
IL-6 in the Ecosystem of Head and Neck Cancer: Possible Therapeutic Perspectives

 ,
,  , , ,
, , ,  and
and 
Abstract
1. Introduction
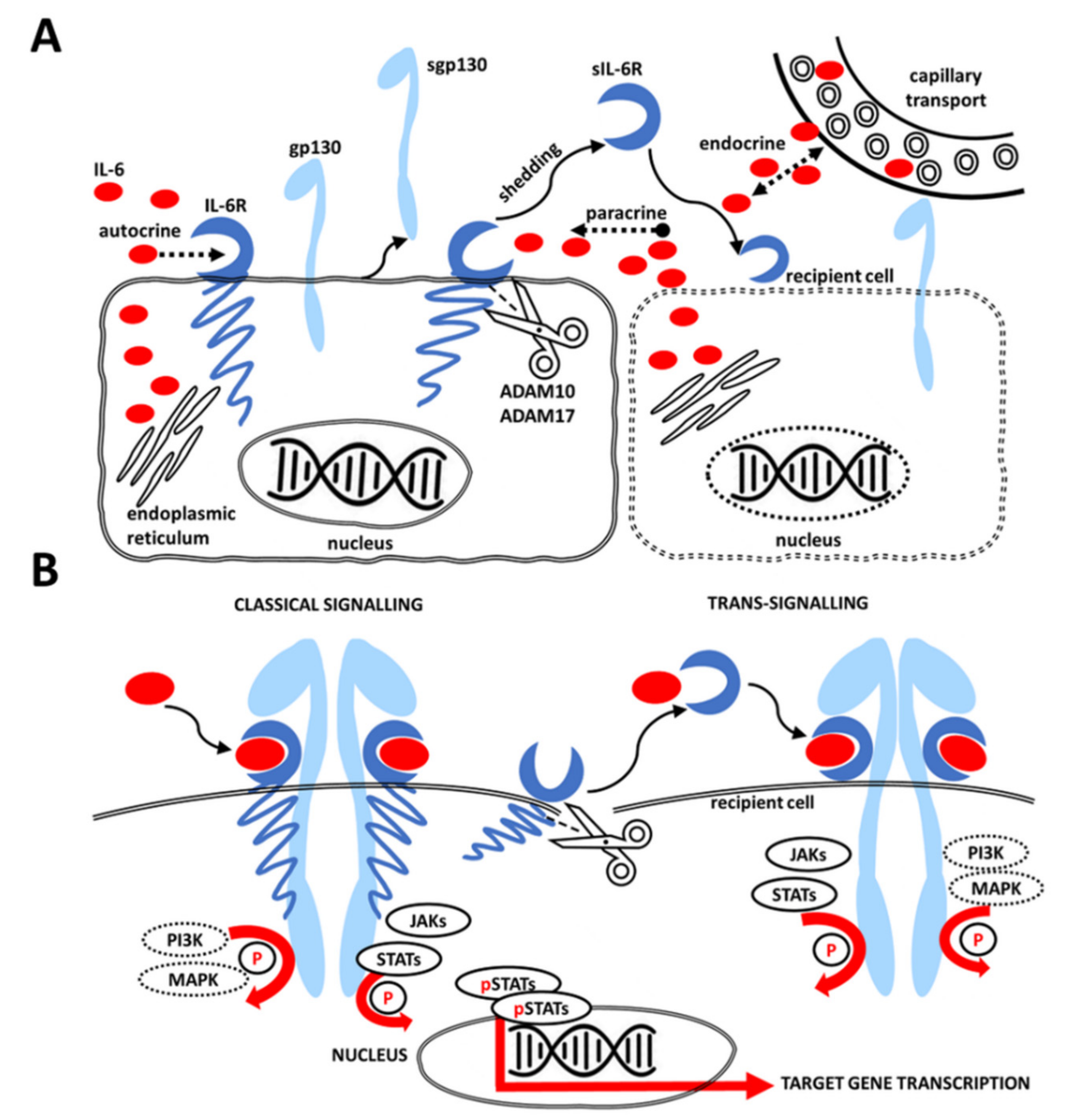
2. IL-6 Signaling Pathway
3. IL-6 in the Context of Head and Neck Cancer
3.1. Tumor Microenvironment
3.2. Cancer-Associated Fibroblasts (CAFs)
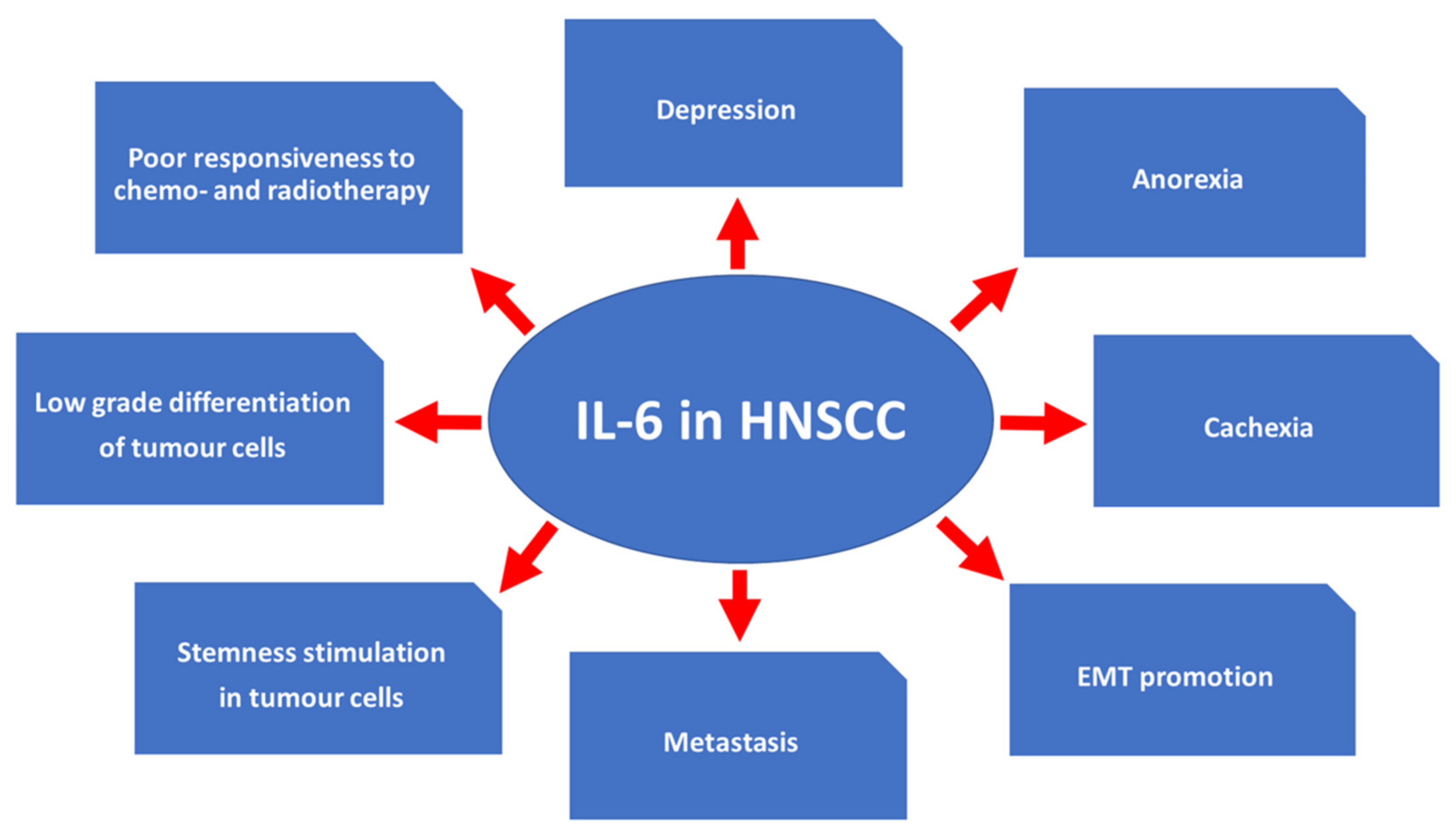
3.3. Effects of IL-6 Expression
3.4. Serum and Salivary Concentrations of IL-6 in HNSCC and Their Potential Role in Diagnostics
3.5. Anti-IL-6 Therapy
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| α-SMA | α-Smooth muscle actin |
| CAF | Cancer-associated fibroblast |
| CBP | CREB-binding protein |
| ECM | Extracellular matrix |
| EMT | Epithelial to mesenchymal transition |
| GAB1 | GRB2-associated-binding protein |
| gp130 | Β-Receptor glycoprotein 130 |
| HNC | Head and neck cancer |
| HNSCC | Head and neck squamous cell carcinoma |
| HPV | Human papilloma virus |
| IFN | Interferon |
| IL-6 | Interleukin 6 |
| IL-8 | Interleukin 8 |
| IL-6R | Receptor for IL-6 |
| JAK | Janus kinase |
| JAK-STAT | Janus kinase and signal transducer and activator of transcription |
| MAPK | Mitogen-activated protein kinase |
| OPMD | Oral potentially malignant disorder |
| OSCC | Oral squamous cell carcinoma |
| PIAS | Protein inhibitor of activated STAT |
| PI3K | Phosphatidylinositol-3-kinase |
| RA | Rheumatoid arthritis |
| SOCS3 | Suppressor of cytokine signaling 3 |
| STAT | Signal transducer and activator of transcription |
| TME | Tumor microenvironment |
| TNF-α | Tumor necrosis factor α |
References
- Warnakulasuriya, S. Global Epidemiology of Oral and Oropharyngeal Cancer. Oral Oncol. 2009, 45, 309–316. [Google Scholar] [CrossRef]
- Vokes, E.E.; Weichselbaum, R.R.; Lippman, S.M.; Hong, W.K. Head and Neck Cancer. N. Engl. J. Med. 1993, 328. [Google Scholar] [CrossRef] [PubMed]
- Novák, Š.; Bandurová, V.; Mifková, A.; Kalfeřt, D.; Fík, Z.; Lukeš, P.; Szabo, P.; Plzák, J.; Smetana, K., Jr. Tumor Microenvironment. Otorinolaryng. A Foniat. 2019, 68, 41–51. [Google Scholar]
- Smetana, K.; Lacina, L.; Szabo, P.; Dvořánková, B.; Brož, P.; Šedo, A. Ageing as an Important Risk Factor for Cancer. Anticancer Res. 2016, 36, 5009–5017. [Google Scholar] [CrossRef] [PubMed]
- Saussez, S.; Duray, A.; Demoulin, S.; Hubert, P.; Delvenne, P. Immune Suppression in Head and Neck Cancers: A Review. Clin. Dev. Immunol. 2010, 2010, 15. [Google Scholar] [CrossRef]
- Lacina, L.; Brábek, J.; Král, V.; Kodet, O.; Smetana, K. Interleukin-6: A Molecule with Complex Biological Impact in Cancer. Histol. Histopathol. 2019, 34, 125–136. [Google Scholar] [CrossRef]
- Hamburger, A.W.; Salmon, S.E. Primary Bioassay of Human Tumor Stem Cells. Science 1977, 197, 461–463. [Google Scholar] [CrossRef]
- Metwaly, H.; Maruyama, S.; Yamazaki, M.; Tsuneki, M.; Abé, T.; Jen, K.Y.; Cheng, J.; Saku, T. Parenchymal-Stromal Switching for Extracellular Matrix Production on Invasion of Oral Squamous Cell Carcinoma. Hum. Pathol. 2012, 43, 1973–1981. [Google Scholar] [CrossRef]
- Polyak, K.; Haviv, I.; Campbell, I.G. Co-Evolution of Tumor Cells and Their Microenvironment. Trends Genet. 2009, 25, 30–38. [Google Scholar] [CrossRef]
- Lacina, L.; Plzak, J.; Kodet, O.; Szabo, P.; Chovanec, M.; Dvorankova, B.; Smetana, K. Cancer Microenvironment: What Can We Learn from the Stem Cell Niche. Int. J. Mol. Sci. 2015, 16, 24094–24110. [Google Scholar] [CrossRef]
- Li, H.; Fan, X.; Houghton, J.M. Tumor Microenvironment: The Role of the Tumor Stroma in Cancer. J. Cell. Biochem. 2007, 101, 805–815. [Google Scholar] [CrossRef]
- Lorusso, G.; Rüegg, C. The Tumor Microenvironment and Its Contribution to Tumor Evolution toward Metastasis. Histochem. Cell Biol. 2008, 130, 1091–1103. [Google Scholar] [CrossRef]
- Plzák, J.; Lacina, L.; Chovanec, M.; Dvořánková, B.; Szabo, P.; Čada, Z.; Smetana, K. Epithelial–Stromal Interaction in Squamous Cell Epithelium-Derived Tumors: An Important New Player in the Control of Tumor Biological Properties. Anticancer Res. 2010, 30. [Google Scholar]
- Fisher, D.T.; Appenheimer, M.M.; Evans, S.S. The Two Faces of IL-6 in the Tumor Microenvironment. Semin. Immunol. 2014, 26, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Kolář, M.; Szabo, P.; Dvořánková, B.; Lacina, L.; Gabius, H.J.; Strnad, H.; Šáchová, J.; Vlček, Č.; Plzák, J.; Chovanec, M.; et al. Upregulation of IL-6, IL-8 and CXCL-1 Production in Dermal Fibroblasts by Normal/Malignant Epithelial Cells in Vitro: Immunohistochemical and Transcriptomic Analyses. Biol. Cell 2012, 104, 738–751. [Google Scholar] [CrossRef] [PubMed]
- Rossi, J.F.; Lu, Z.Y.; Jourdan, M.; Klein, B. Interleukin-6 as a Therapeutic Target. Clin. Cancer Res. 2015, 21, 1248–1257. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, K.; Taga, T.; Hirata, Y.; Yawata, H.; Kawanishi, Y.; Seed, B.; Taniguchi, T.; Hirano, T.; Kishimoto, T. Cloning and Expression of the Human Interleukin-6 (BSF-2/IFNβ 2) Receptor. Science 1988, 241, 825–828. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Yasukawa, K.; Harada, H.; Taga, T.; Watanabe, Y.; Matsuda, T.; Kashiwamura, S.I.; Nakajima, K.; Koyama, K.; Iwamatsu, A.; et al. Complementary DNA for a Novel Human Interleukin (BSF-2) That Induces B Lymphocytes to Produce Immunoglobulin. Nature 1986, 324, 73–76. [Google Scholar] [CrossRef]
- Scheller, J.; Rose-John, S. Interleukin-6 and Its Receptor: From Bench to Bedside. Med. Microbiol. Immunol. 2006, 195, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Rose-John, S. Interleukin-6 Biology Is Coordinated by Membrane-Bound and Soluble Receptors: Role in Inflammation and Cancer. J. Leukoc. Biol. 2006, 80, 227–236. [Google Scholar] [CrossRef]
- Wolf, J.; Rose-John, S.; Garbers, C. Interleukin-6 and Its Receptors: A Highly Regulated and Dynamic System. Cytokine 2014, 70, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Schaper, F.; Rose-John, S. Interleukin-6: Biology, Signaling and Strategies of Blockade. Cytokine Growth Factor Rev. 2015, 26, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, S.; Lange, T.; Benedict, C.; Nowell, M.A.; Jones, S.A.; Scheller, J.; Rose-John, S.; Born, J.; Dimitrov, S.; Lange, T.; et al. Sleep Enhances IL-6 Trans-signaling in Humans. FASEB J. 2006, 20, 2174–2176. [Google Scholar] [CrossRef] [PubMed]
- Scheller, J.; Garbers, C.; Rose-John, S. Interleukin-6: From Basic Biology to Selective Blockade of pro-Inflammatory Activities. Semin. Immunol. 2014, 26, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Jostock, T.; Müllberg, J.; Özbek, S.; Atreya, R.; Blinn, G.; Voltz, N.; Fischer, M.; Neurath, M.F.; Rose-John, S. Soluble Gp130 Is the Natural Inhibitor of Soluble Interleukin-6 Receptor Transsignaling Responses. Eur. J. Biochem. 2001, 268, 160–167. [Google Scholar] [CrossRef]
- Heinrich, P.C.; Behrmann, I.; Haan, S.; Hermanns, H.M.; Müller-Newen, G.; Schaper, F. Principles of Interleukin (IL)-6-Type Cytokine Signalling and Its Regulation. Biochem. J. 2003, 374, 1–20. [Google Scholar] [CrossRef]
- Eulenfeld, R.; Dittrich, A.; Khouri, C.; Müller, P.J.; Mütze, B.; Wolf, A.; Schaper, F. Interleukin-6 Signalling: More than Jaks and STATs. Eur. J. Cell Biol. 2012, 91, 486–495. [Google Scholar] [CrossRef]
- Kretzschmar, A.K.; Dinger, M.C.; Henze, C.; Brocke-Heidrich, K.; Horn, F. Analysis of Stat3 (Signal Transducer and Activator of Transcription 3) Dimerization by Fluorescence Resonance Energy Transfer in Living Cells. Biochem. J. 2004, 377, 289–297. [Google Scholar] [CrossRef]
- Braunstein, J.; Brutsaert, S.; Olson, R.; Schindler, C. STATs Dimerize in the Absence of Phosphorylation. J. Biol. Chem. 2003, 278, 34133–34140. [Google Scholar] [CrossRef]
- Haan, S.; Knotholes, M.; Behrmann, I.; Müller-Esterl, W.; Heinrich, P.C.; Schaper, F. Cytoplasmic STAT Proteins Associate Prior to Activation. Biochem. J. 2000, 345, 417–421. [Google Scholar] [CrossRef]
- Novak, U.; Ji, H.; Kanagasundaram, V.; Simpson, R.; Paradiso, L. STAT3 Forms Stable Homodimers in the Presence of Divalent Cations Prior to Activation. Biochem. Biophys. Res. Commun. 1998, 247, 558–563. [Google Scholar] [CrossRef]
- Vogt, M.; Domoszlai, T.; Kleshchanok, D.; Lehmann, S.; Schmitt, A.; Poli, V.; Richtering, W.; Müller-Newen, G. The Role of the N-Terminal Domain in Dimerization and Nucleocytoplasmic Shuttling of Latent STAT3. J. Cell Sci. 2011, 124, 900–909. [Google Scholar] [CrossRef]
- Pranada, A.L.; Metz, S.; Herrmann, A.; Heinrich, P.C.; Müller-Newen, G. Real Time Analysis of STAT3 Nucleocytoplasmic Shuttling. J. Biol. Chem. 2004, 279, 15114–15123. [Google Scholar] [CrossRef]
- Ogryzko, V.V.; Schiltz, R.L.; Russanova, V.; Howard, B.H.; Nakatani, Y. The Transcriptional Coactivators P300 and CBP Are Histone Acetyltransferases. Cell 1996, 87, 953–959. [Google Scholar] [CrossRef]
- Ray, S.; Boldogh, I.; Brasier, A.R. STAT3 NH2-Terminal Acetylation Is Activated by the Hepatic Acute-Phase Response and Required for IL-6 Induction of Angiotensinogen. Gastroenterology 2005, 129, 1616–1632. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.L.; Guan, Y.J.; Chatterjee, D.; Chin, Y.E. Stat3 Dimerization Regulated by Reversible Acetylation of a Single Lysine Residue. Science 2005, 307, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.; Ray, S.; Lee, C.; Brasier, A.R. The STAT3 NH2-Terminal Domain Stabilizes Enhanceosome Assembly by Interacting with the P300 Bromodomain. J. Biol. Chem. 2008, 283, 30725–30734. [Google Scholar] [CrossRef] [PubMed]
- Fukada, T.; Hibi, M.; Yamanaka, Y.; Takahashi-Tezuka, M.; Fujitani, Y.; Yamaguchi, T.; Nakajima, K.; Hirano, T. Two Signals Are Necessary for Cell Proliferation Induced by a Cytokine Receptor Gp130: Involvement of STAT3 in Anti-Apoptosis. Immunity 1996, 5, 449–460. [Google Scholar] [CrossRef]
- Fukada, T.; Ohtani, T.; Yoshida, Y.; Shirogane, T.; Nishida, K.; Nakajima, K.; Hibi, M.; Hirano, T. STAT3 Orchestrates Contradictory Signals in Cytokine-Induced G1 to S Cell-Cycle Transition. EMBO J. 1998, 17, 6670–6677. [Google Scholar] [CrossRef]
- Judd, L.M.; Alderman, B.M.; Howlett, M.; Shulkes, A.; Dow, C.; Moverley, J.; Grail, D.; Jenkins, B.J.; Ernst, M.; Giraud, A.S. Gastric Cancer Development in Mice Lacking the SHP2 Binding Site on the IL-6 Family Co-Receptor Gp130. Gastroenterology 2004, 126, 196–207. [Google Scholar] [CrossRef]
- Tebbutt, N.C.; Giraud, A.S.; Inglese, M.; Jenkins, B.; Waring, P.; Clay, F.J.; Malki, S.; Alderman, B.M.; Grail, D.; Hollande, F.; et al. Reciprocal Regulation of Gastrointestinal Homeostasis by SHP2 and STAT-Mediated Trefoil Gene Activation in Gp130 Mutant Mice. Nat. Med. 2002, 8, 1089–1097. [Google Scholar] [CrossRef]
- Schaper, F.; Gendo, C.; Eck, M.; Schmitz, J.; Grimm, C.; Anhuf, D.; Kerr, I.M.; Heinrich, P.C. Activation of the Protein Tyrosine Phosphatase SHP2 via the Interleukin-6 Signal Transducing Receptor Protein Gp130 Requires Tyrosine Kinase Jak1 and Limits Acute-Phase Protein Expression. Biochem. J. 1998, 335, 557–565. [Google Scholar] [CrossRef]
- Symes, A.; Stahl, N.; Reeves, S.A.; Farruggella, T.; Servidei, T.; Gearan, T.; Yancopoulos, G.; Stephen Fink, J. The Protein Tyrosine Phosphatase SHP-2 Negatively Regulates Ciliary Neurotrophic Factor Induction of Gene Expression. Curr. Biol. 1997, 7, 697–700. [Google Scholar] [CrossRef]
- Schiemann, W.P.; Bartoe, J.L.; Nathanson, N.M. Box 3-Independent Signaling Mechanisms Are Involved in Leukemia Inhibitory Factor Receptor α- and Gp130-Mediated Stimulation of Mitogen-Activated Protein Kinase. Evidence for Participation of Multiple Signaling Pathways Which Converge at Ras. J. Biol. Chem. 1997, 272, 16631–16636. [Google Scholar] [CrossRef]
- Lai, C.F.; Ripperger, J.; Wang, Y.; Kim, H.; Hawley, R.B.; Baumann, H. The STAT3-Independent Signaling Pathway by Glycoprotein 130 in Hepatic Cells. J. Biol. Chem. 1999, 274, 7793–7802. [Google Scholar] [CrossRef] [PubMed]
- Ernst, M.; Jenkins, B.J. Acquiring Signalling Specificity from the Cytokine Receptor Gp130. Trends Genet. 2004, 20, 23–32. [Google Scholar] [CrossRef]
- Lehmann, U.; Schmitz, J.; Weissenbach, M.; Sobota, R.M.; Hörtner, M.; Friederichs, K.; Behrmann, I.; Tsiaris, W.; Sasaki, A.; Schneider-Mergener, J.; et al. SHP2 and SOCS3 Contribute to Tyr-759-Dependent Attenuation of Interleukin-6 Signaling through Gp130. J. Biol. Chem. 2003, 278, 661–671. [Google Scholar] [CrossRef]
- Kim, H.; Baumann, H. Dual Signaling Role of the Protein Tyrosine Phosphatase SHP-2 in Regulating Expression of Acute-Phase Plasma Proteins by Interleukin-6 Cytokine Receptors in Hepatic Cells. Mol. Cell. Biol. 1999, 19, 5326–5338. [Google Scholar] [CrossRef] [PubMed]
- De Souza, D.; Fabri, L.J.; Nash, A.; Hilton, D.J.; Nicola, N.A.; Baca, M. SH2 Domains from Suppressor of Cytokine Signaling-3 and Protein Tyrosine Phosphatase SHP-2 Have Similar Binding Specificities. Biochemistry 2002, 41, 9229–9236. [Google Scholar] [CrossRef]
- Eulenfeld, R.; Schaper, F. A New Mechanism for the Regulation of Gab1 Recruitment to the Plasma Membrane. J. Cell Sci. 2009, 122, 55–64. [Google Scholar] [CrossRef]
- Takahashi-Tezuka, M.; Yoshida, Y.; Fukada, T.; Ohtani, T.; Yamanaka, Y.; Nishida, K.; Nakajima, K.; Hibi, M.; Hirano, T. Gab1 Acts as an Adapter Molecule Linking the Cytokine Receptor Gp130 to ERK Mitogen-Activated Protein Kinase. Mol. Cell. Biol. 1998, 18, 4109–4117. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Fuller, G.M. Phosphorylation and Internalization of Gp130 Occur after IL-6 Activation of Jak2 Kinase in Hepatocytes. Mol. Biol. Cell 1994, 5, 819–828. [Google Scholar] [CrossRef]
- Zohlnhöfer, D.; Graeve, L.; Rose-John, S.; Schooltink, H.; Dittrich, E.; Heinrich, P.C. The Hepatic Interleukin-6 Receptor Down-Regulation of the Interleukin-6 Binding Subunit (Gp80) by Its Ligand. FEBS Lett. 1992, 306, 219–222. [Google Scholar] [CrossRef]
- Radtke, S.; Wüller, S.; Yang, X.P.; Lippok, B.E.; Mütze, B.; Mais, C.; Schmitz-Van De Leur, H.; Bode, J.G.; Gaestel, M.; Heinrich, P.C.; et al. Cross-Regulation of Cytokine Signalling: Pro-Inflammatory Cytokines Restrict IL-6 Signalling through Receptor Internalisation and Degradation. J. Cell Sci. 2010, 123, 947–959. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.D.; Liao, J.; Liu, B.; Rao, X.; Jay, P.; Berta, P.; Shuai, K. Specific Inhibition of Stat3 Signal Transduction by PIAS3. Science 1997, 278, 1803–1805. [Google Scholar] [CrossRef]
- Scatena, R.; Bottoni, P.; Pontoglio, A.; Giardina, B. Cancer Stem Cells: The Development of New Cancer Therapeutics. Expert Opin. Biol. Ther. 2011, 11, 875–892. [Google Scholar] [CrossRef]
- Korkaya, H.; Liu, S.; Wicha, M.S. Breast Cancer Stem Cells, Cytokine Networks, and the Tumor Microenvironment. J. Clin. Investig. 2011, 121, 3804–3809. [Google Scholar] [CrossRef]
- Gál, P.; Varinská, L.; Fáber, L.; Novák, Š.; Szabo, P.; Mitrengová, P.; Mirossay, A.; Mučaji, P.; Smetana, K. How Signaling Molecules Regulate Tumor Microenvironment: Parallels to Wound Repair. Molecules 2017, 22, 1818. [Google Scholar] [CrossRef]
- Brábek, J.; Jakubek, M.; Vellieux, F.; Novotný, J.; Kolář, M.; Lacina, L.; Szabo, P.; Strnadová, K.; Rösel, D.; Dvořánková, B.; et al. Interleukin-6: Molecule in the Intersection of Cancer, Ageing and COVID-19. Int. J. Mol. Sci. 2020, 21, 7937. [Google Scholar] [CrossRef]
- Östman, A.; Augsten, M. Cancer-Associated Fibroblasts and Tumor Growth - Bystanders Turning into Key Players. Curr. Opin. Genet. Dev. 2009, 19, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Lamertz, L.; Rummel, F.; Polz, R.; Baran, P.; Hansen, S.; Waetzig, G.H.; Moll, J.M.; Floss, D.M.; Scheller, J. Soluble Gp130 Prevents Interleukin-6 and Interleukin-11 Cluster Signaling but Not Intracellular Autocrine Responses. Sci. Signal. 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- Heink, S.; Yogev, N.; Garbers, C.; Herwerth, M.; Aly, L.; Gasperi, C.; Husterer, V.; Croxford, A.L.; Möller-Hackbarth, K.; Bartsch, H.S.; et al. Trans-Presentation of Interleukin-6 by Dendritic Cells Is Required for Priming Pathogenic TH17 Cells. Nat. Immunol. 2017, 18, 74. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.C.; Tsai, L.L.; Wang, M.L.; Yu, C.H.; Lo, W.L.; Chang, Y.C.; Chiou, G.Y.; Chou, M.Y.; Chiou, S.H. MiR145 Targets the SOX9/ADAM17 Axis to Inhibit Tumor-Initiating Cells and IL-6-Mediated Paracrine Effects in Head and Neck Cancer. Cancer Res. 2013, 73, 3425–3440. [Google Scholar] [CrossRef] [PubMed]
- Szabo, P.; Valach, J.; Smetana, K., Jr.; Dvorankova, B. Comparative Analysis of IL-8 and CXCL-1 Production by Normal and Cancer Stromal Fibroblasts. Folia Biol. 2013, 59, 134–137. [Google Scholar]
- Bremnes, R.M.; Dønnem, T.; Al-Saad, S.; Al-Shibli, K.; Andersen, S.; Sirera, R.; Camps, C.; Marinez, I.; Busund, L.T. The Role of Tumor Stroma in Cancer Progression and Prognosis: Emphasis on Carcinoma-Associated Fibroblasts and Non-Small Cell Lung Cancer. J. Thorac. Oncol. 2011, 6, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Novotný, J.; Strnadová, K.; Dvořánková, B.; Kocourková, Š.; Jakša, R.; Dundr, P.; Pačes, V.; Smetana, K.; Kolář, M.; Lacina, L. Single-Cell RNA Sequencing Unravels Heterogeneity of the Stromal Niche in Cutaneous Melanoma Heterogeneous Spheroids. Cancers 2020, 12, 3324. [Google Scholar] [CrossRef]
- Erez, N.; Truitt, M.; Olson, P.; Hanahan, D. Cancer-Associated Fibroblasts Are Activated in Incipient Neoplasia to Orchestrate Tumor-Promoting Inflammation in an NF-ΚB-Dependent Manner. Cancer Cell 2010, 17, 135–147. [Google Scholar] [CrossRef]
- Ganguly, D.; Chandra, R.; Karalis, J.; Teke, M.; Aguilera, T.; Maddipati, R.; Wachsmann, M.B.; Ghersi, D.; Siravegna, G.; Zeh, H.J.; et al. Cancer-Associated Fibroblasts: Versatile Players in the Tumor Microenvironment. Cancers 2020, 12, 2652. [Google Scholar] [CrossRef]
- Lacina, L.; Smetana, K.; Dvořánková, B.; Pytlík, R.; Kideryová, L.; Kučerová, L.; Plzáková, Z.; Štork, J.; Gabius, H.J.; André, S. Stromal Fibroblasts from Basal Cell Carcinoma Affect Phenotype of Normal Keratinocytes. Br. J. Dermatol. 2007, 156, 819–829. [Google Scholar] [CrossRef]
- Lacina, L.; Dvořánkova, B.; Smetana, K.; Chovanec, M.; Plzǎk, J.; Tachezy, R.; Kideryovǎ, L.; Kučerová, L.; Čada, Z.; Bouček, J.; et al. Marker Profiling of Normal Keratinocytes Identifies the Stroma from Squamous Cell Carcinoma of the Oral Cavity as a Modulatory Microenvironment in Co-Culture. Int. J. Radiat. Biol. 2007, 83, 837–848. [Google Scholar] [CrossRef]
- Strnad, H.; Lacina, L.; Kolář, M.; Čada, Z.; Vlček, Č.; Dvořánková, B.; Betka, J.; Plzák, J.; Chovanec, M.; Šáchova, J.; et al. Head and Neck Squamous Cancer Stromal Fibroblasts Produce Growth Factors Influencing Phenotype of Normal Human Keratinocytes. Histochem. Cell Biol. 2010, 133, 201–211. [Google Scholar] [CrossRef]
- Szabó, P.; Kolář, M.; Dvořánková, B.; Lacina, L.; Štork, J.; Vlček, Č.; Strnad, H.; Tvrdek, M.; Smetana, K. Mouse 3T3 Fibroblasts under the Influence of Fibroblasts Isolated from Stroma of Human Basal Cell Carcinoma Acquire Properties of Multipotent Stem Cells. Biol. Cell 2011, 103, 233–248. [Google Scholar] [CrossRef]
- Flier, J.S.; Underhill, L.H.; Dvorak, H.F. Tumors: Wounds That Do Not Heal. N. Engl. J. Med. 1986, 315, 1650–1659. [Google Scholar] [CrossRef]
- Dvoánková, B.; Szabo, P.; Lacina, L.; Gal, P.; Uhrova, J.; Zima, T.; Kaltner, H.; André, S.; Gabius, H.J.; Sykova, E.; et al. Human Galectins Induce Conversion of Dermal Fibroblasts into Myofibroblasts and Production of Extracellular Matrix: Potential Application in Tissue Engineering and Wound Repair. Cells Tissues Organs 2011, 194, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Valach, J.; Fík, Z.; Strnad, H.; Chovanec, M.; Plzák, J.; Čada, Z.; Szabo, P.; Šáchová, J.; Hroudová, M.; Urbanová, M.; et al. Smooth Muscle Actin-Expressing Stromal Fibroblasts in Head and Neck Squamous Cell Carcinoma: Increased Expression of Galectin-1 and Induction of Poor Prognosis Factors. Int. J. Cancer 2012, 131, 2499–2508. [Google Scholar] [CrossRef] [PubMed]
- Hinz, B. Formation and Function of the Myofibroblast during Tissue Repair. J. Investig. Dermatol. 2007, 127, 526–537. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.A.F.; Ghosh, K.; Tonnesen, M.G. Tissue Engineering for Cutaneous Wounds. J. Investig. Dermatol. 2007, 127, 1018–1029. [Google Scholar] [CrossRef]
- Deonarine, K.; Panelli, M.C.; Stashower, M.E.; Jin, P.; Smith, K.; Slade, H.B.; Norwood, C.; Wang, E.; Marincola, F.M.; Stroncek, D.F. Gene Expression Profiling of Cutaneous Wound Healing. J. Transl. Med. 2007, 5, 11. [Google Scholar] [CrossRef]
- Smetana, K.; Szabo, P.; Gál, P.; André, S.; Gabius, H.J.; Kodet, O.; Dvořánková, B. Emerging Role of Tissue Lectins as Microenvironmental Effectors in Tumors and Wounds. Histol. Histopathol. 2015, 30, 293–309. [Google Scholar] [CrossRef]
- Werner, S.; Krieg, T.; Smola, H. Keratinocyte-Fibroblast Interactions in Wound Healing. J. Investig. Dermatol. 2007, 127, 998–1008. [Google Scholar] [CrossRef]
- Slaughter, D.P.; Southwick, H.W.; Smejkal, W. “Field Cancerization” in Oral Stratified Squamous Epithelium. Clinical Implications of Multicentric Origin. Cancer 1953, 6, 963–968. [Google Scholar] [CrossRef]
- Ishii, T.; Suzuki, A.; Kuwata, T.; Hisamitsu, S.; Hashimoto, H.; Ohara, Y.; Yanagihara, K.; Mitsunaga, S.; Yoshino, T.; Kinoshita, T.; et al. Drug-Exposed Cancer-Associated Fibroblasts Facilitate Gastric Cancer Cell Progression Following Chemotherapy. Gastric Cancer 2021, 24. [Google Scholar] [CrossRef]
- Bharti, R.; Dey, G.; Mandal, M. Cancer Development, Chemoresistance, Epithelial to Mesenchymal Transition and Stem Cells: A Snapshot of IL-6 Mediated Involvement. Cancer Lett. 2016, 375, 51–61. [Google Scholar] [CrossRef]
- Krishnamurthy, S.; Warner, K.A.; Dong, Z.; Imai, A.; Nör, C.; Ward, B.B.; Helman, J.I.; Taichman, R.S.; Bellile, E.L.; McCauley, L.K.; et al. Endothelial Interleukin-6 Defines the Tumorigenic Potential of Primary Human Cancer Stem Cells. Stem Cells 2014, 32, 2845–2857. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Chen, Y.-C.; Nör, F.; Warner, K.A.; Andrews, A.; Wagner, V.P.; Zhang, Z.; Zhang, Z.; Martins, M.D.; Pearson, A.T.; et al. Endothelial-Derived Interleukin-6 Induces Cancer Stem Cell Motility by Generating a Chemotactic Gradient towards Blood Vessels. Oncotarget 2017, 8, 100339–100352. [Google Scholar] [CrossRef] [PubMed]
- Jinno, T.; Kawano, S.; Maruse, Y.; Matsubara, R.; Goto, Y.; Sakamoto, T.; Hashiguchi, Y.; Kaneko, N.; Tanaka, H.; Kitamura, R.; et al. Increased Expression of Interleukin-6 Predicts Poor Response to Chemoradiotherapy and Unfavorable Prognosis in Oral Squamous Cell Carcinoma. Oncol. Rep. 2015, 33, 2161–2168. [Google Scholar] [CrossRef]
- Gao, J.; Zhao, S.; Halstensen, T.S. Increased Interleukin-6 Expression Is Associated with Poor Prognosis and Acquired Cisplatin Resistance in Head and Neck Squamous Cell Carcinoma. Oncol. Rep. 2016, 35, 3265–3274. [Google Scholar] [CrossRef][Green Version]
- Matsuoka, Y.; Nakayama, H.; Yoshida, R.; Hirosue, A.; Nagata, M.; Tanaka, T.; Kawahara, K.; Sakata, J.; Arita, H.; Nakashima, H.; et al. IL-6 Controls Resistance to Radiation by Suppressing Oxidative Stress via the Nrf2-Antioxidant Pathway in Oral Squamous Cell Carcinoma. Br. J. Cancer 2016, 115, 1234–1244. [Google Scholar] [CrossRef]
- De Schutter, H.; Landuyt, W.; Verbeken, E.; Goethals, L.; Hermans, R.; Nuyts, S. The Prognostic Value of the Hypoxia Markers CA IX and GLUT I and the Cytokines VEGF and IL 6 in Head and Neck Squamous Cell Carcinoma Treated by Radiotherapy ± Chemotherapy. BMC Cancer 2005, 5, 42. [Google Scholar] [CrossRef] [PubMed]
- Ara, T.; DeClerck, Y.A. Interleukin-6 in Bone Metastasis and Cancer Progression. Eur. J. Cancer 2010, 46, 1223–1231. [Google Scholar] [CrossRef]
- Duffy, S.A.; Taylor, J.M.G.; Terrell, J.E.; Islam, M.; Li, Y.; Fowler, K.E.; Wolf, G.T.; Teknos, T.N. Interleukin-6 Predicts Recurrence and Survival among Head and Neck Cancer Patients. Cancer 2008, 113, 750–757. [Google Scholar] [CrossRef]
- Brailo, V.; Vucicevic-Boras, V.; Lukac, J.; Biocina-Lukenda, D.; Zilic-Alajbeg, I.; Milenovic, A.; Balija, M. Salivary and Serum Interleukin 1 Beta, Interleukin 6 and Tumor Necrosis Factor Alpha in Patients with Leukoplakia and Oral Cancer. Med. Oral Patol. Oral Cir. Bucal 2012, 17, e10. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, X.; Yin, K.E.; Li, T.; Bao, Y.; Chen, Z. Variation and Significance of Secretory Immunoglobulin A, Interleukin 6 and Dendritic Cells in Oral Cancer. Oncol. Lett. 2017, 13, 2297–2303. [Google Scholar] [CrossRef]
- Lotfi, A.; Shahidi, N.; Bayazian, G.; Abdollahi Fakhim, S.; Estakhri, R.; Esfahani, A.; Notash, R. Serum Level of Interleukin-6 in Patients with Oral Tongue Squamous Cell Carcinoma. Iran. J. Otorhinolaryngol. 2015, 27, 207–211. [Google Scholar] [CrossRef]
- Garbers, C.; Hermanns, H.M.; Schaper, F.; Müller-Newen, G.; Grötzinger, J.; Rose-John, S.; Scheller, J. Plasticity and Cross-Talk of Interleukin 6-Type Cytokines. Cytokine Growth Factor Rev. 2012, 23, 85–97. [Google Scholar] [CrossRef]
- Riedel, F.; Zaiss, I.; Herzog, D.; Götte, K.; Naim, R.; Hörmann, K. Serum Levels of Interleukin-6 in Patients with Primary Head and Neck Squamous Cell Carcinoma. Anticancer Res. 2005, 25, 2761–2765. [Google Scholar]
- Andersson, B.-Å.; Lewin, F.; Lundgren, J.; Nilsson, M.; Rutqvist, L.-E.; Löfgren, S.; Laytragoon-Lewin, N. Plasma Tumor Necrosis Factor-α and C-Reactive Protein as Biomarker for Survival in Head and Neck Squamous Cell Carcinoma. J. Cancer Res. Clin. Oncol. 2014, 140, 515–519. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.P.; Kao, H.K.; Wu, C.C.; Fang, K.H.; Chang, Y.L.; Huang, Y.C.; Liu, S.C.; Cheng, M.H. Pretreatment Interleukin-6 Serum Levels Are Associated with Patient Survival for Oral Cavity Squamous Cell Carcinoma. Otolaryngol.-Head Neck Surg. 2013, 148, 786–791. [Google Scholar] [CrossRef]
- Chen, C.C.; Chen, W.C.; Lu, C.H.; Wang, W.H.; Lin, P.Y.; der Lee, K.; Chen, M.F. Significance of Interleukin-6 Signaling in the Resistance of Pharyngeal Cancer to Irradiation and the Epidermal Growth Factor Receptor Inhibitor. Int. J. Radiat. Oncol. Biol. Phys. 2010, 76, 1214–1224. [Google Scholar] [CrossRef] [PubMed]
- Shinagawa, K.; Yanamoto, S.; Naruse, T.; Kawakita, A.; Morishita, K.; Sakamoto, Y.; Rokutanda, S.; Umeda, M. Clinical Roles of Interleukin-6 and STAT3 in Oral Squamous Cell Carcinoma. Pathol. Oncol. Res. 2017, 23, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yan, B.; van Waes, C. Role of the NF-ΚB Transcriptome and Proteome as Biomarkers Human Head and Neck Squamous Cell Carcinomas. Biomark. Med. 2008, 2, 409–429. [Google Scholar] [CrossRef]
- Hao, W.; Zhu, Y.; Zhou, H. Prognostic Value of Interleukin-6 and Interleukin-8 in Laryngeal Squamous Cell Cancer. Med. Oncol. 2013, 30, 333. [Google Scholar] [CrossRef]
- Chen, C.J.; Sung, W.W.; Lin, Y.M.; Chen, M.K.; Lee, C.H.; Lee, H.; Yeh, K.T.; Ko, J.L. Gender Difference in the Prognostic Role of Interleukin 6 in Oral Squamous Cell Carcinoma. PLoS ONE 2012, 7, e50104. [Google Scholar] [CrossRef]
- Schiegnitz, E.; Kämmerer, P.W.; Schön, H.; Blatt, S.; Berres, M.; Sagheb, K.; Al-Nawas, B. Proinflammatory Cytokines as Serum Biomarker in Oral Carcinoma—A Prospective Multi-Biomarker Approach. J. Oral Pathol. Med. 2018, 47, 268–274. [Google Scholar] [CrossRef]
- Li, C.; Zhao, Y.; Zhang, W.; Zhang, W. Increased Prevalence of TH17 Cells in the Peripheral Blood of Patients with Head and Neck Squamous Cell Carcinoma. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endodontol. 2011, 112, 81–89. [Google Scholar] [CrossRef]
- Sparano, A.; Lathers, D.M.; Achille, N.; Petruzzelli, G.J.; Young, M.R. Modulation of Th1 and Th2 Cytokine Profiles and Their Association with Advanced Head and Neck Squamous Cell Carcinoma. Otolaryngol.-Head Neck Surg. Off. J. Am. Acad. Otolaryngol.-Head Neck Surg. 2004, 131, 573–576. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.; Brown, N.V.; Swanson, B.J.; Schmitt, A.C.; Old, M.; Ozer, E.; Agrawal, A.; Schuller, D.E.; Teknos, T.N.; Kumar, P. High Expression of Myoferlin Is Associated with Poor Outcome in Oropharyngeal Squamous Cell Carcinoma Patients and Is Inversely Associated with HPV-Status. Oncotarget 2016, 7, 18665. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.; Yadav, A.; Brown, N.V.; Zhao, S.; Cipolla, M.J.; Wakely, P.E.; Schmitt, A.C.; Baiocchi, R.A.; Teknos, T.N.; Old, M.; et al. Nuclear PRMT5, Cyclin D1 and IL-6 Are Associated with Poor Outcome in Oropharyngeal Squamous Cell Carcinoma Patients and Is Inversely Associated with P16-Status. Oncotarget 2017, 8, 14847. [Google Scholar] [CrossRef] [PubMed]
- St. John, M.A.R.; Li, Y.; Zhou, X.; Denny, P.; Ho, C.M.; Montemagno, C.; Shi, W.; Qi, F.; Wu, B.; Sinha, U.; et al. Interleukin 6 and Interleukin 8 as Potential Biomarkers for Oral Cavity and Oropharyngeal Squamous Cell Carcinoma. Arch. Otolaryngol.-Head Neck Surg. 2004, 130, 929–935. [Google Scholar] [CrossRef]
- Duffy, S.A.; Teknos, T.; Taylor, J.M.G.; Fowler, K.E.; Islam, M.; Wolf, G.T.; McLean, S.; Ghanem, T.A.; Terrell, J.E. Health Behaviors Predict Higher Interleukin-6 Levels Among Patients Newly Diagnosed with Head and Neck Squamous Cell Carcinoma. Cancer Epidemiol. Biomark. Prev. A Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2013, 22, 374. [Google Scholar] [CrossRef]
- Meleti, M.; Cassi, D.; Vescovi, P.; Setti, G.; Pertinhez, T.A.; Pezzi, M.E. Salivary Biomarkers for Diagnosis of Systemic Diseases and Malignant Tumors. A Systematic Review. Med. Oral Patol. Oral Cir. Bucal 2020, 25, e299. [Google Scholar] [CrossRef] [PubMed]
- Setti, G.; Pezzi, M.E.; Viani, M.V.; Pertinhez, T.A.; Cassi, D.; Magnoni, C.; Bellini, P.; Musolino, A.; Vescovi, P.; Meleti, M. Salivary MicroRNA for Diagnosis of Cancer and Systemic Diseases: A Systematic Review. Int. J. Mol. Sci. 2020, 21, 907. [Google Scholar] [CrossRef]
- Meleti, M.; Quartieri, E.; Antonelli, R.; Pezzi, M.E.; Ghezzi, B.; Viani, M.V.; Setti, G.; Casali, E.; Ferrari, E.; Ciociola, T.; et al. Metabolic Profiles of Whole, Parotid and Submandibular/Sublingual Saliva. Metabolites 2020, 10, 318. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, E.; Pezzi, M.E.; Cassi, D.; Pertinhez, T.A.; Spisni, A.; Meleti, M. Salivary Cytokines as Biomarkers for Oral Squamous Cell Carcinoma: A Systematic Review. Int. J. Mol. Sci. 2021, 22, 6795. [Google Scholar] [CrossRef]
- Cristaldi, M.; Mauceri, R.; di Fede, O.; Giuliana, G.; Campisi, G.; Panzarella, V. Salivary Biomarkers for Oral Squamous Cell Carcinoma Diagnosis and Follow-Up: Current Status and Perspectives. Front. Physiol. 2019, 10, 1476. [Google Scholar] [CrossRef] [PubMed]
- Dineshkumar, T.; Ashwini, B.K.; Rameshkumar, A.; Rajashree, P.; Ramya, R.; Rajkumar, K. Salivary and Serum Interleukin-6 Levels in Oral Premalignant Disorders and Squamous Cell Carcinoma: Diagnostic Value and Clinicopathologic Correlations. Asian Pac. J. Cancer Prev. APJCP 2016, 17, 4899. [Google Scholar] [CrossRef]
- Katakura, A.; Kamiyama, I.; Takano, N.; Shibahara, T.; Muramatsu, T.; Ishihara, K.; Takagi, R.; Shouno, T. Comparison of Salivary Cytokine Levels in Oral Cancer Patients and Healthy Subjects. Bull. Tokyo Dent. Coll. 2007, 48, 199–203. [Google Scholar] [CrossRef]
- Rhodus, N.L.; Ho, V.; Miller, C.S.; Myers, S.; Ondrey, F. NF-ΚB Dependent Cytokine Levels in Saliva of Patients with Oral Preneoplastic Lesions and Oral Squamous Cell Carcinoma. Cancer Detect. Prev. 2005, 29, 42–45. [Google Scholar] [CrossRef]
- Juretić, M.; Cerović, R.; Belušić-Gobić, M.; Brekalo Pršo, I.; Kqiku, L.; Špalj, S.; Pezelj-Ribarić, S. Salivary Levels of TNF-α and IL-6 in Patients with Oral Premalignant and Malignant Lesions. Folia Biol. 2014, 59, 99–102. [Google Scholar]
- Korostoff, A.; Reder, L.; Masood, R.; Sinha, U.K. The Role of Salivary Cytokine Biomarkers in Tongue Cancer Invasion and Mortality. Oral Oncol. 2011, 47, 282–287. [Google Scholar] [CrossRef]
- Hamad, A.; Gaphor, S.; Shawagfeh, M.T.; Al-Talabani, N. Study of Serum and Salivary Levels of Proinflammatory Cytokines, Potential Biomarkers in the Diagnosis of Oral Squamous Cell Carcinoma. Acad. J. Cancer Res. 2011, 4, 47–55. [Google Scholar]
- Sahebjamee, M.; Eslami, M.; Atarbashimoghadam, F.; Sarafnejad, A. Salivary Concentration of TNFα, IL1α, IL6, and IL8 in Oral Squamous Cell Carcinoma. Med. Oral Patol. Oral Cir. Bucal 2008, 13, E292–E295. [Google Scholar] [PubMed]
- Cheng, Y.-S.L.; Jordan, L.; Gorugantula, L.M.; Schneiderman, E.; Chen, H.-S.; Rees, T. Salivary Interleukin-6 and -8 in Patients With Oral Cancer and Patients With Chronic Oral Inflammatory Diseases. J. Periodontol. 2014, 85, 956–965. [Google Scholar] [CrossRef] [PubMed]
- Selvam, N.P.; Sadaksharam, J. Salivary Interleukin-6 in the Detection of Oral Cancer and Precancer. Asia-Pac. J. Clin. Oncol. 2015, 11, 236–241. [Google Scholar] [CrossRef]
- Dikova, V.; Jantus-Lewintre, E.; Bagan, J. Potential Non-Invasive Biomarkers for Early Diagnosis of Oral Squamous Cell Carcinoma. J. Clin. Med. 2021, 10, 1658. [Google Scholar] [CrossRef] [PubMed]
- Van der Waal, I. Potentially Malignant Disorders of the Oral and Oropharyngeal Mucosa; Terminology, Classification and Present Concepts of Management. Oral Oncol. 2009, 45, 317–323. [Google Scholar] [CrossRef]
- Sato, J.; Goto, J.; Murata, T.; Kitamori, S.; Yamazaki, Y.; Satoh, A.; Kitagawa, Y. Changes in Saliva Interleukin-6 Levels in Patients with Oral Squamous Cell Carcinoma. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2010, 110, 330–336. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sato, J.; Ohuchi, M.; Abe, K.; Satoh, T.; Abe, T.; Yamazaki, Y.; Satoh, A.; Notani, K.; Kitagawa, Y. Correlation between Salivary Interleukin-6 Levels and Early Locoregional Recurrence in Patients with Oral Squamous Cell Carcinoma: Preliminary Study. Head Neck 2013, 35, 889–894. [Google Scholar] [CrossRef]
- Sato, J.; Ohuchi, M.; Wada, M.; Ohga, N.; Asaka, T.; Yoshikawa, K.; Miyakoshi, M.; Hata, H.; Satoh, A.; Kitagawa, Y. Differences in Sequential Posttreatment Salivary IL-6 Levels between Patients with and Patients without Locoregional Recurrences of Oral Squamous Cell Carcinoma: Part III of a Cohort Study. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2015, 120, 751–760.e2. [Google Scholar] [CrossRef]
- Fan, N.; Luo, Y.; Ou, Y.; He, H. Altered Serum Levels of TNF-α, IL-6, and IL-18 in Depressive Disorder Patients. Hum. Psychopharmacol. 2017, 32, e2588. [Google Scholar] [CrossRef]
- Patel, H.J.; Patel, B.M. TNF-α and Cancer Cachexia: Molecular Insights and Clinical Implications. Life Sci. 2017, 170, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Smetana, K., Jr.; Dvořánková, B.; Lacina, L.; Strnad, H.; Kolář, M.; Chovanec, M.; Plzák, J.; Čada, Z.; Vlček, Č.; Szabo, P.; et al. Combination of Antibodies or Fab Fragments Thereof for Use as Medicament, and Pharmaceutical Composition Containing Said Antibodies or Fab Fragments Thereof. Czech Patent No. B6 303227, 6 June 2012. (In Czech). [Google Scholar]
- Jayatilaka, H.; Tyle, P.; Chen, J.J.; Kwak, M.; Ju, J.; Kim, H.J.; Lee, J.S.H.; Wu, P.H.; Gilkes, D.M.; Fan, R.; et al. Synergistic IL-6 and IL-8 Paracrine Signalling Pathway Infers a Strategy to Inhibit Tumour Cell Migration. Nat. Commun. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wirtz, D.G. and Yatilaka H. Cancer Cell Migration Inhibitors and Their Use in Therapeutic Treatments. U.S. Patent 2017/0165363 A1, 15 June 2017. [Google Scholar]
- Kodet, O.; Dvořánková, B.; Bendlová, B.; Sýkorová, V.; Krajsová, I.; Štork, J.; Kučera, J.; Szabo, P.; Strnad, H.; Kolář, M.; et al. Microenvironment-Driven Resistance to B-Raf Inhibition in a Melanoma Patient Is Accompanied by Broad Changes of Gene Methylation and Expression in Distal Fibroblasts. Int. J. Mol. Med. 2018, 41, 2687–2703. [Google Scholar] [CrossRef] [PubMed]
- Loppnow, H.; Zhang, L.; Buerke, M.; Lautenschläger, M.; Chen, L.; Frister, A.; Schlitt, A.; Luther, T.; Song, N.; Hofmann, B.; et al. Statins Potently Reduce the Cytokine-Mediated IL-6 Release in SMC/MNC Cocultures. J. Cell. Mol. Med. 2011, 15, 994. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, S.; Wagner, T.; Venkatakrishnan, A.J.; Puranik, A.; Hurchik, M.; Agarwal, V.; Conrad, I.; Kirkup, C.; Arunachalam, R.; O’Horo, J.; et al. Plasma IL-6 Levels Following Corticosteroid Therapy as an Indicator of ICU Length of Stay in Critically Ill COVID-19 Patients. Cell Death Discov. 2021, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.K.; Ridker, P.M. Anti-Inflammatory Effects of Statins: Clinical Evidence and Basic Mechanisms. Nat. Rev. Drug Discov. 2005, 4, 977–987. [Google Scholar] [CrossRef]
- Smetana, K.; Smetana, K.; Brábek, J.; Brábek, J. Role of Interleukin-6 in Lung Complications in Patients with COVID-19: Therapeutic Implications. In Vivo 2020, 34, 1589–1592. [Google Scholar] [CrossRef]
- Atal, S.; Fatima, Z. IL-6 Inhibitors in the Treatment of Serious COVID-19: A Promising Therapy? Pharm. Med. 2020, 34, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Gould, S.; Norris, S.L. Contested Effects and Chaotic Policies: The 2020 Story of (Hydroxy) Chloroquine for Treating COVID-19. Cochrane Database Syst. Rev. 2021, 3, ED000151. [Google Scholar] [CrossRef]
- Wozniacka, A.; Lesiak, A.; Narbutt, J.; McCauliffe, D.P.; Sysa-Jedrzejowska, A. Chloroquine Treatment Influences Proinflammatory Cytokine Levels in Systemic Lupus Erythematosus Patients. Lupus 2006, 15, 268–275. [Google Scholar] [CrossRef]
- Van den Borne, B.E.; Dijkmans, B.A.; de Rooij, H.H.; le Cessie, S.; Verweij, C.L. Chloroquine and Hydroxychloroquine Equally Affect Tumor Necrosis Factor-Alpha, Interleukin 6, and Interferon-Gamma Production by Peripheral Blood Mononuclear Cells. J. Rheumatol. 1997, 24, 55–60. [Google Scholar]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-Related Inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef]
- Varisli, L.; Cen, O.; Vlahopoulos, S. Dissecting Pharmacological Effects of Chloroquine in Cancer Treatment: Interference with Inflammatory Signaling Pathways. Immunology 2020, 159, 257–278. [Google Scholar] [CrossRef]
- Bryant, J.; Batis, N.; Franke, A.C.; Clancey, G.; Hartley, M.; Ryan, G.; Brooks, J.; Southam, A.D.; Barnes, N.; Parish, J.; et al. Repurposed Quinacrine Synergizes with Cisplatin, Reducing the Effective Dose Required for Treatment of Head and Neck Squamous Cell Carcinoma. Oncotarget 2019, 10, 5229–5244. [Google Scholar] [CrossRef][Green Version]
- Duarte, D.; Vale, N. New Trends for Antimalarial Drugs: Synergism between Antineoplastics and Antimalarials on Breast Cancer Cells. Biomolecules 2020, 10, 1623. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Cánoves, P.; Scheele, C.; Pedersen, B.K.; Serrano, A.L. Interleukin-6 Myokine Signaling in Skeletal Muscle: A Double-edged Sword? FEBS J. 2013, 280, 4131. [Google Scholar] [CrossRef]
- Kawano, M.; Hirano, T.; Matsuda, T.; Taga, T.; Horii, Y.; Iwato, K.; Asaoku, H.; Tang, B.; Tanabe, O.; Tanaka, H.; et al. Autocrine Generation and Requirement of BSF-2/IL-6 for Human Multiple Myelomas. Nature 1988, 332, 83–85. [Google Scholar] [CrossRef] [PubMed]
- Beck, J.T.; Hsu, S.-M.; Wijdenes, J.; Bataille, R.; Klein, B.; Vesole, D.; Hayden, K.; Jagannath, S.; Barlogie, B. Alleviation of Systemic Manifestations of Castleman’s Disease by Monoclonal Anti-Interleukin-6 Antibody. N. Engl. J. Med. 2010, 330, 602–605. [Google Scholar] [CrossRef]
- Van Rhee, F.; Fayad, L.; Voorhees, P.; Furman, R.; Lonial, S.; Borghaei, H.; Sokol, L.; Crawford, J.; Cornfeld, M.; Qi, M.; et al. Siltuximab, a Novel Anti-Interleukin-6 Monoclonal Antibody, for Castleman’s Disease. J. Clin. Oncol. 2010, 28. [Google Scholar] [CrossRef] [PubMed]
- Choy, E.H.; de Benedetti, F.; Takeuchi, T.; Hashizume, M.; John, M.R.; Kishimoto, T. Translating IL-6 Biology into Effective Treatments. Nat. Rev. Rheumatol. 2020, 16, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Doberer, K.; Duerr, M.; Halloran, P.F.; Eskandary, F.; Budde, K.; Regele, H.; Reeve, J.; Borski, A.; Kozakowski, N.; Reindl-Schwaighofer, R.; et al. A Randomized Clinical Trial of Anti⇓IL-6 Antibody Clazakizumab in Late Antibody-Mediated Kidney Transplant Rejection. J. Am. Soc. Nephrol. 2021, 32, 708–722. [Google Scholar] [CrossRef] [PubMed]
- Shaw, S.; Bourne, T.; Meier, C.; Carrington, B.; Gelinas, R.; Henry, A.; Popplewell, A.; Adams, R.; Baker, T.; Rapecki, S.; et al. Discovery and Characterization of Olokizumab: A Humanized Antibody Targeting Interleukin-6 and Neutralizing Gp130-Signaling. mAbs 2014, 6, 773–781. [Google Scholar] [CrossRef]
- Burger, R.; Günther, A.; Klausz, K.; Staudinger, M.; Peipp, M.; Penas, E.M.M.; Rose-John, S.; Wijdenes, J.; Gramatzki, M. Due to Interleukin-6 Type Cytokine Redundancy Only Glycoprotein 130 Receptor Blockade Efficiently Inhibits Myeloma Growth. Haematologica 2017, 102, 381. [Google Scholar] [CrossRef]
- Nishimoto, N.; Kishimoto, T. Humanized Antihuman IL-6 Receptor Antibody, Tocilizumab. Handb. Exp. Pharmacol. 2008, 181, 151–160. [Google Scholar] [CrossRef]
- Genovese, M.C.; McKay, J.D.; Nasonov, E.L.; Mysler, E.F.; da Silva, N.A.; Alecock, E.; Woodworth, T.; Gomez-Reino, J.J. Interleukin-6 Receptor Inhibition with Tocilizumab Reduces Disease Activity in Rheumatoid Arthritis with Inadequate Response to Disease-Modifying Antirheumatic Drugs: The Tocilizumab in Combination with Traditional Disease-Modifying Antirheumatic Drug Therapy Study. Arthritis Rheum. 2008, 58, 2968–2980. [Google Scholar] [CrossRef]
- Kim, N.; Kim, S.; Kim, D.; Zhang, D.; Park, J.; Yi, H.; Kim, J.; Shin, H. Anti-proliferative Action of IL-6R-Targeted Antibody Tocilizumab for Non-Small Cell Lung Cancer Cells. Oncol. Lett. 2015, 9, 2283–2288. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Alraouji, N.N.; Al-Mohanna, F.H.; Ghebeh, H.; Arafah, M.; Almeer, R.; Al-Tweigeri, T.; Aboussekhra, A. Tocilizumab Potentiates Cisplatin Cytotoxicity and Targets Cancer Stem Cells in Triple-Negative Breast Cancer. Mol. Carcinog. 2020, 59, 1041–1051. [Google Scholar] [CrossRef] [PubMed]
- Varra, V.; Smile, T.D.; Geiger, J.L.; Koyfman, S.A. Recent and Emerging Therapies for Cutaneous Squamous Cell Carcinomas of the Head and Neck. Curr. Treat. Options Oncol. 2020, 21, 1–12. [Google Scholar] [CrossRef]
- Migden, M.R.; Rischin, D.; Schmults, C.D.; Guminski, A.; Hauschild, A.; Lewis, K.D.; Chung, C.H.; Hernandez-Aya, L.; Lim, A.M.; Chang, A.L.S.; et al. PD-1 Blockade with Cemiplimab in Advanced Cutaneous Squamous-Cell Carcinoma. N. Engl. J. Med. 2018, 379, 341–351. [Google Scholar] [CrossRef]
- Tsukamoto, H.; Fujieda, K.; Miyashita, A.; Fukushima, S.; Ikeda, T.; Kubo, Y.; Senju, S.; Ihn, H.; Nishimura, Y.; Oshiumi, H. Combined Blockade of IL6 and PD-1/PD-L1 Signaling Abrogates Mutual Regulation of Their Immunosuppressive Effects in the Tumor Microenvironment. Cancer Res. 2018, 78, 5011–5022. [Google Scholar] [CrossRef]
- Boyce, E.G.; Rogan, E.L.; Vyas, D.; Prasad, N.; Mai, Y. Sarilumab: Review of a Second IL-6 Receptor Antagonist Indicated for the Treatment of Rheumatoid Arthritis. Ann. Pharmacother. 2018, 52, 780–791. [Google Scholar] [CrossRef]
- Heo, Y.A. Satralizumab: First Approval. Drugs 2020, 80, 1477–1482. [Google Scholar] [CrossRef] [PubMed]
- Glicklich, A.; Grayson, P.; Blanchetot, C.; Zhou, Q.; Kretz-Rommel, A. The Development of a New Anti–Interleukin 6 Blocker for Rheumatoid Arthritis Patients-ACR Meeting Abstracts. In Arthritis & Rheumatology; Wiley: Hoboken, NJ, USA, 2016; Volume 68. [Google Scholar]
- Rinaldi, M.; van Bogaert, T.; van Roy, M.; Bontinck, L.; Hohlbaum, A.; Snoeck, V.; Dombrecht, E.; van Beneden, K.; Schoen, P.; Ulrichts, H. Assessment of Dose Dependent Effects of Vobarilizumab, an Anti-IL6 Receptor (IL-6R) Nanobody®, on Systemic Biomarkers in Patients with Moderate-to-Severe Rheumatoid Arthritis (RA): Results of Two Phase 2b Studies-ACR Meeting Abstracts. Arthritis Rheumatol. 2017, 69, 2476. [Google Scholar]
- Chevalier, S.; Fourcin, M.; Robledo, O.; Wijdenes, J.; Pouplard-Barthelaix, A.; Gascan, H. Interleukin-6 Family of Cytokines Induced Activation of Different Functional Sites Expressed by Gp130 Transducing Protein. J. Biol. Chem. 1996, 271. [Google Scholar] [CrossRef]
- Yamamoto, D.; Sunazuka, T.; Hirose, T.; Kojima, N.; Kaji, E.; Omura, S. Design, Synthesis, and Biological Activities of Madindoline Analogues. Bioorg. Med. Chem. Lett. 2006, 16, 2807–2811. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Grande, F.; Garofalo, A.; Neamati, N. Discovery of a Novel Orally Active Small-Molecule Gp130 Inhibitor for the Treatment of Ovarian Cancer. Mol. Cancer Ther. 2013, 12, 937–949. [Google Scholar] [CrossRef]
- Hong, S.-S.; Choi, J.H.; Lee, S.Y.; Park, Y.-H.; Park, K.-Y.; Lee, J.Y.; Kim, J.; Gajulapati, V.; Goo, J.-I.; Singh, S.; et al. A Novel Small-Molecule Inhibitor Targeting the IL-6 Receptor β Subunit, Glycoprotein 130. J. Immunol. 2015, 195, 237–245. [Google Scholar] [CrossRef]
- Li, H.; Xiao, H.; Lin, L.; Jou, D.; Kumari, V.; Lin, J.; Li, C. Drug Design Targeting Protein-Protein Interactions (PPIs) Using Multiple Ligand Simultaneous Docking (MLSD) and Drug Repositioning: Discovery of Raloxifene and Bazedoxifene as Novel Inhibitors of IL-6/GP130 Interface. J. Med. Chem. 2014, 57, 632–641. [Google Scholar] [CrossRef]
- Thilakasiri, P.; Huynh, J.; Poh, A.R.; Tan, C.W.; Nero, T.L.; Tran, K.; Parslow, A.C.; Afshar-Sterle, S.; Baloyan, D.; Hannan, N.J.; et al. Repurposing the Selective Estrogen Receptor Modulator Bazedoxifene to Suppress Gastrointestinal Cancer Growth. EMBO Mol. Med. 2019, 11, e9539. [Google Scholar] [CrossRef]
- Heise, D.; Derrac Soria, A.; Hansen, S.; Dambietz, C.; Akbarzadeh, M.; Berg, A.F.; Waetzig, G.H.; Jones, S.A.; Dvorsky, R.; Ahmadian, M.R.; et al. Selective Inhibition of IL-6 Trans-Signaling by a Miniaturized, Optimized Chimeric Soluble Gp130 Inhibits T H 17 Cell Expansion. Sci. Signal. 2021, 14, eabc3480. [Google Scholar] [CrossRef]
- Schreiber, S.; Aden, K.; Bernardes, J.P.; Conrad, C.; Tran, F.; Höper, H.; Volk, V.; Mishra, N.; Blase, J.I.; Nikolaus, S.; et al. Therapeutic Interleukin-6 Trans-Signaling Inhibition by Olamkicept (Sgp130Fc) in Patients With Active Inflammatory Bowel Disease. Gastroenterology 2021, 160, 2354–2366.e11. [Google Scholar] [CrossRef]
- Uz, U.; Eskiizmir, G. Association between Interleukin-6 and Head and Neck Squamous Cell Carcinoma: A Systematic Review. Clin. Exp. Otorhinolaryngol. 2021, 14, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Cui, N.; Nomura, T.; Noma, H.; Yokoo, K.; Takagi, R.; Hashimoto, S.; Okamoto, M.; Sato, M.; Yu, G.; Guo, C.; et al. Effect of YM529 on a Model of Mandibular Invasion by Oral Squamous Cell Carcinoma in Mice. Clin. Cancer Res. 2005, 11, 2713–2719. [Google Scholar] [CrossRef] [PubMed]
- Wichmann, G.; Cedra, S.; Schlegel, D.; Kolb, M.; Wiegand, S.; Boehm, A.; Hofer, M.; Dietz, A. Cilengitide and Cetuximab Reduce Cytokine Production and Colony Formation of Head and Neck Squamous Cell Carcinoma Cells Ex Vivo. Anticancer Res. 2017, 37, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Wolf, J.S.; Li, G.; Varadhachary, A.; Petrak, K.; Schneyer, M.; Li, D.; Ongkasuwan, J.; Zhang, X.; Taylor, R.J.; Strome, S.E.; et al. Oral Lactoferrin Results in T Cell-Dependent Tumor Inhibition of Head and Neck Squamous Cell Carcinoma in Vivo. Clin. Cancer Res. 2007, 13, 1601–1610. [Google Scholar] [CrossRef] [PubMed]
- Tu, D.G.; Lin, W.T.; Yu, C.C.; Lee, S.S.; Peng, C.Y.; Lin, T.; Yu, C.H. Chemotherapeutic Effects of Luteolin on Radio-Sensitivity Enhancement and Interleukin-6/Signal Transducer and Activator of Transcription 3 Signaling Repression of Oral Cancer Stem Cells. J. Formos. Med. Assoc. 2016, 115, 1032–1038. [Google Scholar] [CrossRef] [PubMed]
- Tamatani, T.; Azuma, M.; Motegi, K.; Takamaru, N.; Kawashima, Y.; Bando, T. Cepharanthin-Enhanced Radiosensitivity through the Inhibition of Radiation-Induced Nuclear Factor-ΚB Activity in Human Oral Squamous Cell Carcinoma Cells. Int. J. Oncol. 2007, 31, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Macha, M.A.; Matta, A.; Chauhan, S.S.; Michael Siu, K.W.; Ralhan, R. Guggulsterone (GS) Inhibits Smokeless Tobacco and Nicotine-Induced NF-ΚB and STAT3 Pathways in Head and Neck Cancer Cells. Carcinogenesis 2011, 32, 368–380. [Google Scholar] [CrossRef]
- Lin, H.Y.; Hou, S.C.; Chen, S.C.; Kao, M.C.; Yu, C.C.; Funayama, S.; Ho, C.T.; der Way, T. (-)-Epigallocatechin Gallate Induces Fas/CD95-Mediated Apoptosis through Inhibiting Constitutive and IL-6-Induced JAK/STAT3 Signaling in Head and Neck Squamous Cell Carcinoma Cells. J. Agric. Food Chem. 2012, 60, 2480–2489. [Google Scholar] [CrossRef]
- Cohen, A.N.; Veena, M.S.; Srivatsan, E.S.; Wang, M.B. Suppression of Interleukin 6 and 8 Production in Head and Neck Cancer Cells with Curcumin via Inhibition of Iκβ Kinase. Arch. Otolaryngol.-Head Neck Surg. 2009, 135, 190–197. [Google Scholar] [CrossRef]
- Chakravarti, N.; Myers, J.N.; Aggarwal, B.B. Targeting Constitutive and Interleukin-6-Inducible Signal Transducers and Activators of Transcription 3 Pathway in Head and Neck Squamous Cell Carcinoma Cells by Curcumin (Diferuloylmethane). Int. J. Cancer 2006, 119, 1268–1275. [Google Scholar] [CrossRef]
- DiNatale, B.C.; Schroeder, J.C.; Perdew, G.H. Ah Receptor Antagonism Inhibits Constitutive and Cytokine Inducible IL6 Production in Head and Neck Tumor Cell Lines. Mol. Carcinog. 2011, 50, 173–183. [Google Scholar] [CrossRef] [PubMed]
- DiNatale, B.C.; Smith, K.; John, K.; Krishnegowda, G.; Amin, S.G.; Perdew, G.H. Ah Receptor Antagonism Represses Head and Neck Tumor Cell Aggressive Phenotype. Mol. Cancer Res. 2012, 10, 1369–1379. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.H.; Chuang, J.Y.; Fong, Y.C.; Maa, M.C.; der Way, T.; Hung, C.H. Bone-Derived SDF-1 Stimulates IL-6 Release via CXCR4, ERK and NF-ΚB Pathways and Promotes Osteoclastogenesis in Human Oral Cancer Cells. Carcinogenesis 2008, 29, 1483–1492. [Google Scholar] [CrossRef] [PubMed]
- Siavash, H.; Nikitakis, N.G.; Sauk, J.J. Abrogation of IL-6-Mediated JAK Signalling by the Cyclopentenone Prostaglandin 15d-PGJ2 in Oral Squamous Carcinoma Cells. Br. J. Cancer 2004, 91, 1074–1080. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Poth, K.J.; Guminski, A.D.; Thomas, G.P.; Leo, P.J.; Jabbar, I.A.; Saunders, N.A. Cisplatin Treatment Induces a Transient Increase in Tumorigenic Potential Associated with High Interleukin-6 Expression in Head and Neck Squamous Cell Carcinoma. Mol. Cancer Ther. 2010, 9, 2430–2439. [Google Scholar] [CrossRef] [PubMed]
- Stanam, A.; Love-Homan, L.; Joseph, T.S.; Espinosa-Cotton, M.; Simons, A.L. Upregulated Interleukin-6 Expression Contributes to Erlotinib Resistance in Head and Neck Squamous Cell Carcinoma. Mol. Oncol. 2015, 9, 1371–1383. [Google Scholar] [CrossRef]
- Shinriki, S.; Jono, H.; Ueda, M.; Ota, K.; Ota, T.; Sueyoshi, T.; Oike, Y.; Ibusuki, M.; Hiraki, A.; Nakayama, H.; et al. Interleukin-6 Signalling Regulates Vascular Endothelial Growth Factor-C Synthesis and Lymphangiogenesis in Human Oral Squamous Cell Carcinoma. J. Pathol. 2011, 225, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Shinriki, S.; Jono, H.; Ota, K.; Ueda, M.; Kudo, M.; Ota, T.; Oike, Y.; Endo, M.; Ibusuki, M.; Hiraki, A.; et al. Humanized Anti-Interleukin-6 Receptor Antibody Suppresses Tumor Angiogenesis and in Vivo Growth of Human Oral Squamous Cell Carcinoma. Clin. Cancer Res. 2009, 15, 5426–5434. [Google Scholar] [CrossRef]
- Islam, M.; Sharma, S.; Teknos, T.N. RhoC Regulates Cancer Stem Cells in Head and Neck Squamous Cell Carcinoma by Overexpressing IL-6 and Phosphorylation of STAT3. PLoS ONE 2014, 9, 88527. [Google Scholar] [CrossRef]
- Hwang, Y.S.; Ahn, S.Y.; Moon, S.; Zheng, Z.; Cha, I.H.; Kim, J.; Zhang, X. Insulin-like Growth Factor-II MRNA Binding Protein-3 and Podoplanin Expression Are Associated with Bone Invasion and Prognosis in Oral Squamous Cell Carcinoma. Arch. Oral Biol. 2016, 69, 25–32. [Google Scholar] [CrossRef]
- Finkel, K.A.; Warner, K.A.; Kerk, S.; Bradford, C.R.; McLean, S.A.; Prince, M.E.; Zhong, H.; Hurt, E.M.; Hollingsworth, R.E.; Wicha, M.S.; et al. IL-6 Inhibition With MEDI5117 Decreases The Fraction of Head and Neck Cancer Stem Cells and Prevents Tumor Recurrence. Neoplasia 2016, 18, 273–281. [Google Scholar] [CrossRef]
- Ekshyyan, O.; Khandelwal, A.R.; Rong, X.; Moore-Medlin, T.; Ma, X.; Alexander, J.S.; Nathan, C.A.O. Rapamycin Targets Interleukin 6 (IL-6) Expression and Suppresses Endothelial Cell Invasion Stimulated by Tumor Cells. Am. J. Transl. Res. 2016, 8, 4822–4830. [Google Scholar]
- Riebe, C.; Pries, R.; Kemkers, A.; Wollenberg, B. Increased Cytokine Secretion in Head and Neck Cancer upon P38 Mitogen-Activated Protein Kinase Activation. Int. J. Mol. Med. 2007, 20, 883–887. [Google Scholar] [CrossRef][Green Version]
- Jing, Z.; Xu, H.; Chen, X.; Zhong, Q.; Huang, J.; Zhang, Y.; Guo, W.; Yang, Z.; Ding, S.; Chen, P.; et al. The Proton-Sensing G-Protein Coupled Receptor GPR4 Promotes Angiogenesis in Head and Neck Cancer. PLoS ONE 2016, 11, e0152789. [Google Scholar] [CrossRef] [PubMed]
- Kross, K.W.; Heimdal, J.H.; Olsnes, C.; Olofson, J.; Aarstad, H.J. Tumour-Associated Macrophages Secrete IL-6 and MCP-1 in Head and Neck Squamous Cell Carcinoma Tissue. Acta Oto-Laryngol. 2007, 127, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.B.; Lobo, A.S.; Joshi, K.S.; Rathos, M.J.; Kumar, G.A.; Padigaru, M. Molecular Mechanisms of Anti-Tumor Properties of P276-00 in Head and Neck Squamous Cell Carcinoma. J. Transl. Med. 2013, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sen, M.; Johnston, P.A.; Pollock, N.I.; DeGrave, K.; Joyce, S.C.; Freilino, M.L.; Hua, Y.; Camarco, D.P.; Close, D.A.; Huryn, D.M.; et al. Mechanism of Action of Selective Inhibitors of IL-6 Induced STAT3 Pathway in Head and Neck Cancer Cell Lines. J. Chem. Biol. 2017, 10, 129–141. [Google Scholar] [CrossRef]
- Teknos, T.N.; Islam, M.; Arenberg, D.A.; Pan, Q.; Carskadon, S.L.; Abarbanell, A.M.; Marcus, B.; Paul, S.; Vandenberg, C.D.; Carron, M.; et al. The Effect of Tetrathiomolybdate on Cytokine Expression, Angiogenesis, and Tumor Growth in Squamous Cell Carcinoma of the Head and Neck. Arch. Otolaryngol.-Head Neck Surg. 2005, 131, 204–211. [Google Scholar] [CrossRef]
- Van Tubergen, E.; vander Broek, R.; Lee, J.; Wolf, G.; Carey, T.; Bradford, C.; Prince, M.; Kirkwood, K.L.; D’Silva, N.J. Tristetraprolin Regulates Interleukin-6, Which Is Correlated with Tumor Progression in Patients with Head and Neck Squamous Cell Carcinoma. Cancer 2011, 117, 2677–2689. [Google Scholar] [CrossRef]
- Van Tubergen, E.A.; Banerjee, R.; Liu, M.; vander Broek, R.; Light, E.; Kuo, S.; Feinberg, S.E.; Willis, A.L.; Wolf, G.; Carey, T.; et al. Inactivation or Loss of TTP Promotes Invasion in Head and Neck Cancer via Transcript Stabilization and Secretion of MMP9, MMP2, and IL-6. Clin. Cancer Res. 2013, 19, 1169–1179. [Google Scholar] [CrossRef]
- Zhou, X.; Ren, Y.; Liu, A.; Han, L.; Zhang, K.; Li, S.; Li, P.; Li, P.; Kang, C.; Wang, X.; et al. STAT3 Inhibitor WP1066 Attenuates MiRNA-21 to Suppress Human Oral Squamous Cell Carcinoma Growth in Vitro and in Vivo. Oncol. Rep. 2014, 31, 2173–2180. [Google Scholar] [CrossRef]
- Te Chang, M.; Lee, S.P.; Fang, C.Y.; Hsieh, P.L.; Liao, Y.W.; Lu, M.Y.; Tsai, L.L.; Yu, C.C.; Liu, C.M. Chemosensitizing Effect of Honokiol in Oral Carcinoma Stem Cells via Regulation of IL-6/Stat3 Signaling. Environ. Toxicol. 2018, 33, 1105–1112. [Google Scholar] [CrossRef]
- Wang, T.H.; Fang, J.Y.; Wu, S.J.; Liu, Y.W.; Chan, C.W.; Chuang, S.Y.; Chen, C.Y. 2-O-Methylmagnolol Induces Apoptosis and Inhibits IL-6/STAT3 Signaling in Oral Squamous Cell Carcinoma. Cell. Physiol. Biochem. 2018, 50, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Gehrke, T.; Scherzad, A.; Hackenberg, S.; Ickrath, P.; Schendzielorz, P.; Hagen, R.; Kleinsasser, N. Additive Antitumor Effects of Celecoxib and Simvastatin on Head and Neck Squamous Cell Carcinoma in Vitro. Int. J. Oncol. 2017, 51, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.; Kumar, B.; Teknos, T.N.; Kumar, P. Bazedoxifene Enhances the Anti-Tumor Effects of Cisplatin and Radiation Treatment by Blocking IL-6 Signaling in Head and Neck Cancer. Oncotarget 2017, 8, 66912. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.; Pries, R.; Wollenberg, B. Established and Novel NF-ΚB Inhibitors Lead to Downregulation of TLR3 and the Proliferation and Cytokine Secretion in HNSCC. Oral Oncol. 2011, 47, 818–826. [Google Scholar] [CrossRef] [PubMed]
- Ruan, M.; Thorn, K.; Liu, S.; Li, S.; Yang, W.; Zhang, C.; Zhang, C. The Secretion of IL-6 by CpG-ODN-Treated Cancer Cells Promotes T-Cell Immune Responses Partly through the TLR-9/AP-1 Pathway in Oral Squamous Cell Carcinoma. Int. J. Oncol. 2014, 45, 2103–2110. [Google Scholar] [CrossRef] [PubMed]
- A First-in-Humans Dose Finding Study for an Aryl Hydrocarbon Receptor Inhibitor (AhRi) in Patients With Advanced Cancer-Full Text View-ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/show/study/NCT04069026?term=IL-6&cond=HNSCC&draw=2&rank=9 (accessed on 12 September 2021).
- Zafar, E.; Maqbool, M.F.; Iqbal, A.; Maryam, A.; Shakir, H.A.; Irfan, M.; Khan, M.; Li, Y.; Ma, T. A Comprehensive Review on Anticancer Mechanism of Bazedoxifene. Biotechnol. Appl. Biochem. 2021. [Google Scholar] [CrossRef] [PubMed]
- Erbitux (Cetuximab)-Summary of Product Characteristics. Available online: https://www.ema.europa.eu/en/documents/product-information/erbitux-epar-product-information_en.pdf (accessed on 10 October 2021).
- The Effect of Curcumin for Treatment of Cancer Anorexia-Cachexia Syndrome in Patients with Stage III-IV of Head and Neck Cancer-Full Text View-ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/show/NCT04208334?cond=head+and+neck+cancer+curcumin&draw=2&rank=2 (accessed on 12 September 2021).
- Kotha, R.R.; Luthria, D.L. Curcumin: Biological, Pharmaceutical, Nutraceutical, and Analytical Aspects. Molecules 2019, 24, 2930. [Google Scholar] [CrossRef]
- Celebrex (Celecoxib) Information | FDA. Available online: https://www.fda.gov/drugs/postmarket-drug-safety-information-patients-and-providers/celebrex-celecoxib-information (accessed on 12 September 2021).
- Simvastatin Information | FDA. Available online: https://www.fda.gov/drugs/postmarket-drug-safety-information-patients-and-providers/simvastatin-information (accessed on 12 September 2021).
- Du, G.-J.; Zhang, Z.; Wen, X.-D.; Yu, C.; Calway, T.; Yuan, C.-S.; Wang, C.-Z. Epigallocatechin Gallate (EGCG) Is the Most Effective Cancer Chemopreventive Polyphenol in Green Tea. Nutrients 2012, 4, 1679. [Google Scholar] [CrossRef]
- Leeman-Neill, R.J.; Wheeler, S.E.; Singh, S.V.; Thomas, S.M.; Seethala, R.R.; Neill, D.B.; Panahandeh, M.C.; Hahm, E.-R.; Joyce, S.C.; Sen, M.; et al. Guggulsterone Enhances Head and Neck Cancer Therapies via Inhibition of Signal Transducer and Activator of Transcription-3. Carcinogenesis 2009, 30, 1848. [Google Scholar] [CrossRef] [PubMed]
- Rauf, A.; Olatunde, A.; Imran, M.; Alhumaydhi, F.A.; Aljohani, A.S.M.; Khan, S.A.; Uddin, M.S.; Mitra, S.; bin Emran, T.; Khayrullin, M.; et al. Honokiol: A Review of Its Pharmacological Potential and Therapeutic Insights. Phytomedicine 2021, 90, 153647. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Shi, R.; Wang, X.; Shen, H.-M. Luteolin, a Flavonoid with Potentials for Cancer Prevention and Therapy. Curr. Cancer Drug Targets 2008, 8, 634. [Google Scholar] [CrossRef]
- Rapamune-SPC. Available online: https://www.ema.europa.eu/en/documents/product-information/rapamune-epar-product-information_en.pdf (accessed on 19 September 2021).
- Angevin, E.; Tabernero, J.; Elez, E.; Cohen, S.J.; Bahleda, R.; van Laethem, J.-L.; Ottensmeier, C.; Lopez-Martin, J.A.; Clive, S.; Joly, F.; et al. A Phase I/II, Multiple-Dose, Dose-Escalation Study of Siltuximab, an Anti-Interleukin-6 Monoclonal Antibody, in Patients with Advanced Solid Tumors. Clin. Cancer Res. 2014, 20, 2192–2204. [Google Scholar] [CrossRef]
- A Safety, Efficacy and Pharmacokinetic Study of Siltuximab (CNTO 328) in Participants with Solid Tumors-Full Text View-ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/show/NCT00841191 (accessed on 13 September 2021).
- Interleukin-6 Inhibitors | COVID-19 Treatment Guidelines. Available online: https://www.covid19treatmentguidelines.nih.gov/therapies/immunomodulators/interleukin-6-inhibitors/ (accessed on 12 September 2021).
- Actemra-Highlights of Prescribing Information. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2017/125276s114lbl.pdf (accessed on 19 September 2021).
- Kerschbaumer, A.; Sepriano, A.; Smolen, J.S.; van der Heijde, D.; Dougados, M.; van Vollenhoven, R.; McInnes, I.B.; Bijlsma, J.W.J.; Burmester, G.R.; de Wit, M.; et al. Efficacy of Pharmacological Treatment in Rheumatoid Arthritis: A Systematic Literature Research Informing the 2019 Update of the EULAR Recommendations for Management of Rheumatoid Arthritis. Ann. Rheum. Dis. 2020, 79, 744. [Google Scholar] [CrossRef] [PubMed]
- Fleischmann, R.; van Adelsberg, J.; Lin, Y.; da Castelar-Pinheiro, G.R.; Brzezicki, J.; Hrycaj, P.; Graham, N.M.H.; van Hoogstraten, H.; Bauer, D.; Burmester, G.R. Sarilumab and Nonbiologic Disease-Modifying Antirheumatic Drugs in Patients With Active Rheumatoid Arthritis and Inadequate Response or Intolerance to Tumor Necrosis Factor Inhibitors. Arthritis Rheumatol. 2017, 69, 277–290. [Google Scholar] [CrossRef]
- Kevzara-Highlights of Prescribing Information. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2017/761037s000lbl.pdf (accessed on 19 September 2021).
- Roche-Enspryng (Satralizumab). Available online: https://www.roche.com/products/product-details.htm?productId=6f4b47a2-7ca9-4768-bf1a-6d813702f5b6 (accessed on 16 September 2021).
- Eskandary, F.; Dürr, M.; Budde, K.; Doberer, K.; Reindl-Schwaighofer, R.; Waiser, J.; Wahrmann, M.; Regele, H.; Spittler, A.; Lachmann, N.; et al. Clazakizumab in Late Antibody-Mediated Rejection: Study Protocol of a Randomized Controlled Pilot Trial. Trials 2019, 20, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Rovin, B.H.; van Vollenhoven, R.F.; Aranow, C.; Wagner, C.; Gordon, R.; Zhuang, Y.; Belkowski, S.; Hsu, B. A Multicenter, Randomized, Double-Blind, Placebo-Controlled Study to Evaluate the Efficacy and Safety of Treatment With Sirukumab (CNTO 136) in Patients With Active Lupus Nephritis. Arthritis Rheumatol. 2016, 68, 2174–2183. [Google Scholar] [CrossRef]
- Reeh, H.; Rudolph, N.; Billing, U.; Christen, H.; Streif, S.; Bullinger, E.; Schliemann-Bullinger, M.; Findeisen, R.; Schaper, F.; Huber, H.J.; et al. Response to IL-6 Trans- and IL-6 Classic Signalling Is Determined by the Ratio of the IL-6 Receptor α to Gp130 Expression: Fusing Experimental Insights and Dynamic Modelling. Cell Commun. Signal. 2019, 17, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Jakafi-Highlights of Prescribing Information. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2019/202192s017lbl.pdf (accessed on 19 September 2021).
- Xeljanz-Highlights of Prescribing Information. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2018/203214s018lbl.pdf (accessed on 19 September 2021).
- Goel, P.; Gerriets, V. Chloroquine. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021; pp. 1–7. [Google Scholar]
- Jang, C.-H.; Choi, J.-H.; Byun, M.-S.; Jue, D.-M. Chloroquine Inhibits Production of TNF-α, IL-1β and IL-6 from Lipopolysaccharide-Stimulated Human Monocytes/Macrophages by Different Modes. Rheumatology 2006, 45, 703–710. [Google Scholar] [CrossRef]
- Aralen. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2017/006002s044lbl.pdf (accessed on 19 September 2021).
- Bansal, P.; Goyal, A.; Cusick, A., IV; Lahan, S.; Dhaliwal, H.S.; Bhyan, P.; Bhattad, P.B.; Aslam, F.; Ranka, S.; Dalia, T.; et al. Hydroxychloroquine: A Comprehensive Review and Its Controversial Role in Coronavirus Disease 2019. Ann. Med. 2021, 53, 117. [Google Scholar] [CrossRef] [PubMed]
- Plaquenil. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2017/009768s037s045s047lbl.pdf (accessed on 19 September 2021).
- Wang, Y.; Huang, Z.; Wang, L.; Meng, S.; Fan, Y.; Chen, T.; Cao, J.; Jiang, R.; Wang, C. The Anti-Malarial Artemisinin Inhibits pro-Inflammatory Cytokines via the NF-ΚB Canonical Signaling Pathway in PMA-Induced THP-1 Monocytes. Int. J. Mol. Med. 2011, 27, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Artesunate-Highlights of Prescribing Information. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2020/213036s000lbl.pdf (accessed on 19 September 2021).
- Lin, L.; Benson, D.M.; DeAngelis, S.; Bakan, C.E.; Li, P.-K.; Li, C.; Lin, J. A Small Molecule, LLL12 Inhibits Constitutive STAT3 and IL-6-Induced STAT3 Signaling and Exhibits Potent Growth Suppressive Activity in Human Multiple Myeloma Cells. Int. J. Cancer 2012, 130, 1459–1469. [Google Scholar] [CrossRef]
- Shi, W.; Yan, D.; Zhao, C.; Xiao, M.; Wang, Y.; Ma, H.; Liu, T.; Qin, H.; Zhang, C.; Li, C.; et al. Inhibition of IL-6/STAT3 Signaling in Human Cancer Cells Using Evista. Biochem. Biophys. Res. Commun. 2017, 491, 159–165. [Google Scholar] [CrossRef]
- Evista-Highlights of Prescribing Information. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2007/020815s018lbl.pdf (accessed on 19 September 2021).
- Hayashi, M.; Rho, M.-C.; Enomoto, A.; Fukami, A.; Kim, Y.-P.; Kikuchi, Y.; Sunazuka, T.; Hirose, T.; Komiyama, K.; Omura, S. Suppression of Bone Resorption by Madindoline A, a Novel Nonpeptide Antagonist to Gp130. Proc. Natl. Acad. Sci. USA 2002, 99, 14728. [Google Scholar] [CrossRef]
- Singh, S.; Gajulapati, V.; Gajulapati, K.; Goo, J.; Park, Y.H.; Jung, H.Y.; Lee, S.Y.; Choi, J.H.; Kim, Y.K.; Lee, K.; et al. Structure–Activity Relationship Study of a Series of Novel Oxazolidinone Derivatives as IL-6 Signaling Blockers. Bioorganic Med. Chem. Lett. 2016, 26, 1282–1286. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, L.; Xie, N.; Nice, E.C.; Zhang, T.; Cui, Y.; Huang, C. Overcoming Cancer Therapeutic Bottleneck by Drug Repurposing. Signal Transduct. Target. Ther. 2020, 5, 1–25. [Google Scholar] [CrossRef]
- Gyebi, G.A.; Ogunyemi, O.M.; Ibrahim, I.M.; Afolabi, S.O.; Adebayo, J.O. Dual Targeting of Cytokine Storm and Viral Replication in COVID-19 by Plant-Derived Steroidal Pregnanes: An in Silico Perspective. Comput. Biol. Med. 2021, 134, 104406. [Google Scholar] [CrossRef]
- Results of a Phase IIb Study of Vobarilizumab, an Anti-Interleukin 6 Receptor Nanobody®, in Patients with Moderate-to-Severe Rheumatoid Arthritis Despite Treatment with Methotrexate. Available online: https://www.ablynx.com/uploads/data/files/cra2017_alx-0061_abstract%20210_poster_final.pdf (accessed on 3 October 2021).
- Genovese, M.C.; Durez, P.; Fleischmann, R.; Tanaka, Y.; Furst, D.; Yamanaka, H.; Korneva, E.; Vasyutin, I.; Takeuchi, T. Long-Term Safety and Efficacy of Olokizumab in Patients with Rheumatoid Arthritis and Inadequate Response to Tumor Necrosis Factor Inhibitor Therapy in Phase II Studies. Eur. J. Rheumatol. 2021, 8, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Baricitinib Letter of Authorization Revised July 28 2021. Available online: https://www.fda.gov/media/143822/download (accessed on 3 October 2021).
- FDA Briefing Document Pharmacy Compounding Advisory Committee (PCAC) Meeting. Available online: https://www.fda.gov/media/95976/download (accessed on 3 October 2021).



{kind=link}
{kind=link}
| Agent | Mode of Action | Tested in HNSCC | Approved for Human Application in (Primary Indication): | ||
|---|---|---|---|---|---|
| In Vitro | In Vivo (Animal Study) | Clinical Trial (Registered on Clinicaltrials.gov) | |||
| 2-O-Methylmagnolol | ◼ downstream signaling inhibition | Wang et al., 2018 [207] | Wang et al., 2018 [207] | - | - |
| AMD3100 (CXCR4 inhibitor) | ✼ inhibition of IL-6 production | Tang et al., 2008 [187] | - | - | - |
| PD98059 (MEK inhibitor) | ✼ inhibition of IL-6 production | Tang et al., 2008 [187] | - | - | - |
| Ammonium pyrrolidinecarbodithioate (PDTC) | ✼ inhibition of IL-6 production | Tang et al., 2008 [187] | - | - | - |
| Aryl hydrocarbon receptor antagonist (BAY 2416964) | ✼ inhibition of IL-6 production | DiNatale et al., 2011, 2012 [185,186] | - | NCT04069026 § [212] | - |
| Bazedoxifene | ◆ gp130 blockade | Yadav et al., 2017 [209] | Yadav et al., 2017 [209] | - | Postmenopausal osteoporosis [213] |
| Cepharanthin | ✼ inhibition of IL-6 production | Tamatani et al., 2007 [180] | - | - | - |
| Cilengitide (EMD 121974) + cetuximab | ✼ inhibition of IL-6 production | Wichmann et al., 2017 [177] | - | - | Cetuximab–metastatic colorectal cancer, advanced HNSCC [214] |
| Curcumin | ✼ inhibition of IL-6 production ◼ downstream signaling inhibition | Chakravarti et al., 2006 [184] Cohen et al., 2009 [183] Meyer et al., 2011 [210] | Yu et al., 2013 [63] | NCT04208334; Thambamroong et al., 2016 [215] | Dietary supplement [216] |
| Celecoxib + Simvastatin | ✼ inhibition of IL-6 production | Gehrke et al. 2017 [208] | - | - | Celecoxib - NSAID (non-steroidal anti-inflammatory drug) [217] Simvastatin - reduction of LDL (low-density lipoprotein) cholesterol blood levels [218] |
| Cyclopentenone prostaglandin 15d-PGJ2 | ◼ downstream signaling inhibition | Siavash et al., 2004 [188] | - | - | - |
| Epigallocatechin gallate | ◼ downstream signaling inhibition | Lin et al., 2012 [182] | - | - | Dietary supplement [219] |
| Guggulsterone | ◼ downstream signaling inhibition | Macha et al., 2011 [181] Leeman-Neill et al. [220] | Leeman-Neill et al. [220] | - | Dietary supplement [220] |
| Honokiol | ✼ inhibition of IL-6 production ◼ downstream signaling inhibition | Chang et al., 2018 [206] | - | - | Dietary supplement [221] |
| Lactoferrin (Human, recombinant) | ✼ inhibition of IL-6 production | Wolf et al. [178] | Wolf et al. [178] | - | - |
| RhoC knockdown (lentiviral vector-based shRNA) | ✼ inhibition of IL-6 production | Islam et al. [193] | - | - | - |
| L-leucine-methylester (lysosomotropic agent) | ✼ inhibition of IL-6 production | Kross et al. [199] | - | - | - |
| Luteolin | ◼ downstream signaling inhibition | Tu et al. [179] | - | - | Dietary supplement, popular in chinese traditional medicine [222] |
| MEDI5117 (anti-IL-6 humanized monoclonal antibody, IgG1, with Enhanced Serum Half-Life; also known as WBP216) | ♢ IL-6 neutralization | Finkel et al., 2016 [195] | Finkel et al., 2016 [195] | - | - |
| P276-00 (cyclin-dependent kinase inhibitor) | ✼ inhibition of IL-6 production | Mishra et al., 2013 [200] | Mishra et al., 2013 [200] | - | - |
| Rapamycin | ✼ inhibition of IL-6 production | Ekshyyan et al., 2016 [196] | - | - | Immunosuppressive drug [223] |
| SB203580 (p38 MAPK inhibitor) | ✼ inhibition of IL-6 production | Riebe et al., 2007 [197] Jing et al., 2016 [198] | Jing et al., 2016 [198] | - | - |
| Siltuximab (anti-IL-6 chimeric (human-murine) monoclonal recombinant antibody (IgG1κ)) | ♢ IL-6 neutralization | - | - | NCT00841191; Angevin et al., 2014 [224,225] | Management of systemic inflammation in COVID-19 [226] |
| Insulin-like growth factor-II mRNA binding protein-3 and podoplanin knockdown (lentiviral vector-based shRNA) | ✼ inhibition of IL-6 production | Hwang et al., 2016 [194] | Hwang et al., 2016 [194] | - | - |
| TLR-9- knockdown (siRNAs) | ✼ inhibition of IL-6 production | Ruan et al., 2014 [211] | - | - | - |
| Tristetraprolin knockdown (lentiviral vector-based shRNA) | ✼ inhibition of IL-6 production | Van Tubergen et al., 2011, 2013 [203,204] | Van Tubergen et al., 2013 [204] | - | - |
| Tetrathiomolybdate | ✼ inhibition of IL-6 production | Teknos et al., 2005 [202] | Teknos et al., 2005 [202] | - | - |
| Tocilizumab (anti-IL-6R humanised IgG1 monoclonal recombinant antibody ) | ❖ IL-6R blockade | Matsuoka et al., 2016 [88] Stanam et al., 2015 [190] Shinriki et al., 2009, 2011 [191,192] | Poth et al., 2010 [189] Stanam et al., 2015 [190] Shinriki et al., 2009, 2011 [191,192] | - | Rheumatoid arthritis [227] Giant cell arteritis [227] Polyarticular Juvenile idiopathic arthritis [227] Systemic Juvenile Idiopathic arthritis [227] Cytokine Release syndrome [227] Management of systemic inflammation in COVID-19 [226] |
| Triazolothiadiazine | ◆ gp130 blockade | Sen et al., 2017 [201] | - | - | - |
| Oxazole-piperazine | ◆ gp130 blockade | Sen et al., 2017 [201] | - | ||
| WP1066 (JAK inhibitor) | ◼ Downstream signaling inhibition | Zhou et al., 2014 [205] | Zhou et al., 2014 [205] | - | - |
| YM529 (third-generation bisphosphonate, Minodronic acid) | ✼ inhibition of IL-6 production | - | Cui et al., 2005 [176] | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Španko, M.; Strnadová, K.; Pavlíček, A.J.; Szabo, P.; Kodet, O.; Valach, J.; Dvořánková, B.; Smetana, K., Jr.; Lacina, L. IL-6 in the Ecosystem of Head and Neck Cancer: Possible Therapeutic Perspectives. Int. J. Mol. Sci. 2021, 22, 11027. https://doi.org/10.3390/ijms222011027
Španko M, Strnadová K, Pavlíček AJ, Szabo P, Kodet O, Valach J, Dvořánková B, Smetana K Jr., Lacina L. IL-6 in the Ecosystem of Head and Neck Cancer: Possible Therapeutic Perspectives. International Journal of Molecular Sciences. 2021; 22(20):11027. https://doi.org/10.3390/ijms222011027
Chicago/Turabian StyleŠpanko, Michal, Karolína Strnadová, Aleš Jan Pavlíček, Pavol Szabo, Ondřej Kodet, Jaroslav Valach, Barbora Dvořánková, Karel Smetana, Jr., and Lukáš Lacina. 2021. "IL-6 in the Ecosystem of Head and Neck Cancer: Possible Therapeutic Perspectives" International Journal of Molecular Sciences 22, no. 20: 11027. https://doi.org/10.3390/ijms222011027
APA StyleŠpanko, M., Strnadová, K., Pavlíček, A. J., Szabo, P., Kodet, O., Valach, J., Dvořánková, B., Smetana, K., Jr., & Lacina, L. (2021). IL-6 in the Ecosystem of Head and Neck Cancer: Possible Therapeutic Perspectives. International Journal of Molecular Sciences, 22(20), 11027. https://doi.org/10.3390/ijms222011027