
Glucocorticoid Receptor Signaling in Diabetes


Abstract
1. Introduction
1.1. Stress, Depression and Type 2 Diabetes
1.2. Hypothalamic-Pituitary-Adrenal Axis Hyperactivation and Hypercortisolism in Type 2 Diabetes Mellitus
2. Glucocorticoid Receptor
2.1. Mechanism of Action
2.2. Glucocorticoid Receptor and Metabolism
3. Glucocorticoid Receptor Signaling in Immune Cells in Type 2 Diabetes Mellitus
4. Glucocorticoid Receptor Polymorphisms in Type 2 Diabetes Mellitus
5. The Effects of Micro-RNAs on Glucocorticoid Receptor in Type 2 Diabetes Mellitus
6. Glucocorticoid Receptor and Stevia Glycosides
7. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- American Diabetes Association. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2021. Diabetes Care 2021, 44, S15–S33. [Google Scholar] [CrossRef]
- Chawla, A.; Chawla, R.; Jaggi, S. Microvasular and macrovascular complications in diabetes mellitus: Distinct or continuum? Indian J. Endocrinol. Metab. 2016, 20, 546–551. [Google Scholar] [CrossRef]
- Tuomi, T.; Santoro, N.; Caprio, S.; Cai, M.; Weng, J.; Groop, L. The many faces of diabetes: A disease with increasing heterogeneity. Lancet 2014, 383, 1084–1094. [Google Scholar] [CrossRef]
- Hu, F.B.; Manson, J.E.; Stampfer, M.J.; Colditz, G.; Liu, S.; Solomon, C.G.; Willett, W.C. Diet, lifestyle, and the risk of type 2 diabetes mellitus in women. N. Engl. J. Med. 2001, 345, 790–797. [Google Scholar] [CrossRef]
- Xue, A.; Wu, Y.; Zhu, Z.; Zhang, F.; Kemper, K.E.; Zheng, Z.; Yengo, L.; Lloyd-Jones, L.R.; Sidorenko, J.; Wu, Y.; et al. Genome-wide association analyses identify 143 risk variants and putative regulatory mechanisms for type 2 diabetes. Nat. Commun. 2018, 9, 2941. [Google Scholar] [CrossRef]
- Kivimaki, M.; Virtanen, M.; Kawachi, I.; Nyberg, S.T.; Alfredsson, L.; Batty, G.D.; Bjorner, J.B.; Borritz, M.; Brunner, E.J.; Burr, H.; et al. Long working hours, socioeconomic status, and the risk of incident type 2 diabetes: A meta-analysis of published and unpublished data from 222 120 individuals. Lancet Diabetes Endocrinol. 2015, 3, 27–34. [Google Scholar] [CrossRef]
- Nyberg, S.T.; Fransson, E.I.; Heikkila, K.; Ahola, K.; Alfredsson, L.; Bjorner, J.B.; Borritz, M.; Burr, H.; Dragano, N.; Goldberg, M.; et al. Job strain as a risk factor for type 2 diabetes: A pooled analysis of 124,808 men and women. Diabetes Care 2014, 37, 2268–2275. [Google Scholar] [CrossRef]
- Huang, H.; Yan, P.; Shan, Z.; Chen, S.; Li, M.; Luo, C.; Gao, H.; Hao, L.; Liu, L. Adverse childhood experiences and risk of type 2 diabetes: A systematic review and meta-analysis. Metabolism 2015, 64, 1408–1418. [Google Scholar] [CrossRef]
- Demakakos, P.; Zaninotto, P.; Nouwen, A. Is the association between depressive symptoms and glucose metabolism bidirectional? Evidence from the English Longitudinal Study of Ageing. Psychosom. Med. 2014, 76, 555–561. [Google Scholar] [CrossRef]
- Knol, M.J.; Twisk, J.W.; Beekman, A.T.; Heine, R.J.; Snoek, F.J.; Pouwer, F. Depression as a risk factor for the onset of type 2 diabetes mellitus. A meta-analysis. Diabetologia 2006, 49, 837–845. [Google Scholar] [CrossRef]
- Mezuk, B.; Eaton, W.W.; Albrecht, S.; Golden, S.H. Depression and type 2 diabetes over the lifespan: A meta-analysis. Diabetes Care 2008, 31, 2383–2390. [Google Scholar] [CrossRef] [PubMed]
- Rotella, F.; Mannucci, E. Depression as a risk factor for diabetes: A meta-analysis of longitudinal studies. J. Clin. Psychiatry 2013, 74, 31–37. [Google Scholar] [CrossRef]
- Bergmans, R.S.; Rapp, A.; Kelly, K.M.; Weiss, D.; Mezuk, B. Understanding the relationship between type 2 diabetes and depression: Lessons from genetically informative study designs. Diabet. Med. 2021, 38, e14399. [Google Scholar] [CrossRef]
- Inoue, K.; Beekley, J.; Goto, A.; Jeon, C.Y.; Ritz, B.R. Depression and cardiovascular disease events among patients with type 2 diabetes: A systematic review and meta-analysis with bias analysis. J. Diabetes Complic. 2020, 34, 107710. [Google Scholar] [CrossRef]
- Loizzo, S.; Campana, G.; Vella, S.; Fortuna, A.; Galietta, G.; Guarino, I.; Costa, L.; Capasso, A.; Renzi, P.; Frajese, G.V.; et al. Post-natal stress-induced endocrine and metabolic alterations in mice at adulthood involve different pro-opiomelanocortin-derived peptides. Peptides 2010, 31, 2123–2129. [Google Scholar] [CrossRef]
- Maniam, J.; Antoniadis, C.P.; Morris, M.J. The effect of early-life stress and chronic high-sucrose diet on metabolic outcomes in female rats. Stress 2015, 18, 524–537. [Google Scholar] [CrossRef]
- Hackett, R.A.; Steptoe, A.; Kumari, M. Association of diurnal patterns in salivary cortisol with type 2 diabetes in the Whitehall II study. J. Clin. Endocrinol. Metab. 2014, 99, 4625–4631. [Google Scholar] [CrossRef]
- Steptoe, A.; Hackett, R.A.; Lazzarino, A.I.; Bostock, S.; La Marca, R.; Carvalho, L.A.; Hamer, M. Disruption of multisystem responses to stress in type 2 diabetes: Investigating the dynamics of allostatic load. Proc. Natl. Acad. Sci. USA 2014, 111, 15693–15698. [Google Scholar] [CrossRef]
- Hackett, R.A.; Steptoe, A. Type 2 diabetes mellitus and psychological stress—A modifiable risk factor. Nat. Rev. Endocrinol. 2017, 13, 547–560. [Google Scholar] [CrossRef]
- Joseph, J.J.; Golden, S.H. Cortisol dysregulation: The bidirectional link between stress, depression, and type 2 diabetes mellitus. Ann. N. Y. Acad. Sci. 2017, 1391, 20–34. [Google Scholar] [CrossRef]
- Tsigos, C.; Chrousos, G.P. Hypothalamic-pituitary-adrenal axis, neuroendocrine factors and stress. J. Psychosom. Res. 2002, 53, 865–871. [Google Scholar] [CrossRef]
- Ramamoorthy, S.; Cidlowski, J.A. Corticosteroids: Mechanisms of Action in Health and Disease. Rheum. Dis. Clin. N. Am. 2016, 42, 15–31. [Google Scholar] [CrossRef]
- Nicolaides, N.C.; Charmandari, E.; Chrousos, G.P.; Kino, T. Recent advances in the molecular mechanisms determining tissue sensitivity to glucocorticoids: Novel mutations, circadian rhythm and ligand-induced repression of the human glucocorticoid receptor. BMC Endocr. Disord. 2014, 14, 71. [Google Scholar] [CrossRef] [PubMed]
- Breuner, C.W.; Orchinik, M. Plasma binding proteins as mediators of corticosteroid action in vertebrates. J. Endocrinol. 2002, 175, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Seckl, J.R. 11beta-hydroxysteroid dehydrogenases: Changing glucocorticoid action. Curr. Opin. Pharmacol. 2004, 4, 597–602. [Google Scholar] [CrossRef] [PubMed]
- Schibler, U.; Sassone-Corsi, P. A web of circadian pacemakers. Cell 2002, 111, 919–922. [Google Scholar] [CrossRef]
- Takahashi, J.S.; Hong, H.K.; Ko, C.H.; McDearmon, E.L. The genetics of mammalian circadian order and disorder: Implications for physiology and disease. Nat. Rev. Genet. 2008, 9, 764–775. [Google Scholar] [CrossRef]
- Kadmiel, M.; Cidlowski, J.A. Glucocorticoid receptor signaling in health and disease. Trends Pharmacol. Sci. 2013, 34, 518–530. [Google Scholar] [CrossRef]
- Chrousos, G.P. Stress and disorders of the stress system. Nat. Rev. Endocrinol. 2009, 5, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Kyrou, I.; Chrousos, G.P.; Tsigos, C. Stress, visceral obesity, and metabolic complications. Ann. N. Y. Acad. Sci. 2006, 1083, 77–110. [Google Scholar] [CrossRef]
- Chiodini, I.; Adda, G.; Scillitani, A.; Coletti, F.; Morelli, V.; Di Lembo, S.; Epaminonda, P.; Masserini, B.; Beck-Peccoz, P.; Orsi, E.; et al. Cortisol secretion in patients with type 2 diabetes: Relationship with chronic complications. Diabetes Care 2007, 30, 83–88. [Google Scholar] [CrossRef]
- Prpic-Krizevac, I.; Canecki-Varzic, S.; Bilic-Curcic, I. Hyperactivity of the hypothalamic-pituitary-adrenal axis in patients with type 2 diabetes and relations with insulin resistance and chronic complications. Wien. Klin. Wochenschr. 2012, 124, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Bruehl, H.; Rueger, M.; Dziobek, I.; Sweat, V.; Tirsi, A.; Javier, E.; Arentoft, A.; Wolf, O.T.; Convit, A. Hypothalamic-pituitary-adrenal axis dysregulation and memory impairments in type 2 diabetes. J. Clin. Endocrinol. Metab. 2007, 92, 2439–2445. [Google Scholar] [CrossRef]
- Steffensen, C.; Thomsen, H.H.; Dekkers, O.M.; Christiansen, J.S.; Rungby, J.; Jorgensen, J.O. Low positive predictive value of midnight salivary cortisol measurement to detect hypercortisolism in type 2 diabetes. Clin. Endocrinol. 2016, 85, 202–206. [Google Scholar] [CrossRef]
- Tsigos, C.; Young, R.J.; White, A. Diabetic neuropathy is associated with increased activity of the hypothalamic-pituitary-adrenal axis. J. Clin. Endocrinol. Metab. 1993, 76, 554–558. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.S.; Roy, A.; Brown, S. Increased urinary-free cortisol outputs in diabetic patients. J. Diabetes Complicat. 1998, 12, 24–27. [Google Scholar] [CrossRef]
- Champaneri, S.; Xu, X.; Carnethon, M.R.; Bertoni, A.G.; Seeman, T.; Diez Roux, A.; Golden, S.H. Diurnal salivary cortisol and urinary catecholamines are associated with diabetes mellitus: The Multi-Ethnic Study of Atherosclerosis. Metabolism 2012, 61, 986–995. [Google Scholar] [CrossRef] [PubMed]
- Oltmanns, K.M.; Dodt, B.; Schultes, B.; Raspe, H.H.; Schweiger, U.; Born, J.; Fehm, H.L.; Peters, A. Cortisol correlates with metabolic disturbances in a population study of type 2 diabetic patients. Eur. J. Endocrinol. 2006, 154, 325–331. [Google Scholar] [CrossRef]
- Direk, N.; Dekker, M.J.; Luik, A.I.; Kirschbaum, C.; de Rijke, Y.B.; Hofman, A.; Hoogendijk, W.J.; Tiemeier, H. The Very Low-Dose Dexamethasone Suppression Test in the General Population: A Cross-Sectional Study. PLoS ONE 2016, 11, e0164348. [Google Scholar] [CrossRef] [PubMed]
- Ryan, R.; Booth, S.; Spathis, A.; Mollart, S.; Clow, A. Use of Salivary Diurnal Cortisol as an Outcome Measure in Randomised Controlled Trials: A Systematic Review. Ann. Behav. Med. 2016, 50, 210–236. [Google Scholar] [CrossRef] [PubMed]
- Kudielka, B.M.; Wust, S. Human models in acute and chronic stress: Assessing determinants of individual hypothalamus-pituitary-adrenal axis activity and reactivity. Stress 2010, 13, 1–14. [Google Scholar] [CrossRef]
- Pariante, C.M.; Lightman, S.L. The HPA axis in major depression: Classical theories and new developments. Trends Neurosci. 2008, 31, 464–468. [Google Scholar] [CrossRef]
- Khoury, J.E.; Gonzalez, A.; Levitan, R.D.; Pruessner, J.C.; Chopra, K.; Basile, V.S.; Masellis, M.; Goodwill, A.; Atkinson, L. Summary cortisol reactivity indicators: Interrelations and meaning. Neurobiol. Stress 2015, 2, 34–43. [Google Scholar] [CrossRef]
- Lederbogen, F.; Hummel, J.; Fademrecht, C.; Krumm, B.; Kuhner, C.; Deuschle, M.; Ladwig, K.H.; Meisinger, C.; Wichmann, H.E.; Lutz, H.; et al. Flattened circadian cortisol rhythm in type 2 diabetes. Exp. Clin. Endocrinol. Diabetes 2011, 119, 573–575. [Google Scholar] [CrossRef]
- Bruehl, H.; Wolf, O.T.; Convit, A. A blunted cortisol awakening response and hippocampal atrophy in type 2 diabetes mellitus. Psychoneuroendocrinology 2009, 34, 815–821. [Google Scholar] [CrossRef]
- Johar, H.; Emeny, R.T.; Bidlingmaier, M.; Kruse, J.; Ladwig, K.H. Sex-related differences in the association of salivary cortisol levels and type 2 diabetes. Findings from the cross-sectional population based KORA-age study. Psychoneuroendocrinology 2016, 69, 133–141. [Google Scholar] [CrossRef]
- Spanakis, E.K.; Wang, X.; Sanchez, B.N.; Diez Roux, A.V.; Needham, B.L.; Wand, G.S.; Seeman, T.; Golden, S.H. Lack of significant association between type 2 diabetes mellitus with longitudinal change in diurnal salivary cortisol: The multiethnic study of atherosclerosis. Endocrine 2016, 53, 227–239. [Google Scholar] [CrossRef]
- Bellastella, G.; Maiorino, M.I.; De Bellis, A.; Vietri, M.T.; Mosca, C.; Scappaticcio, L.; Pasquali, D.; Esposito, K.; Giugliano, D. Serum but not salivary cortisol levels are influenced by daily glycemic oscillations in type 2 diabetes. Endocrine 2016, 53, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Nicolaides, N.C.; Galata, Z.; Kino, T.; Chrousos, G.P.; Charmandari, E. The human glucocorticoid receptor: Molecular basis of biologic function. Steroids 2010, 75, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Oakley, R.H.; Cidlowski, J.A. The biology of the glucocorticoid receptor: New signaling mechanisms in health and disease. J. Allergy Clin. Immunol. 2013, 132, 1033–1044. [Google Scholar] [CrossRef] [PubMed]
- Vandevyver, S.; Dejager, L.; Libert, C. Comprehensive overview of the structure and regulation of the glucocorticoid receptor. Endocr. Rev. 2014, 35, 671–693. [Google Scholar] [CrossRef]
- Lu, N.Z.; Cidlowski, J.A. Glucocorticoid receptor isoforms generate transcription specificity. Trends Cell Biol. 2006, 16, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Bamberger, C.M.; Bamberger, A.M.; de Castro, M.; Chrousos, G.P. Glucocorticoid receptor beta, a potential endogenous inhibitor of glucocorticoid action in humans. J. Clin. Investig. 1995, 95, 2435–2441. [Google Scholar] [CrossRef]
- Rao, N.A.; McCalman, M.T.; Moulos, P.; Francoijs, K.J.; Chatziioannou, A.; Kolisis, F.N.; Alexis, M.N.; Mitsiou, D.J.; Stunnenberg, H.G. Coactivation of GR and NFKB alters the repertoire of their binding sites and target genes. Genome Res. 2011, 21, 1404–1416. [Google Scholar] [CrossRef]
- Biddie, S.C.; John, S.; Sabo, P.J.; Thurman, R.E.; Johnson, T.A.; Schiltz, R.L.; Miranda, T.B.; Sung, M.H.; Trump, S.; Lightman, S.L.; et al. Transcription factor AP1 potentiates chromatin accessibility and glucocorticoid receptor binding. Mol. Cell 2011, 43, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Stocklin, E.; Wissler, M.; Gouilleux, F.; Groner, B. Functional interactions between Stat5 and the glucocorticoid receptor. Nature 1996, 383, 726–728. [Google Scholar] [CrossRef]
- Vandevyver, S.; Dejager, L.; Tuckermann, J.; Libert, C. New insights into the anti-inflammatory mechanisms of glucocorticoids: An emerging role for glucocorticoid-receptor-mediated transactivation. Endocrinology 2013, 154, 993–1007. [Google Scholar] [CrossRef] [PubMed]
- Galigniana, M.D.; Piwien-Pilipuk, G.; Assreuy, J. Inhibition of glucocorticoid receptor binding by nitric oxide. Mol. Pharmacol. 1999, 55, 317–323. [Google Scholar] [CrossRef]
- Le Drean, Y.; Mincheneau, N.; Le Goff, P.; Michel, D. Potentiation of glucocorticoid receptor transcriptional activity by sumoylation. Endocrinology 2002, 143, 3482–3489. [Google Scholar] [CrossRef]
- Nader, N.; Chrousos, G.P.; Kino, T. Circadian rhythm transcription factor CLOCK regulates the transcriptional activity of the glucocorticoid receptor by acetylating its hinge region lysine cluster: Potential physiological implications. FASEB J. 2009, 23, 1572–1583. [Google Scholar] [CrossRef]
- Okamoto, K.; Tanaka, H.; Ogawa, H.; Makino, Y.; Eguchi, H.; Hayashi, S.; Yoshikawa, N.; Poellinger, L.; Umesono, K.; Makino, I. Redox-dependent regulation of nuclear import of the glucocorticoid receptor. J. Biol. Chem. 1999, 274, 10363–10371. [Google Scholar] [CrossRef]
- Wallace, A.D.; Cidlowski, J.A. Proteasome-mediated glucocorticoid receptor degradation restricts transcriptional signaling by glucocorticoids. J. Biol. Chem. 2001, 276, 42714–42721. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, W.; Kono, E.; Dang, T.; Garabedian, M.J. Modulation of glucocorticoid receptor phosphorylation and transcriptional activity by a C-terminal-associated protein phosphatase. Mol. Endocrinol. 2007, 21, 625–634. [Google Scholar] [CrossRef]
- Chrousos, G.P.; Kino, T. Intracellular glucocorticoid signaling: A formerly simple system turns stochastic. Sci. Signal. 2005, 2005, pe48. [Google Scholar] [CrossRef]
- Cain, D.W.; Cidlowski, J.A. Specificity and sensitivity of glucocorticoid signaling in health and disease. Best Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 545–556. [Google Scholar] [CrossRef]
- Ito, K.; Yamamura, S.; Essilfie-Quaye, S.; Cosio, B.; Ito, M.; Barnes, P.J.; Adcock, I.M. Histone deacetylase 2-mediated deacetylation of the glucocorticoid receptor enables NF-kappaB suppression. J. Exp. Med. 2006, 203, 7–13. [Google Scholar] [CrossRef]
- Irusen, E.; Matthews, J.G.; Takahashi, A.; Barnes, P.J.; Chung, K.F.; Adcock, I.M. p38 Mitogen-activated protein kinase-induced glucocorticoid receptor phosphorylation reduces its activity: Role in steroid-insensitive asthma. J. Allergy Clin. Immunol. 2002, 109, 649–657. [Google Scholar] [CrossRef]
- Szatmary, Z.; Garabedian, M.J.; Vilcek, J. Inhibition of glucocorticoid receptor-mediated transcriptional activation by p38 mitogen-activated protein (MAP) kinase. J. Biol. Chem. 2004, 279, 43708–43715. [Google Scholar] [CrossRef]
- Shahidi, H.; Vottero, A.; Stratakis, C.A.; Taymans, S.E.; Karl, M.; Longui, C.A.; Chrousos, G.P.; Daughaday, W.H.; Gregory, S.A.; Plate, J.M. Imbalanced expression of the glucocorticoid receptor isoforms in cultured lymphocytes from a patient with systemic glucocorticoid resistance and chronic lymphocytic leukemia. Biochem. Biophys. Res. Commun. 1999, 254, 559–565. [Google Scholar] [CrossRef]
- Koga, Y.; Matsuzaki, A.; Suminoe, A.; Hattori, H.; Kanemitsu, S.; Hara, T. Differential mRNA expression of glucocorticoid receptor alpha and beta is associated with glucocorticoid sensitivity of acute lymphoblastic leukemia in children. Pediatr. Blood Cancer 2005, 45, 121–127. [Google Scholar] [CrossRef]
- Tienrungroj, W.; Meshinchi, S.; Sanchez, E.R.; Pratt, S.E.; Grippo, J.F.; Holmgren, A.; Pratt, W.B. The role of sulfhydryl groups in permitting transformation and DNA binding of the glucocorticoid receptor. J. Biol. Chem. 1987, 262, 6992–7000. [Google Scholar] [CrossRef]
- Silva, C.M.; Cidlowski, J.A. Direct evidence for intra- and intermolecular disulfide bond formation in the human glucocorticoid receptor. Inhibition of DNA binding and identification of a new receptor-associated protein. J. Biol. Chem. 1989, 264, 6638–6647. [Google Scholar] [CrossRef]
- Hutchison, K.A.; Matic, G.; Czar, M.J.; Pratt, W.B. DNA-binding and non-DNA-binding forms of the transformed glucocorticoid receptor. J. Steroid Biochem. Mol. Biol. 1992, 41, 715–718. [Google Scholar] [CrossRef][Green Version]
- Hutchison, K.A.; Matic, G.; Meshinchi, S.; Bresnick, E.H.; Pratt, W.B. Redox manipulation of DNA binding activity and BuGR epitope reactivity of the glucocorticoid receptor. J. Biol. Chem. 1991, 266, 10505–10509. [Google Scholar] [CrossRef]
- Ashida, H.; Ohue, K.; Kanazawa, K.; Danno, G. Effects of dietary lipid peroxidation products on hormonal responses in primary cultured hepatocytes of rats. Biosci. Biotechnol. Biochem. 1997, 61, 2089–2094. [Google Scholar] [CrossRef]
- Esposito, F.; Cuccovillo, F.; Morra, F.; Russo, T.; Cimino, F. DNA binding activity of the glucocorticoid receptor is sensitive to redox changes in intact cells. Biochim. Biophys. Acta 1995, 1260, 308–314. [Google Scholar] [CrossRef]
- Makino, Y.; Okamoto, K.; Yoshikawa, N.; Aoshima, M.; Hirota, K.; Yodoi, J.; Umesono, K.; Makino, I.; Tanaka, H. Thioredoxin: A redox-regulating cellular cofactor for glucocorticoid hormone action. Cross talk between endocrine control of stress response and cellular antioxidant defense system. J. Clin. Investig. 1996, 98, 2469–2477. [Google Scholar] [CrossRef]
- Okamoto, K.; Tanaka, H.; Makino, Y.; Makino, I. Restoration of the glucocorticoid receptor function by the phosphodiester compound of vitamins C and E, EPC-K1 (L-ascorbic acid 2-[3,4-dihydro-2,5,7,8-tetramethyl-2-(4,8,12-trimethyltridecyl)-2H-1-benzopyran-6-yl hydrogen phosphate] potassium salt), via a redox-dependent mechanism. Biochem. Pharmacol. 1998, 56, 79–86. [Google Scholar] [CrossRef]
- Makino, Y.; Yoshikawa, N.; Okamoto, K.; Hirota, K.; Yodoi, J.; Makino, I.; Tanaka, H. Direct association with thioredoxin allows redox regulation of glucocorticoid receptor function. J. Biol. Chem. 1999, 274, 3182–3188. [Google Scholar] [CrossRef]
- Hafezi-Moghadam, A.; Simoncini, T.; Yang, Z.; Limbourg, F.P.; Plumier, J.C.; Rebsamen, M.C.; Hsieh, C.M.; Chui, D.S.; Thomas, K.L.; Prorock, A.J.; et al. Acute cardiovascular protective effects of corticosteroids are mediated by non-transcriptional activation of endothelial nitric oxide synthase. Nat. Med. 2002, 8, 473–479. [Google Scholar] [CrossRef]
- Dimmeler, S.; Fleming, I.; Fisslthaler, B.; Hermann, C.; Busse, R.; Zeiher, A.M. Activation of nitric oxide synthase in endothelial cells by Akt-dependent phosphorylation. Nature 1999, 399, 601–605. [Google Scholar] [CrossRef]
- Li, X.; Qiu, J.; Wang, J.; Zhong, Y.; Zhu, J.; Chen, Y. Corticosterone-induced rapid phosphorylation of p38 and JNK mitogen-activated protein kinases in PC12 cells. FEBS Lett. 2001, 492, 210–214. [Google Scholar] [CrossRef]
- Qiu, J.; Wang, P.; Jing, Q.; Zhang, W.; Li, X.; Zhong, Y.; Sun, G.; Pei, G.; Chen, Y. Rapid activation of ERK1/2 mitogen-activated protein kinase by corticosterone in PC12 cells. Biochem. Biophys. Res. Commun. 2001, 287, 1017–1024. [Google Scholar] [CrossRef]
- Panettieri, R.A.; Schaafsma, D.; Amrani, Y.; Koziol-White, C.; Ostrom, R.; Tliba, O. Non-genomic Effects of Glucocorticoids: An Updated View. Trends Pharmacol. Sci. 2019, 40, 38–49. [Google Scholar] [CrossRef]
- Zhang, Y.; Sheng, H.; Qi, J.; Ma, B.; Sun, J.; Li, S.; Ni, X. Glucocorticoid acts on a putative G protein-coupled receptor to rapidly regulate the activity of NMDA receptors in hippocampal neurons. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E747–E758. [Google Scholar] [CrossRef]
- Kokkinopoulou, I.; Moutsatsou, P. Mitochondrial Glucocorticoid Receptors and Their Actions. Int. J. Mol. Sci. 2021, 22, 6054. [Google Scholar] [CrossRef]
- Demonacos, C.; Djordjevic-Markovic, R.; Tsawdaroglou, N.; Sekeris, C.E. The mitochondrion as a primary site of action of glucocorticoids: The interaction of the glucocorticoid receptor with mitochondrial DNA sequences showing partial similarity to the nuclear glucocorticoid responsive elements. J. Steroid Biochem. Mol. Biol. 1995, 55, 43–55. [Google Scholar] [CrossRef]
- Leistner, C.; Menke, A. How to measure glucocorticoid receptor’s sensitivity in patients with stress-related psychiatric disorders. Psychoneuroendocrinology 2018, 91, 235–260. [Google Scholar] [CrossRef] [PubMed]
- Vegiopoulos, A.; Herzig, S. Glucocorticoids, metabolism and metabolic diseases. Mol. Cell. Endocrinol. 2007, 275, 43–61. [Google Scholar] [CrossRef]
- Biering, H.; Knappe, G.; Gerl, H.; Lochs, H. Prevalence of diabetes in acromegaly and Cushing syndrome. Acta Med. Austriaca 2000, 27, 27–31. [Google Scholar] [CrossRef]
- Catargi, B.; Rigalleau, V.; Poussin, A.; Ronci-Chaix, N.; Bex, V.; Vergnot, V.; Gin, H.; Roger, P.; Tabarin, A. Occult Cushing’s syndrome in type-2 diabetes. J. Clin. Endocrinol. Metab. 2003, 88, 5808–5813. [Google Scholar] [CrossRef]
- Henriksen, J.E.; Alford, F.; Ward, G.M.; Beck-Nielsen, H. Risk and mechanism of dexamethasone-induced deterioration of glucose tolerance in non-diabetic first-degree relatives of NIDDM patients. Diabetologia 1997, 40, 1439–1448. [Google Scholar] [CrossRef]
- Rahimi, L.; Rajpal, A.; Ismail-Beigi, F. Glucocorticoid-Induced Fatty Liver Disease. Diabetes Metab. Syndr. Obes. 2020, 13, 1133–1145. [Google Scholar] [CrossRef] [PubMed]
- Sato, A.; Funder, J.W.; Okubo, M.; Kubota, E.; Saruta, T. Glucocorticoid-induced hypertension in the elderly. Relation to serum calcium and family history of essential hypertension. Am. J. Hypertens. 1995, 8, 823–828. [Google Scholar] [CrossRef]
- Smith, O.L.; Wong, C.Y.; Gelfand, R.A. Influence of glucocorticoids on skeletal muscle proteolysis in normal and diabetic-adrenalectomized eviscerated rats. Metabolism 1990, 39, 641–646. [Google Scholar] [CrossRef]
- Kuo, T.; McQueen, A.; Chen, T.C.; Wang, J.C. Regulation of Glucose Homeostasis by Glucocorticoids. Adv. Exp. Med. Biol. 2015, 872, 99–126. [Google Scholar] [CrossRef] [PubMed]
- Berdanier, C.D. Role of glucocorticoids in the regulation of lipogenesis. FASEB J. 1989, 3, 2179–2183. [Google Scholar] [CrossRef]
- Coderre, L.; Srivastava, A.K.; Chiasson, J.L. Effect of hypercorticism on regulation of skeletal muscle glycogen metabolism by insulin. Am. J. Physiol. 1992, 262, E427–E433. [Google Scholar] [CrossRef]
- Dimitriadis, G.; Leighton, B.; Parry-Billings, M.; Sasson, S.; Young, M.; Krause, U.; Bevan, S.; Piva, T.; Wegener, G.; Newsholme, E.A. Effects of glucocorticoid excess on the sensitivity of glucose transport and metabolism to insulin in rat skeletal muscle. Biochem. J. 1997, 321 Pt 3, 707–712. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, D.P.; Forbes, S.; Walker, B.R. Glucocorticoids and fatty acid metabolism in humans: Fuelling fat redistribution in the metabolic syndrome. J. Endocrinol. 2008, 197, 189–204. [Google Scholar] [CrossRef]
- Lee, M.J.; Pramyothin, P.; Karastergiou, K.; Fried, S.K. Deconstructing the roles of glucocorticoids in adipose tissue biology and the development of central obesity. Biochim. Biophys. Acta 2014, 1842, 473–481. [Google Scholar] [CrossRef]
- Longano, C.A.; Fletcher, H.P. Insulin release after acute hydrocortisone treatment in mice. Metabolism 1983, 32, 603–608. [Google Scholar] [CrossRef]
- Wise, J.K.; Hendler, R.; Felig, P. Influence of glucocorticoids on glucagon secretion and plasma amino acid concentrations in man. J. Clin. Investig. 1973, 52, 2774–2782. [Google Scholar] [CrossRef] [PubMed]
- Lambillotte, C.; Gilon, P.; Henquin, J.C. Direct glucocorticoid inhibition of insulin secretion. An in vitro study of dexamethasone effects in mouse islets. J. Clin. Investig. 1997, 99, 414–423. [Google Scholar] [CrossRef]
- Beaupere, C.; Liboz, A.; Feve, B.; Blondeau, B.; Guillemain, G. Molecular Mechanisms of Glucocorticoid-Induced Insulin Resistance. Int. J. Mol. Sci. 2021, 22, 623. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.Z.; Zhu, Y.M.; Zou, G.H.; Sun, Y.X.; Xiu, X.L.; Huang, X.; Zhang, Q.H. Relationship between Glucocorticoids and Insulin Resistance in Healthy Individuals. Med. Sci. Monit. 2016, 22, 1887–1894. [Google Scholar] [CrossRef] [PubMed]
- Hiraiwa, H.; Chou, J.Y. Glucocorticoids activate transcription of the gene for the glucose-6-phosphate transporter, deficient in glycogen storage disease type 1b. DNA Cell Biol. 2001, 20, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Imai, E.; Stromstedt, P.E.; Quinn, P.G.; Carlstedt-Duke, J.; Gustafsson, J.A.; Granner, D.K. Characterization of a complex glucocorticoid response unit in the phosphoenolpyruvate carboxykinase gene. Mol. Cell. Biol. 1990, 10, 4712–4719. [Google Scholar] [CrossRef]
- Jantzen, H.M.; Strahle, U.; Gloss, B.; Stewart, F.; Schmid, W.; Boshart, M.; Miksicek, R.; Schutz, G. Cooperativity of glucocorticoid response elements located far upstream of the tyrosine aminotransferase gene. Cell 1987, 49, 29–38. [Google Scholar] [CrossRef]
- Pierreux, C.E.; Urso, B.; De Meyts, P.; Rousseau, G.G.; Lemaigre, F.P. Inhibition by insulin of glucocorticoid-induced gene transcription: Involvement of the ligand-binding domain of the glucocorticoid receptor and independence from the phosphatidylinositol 3-kinase and mitogen-activated protein kinase pathways. Mol. Endocrinol. 1998, 12, 1343–1354. [Google Scholar] [CrossRef][Green Version]
- Vander Kooi, B.T.; Onuma, H.; Oeser, J.K.; Svitek, C.A.; Allen, S.R.; Vander Kooi, C.W.; Chazin, W.J.; O’Brien, R.M. The glucose-6-phosphatase catalytic subunit gene promoter contains both positive and negative glucocorticoid response elements. Mol. Endocrinol. 2005, 19, 3001–3022. [Google Scholar] [CrossRef] [PubMed]
- Ngo, S.; Barry, J.B.; Nisbet, J.C.; Prins, J.B.; Whitehead, J.P. Reduced phosphorylation of AS160 contributes to glucocorticoid-mediated inhibition of glucose uptake in human and murine adipocytes. Mol. Cell. Endocrinol. 2009, 302, 33–40. [Google Scholar] [CrossRef]
- Connaughton, S.; Chowdhury, F.; Attia, R.R.; Song, S.; Zhang, Y.; Elam, M.B.; Cook, G.A.; Park, E.A. Regulation of pyruvate dehydrogenase kinase isoform 4 (PDK4) gene expression by glucocorticoids and insulin. Mol. Cell. Endocrinol. 2010, 315, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Kuo, T.; Lew, M.J.; Mayba, O.; Harris, C.A.; Speed, T.P.; Wang, J.C. Genome-wide analysis of glucocorticoid receptor-binding sites in myotubes identifies gene networks modulating insulin signaling. Proc. Natl. Acad. Sci. USA 2012, 109, 11160–11165. [Google Scholar] [CrossRef]
- Beaudry, J.L.; Dunford, E.C.; Teich, T.; Zaharieva, D.; Hunt, H.; Belanoff, J.K.; Riddell, M.C. Effects of selective and non-selective glucocorticoid receptor II antagonists on rapid-onset diabetes in young rats. PLoS ONE 2014, 9, e91248. [Google Scholar] [CrossRef] [PubMed]
- Watts, L.M.; Manchem, V.P.; Leedom, T.A.; Rivard, A.L.; McKay, R.A.; Bao, D.; Neroladakis, T.; Monia, B.P.; Bodenmiller, D.M.; Cao, J.X.; et al. Reduction of hepatic and adipose tissue glucocorticoid receptor expression with antisense oligonucleotides improves hyperglycemia and hyperlipidemia in diabetic rodents without causing systemic glucocorticoid antagonism. Diabetes 2005, 54, 1846–1853. [Google Scholar] [CrossRef]
- Liang, Y.; Osborne, M.C.; Monia, B.P.; Bhanot, S.; Watts, L.M.; She, P.; DeCarlo, S.O.; Chen, X.; Demarest, K. Antisense oligonucleotides targeted against glucocorticoid receptor reduce hepatic glucose production and ameliorate hyperglycemia in diabetic mice. Metabolism 2005, 54, 848–855. [Google Scholar] [CrossRef]
- Jacobson, P.B.; von Geldern, T.W.; Ohman, L.; Osterland, M.; Wang, J.; Zinker, B.; Wilcox, D.; Nguyen, P.T.; Mika, A.; Fung, S.; et al. Hepatic glucocorticoid receptor antagonism is sufficient to reduce elevated hepatic glucose output and improve glucose control in animal models of type 2 diabetes. J. Pharmacol. Exp. Ther. 2005, 314, 191–200. [Google Scholar] [CrossRef]
- Liu, Y.; Nakagawa, Y.; Wang, Y.; Sakurai, R.; Tripathi, P.V.; Lutfy, K.; Friedman, T.C. Increased glucocorticoid receptor and 11{beta}-hydroxysteroid dehydrogenase type 1 expression in hepatocytes may contribute to the phenotype of type 2 diabetes in db/db mice. Diabetes 2005, 54, 32–40. [Google Scholar] [CrossRef]
- Opherk, C.; Tronche, F.; Kellendonk, C.; Kohlmuller, D.; Schulze, A.; Schmid, W.; Schutz, G. Inactivation of the glucocorticoid receptor in hepatocytes leads to fasting hypoglycemia and ameliorates hyperglycemia in streptozotocin-induced diabetes mellitus. Mol. Endocrinol. 2004, 18, 1346–1353. [Google Scholar] [CrossRef]
- Xu, X.; Chen, Y.; Zhu, D.; Zhao, T.; Xu, R.; Wang, J.; Hu, L.; Shen, X. FX5 as a non-steroidal GR antagonist improved glucose homeostasis in type 2 diabetic mice via GR/HNF4alpha/miR-122-5p pathway. Aging 2020, 13, 2436–2458. [Google Scholar] [CrossRef]
- Chen, G.; Wang, R.; Chen, H.; Wu, L.; Ge, R.S.; Wang, Y. Gossypol ameliorates liver fibrosis in diabetic rats induced by high-fat diet and streptozocin. Life Sci. 2016, 149, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Aylward, A.; Okino, M.L.; Benaglio, P.; Chiou, J.; Beebe, E.; Padilla, J.A.; Diep, S.; Gaulton, K.J. Glucocorticoid signaling in pancreatic islets modulates gene regulatory programs and genetic risk of type 2 diabetes. PLoS Genet. 2021, 17, e1009531. [Google Scholar] [CrossRef] [PubMed]
- Lowy, M.T. Reserpine-induced decrease in type I and II corticosteroid receptors in neuronal and lymphoid tissues of adrenalectomized rats. Neuroendocrinology 1990, 51, 190–196. [Google Scholar] [CrossRef]
- Spencer, R.L.; Miller, A.H.; Stein, M.; McEwen, B.S. Corticosterone regulation of type I and type II adrenal steroid receptors in brain, pituitary, and immune tissue. Brain Res. 1991, 549, 236–246. [Google Scholar] [CrossRef]
- Panagiotou, C.; Lambadiari, V.; Maratou, E.; Geromeriati, C.; Artemiadis, A.; Dimitriadis, G.; Moutsatsou, P. Insufficient glucocorticoid receptor signaling and flattened salivary cortisol profile are associated with metabolic and inflammatory indices in type 2 diabetes. J. Endocrinol. Investig. 2021, 44, 37–48. [Google Scholar] [CrossRef]
- Carvalho, L.A.; Urbanova, L.; Hamer, M.; Hackett, R.A.; Lazzarino, A.I.; Steptoe, A. Blunted glucocorticoid and mineralocorticoid sensitivity to stress in people with diabetes. Psychoneuroendocrinology 2015, 51, 209–218. [Google Scholar] [CrossRef]
- Ross, K.M.; Murphy, M.L.M.; Adam, E.K.; Chen, E.; Miller, G.E. How stable are diurnal cortisol activity indices in healthy individuals? Evidence from three multi-wave studies. Psychoneuroendocrinology 2014, 39, 184–193. [Google Scholar] [CrossRef]
- Smit, P.; Russcher, H.; de Jong, F.H.; Brinkmann, A.O.; Lamberts, S.W.; Koper, J.W. Differential regulation of synthetic glucocorticoids on gene expression levels of glucocorticoid-induced leucine zipper and interleukin-2. J. Clin. Endocrinol. Metab. 2005, 90, 2994–3000. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Campbell, J.E.; Kiraly, M.A.; Atkinson, D.J.; D’Souza, A.M.; Vranic, M.; Riddell, M.C. Regular exercise prevents the development of hyperglucocorticoidemia via adaptations in the brain and adrenal glands in male Zucker diabetic fatty rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R168–R176. [Google Scholar] [CrossRef]
- Chan, O.; Chan, S.; Inouye, K.; Shum, K.; Matthews, S.G.; Vranic, M. Diabetes impairs hypothalamo-pituitary-adrenal (HPA) responses to hypoglycemia, and insulin treatment normalizes HPA but not epinephrine responses. Diabetes 2002, 51, 1681–1689. [Google Scholar] [CrossRef]
- Chan, O.; Inouye, K.; Akirav, E.M.; Park, E.; Riddell, M.C.; Matthews, S.G.; Vranic, M. Hyperglycemia does not increase basal hypothalamo-pituitary-adrenal activity in diabetes but it does impair the HPA response to insulin-induced hypoglycemia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R235–R246. [Google Scholar] [CrossRef]
- Slag, M.F.; Ahmad, M.; Gannon, M.C.; Nuttall, F.Q. Meal stimulation of cortisol secretion: A protein induced effect. Metabolism 1981, 30, 1104–1108. [Google Scholar] [CrossRef]
- Follenius, M.; Brandenberger, G.; Hietter, B. Diurnal cortisol peaks and their relationships to meals. J. Clin. Endocrinol. Metab. 1982, 55, 757–761. [Google Scholar] [CrossRef]
- Iranmanesh, A.; Lawson, D.; Dunn, B.; Veldhuis, J.D. Glucose ingestion selectively amplifies ACTH and cortisol secretory-burst mass and enhances their joint synchrony in healthy men. J. Clin. Endocrinol. Metab. 2011, 96, 2882–2888. [Google Scholar] [CrossRef] [PubMed]
- Kassi, E.; Papavassiliou, A.G. Glucose can promote a glucocorticoid resistance state. J. Cell. Mol. Med. 2012, 16, 1146–1149. [Google Scholar] [CrossRef]
- Vincent, A.M.; Russell, J.W.; Low, P.; Feldman, E.L. Oxidative stress in the pathogenesis of diabetic neuropathy. Endocr. Rev. 2004, 25, 612–628. [Google Scholar] [CrossRef]
- Volpe, C.M.O.; Villar-Delfino, P.H.; Dos Anjos, P.M.F.; Nogueira-Machado, J.A. Cellular death, reactive oxygen species (ROS) and diabetic complications. Cell Death Dis. 2018, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- van Rossum, E.F.; Koper, J.W.; Huizenga, N.A.; Uitterlinden, A.G.; Janssen, J.A.; Brinkmann, A.O.; Grobbee, D.E.; de Jong, F.H.; van Duyn, C.M.; Pols, H.A.; et al. A polymorphism in the glucocorticoid receptor gene, which decreases sensitivity to glucocorticoids in vivo, is associated with low insulin and cholesterol levels. Diabetes 2002, 51, 3128–3134. [Google Scholar] [CrossRef] [PubMed]
- van Raalte, D.H.; van Leeuwen, N.; Simonis-Bik, A.M.; Nijpels, G.; van Haeften, T.W.; Schafer, S.A.; Boomsma, D.I.; Kramer, M.H.; Heine, R.J.; Maassen, J.A.; et al. Glucocorticoid receptor gene polymorphisms are associated with reduced first-phase glucose-stimulated insulin secretion and disposition index in women, but not in men. Diabet. Med. 2012, 29, e211–e216. [Google Scholar] [CrossRef]
- van Rossum, E.F.; Voorhoeve, P.G.; te Velde, S.J.; Koper, J.W.; Delemarre-van de Waal, H.A.; Kemper, H.C.; Lamberts, S.W. The ER22/23EK polymorphism in the glucocorticoid receptor gene is associated with a beneficial body composition and muscle strength in young adults. J. Clin. Endocrinol. Metab. 2004, 89, 4004–4009. [Google Scholar] [CrossRef] [PubMed]
- Rosmond, R.; Chagnon, Y.C.; Chagnon, M.; Perusse, L.; Bouchard, C.; Bjorntorp, P. A polymorphism of the 5’-flanking region of the glucocorticoid receptor gene locus is associated with basal cortisol secretion in men. Metabolism 2000, 49, 1197–1199. [Google Scholar] [CrossRef] [PubMed]
- van Rossum, E.F.; Roks, P.H.; de Jong, F.H.; Brinkmann, A.O.; Pols, H.A.; Koper, J.W.; Lamberts, S.W. Characterization of a promoter polymorphism in the glucocorticoid receptor gene and its relationship to three other polymorphisms. Clin. Endocrinol. 2004, 61, 573–581. [Google Scholar] [CrossRef]
- Rosmond, R.; Holm, G. A 5-year follow-up study of 3 polymorphisms in the human glucocorticoid receptor gene in relation to obesity, hypertension, and diabetes. J. Cardiometab. Syndr. 2008, 3, 132–135. [Google Scholar] [CrossRef] [PubMed]
- Rosmond, R.; Chagnon, Y.C.; Holm, G.; Chagnon, M.; Perusse, L.; Lindell, K.; Carlsson, B.; Bouchard, C.; Bjorntorp, P. A glucocorticoid receptor gene marker is associated with abdominal obesity, leptin, and dysregulation of the hypothalamic-pituitary-adrenal axis. Obes. Res. 2000, 8, 211–218. [Google Scholar] [CrossRef]
- Weaver, J.U.; Hitman, G.A.; Kopelman, P.G. An association between a Bc1I restriction fragment length polymorphism of the glucocorticoid receptor locus and hyperinsulinaemia in obese women. J Mol. Endocrinol. 1992, 9, 295–300. [Google Scholar] [CrossRef]
- Geelen, C.C.; van Greevenbroek, M.M.; van Rossum, E.F.; Schaper, N.C.; Nijpels, G.; t Hart, L.M.; Schalkwijk, C.G.; Ferreira, I.; van der Kallen, C.J.; Sauerwein, H.P.; et al. BclI glucocorticoid receptor polymorphism is associated with greater body fatness: The Hoorn and CODAM studies. J. Clin. Endocrinol. Metab. 2013, 98, E595–E599. [Google Scholar] [CrossRef] [PubMed][Green Version]
- van Rossum, E.F.; Koper, J.W.; van den Beld, A.W.; Uitterlinden, A.G.; Arp, P.; Ester, W.; Janssen, J.A.; Brinkmann, A.O.; de Jong, F.H.; Grobbee, D.E.; et al. Identification of the BclI polymorphism in the glucocorticoid receptor gene: Association with sensitivity to glucocorticoids in vivo and body mass index. Clin. Endocrinol. 2003, 59, 585–592. [Google Scholar] [CrossRef]
- Giordano, R.; Marzotti, S.; Berardelli, R.; Karamouzis, I.; Brozzetti, A.; D’Angelo, V.; Mengozzi, G.; Mandrile, G.; Giachino, D.; Migliaretti, G.; et al. BClI polymorphism of the glucocorticoid receptor gene is associated with increased obesity, impaired glucose metabolism and dyslipidaemia in patients with Addison’s disease. Clin. Endocrinol. 2012, 77, 863–870. [Google Scholar] [CrossRef]
- Koeijvoets, K.C.; van der Net, J.B.; van Rossum, E.F.; Steyerberg, E.W.; Defesche, J.C.; Kastelein, J.J.; Lamberts, S.W.; Sijbrands, E.J. Two common haplotypes of the glucocorticoid receptor gene are associated with increased susceptibility to cardiovascular disease in men with familial hypercholesterolemia. J. Clin. Endocrinol. Metab. 2008, 93, 4902–4908. [Google Scholar] [CrossRef] [PubMed]
- van Moorsel, D.; van Greevenbroek, M.M.; Schaper, N.C.; Henry, R.M.; Geelen, C.C.; van Rossum, E.F.; Nijpels, G.; t Hart, L.M.; Schalkwijk, C.G.; van der Kallen, C.J.; et al. BclI glucocorticoid receptor polymorphism in relation to cardiovascular variables: The Hoorn and CODAM studies. Eur. J. Endocrinol. 2015, 173, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Huizenga, N.A.; Koper, J.W.; De Lange, P.; Pols, H.A.; Stolk, R.P.; Burger, H.; Grobbee, D.E.; Brinkmann, A.O.; De Jong, F.H.; Lamberts, S.W. A polymorphism in the glucocorticoid receptor gene may be associated with and increased sensitivity to glucocorticoids in vivo. J. Clin. Endocrinol. Metab. 1998, 83, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Bayramci, N.S.; Acik, L.; Kalkan, C.; Yetkin, I. Investigation of glucocorticoid receptor and calpain-10 gene polymorphisms in Turkish patients with type 2 diabetes mellitus. Turk. J. Med. Sci. 2017, 47, 1568–1575. [Google Scholar] [CrossRef]
- Roussel, R.; Reis, A.F.; Dubois-Laforgue, D.; Bellanne-Chantelot, C.; Timsit, J.; Velho, G. The N363S polymorphism in the glucocorticoid receptor gene is associated with overweight in subjects with type 2 diabetes mellitus. Clin. Endocrinol. 2003, 59, 237–241. [Google Scholar] [CrossRef]
- Lin, R.C.; Wang, W.Y.; Morris, B.J. High penetrance, overweight, and glucocorticoid receptor variant: Case-control study. BMJ 1999, 319, 1337–1338. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jewell, C.M.; Katen, K.S.; Barber, L.M.; Cannon, C.; Garantziotis, S.; Cidlowski, J.A. Healthy glucocorticoid receptor N363S carriers dysregulate gene expression associated with metabolic syndrome. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E741–E748. [Google Scholar] [CrossRef][Green Version]
- Moller, H.J.; Frikke-Schmidt, R.; Moestrup, S.K.; Nordestgaard, B.G.; Tybjaerg-Hansen, A. Serum soluble CD163 predicts risk of type 2 diabetes in the general population. Clin. Chem. 2011, 57, 291–297. [Google Scholar] [CrossRef]
- Dasu, M.R.; Ramirez, S.; Isseroff, R.R. Toll-like receptors and diabetes: A therapeutic perspective. Clin. Sci. 2012, 122, 203–214. [Google Scholar] [CrossRef]
- Derijk, R.H.; Schaaf, M.J.; Turner, G.; Datson, N.A.; Vreugdenhil, E.; Cidlowski, J.; de Kloet, E.R.; Emery, P.; Sternberg, E.M.; Detera-Wadleigh, S.D. A human glucocorticoid receptor gene variant that increases the stability of the glucocorticoid receptor beta-isoform mRNA is associated with rheumatoid arthritis. J. Rheumatol. 2001, 28, 2383–2388. [Google Scholar]
- Syed, A.A.; Irving, J.A.; Redfern, C.P.; Hall, A.G.; Unwin, N.C.; White, M.; Bhopal, R.S.; Weaver, J.U. Association of glucocorticoid receptor polymorphism A3669G in exon 9beta with reduced central adiposity in women. Obesity 2006, 14, 759–764. [Google Scholar] [CrossRef]
- Trementino, L.; Appolloni, G.; Concettoni, C.; Cardinaletti, M.; Boscaro, M.; Arnaldi, G. Association of glucocorticoid receptor polymorphism A3669G with decreased risk of developing diabetes in patients with Cushing’s syndrome. Eur. J. Endocrinol. 2012, 166, 35–42. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Paul, P.; Chakraborty, A.; Sarkar, D.; Langthasa, M.; Rahman, M.; Bari, M.; Singha, R.S.; Malakar, A.K.; Chakraborty, S. Interplay between miRNAs and human diseases. J. Cell. Physiol. 2018, 233, 2007–2018. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.; Li, Y.Q.; Liang, Y.Z.; Dong, J.; He, Y.; Zhang, L.; Yan, Y.X. Expression of miR-18a and miR-34c in circulating monocytes associated with vulnerability to type 2 diabetes mellitus and insulin resistance. J. Cell. Mol. Med. 2017, 21, 3372–3380. [Google Scholar] [CrossRef]
- Wang, Z.; Miu, K.K.; Zhang, X.; Wan, A.T.; Lu, G.; Cheung, H.H.; Lee, H.M.; Kong, A.P.; Chan, J.C.; Chan, W.Y. Hepatic miR-192-3p reactivation alleviates steatosis by targeting glucocorticoid receptor. JHEP Rep. 2020, 2, 100179. [Google Scholar] [CrossRef]
- Yadav, S.K.; Guleria, P. Steviol glycosides from Stevia: Biosynthesis pathway review and their application in foods and medicine. Crit. Rev. Food Sci. Nutr. 2012, 52, 988–998. [Google Scholar] [CrossRef]
- Boonkaewwan, C.; Burodom, A. Anti-inflammatory and immunomodulatory activities of stevioside and steviol on colonic epithelial cells. J. Sci. Food Agric. 2013, 93, 3820–3825. [Google Scholar] [CrossRef]
- Brahmachari, G.; Mandal, L.C.; Roy, R.; Mondal, S.; Brahmachari, A.K. Stevioside and related compounds—Molecules of pharmaceutical promise: A critical overview. Arch. Pharm. 2011, 344, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Chatsudthipong, V.; Muanprasat, C. Stevioside and related compounds: Therapeutic benefits beyond sweetness. Pharmacol. Ther. 2009, 121, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Sehar, I.; Kaul, A.; Bani, S.; Pal, H.C.; Saxena, A.K. Immune up regulatory response of a non-caloric natural sweetener, stevioside. Chem. Biol. Interact. 2008, 173, 115–121. [Google Scholar] [CrossRef]
- Chang, S.F.; Yang, L.M.; Hsu, F.L.; Hsu, J.Y.; Liaw, J.H.; Lin, S.J. Transformation of steviol-16alpha,17-epoxide by Streptomyces griseus and Cunninghamella bainieri. J. Nat. Prod. 2006, 69, 1450–1455. [Google Scholar] [CrossRef] [PubMed]
- Panagiotou, C.; Mihailidou, C.; Brauhli, G.; Katsarou, O.; Moutsatsou, P. Effect of steviol, steviol glycosides and stevia extract on glucocorticoid receptor signaling in normal and cancer blood cells. Mol. Cell. Endocrinol. 2018, 460, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Dusek, J.; Carazo, A.; Trejtnar, F.; Hyrsova, L.; Holas, O.; Smutny, T.; Micuda, S.; Pavek, P. Steviol, an aglycone of steviol glycoside sweeteners, interacts with the pregnane X (PXR) and aryl hydrocarbon (AHR) receptors in detoxification regulation. Food Chem. Toxicol. 2017, 109, 130–142. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, Z.; Pavek, P. Regulation of drug-metabolizing cytochrome P450 enzymes by glucocorticoids. Drug Metab. Rev. 2010, 42, 621–635. [Google Scholar] [CrossRef]
- Corcuff, J.B.; Brossaud, J. Rebaudioside A and cortisol metabolism: Sweet news for consumers. Clin. Chim. Acta 2014, 431, 276–277. [Google Scholar] [CrossRef]
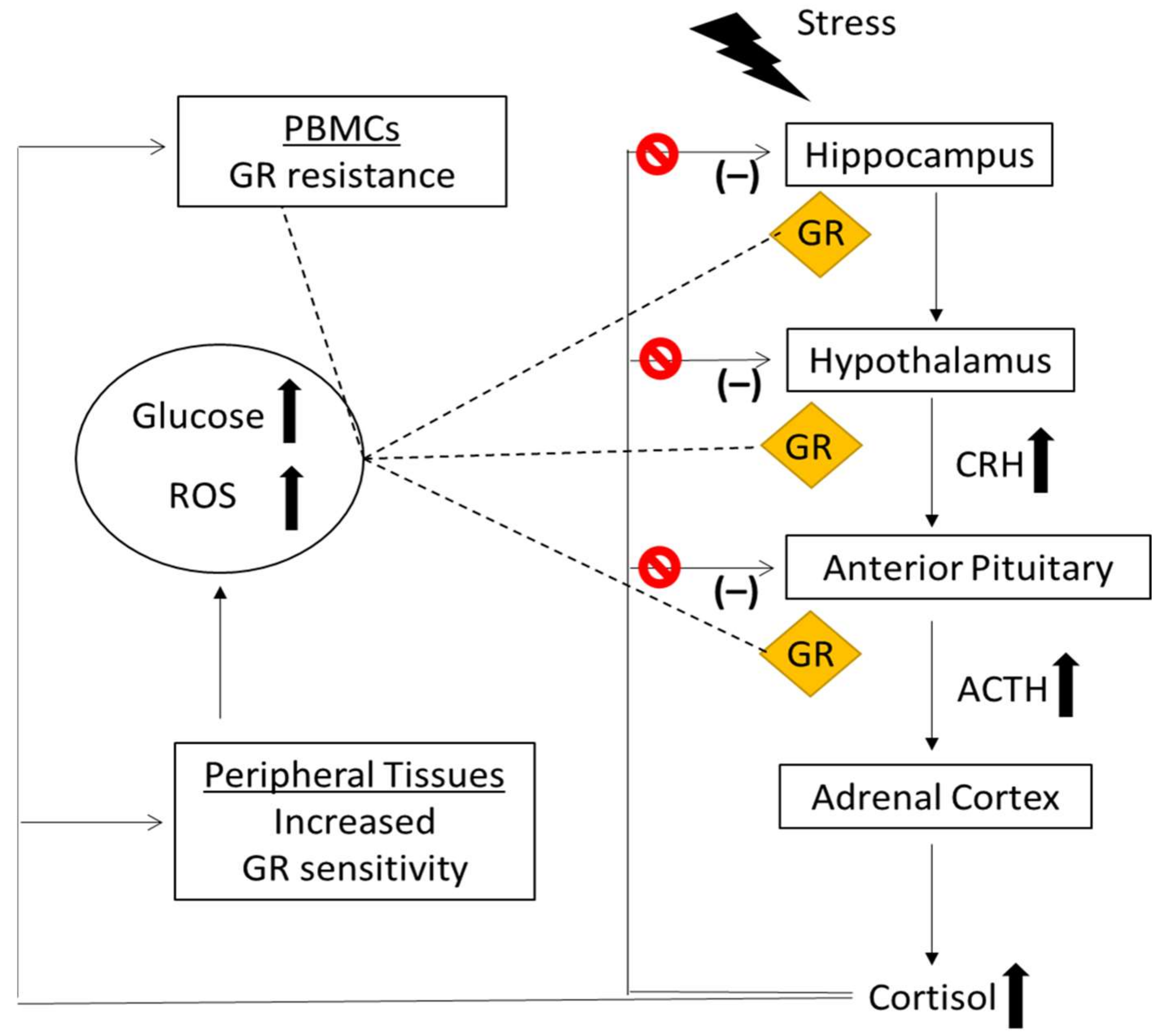
 increased levels,
increased levels,  impairment (blockade) of the HPA axis negative feedback, GR; Glucocor-ticoid Receptor, CRH; Corticotropin Releasing Hormone, ACTH; Adrenocorticotropic Hormone, PBMCs; Peripheral Blood Mononuclear Cells, ROS; Reactive Oxygen Species.
increased levels, impairment (blockade) of the HPA axis negative feedback, GR; Glucocor-ticoid Receptor, CRH; Corticotropin Releasing Hormone, ACTH; Adrenocorticotropic Hormone, PBMCs; Peripheral Blood Mononuclear Cells, ROS; Reactive Oxygen Species.
impairment (blockade) of the HPA axis negative feedback, GR; Glucocor-ticoid Receptor, CRH; Corticotropin Releasing Hormone, ACTH; Adrenocorticotropic Hormone, PBMCs; Peripheral Blood Mononuclear Cells, ROS; Reactive Oxygen Species.
increased levels, impairment (blockade) of the HPA axis negative feedback, GR; Glucocor-ticoid Receptor, CRH; Corticotropin Releasing Hormone, ACTH; Adrenocorticotropic Hormone, PBMCs; Peripheral Blood Mononuclear Cells, ROS; Reactive Oxygen Species.

{kind=link}
| Patients/Sample | GR 1 Signaling Parameters | GC 2 Sensitivity Assay | GC Response | Ref. |
|---|---|---|---|---|
| T2D 3 patients vs. controls (PBMCs 4) | pGR-S211 protein levels ↓ | [126] | ||
| GRβ mRNA levels ↑ | ||||
| Basal GILZ mRNA levels ↓ Dex 5-induced GILZ mRNA levels ↓ | Basal and Dex induced expression of GR-stimulated genes | Reduced GC sensitivity (GC resistance) | ||
| Basal FKBP5 mRNA levels ↓ Dex-induced FKBP5 mRNA levels ↓ | Basal and Dex induced expression of GR-stimulated genes | Reduced GC sensitivity (GC resistance) | ||
| T2D patients vs controls (Whole Blood Cells) | Less capable of producing IL-6 after LPS 6 stimulation Lower Dex concentration needed to inhibit LPS-induced IL-6 | Dex inhibition of LPS-induced IL-6 production | Increased GC sensitivity | [127] |
| GR 1 Polymorphisms | GC 2 Response | Refs | Metabolic Indices | Population | Ref. |
|---|---|---|---|---|---|
| ER22/23EK | GC resistance | [139] | total and LDL 3 cholesterol levels, fasting insulin levels ↓ | Healthy elderly subjects | [139] |
| insulin secretion, disposition index ↓ | Women with normal/impaired glucose tolerance | [140] | |||
| muscle strength and beneficial body composition ↑ | Healthy subjects | [141] | |||
| Tth111I | basal and bedtime salivary cortisol levels ↑ | Middle-aged men | [142] | ||
| ER22/23EK and Tth111I | GC resistance | [143] | fasting insulin levels, total and LDL cholesterol levels ↓ | Elderly subjects | [143] |
| BclI | Increased GC sensitivity | [148] | body weight, BMI 4 ↑ abdominal obesity fasting plasma glucose, insulin, HOMA 5 ↑ | Swedish men born in 1944 | [144] |
| salivary cortisol values after stimulation by a standardized lunch ↑ abdominal obesity | Middle-aged men | [145] | |||
| fasting insulin, HOMA↑ | Women with obesity | [146] | |||
| total body fatness, insulin resistance ↑ | Participants from CODAM 8 and the Hoorn study | [147] | |||
| central adiposity, impaired glucose tolerance, diabetes mellitus, dyslipidemia | Patients with Addison’s disease | [149] | |||
| N363S | Increased GC sensitivity | [152] | obesity, weight, BMI, HDL 6 ↑ | Patients with T2D | [153] |
| overweight | Patients with T2D | [154] | |||
| obesity | Non-diabetic white subjects of British descent | [155] | |||
| mRNA levels HSD11B1 ↑ mRNA levels of IGF-1 ↓ mRNA levels of IRS1 ↓ | Healthy subjects | [156] | |||
| disposition index ↓ | Women with normal/impaired glucose tolerance | [140] | |||
| GR-9β | GC resistance | [159] | risk of obesity ↓ total cholesterol levels ↓ HDL↑ | Europid women | [160] |
| risk of developing T2D 7 ↓ | Patients with Cushing’s syndrome | [161] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kokkinopoulou, I.; Diakoumi, A.; Moutsatsou, P. Glucocorticoid Receptor Signaling in Diabetes. Int. J. Mol. Sci. 2021, 22, 11173. https://doi.org/10.3390/ijms222011173
Kokkinopoulou I, Diakoumi A, Moutsatsou P. Glucocorticoid Receptor Signaling in Diabetes. International Journal of Molecular Sciences. 2021; 22(20):11173. https://doi.org/10.3390/ijms222011173
Chicago/Turabian StyleKokkinopoulou, Ioanna, Andriana Diakoumi, and Paraskevi Moutsatsou. 2021. "Glucocorticoid Receptor Signaling in Diabetes" International Journal of Molecular Sciences 22, no. 20: 11173. https://doi.org/10.3390/ijms222011173
APA StyleKokkinopoulou, I., Diakoumi, A., & Moutsatsou, P. (2021). Glucocorticoid Receptor Signaling in Diabetes. International Journal of Molecular Sciences, 22(20), 11173. https://doi.org/10.3390/ijms222011173