
S100A8/A9 Enhances Immunomodulatory and Tissue-Repairing Properties of Human Amniotic Mesenchymal Stem Cells in Myocardial Ischemia-Reperfusion Injury


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
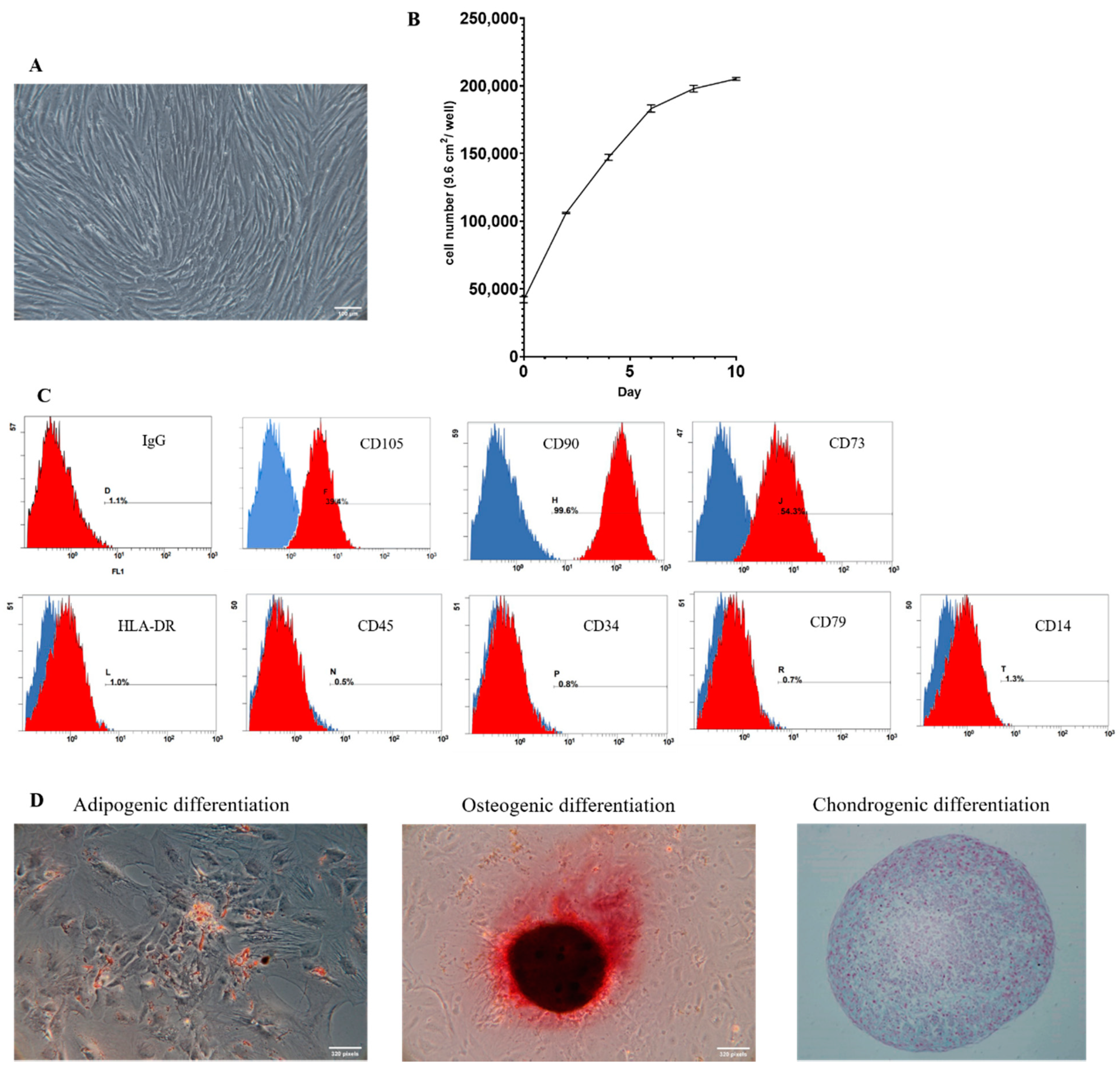
2.1. Identification and Characterization of hAMSCs
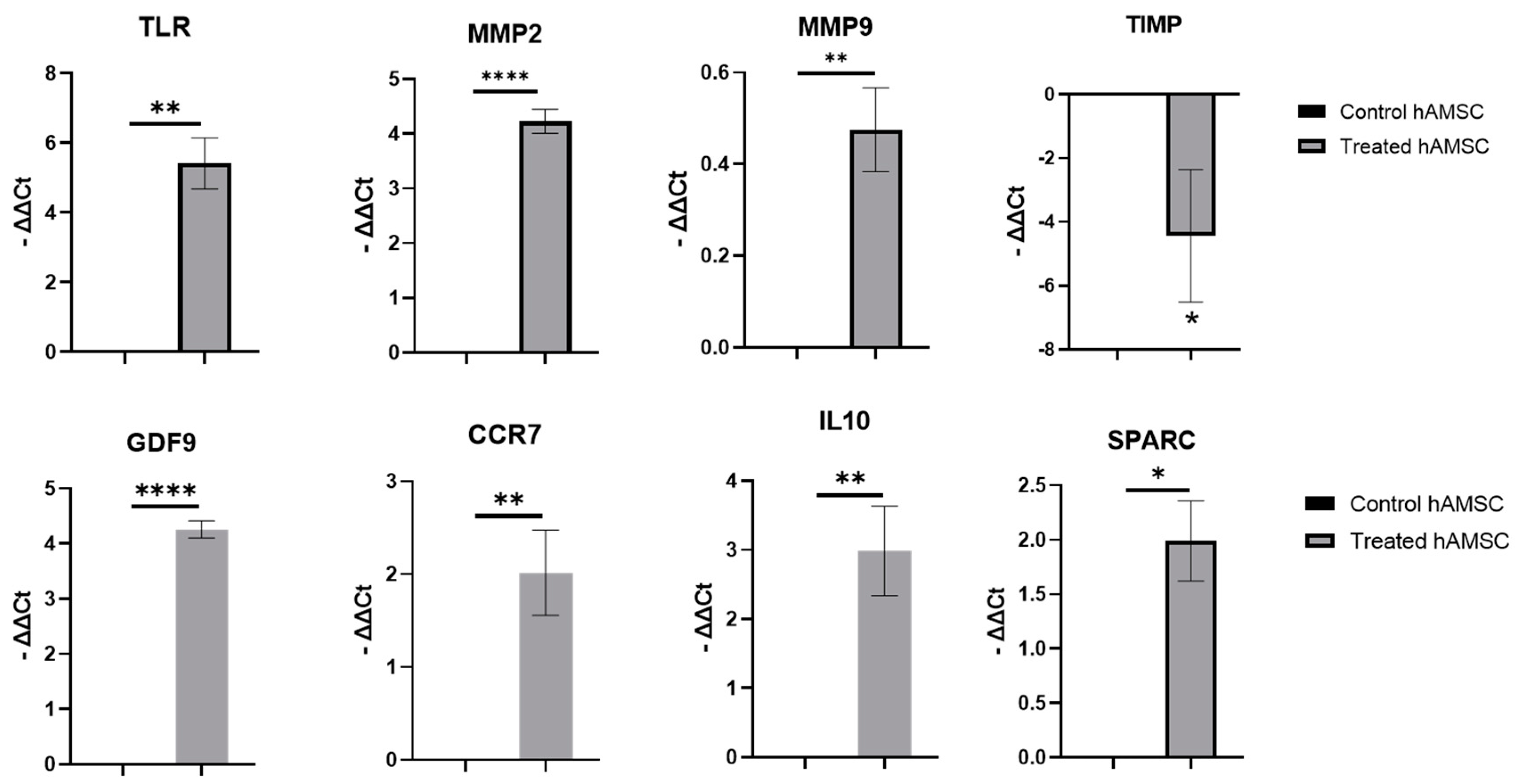
2.2. S100A8/A9 Pretreatment of hAMSCs
2.3. H/R Injury of nCMs In Vitro
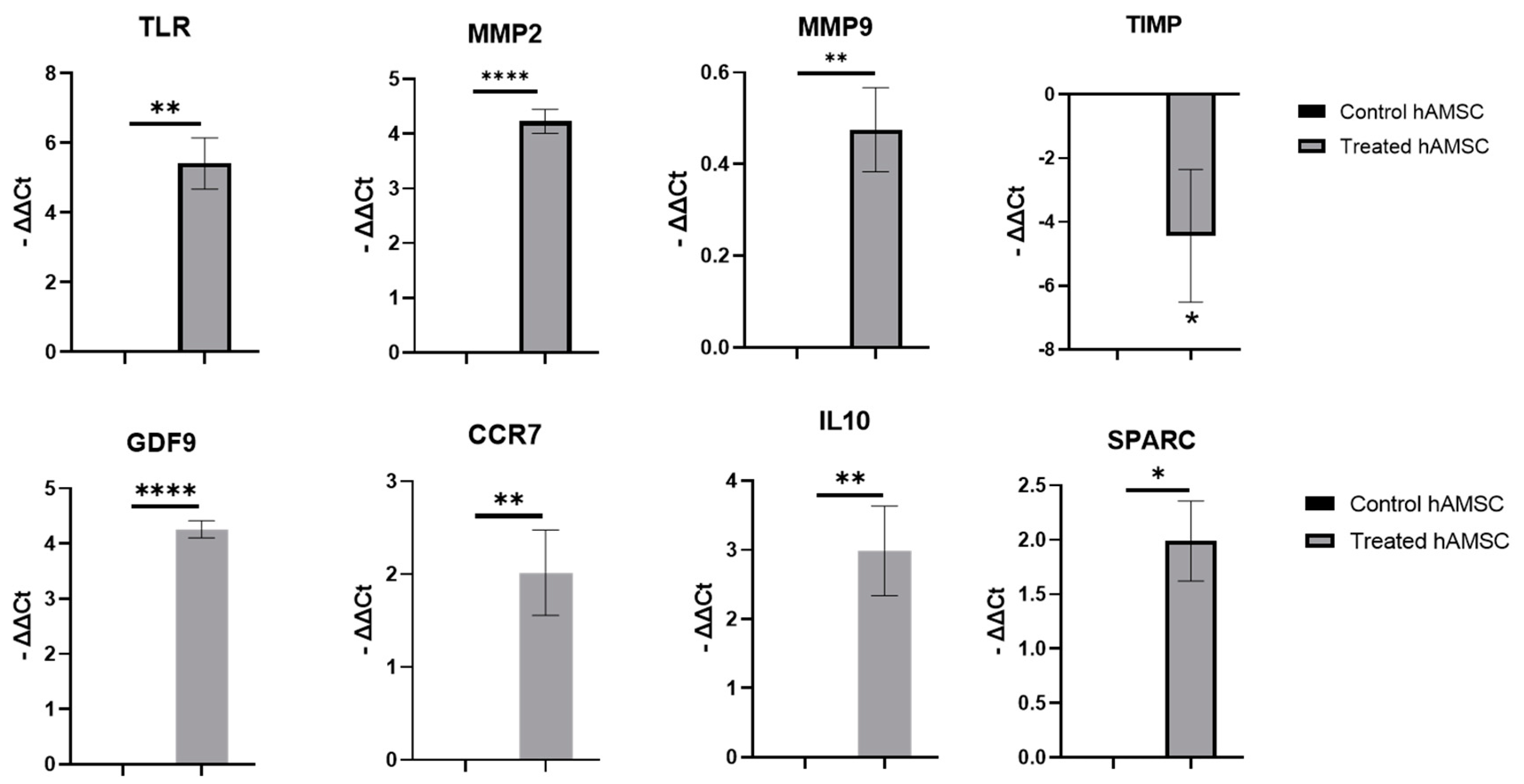
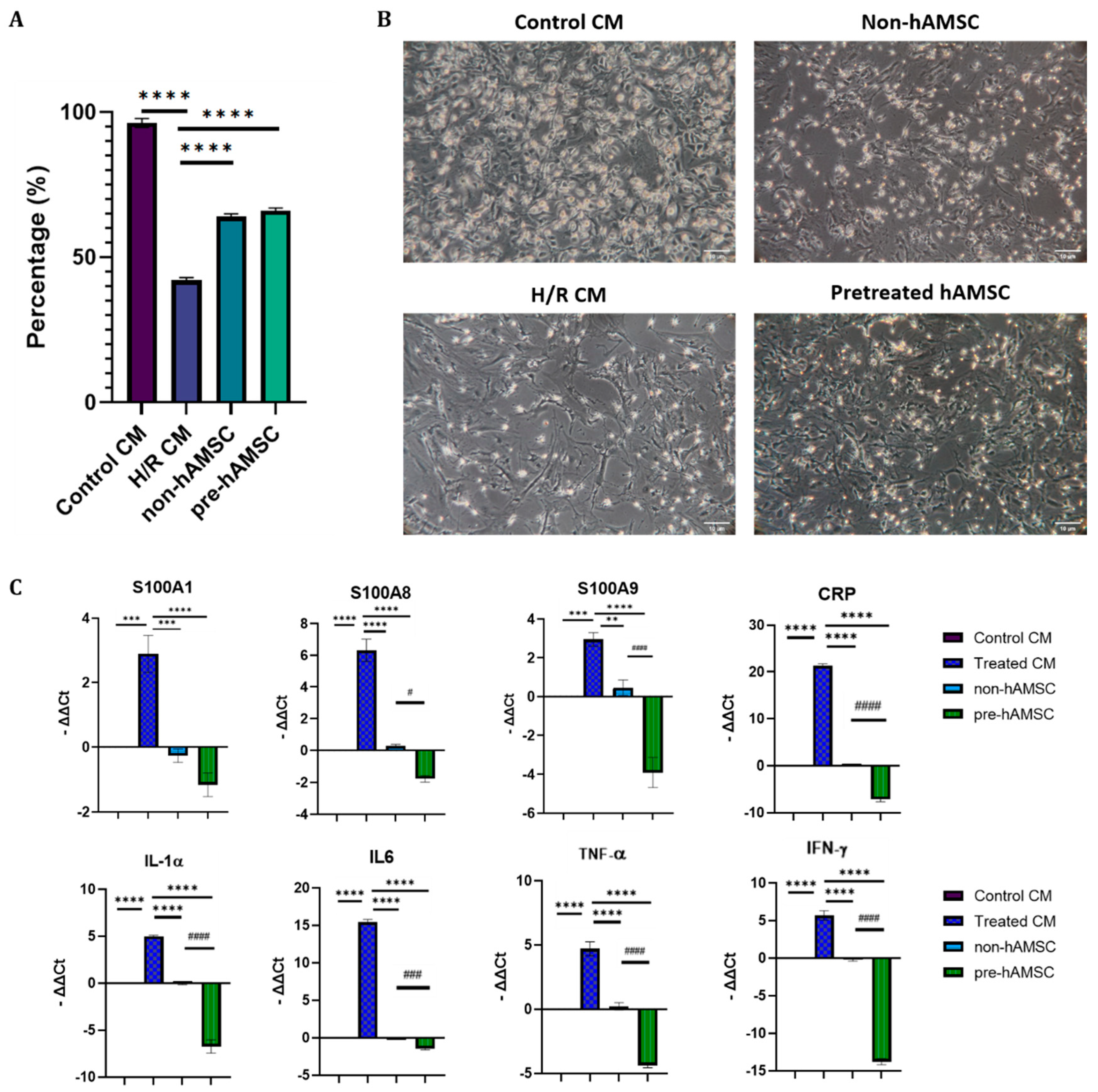
2.4. S100A8/A9 Pretreatment Enhanced the Anti-Inflammatory Effects of hAMSC-Derived Conditioned Medium in the H/R Model
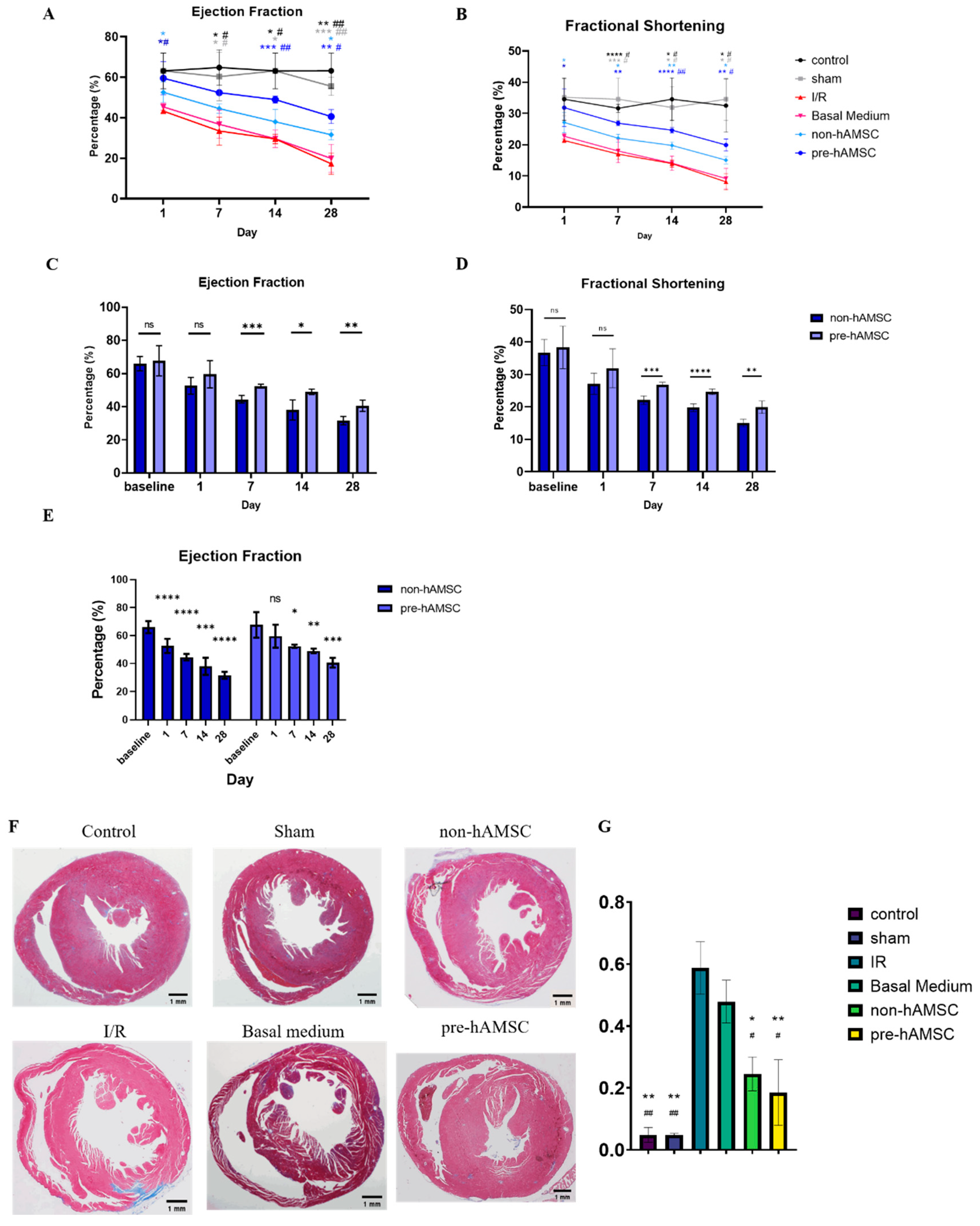
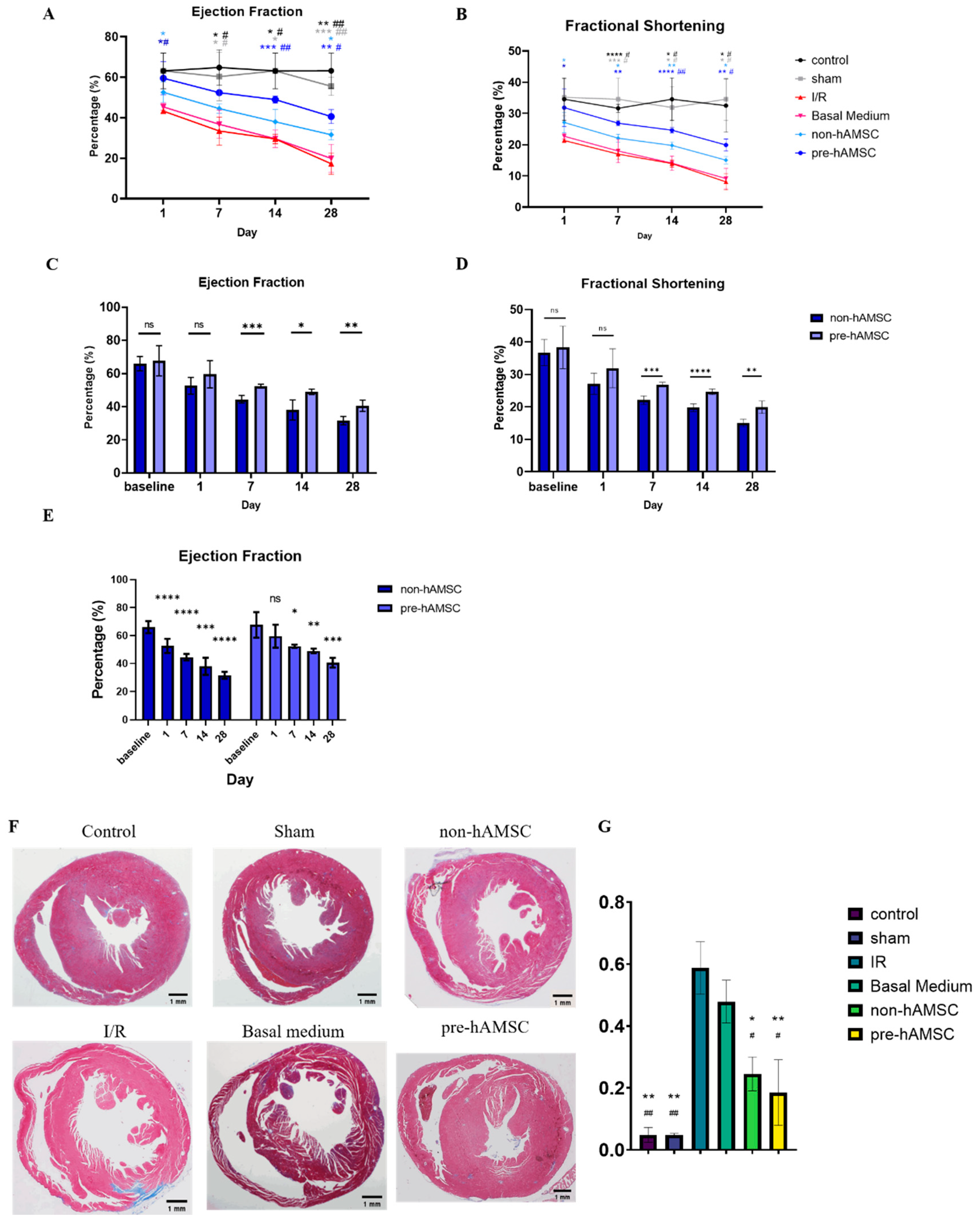
2.5. S100A8/A9 Pretreatment Enhanced the Therapeutic Effects of hAMSC-Derived Conditioned Medium in the Murine I/R Model
3. Discussion
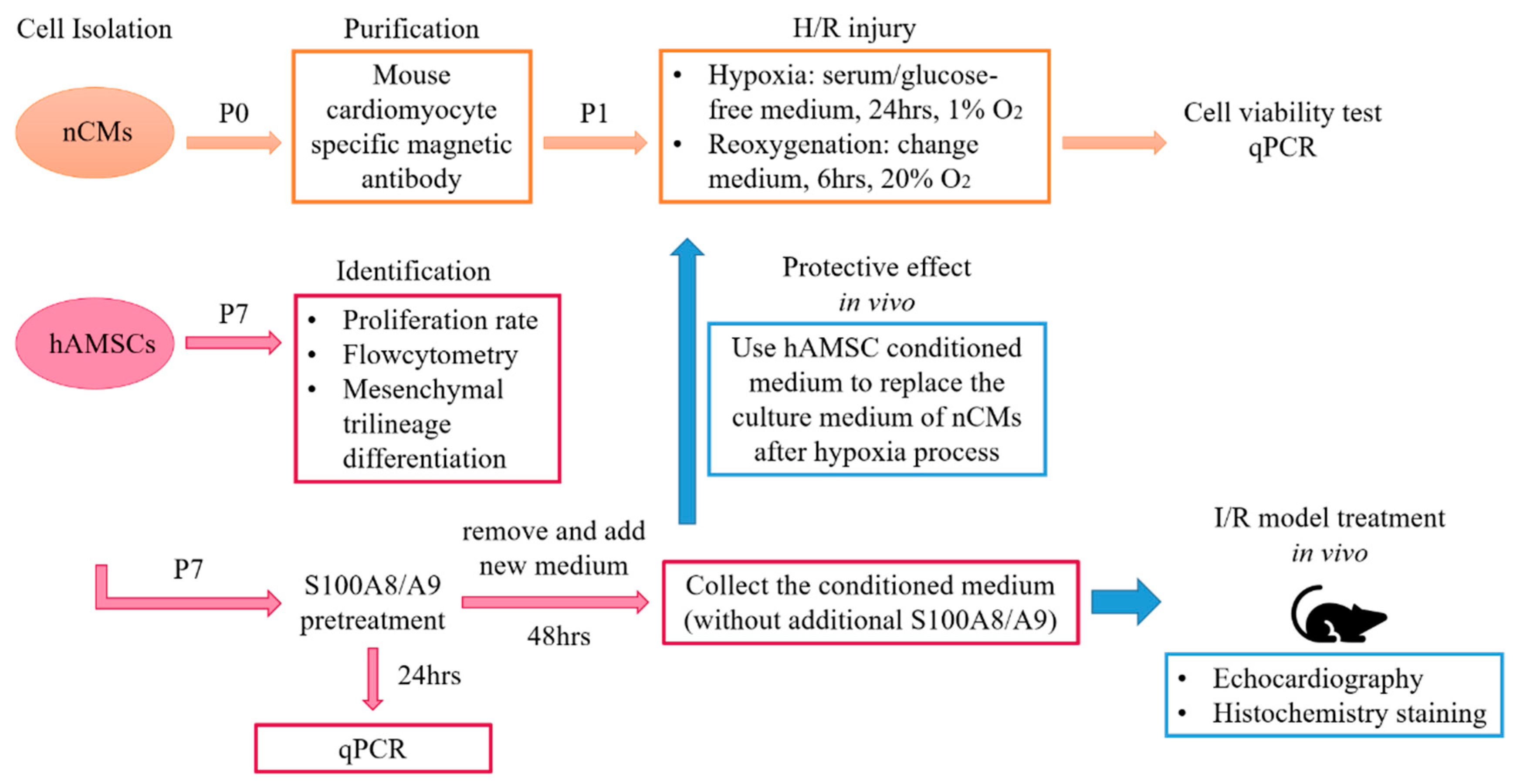
4. Materials & Methods
4.1. Animals
4.2. Preparation of hAMSCs
4.3. Preparation of nCMs
4.4. H/R Injury Induction of nCMs
4.5. S100A8/A9 Pretreatment of hAMSCs
4.6. Cell Proliferation Test
4.7. Mesenchymal Trilineage Differentiation Tests of hAMSCs
4.8. Flow Cytometry
4.9. RNA Isolation
4.10. Quantitative Real-Time PCR
4.11. Preparation of S1008/A9-Pretreated hAMSC-Derived Conditioned Medium
4.12. Murine Myocardial I/R Model
4.13. Echocardiography
4.14. Histological Staining and Quantification Analysis
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MI | myocardial infarction |
| DAMPs | Danger Associated Molecular Patterns |
| hMSCs | human mesenchymal stem cells |
| hAMSCs | human amniotic membrane stem cells |
| nCMs | neonatal cardiomyocytes |
| H/R | hypoxia/reoxygenation |
| I/R | ischemia/reoxygenation |
| IHD | ischemic heart disease |
| ECG | Electrocardiography |
| LVH | left ventricular hypertrophy |
| MSC | mesenchymal stem cell |
| LAD | left anterior descending artery |
| LVDd | left ventricular end-diastolic dimension |
| LVDs | left ventricular end-systolic dimension |
| FS | fractional shortening |
| EF | ejection fraction |
| EDvol | end-diastolic volume |
| ESvol | end-systolic volume |
| H&E | hematoxylin and eosin |
References
- Mendis, S.; Puska, P.; Norrving, B. Global Atlas on Cardiovascular Disease Prevention and Control; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Moore, K.L.; Dalley, A.F. Clinically Oriented Anatomy; Wolters Kluwer India Pvt Ltd.: Toronto, ON, Canada, 2018. [Google Scholar]
- Pascual-Gil, S.; Garbayo, E.; Díaz-Herráez, P.; Prosper, F.; Blanco-Prieto, M.J. Heart regeneration after myocardial infarction using synthetic biomaterials. J. Control Release 2015, 203, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, S.D.; Frangogiannis, N.G. The biological basis for cardiac repair after myocardial infarction: From inflammation to fibrosis. Circ. Res. 2016, 119, 91–112. [Google Scholar] [CrossRef]
- Vénéreau, E.; Ceriotti, C.; Bianchi, M.E. DAMPs from cell death to new life. Front. Immunol. 2015, 6, 422. [Google Scholar] [CrossRef] [Green Version]
- Gentek, R.; Hoeffel, G. The innate immune response in myocardial infarction, repair, and regeneration. Adv. Exp. Med. Biol. 2017, 1003, 251–272. [Google Scholar]
- Cashman, T.J.; Gouon-Evans, V.; Costa, K.D. Mesenchymal stem cells for cardiac therapy: Practical challenges and potential mechanisms. Stem Cell Rev. Rep. 2013, 9, 254–265. [Google Scholar] [CrossRef] [Green Version]
- Pistoia, V.; Raffaghello, L. Damage-associated molecular patterns (DAMPs) and mesenchymal stem cells: A matter of attraction and excitement. Eur. J. Immunol. 2011, 41, 1828–1831. [Google Scholar] [CrossRef]
- Golpanian, S.; Wolf, A.; Hatzistergos, K.E.; Hare, J.M. Rebuilding the damaged heart: Mesenchymal stem cells, cell-based therapy, and engineered heart tissue. Physiol. Rev. 2016, 96, 1127–1168. [Google Scholar] [CrossRef]
- Zakrzewski, W.; Dobrzyński, M.; Szymonowicz, M.; Rybak, Z. Stem cells: Past, present, and future. Stem Cell Res. Ther. 2019, 10, 1–22. [Google Scholar] [CrossRef]
- Timmers, L.; Lim, S.K.; Arslan, F.; Armstrong, J.S.; Hoefer, I.E.; Doevendans, P.A.; Piek, J.J.; El Oakley, R.M.; Choo, A.; Lee, C.N. Reduction of myocardial infarct size by human mesenchymal stem cell conditioned medium. Stem Cell Res. 2008, 1, 129–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.-W.; Huang, Q.-M.; Wu, H.-Y.; Zuo, G.-S.-L.; Gu, H.-C.; Deng, K.-Y.; Xin, H.-B. Characteristics and therapeutic potential of human amnion-derived stem cells. Int. J. Mol. Sci. 2021, 22, 970. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-W.; Zhang, H.-Z.; Kim, C.-E.; Kim, J.-M.; Kim, M.H. Amniotic mesenchymal stem cells with robust chemotactic properties are effective in the treatment of a myocardial infarction model. Int. J. Cardiol. 2013, 168, 1062–1069. [Google Scholar] [CrossRef]
- Kim, P.J.; Mahmoudi, M.; Ge, X.; Matsuura, Y.; Toma, I.; Metzler, S.; Kooreman, N.G.; Ramunas, J.; Holbrook, C.; McConnell, M.V. Direct evaluation of myocardial viability and stem cell engraftment demonstrates salvage of the injured myocardium. Circ. Res. 2015, 116, e40–e50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maleki, S.N.; Aboutaleb, N.; Nazarinia, D.; Beik, S.A.; Qolamian, A.; Nobakht, M. Conditioned medium obtained from human amniotic membrane-derived mesenchymal stem cell attenuates heart failure injury in rats. Iran. J. Basic Med. Sci. 2019, 22, 1253. [Google Scholar]
- Basu, A.; Munir, S.; Mulaw, M.A.; Singh, K.; Crisan, D.; Sindrilaru, A.; Treiber, N.; Wlaschek, M.; Huber-Lang, M.; Gebhard, F. A novel S100A8/A9 induced fingerprint of mesenchymal stem cells associated with enhanced wound healing. Sci. Rep. 2018, 8, 1–16. [Google Scholar]
- Viola, M.; de Jager, S.C.; Sluijter, J.P. Targeting Inflammation after Myocardial Infarction: A Therapeutic Opportunity for Extracellular Vesicles? Int. J. Mol. Sci. 2021, 22, 7831. [Google Scholar] [CrossRef] [PubMed]
- Sagaradze, G.; Grigorieva, O.; Nimiritsky, P.; Basalova, N.; Kalinina, N.; Akopyan, Z.; Efimenko, A. Conditioned medium from human mesenchymal stromal cells: Towards the clinical translation. Int. J. Mol. Sci. 2019, 20, 1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, M.; Liang, Y.; Wang, C.; Naruse, K.; Takahashi, K. Treatment of oxidative stress with exosomes in myocardial ischemia. Int. J. Mol. Sci. 2021, 22, 1729. [Google Scholar] [CrossRef] [PubMed]
- Luo, R.; Lu, Y.; Liu, J.; Cheng, J.; Chen, Y. Enhancement of the efficacy of mesenchymal stem cells in the treatment of ischemic diseases. Biomed. Pharmacother. 2019, 109, 2022–2034. [Google Scholar] [CrossRef]
- Ehrchen, J.M.; Sunderkötter, C.; Foell, D.; Vogl, T.; Roth, J. The endogenous toll–like receptor 4 agonist S100A8/S100A9 (calprotectin) as innate amplifier of infection, autoimmunity, and cancer. J. Leukoc. Biol. 2009, 86, 557–566. [Google Scholar] [CrossRef]
- Sasaki, M.; Abe, R.; Fujita, Y.; Ando, S.; Inokuma, D.; Shimizu, H. Mesenchymal stem cells are recruited into wounded skin and contribute to wound repair by transdifferentiation into multiple skin cell type. J. Immunol. 2008, 180, 2581–2587. [Google Scholar] [CrossRef]
- Cal, S.; Quesada, V.; Llamazares, M.; Díaz-Perales, A.; Garabaya, C.; López-Otín, C. Human polyserase-2, a novel enzyme with three tandem serine protease domains in a single polypeptide chain. J. Biol. Chem. 2005, 280, 1953–1961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cominelli, A.; Gaide Chevronnay, H.P.; Lemoine, P.; Courtoy, P.J.; Marbaix, E.; Henriet, P. Matrix metalloproteinase-27 is expressed in CD163+/CD206+ M2 macrophages in the cycling human endometrium and in superficial endometriotic lesions. Mol. Hum. Reprod. 2014, 20, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Buga, A.-M.; Scholz, C.J.; Kumar, S.; Herndon, J.G.; Alexandru, D.; Cojocaru, G.R.; Dandekar, T.; Popa-Wagner, A. Identification of new therapeutic targets by genome-wide analysis of gene expression in the ipsilateral cortex of aged rats after stroke. PLoS ONE 2012, 7, e50985. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Kligman, L.H.; Samulewicz, S.J.; Howe, C.C. Impaired wound healing in mice deficient in a matricellular protein SPARC (osteonectin, BM-40). BMC Cell Biol. 2001, 2, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, M.; Nito, C.; Sowa, K.; Suda, S.; Nishiyama, Y.; Nakamura-Takahashi, A.; Nitahara-Kasahara, Y.; Imagawa, K.; Hirato, T.; Ueda, M. Mesenchymal stem cells overexpressing interleukin-10 promote neuroprotection in experimental acute ischemic stroke. Mol. Ther. Methods Clin. Dev. 2017, 6, 102–111. [Google Scholar] [CrossRef] [Green Version]
- Bolli, R.; Becker, L.; Gross, G.; Mentzer, R., Jr.; Balshaw, D.; Lathrop, D.A. Myocardial protection at a crossroads: The need for translation into clinical therapy. Circ. Res. 2004, 95, 125–134. [Google Scholar] [CrossRef]
- Díaz-Prado, S.; Muiños-López, E.; Hermida-Gómez, T.; Rendal-Vázquez, M.E.; Fuentes-Boquete, I.; de Toro, F.J.; Blanco, F.J. Isolation and characterization of mesenchymal stem cells from human amniotic membrane. Tissue Eng. Part C Methods 2011, 17, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Vidyasekar, P.; Shyamsunder, P.; Santhakumar, R.; Arun, R.; Verma, R.S. A simplified protocol for the isolation and culture of cardiomyocytes and progenitor cells from neonatal mouse ventricles. Eur. J. Cell Biol. 2015, 94, 444–452. [Google Scholar] [CrossRef]
- Portal, L.; Martin, V.; Assaly, R.; d’Anglemont de Tassigny, A.; Michineau, S.; Berdeaux, A.; Ghaleh, B.; Pons, S. A model of hypoxia-reoxygenation on isolated adult mouse cardiomyocytes: Characterization, comparison with ischemia-reperfusion, and application to the cardioprotective effect of regular treadmill exercise. J. Cardiovasc. Pharmacol. Ther. 2013, 18, 367–375. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, T.-J.; Yeh, Y.-T.; Peng, F.-S.; Li, A.-H.; Wu, S.-C. S100A8/A9 Enhances Immunomodulatory and Tissue-Repairing Properties of Human Amniotic Mesenchymal Stem Cells in Myocardial Ischemia-Reperfusion Injury. Int. J. Mol. Sci. 2021, 22, 11175. https://doi.org/10.3390/ijms222011175
Chen T-J, Yeh Y-T, Peng F-S, Li A-H, Wu S-C. S100A8/A9 Enhances Immunomodulatory and Tissue-Repairing Properties of Human Amniotic Mesenchymal Stem Cells in Myocardial Ischemia-Reperfusion Injury. International Journal of Molecular Sciences. 2021; 22(20):11175. https://doi.org/10.3390/ijms222011175
Chicago/Turabian StyleChen, Tzu-Jou, Yen-Ting Yeh, Fu-Shiang Peng, Ai-Hsien Li, and Shinn-Chih Wu. 2021. "S100A8/A9 Enhances Immunomodulatory and Tissue-Repairing Properties of Human Amniotic Mesenchymal Stem Cells in Myocardial Ischemia-Reperfusion Injury" International Journal of Molecular Sciences 22, no. 20: 11175. https://doi.org/10.3390/ijms222011175
APA StyleChen, T.-J., Yeh, Y.-T., Peng, F.-S., Li, A.-H., & Wu, S.-C. (2021). S100A8/A9 Enhances Immunomodulatory and Tissue-Repairing Properties of Human Amniotic Mesenchymal Stem Cells in Myocardial Ischemia-Reperfusion Injury. International Journal of Molecular Sciences, 22(20), 11175. https://doi.org/10.3390/ijms222011175