asthma model. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.Y.; Lee, H.Y.; Hur, J.; Kim, K.H.; Kang, J.Y.; Rhee, C.K.; Lee, S.Y. TRPV1 blocking alleviates airway inflammation and remodeling in a chronic asthma murine model. Allergy Asthma Immunol. Res. 2018, 10, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Ninomiya, Y.; Tanuma, S.-I.; Tsukimoto, M. Differences in the effects of four TRPV1 channel antagonists on lipopolysaccharide-induced cytokine production and COX-2 expression in murine macrophages. Biochem. Biophys. Res. Commun. 2017, 484, 668–674. [Google Scholar] [CrossRef]
- Wozniak, K.M.; Rojas, C.; Wu, Y.W.; Slusher, B.S. The role of glutamate signaling in pain processes and its regulation by GCP II inhibition. Curr. Med. Chem. 2012, 19, 1323–1334. [Google Scholar] [CrossRef]
- Jin, Y.; Nishioka, H.; Wakabayashi, K.; Fujita, T.; Yonehara, N. Effect of morphine on the release of excitatory amino acids in the rat hind instep: Pain is modulated by the interaction between the peripheral opioid and glutamate systems. Neuroscience 2006, 138, 1329–1339. [Google Scholar] [CrossRef]
- Arundine, M.; Tymianski, M. Molecular mechanisms of calcium-dependent neurodegeneration in excitotoxicity. Cell Calcium 2003, 34, 325–337. [Google Scholar] [CrossRef]
- Petrenko, A.B.; Yamakura, T.; Baba, H.; Shimoji, K. The role of N-Methyl-d-Aspartate (NMDA) receptors in pain: A review. Anesth. Analg. 2003, 97, 1108–1116. [Google Scholar] [CrossRef] [PubMed]
- Rygh, L.J.; Svendsen, F.; Hole, K.; Tjølsen, A. Increased spinal N -methyl-D-aspartate receptor function after 20 h of carrageenan-induced inflammation. Pain 2001, 93, 15–21. [Google Scholar] [CrossRef]
- Eisenberg, E.; LaCross, S.; Strassman, A.M. The effects of the clinically tested NMDA receptor antagonist memantine on carrageenan-induced thermal hyperalgesia in rats. Eur. J. Pharmacol. 1994, 255, 123–129. [Google Scholar] [CrossRef]
- Kang, S.-Y.; Roh, D.-H.; Yoon, S.-Y.; Moon, J.-Y.; Kim, H.-W.; Lee, H.-J.; Beitz, A.J.; Lee, J.-H. Repetitive treatment with diluted bee venom reduces neuropathic pain via potentiation of locus coeruleus noradrenergic neuronal activity and modulation of spinal NR1 phosphorylation in rats. J. Pain 2012, 13, 155–166. [Google Scholar] [CrossRef]
- Choi, J.G.; Kang, S.Y.; Kim, J.M.; Roh, D.H.; Yoon, S.Y.; Park, J.B.; Lee, J.H.; Kim, H.W. Antinociceptive effect of cyperi rhizoma and corydalis tuber extracts on neuropathic pain in rats. Korean J. Physiol. Pharmacol. 2012, 16, 387–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caudle, R.M.; Perez, F.M.; Del Valle-Pinero, A.Y.; Iadarola, M.J. Spinal cord NR1 serine phosphorylation and NR2B subunit suppression following peripheral inflammation. Mol. Pain 2005, 1, 25. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Saloman, J.L.; Weiland, G.; Auh, Q.-S.; Chung, M.-K.; Ro, J.Y. Functional interactions between NMDA receptors and TRPV1 in trigeminal sensory neurons mediate mechanical hyperalgesia in the rat masseter muscle. Pain 2012, 153, 1514–1524. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.S.; Roh, D.H.; Yoon, S.Y.; Moon, J.Y.; Choi, S.R.; Kwon, S.G.; Kang, S.Y.; Han, H.J.; Kim, H.W.; Beitz, A.J.; et al. Microglial interleukin-1beta in the ipsilateral dorsal horn inhibits the development of mirror-image contralateral mechanical allodynia through astrocyte activation in a rat model of inflammatory pain. Pain 2015, 156, 1046–1059. [Google Scholar] [CrossRef]
- de Almeida, T.B.; de Oliveira, R.G.; Gonçalves, G.M.; de Oliveira, J.M.; dos Santos, B.B.N.; Laureano-Melo, R.; da Silva Côrtes, W.; do Nascimento França, T.; de Almeida Vasconcellos, M.L.A.; Marinho, B.G. Evaluation of the antinociceptive and anti-inflammatory activities of piperic acid: Involvement of the cholinergic and vanilloid systems. Eur. J. Pharmacol. 2018, 834, 54–64. [Google Scholar]
- Kang, S.-Y.; Roh, D.-H.; Choi, J.-W.; Ryu, Y.; Lee, J.-H. repetitive treatment with diluted bee venom attenuates the induction of below-level neuropathic pain behaviors in a rat spinal cord injury model. Toxins 2015, 7, 2571–2585. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, S.-Y.; Seo, S.Y.; Bang, S.K.; Cho, S.J.; Choi, K.-H.; Ryu, Y. Inhibition of Spinal TRPV1 Reduces NMDA Receptor 2B Phosphorylation and Produces Anti-Nociceptive Effects in Mice with Inflammatory Pain. Int. J. Mol. Sci. 2021, 22, 11177. https://doi.org/10.3390/ijms222011177
Kang S-Y, Seo SY, Bang SK, Cho SJ, Choi K-H, Ryu Y. Inhibition of Spinal TRPV1 Reduces NMDA Receptor 2B Phosphorylation and Produces Anti-Nociceptive Effects in Mice with Inflammatory Pain. International Journal of Molecular Sciences. 2021; 22(20):11177. https://doi.org/10.3390/ijms222011177
Chicago/Turabian StyleKang, Suk-Yun, Su Yeon Seo, Se Kyun Bang, Seong Jin Cho, Kwang-Ho Choi, and Yeonhee Ryu. 2021. "Inhibition of Spinal TRPV1 Reduces NMDA Receptor 2B Phosphorylation and Produces Anti-Nociceptive Effects in Mice with Inflammatory Pain" International Journal of Molecular Sciences 22, no. 20: 11177. https://doi.org/10.3390/ijms222011177
APA StyleKang, S.-Y., Seo, S. Y., Bang, S. K., Cho, S. J., Choi, K.-H., & Ryu, Y. (2021). Inhibition of Spinal TRPV1 Reduces NMDA Receptor 2B Phosphorylation and Produces Anti-Nociceptive Effects in Mice with Inflammatory Pain. International Journal of Molecular Sciences, 22(20), 11177. https://doi.org/10.3390/ijms222011177