
Female Mice with Selenocysteine tRNA Deletion in Agrp Neurons Maintain Leptin Sensitivity and Resist Weight Gain While on a High-Fat Diet

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Results
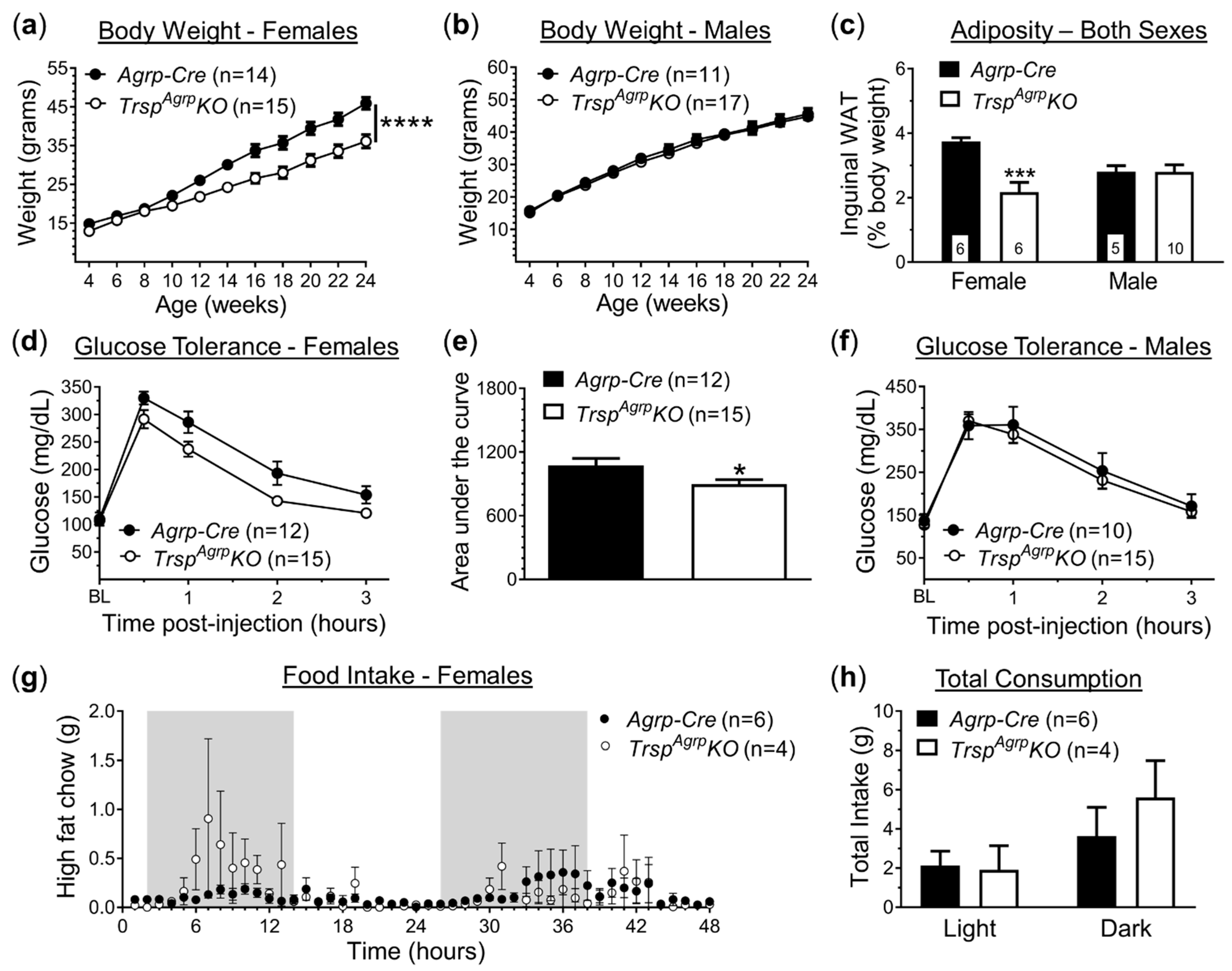
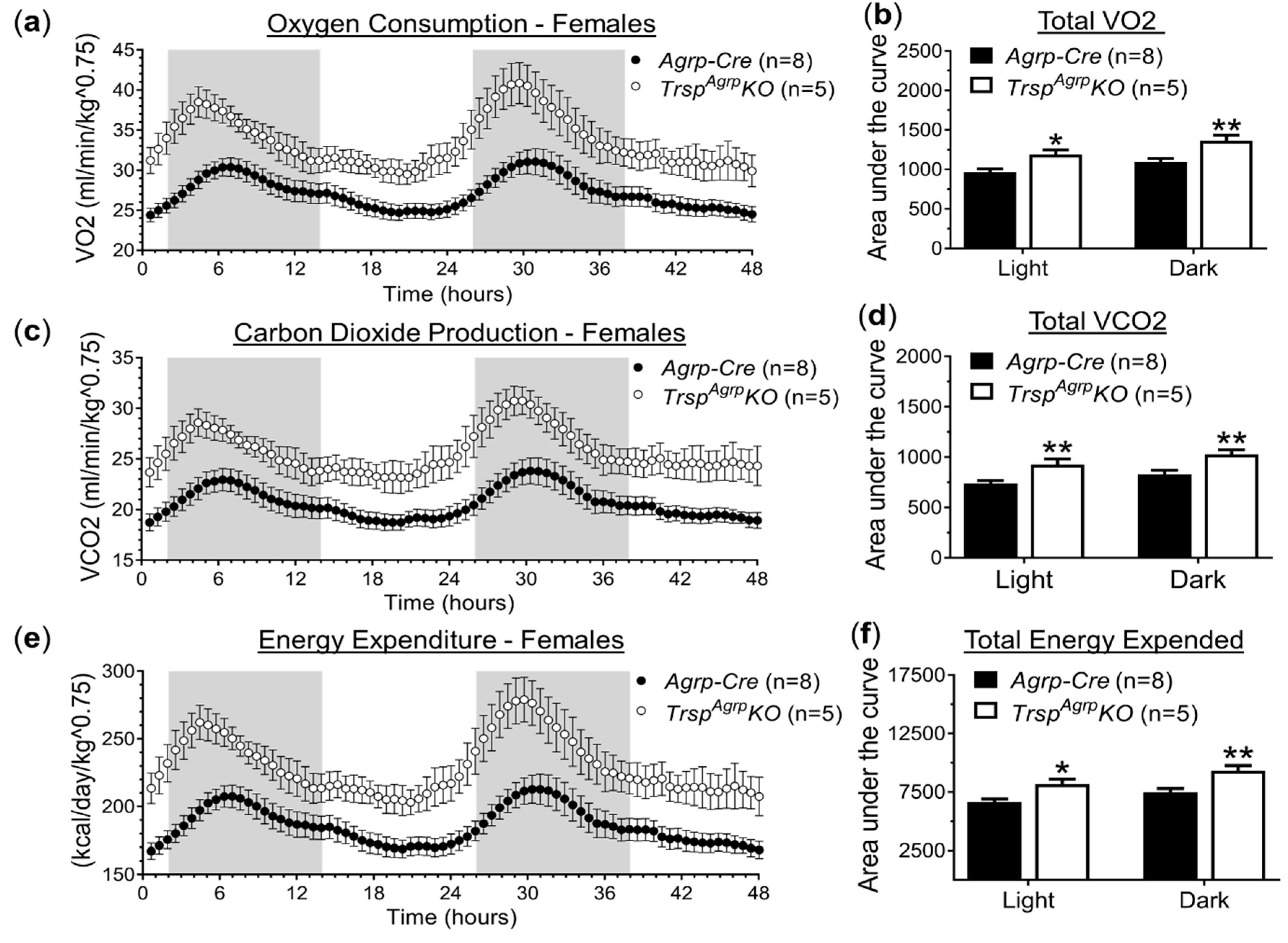
2.1. In Vivo Metabolic Assessment of Mice with Agrp Neuron-specific Ablation of Trsp
2.2. Measurement of Hormones in Serum from TrspAgrpKO Mice
2.3. Histological Analysis of TrspAgrpKO Mouse Brown Adipose Tissue
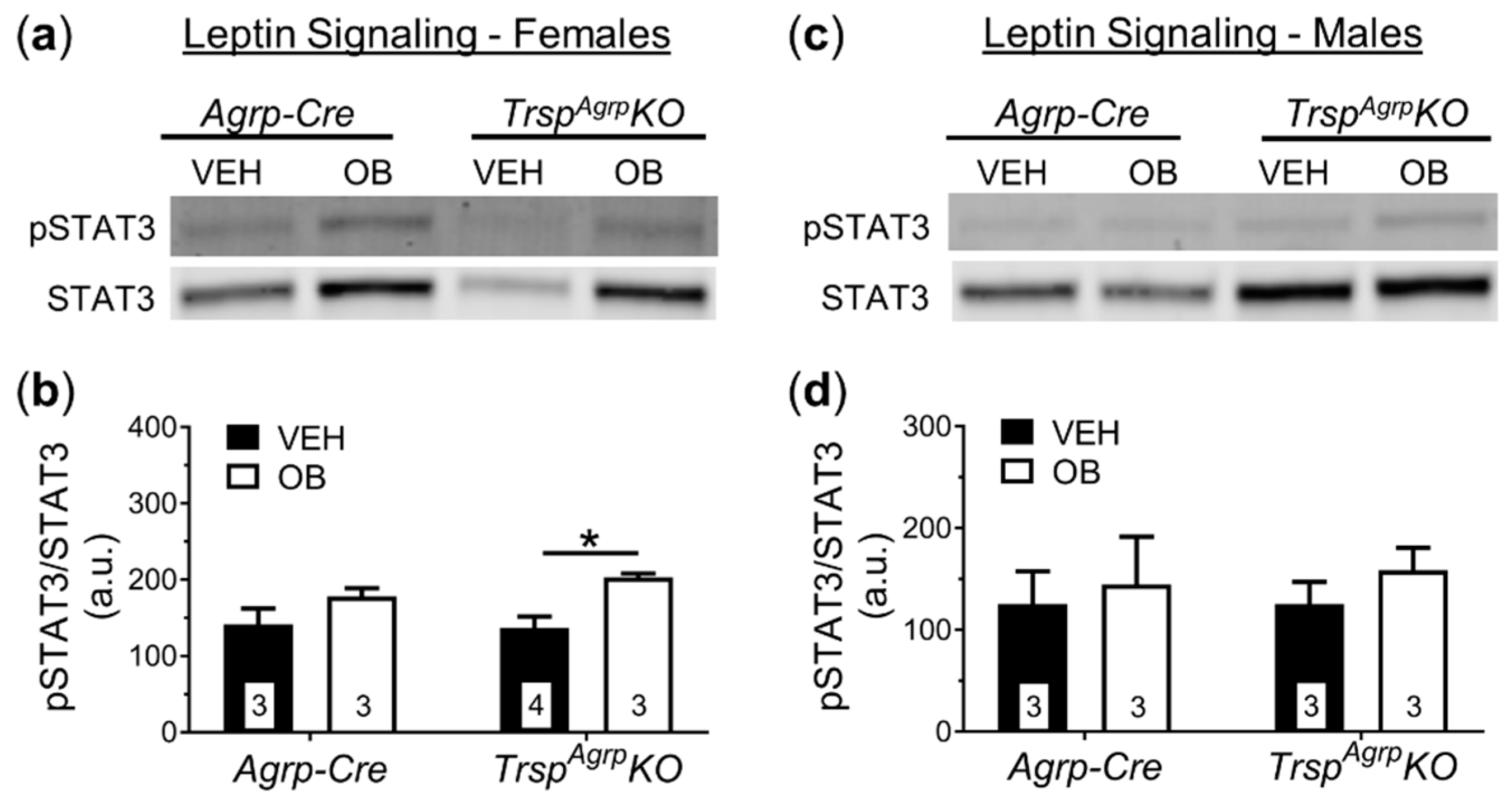
2.4. Assessment of Leptin Sensitivity in the Hypothalamus of TrspAgrpKO Mice
2.5. Analysis of Neuropeptide Expression in the TrspAgrpKO Mouse Hypothalamus
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Design
4.3. Metabolic Chambers
4.4. Glucose Tolerance Test
4.5. Leptin Challenge and Tissue Collection
4.6. Gel Electrophoresis and Western Blotting
4.7. Immunohistochemistry and Histology
4.8. Stereology and Data Quantification
4.9. Measurement of Serum Hormone Levels
4.10. Antibodies
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Agrp | agouti-related peptide |
| Arc | arcuate nucleus |
| BAT | brown adipose tissue |
| CRH | corticotropin-releasing hormone |
| DAB | 3,3′-diaminobenzidine |
| ER | endoplasmic reticulum |
| ERα | estrogen receptor α |
| FSH | follicle-stimulating hormone |
| GABA | γ-Aminobutyric acid |
| GH | growth hormone |
| DIO | diet-induced obesity |
| DMH | dorsomedial hypothalamus |
| HFD | high-fat diet |
| ME | median eminence |
| Npy | neuropeptide y |
| Pomc | pro-opiomelanocortin |
| RIP | rat-insulin-promoter |
| ROS | reactive oxygen species |
| Scly | selenocysteine lyase |
| SELENOM | selenoprotein M |
| STAT3 | signal transducer and activator of transcription 3 |
| TRH | thyrotropin-releasing hormone |
| Trsp | selenocysteine tRNA[Ser]Sec gene |
| UCP1 | uncoupling protein-1 |
| WAT | white adipose tissue |
References
- Caballero, B. Humans against Obesity: Who Will Win? Adv. Nutr. 2019, 10, S4–S9. [Google Scholar] [CrossRef]
- Zagmutt, S.; Mera, P.; Soler-Vazquez, M.C.; Herrero, L.; Serra, D. Targeting AgRP neurons to maintain energy balance: Lessons from animal models. Biochem. Pharmacol. 2018, 155, 224–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, T.; Torres, D.J.; Berry, M.J.; Pitts, M.W. Hypothalamic redox balance and leptin signaling—Emerging role of selenoproteins. Free Radic. Biol. Med. 2018, 127, 172–181. [Google Scholar] [CrossRef]
- Schomburg, L. Selenium, selenoproteins and the thyroid gland: Interactions in health and disease. Nat Rev. Endocrinol 2011, 8, 160–171. [Google Scholar] [CrossRef]
- Youn, H.S.; Lim, H.J.; Choi, Y.J.; Lee, J.Y.; Lee, M.Y.; Ryu, J.H. Selenium suppresses the activation of transcription factor NF-kappa B and IRF3 induced by TLR3 or TLR4 agonists. Int. Immunopharmacol. 2008, 8, 495–501. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, Y.; Schweizer, U.; Savaskan, N.E.; Hua, D.; Kipnis, J.; Hatfield, D.L.; Gladyshev, V.N. Comparative analysis of selenocysteine machinery and selenoproteome gene expression in mouse brain identifies neurons as key functional sites of selenium in mammals. J. Biol. Chem. 2008, 283, 2427–2438. [Google Scholar] [CrossRef] [Green Version]
- Henry, F.E.; Sugino, K.; Tozer, A.; Branco, T.; Sternson, S.M. Cell type-specific transcriptomics of hypothalamic energy-sensing neuron responses to weight-loss. eLife 2015, 4, e09800. [Google Scholar] [CrossRef]
- Yagishita, Y.; Uruno, A.; Fukutomi, T.; Saito, R.; Saigusa, D.; Pi, J.; Fukamizu, A.; Sugiyama, F.; Takahashi, S.; Yamamoto, M. Nrf2 Improves Leptin and Insulin Resistance Provoked by Hypothalamic Oxidative Stress. Cell. Rep. 2017, 18, 2030–2044. [Google Scholar] [CrossRef] [Green Version]
- Schriever, S.C.; Zimprich, A.; Pfuhlmann, K.; Baumann, P.; Giesert, F.; Klaus, V.; Kabra, D.G.; Hafen, U.; Romanov, A.; Tschop, M.H.; et al. Alterations in neuronal control of body weight and anxiety behavior by glutathione peroxidase 4 deficiency. Neuroscience 2017, 357, 241–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, D.J.; Pitts, M.W.; Hashimoto, A.C.; Berry, M.J. Agrp-Specific Ablation of Scly Protects against Diet-Induced Obesity and Leptin Resistance. Nutrients 2019, 11, 1693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deem, J.D.; Faber, C.L.; Morton, G.J. AgRP neurons: Regulators of feeding, energy expenditure, and behavior. FEBS J. 2021. [Google Scholar] [CrossRef] [PubMed]
- Cavaliere, G.; Viggiano, E.; Trinchese, G.; De Filippo, C.; Messina, A.; Monda, V.; Valenzano, A.; Cincione, R.I.; Zammit, C.; Cimmino, F.; et al. Long Feeding High-Fat Diet Induces Hypothalamic Oxidative Stress and Inflammation, and Prolonged Hypothalamic AMPK Activation in Rat Animal Model. Front. Physiol. 2018, 9, 818. [Google Scholar] [CrossRef]
- Gong, T.; Hashimoto, A.C.; Sasuclark, A.R.; Khadka, V.S.; Gurary, A.; Pitts, M.W. Selenoprotein M Promotes Hypothalamic Leptin Signaling and Thioredoxin Antioxidant Activity. Antioxid. Redox. Signal. 2019, 35, 775–787. [Google Scholar] [CrossRef]
- Pitts, M.W.; Reeves, M.A.; Hashimoto, A.C.; Ogawa, A.; Kremer, P.; Seale, L.A.; Berry, M.J. Deletion of selenoprotein M leads to obesity without cognitive deficits. J. Biol. Chem. 2013, 288, 26121–26134. [Google Scholar] [CrossRef] [Green Version]
- Phillips, K.J. Beige Fat, Adaptive Thermogenesis, and Its Regulation by Exercise and Thyroid Hormone. Biology 2019, 8, 57. [Google Scholar] [CrossRef] [Green Version]
- Orthofer, M.; Valsesia, A.; Magi, R.; Wang, Q.P.; Kaczanowska, J.; Kozieradzki, I.; Leopoldi, A.; Cikes, D.; Zopf, L.M.; Tretiakov, E.O.; et al. Identification of ALK in Thinness. Cell 2020, 181, 1246–1262.e22. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Ji, Y.; Yuen, T.; Rendina-Ruedy, E.; DeMambro, V.E.; Dhawan, S.; Abu-Amer, W.; Izadmehr, S.; Zhou, B.; Shin, A.C.; et al. Blocking FSH induces thermogenic adipose tissue and reduces body fat. Nature 2017, 546, 107–112. [Google Scholar] [CrossRef]
- Oldfield, B.J.; Giles, M.E.; Watson, A.; Anderson, C.; Colvill, L.M.; McKinley, M.J. The neurochemical characterisation of hypothalamic pathways projecting polysynaptically to brown adipose tissue in the rat. Neuroscience 2002, 110, 515–526. [Google Scholar] [CrossRef]
- Bamshad, M.; Song, C.K.; Bartness, T.J. CNS origins of the sympathetic nervous system outflow to brown adipose tissue. Am. J. Physiol. 1999, 276, R1569–R1578. [Google Scholar] [CrossRef]
- Zaretskaia, M.V.; Zaretsky, D.V.; Shekhar, A.; DiMicco, J.A. Chemical stimulation of the dorsomedial hypothalamus evokes non-shivering thermogenesis in anesthetized rats. Brain Res. 2002, 928, 113–125. [Google Scholar] [CrossRef]
- Ye, Z.; Liu, G.; Guo, J.; Su, Z. Hypothalamic endoplasmic reticulum stress as a key mediator of obesity-induced leptin resistance. Obes. Rev. 2018, 19, 770–785. [Google Scholar] [CrossRef]
- Olofsson, L.E.; Unger, E.K.; Cheung, C.C.; Xu, A.W. Modulation of AgRP-neuronal function by SOCS3 as an initiating event in diet-induced hypothalamic leptin resistance. Proc. Natl Acad. Sci. USA 2013, 110, E697–E706. [Google Scholar] [CrossRef] [Green Version]
- Friedman, J.M. Leptin and the endocrine control of energy balance. Nat. Metab. 2019, 1, 754–764. [Google Scholar] [CrossRef] [PubMed]
- Pierce, A.A.; Xu, A.W. De novo neurogenesis in adult hypothalamus as a compensatory mechanism to regulate energy balance. J. Neurosci. 2010, 30, 723–730. [Google Scholar] [CrossRef] [Green Version]
- Luquet, S.; Perez, F.A.; Hnasko, T.S.; Palmiter, R.D. NPY/AgRP neurons are essential for feeding in adult mice but can be ablated in neonates. Science 2005, 310, 683–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, A.W.; Kaelin, C.B.; Morton, G.J.; Ogimoto, K.; Stanhope, K.; Graham, J.; Baskin, D.G.; Havel, P.; Schwartz, M.W.; Barsh, G.S. Effects of hypothalamic neurodegeneration on energy balance. PLoS Biol. 2005, 3, 415. [Google Scholar] [CrossRef] [PubMed]
- Bosl, M.R.; Takaku, K.; Oshima, M.; Nishimura, S.; Taketo, M.M. Early embryonic lethality caused by targeted disruption of the mouse selenocysteine tRNA gene (Trsp). Proc. Natl Acad. Sci. USA 1997, 94, 5531–5534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirth, E.K.; Conrad, M.; Winterer, J.; Wozny, C.; Carlson, B.A.; Roth, S.; Schmitz, D.; Bornkamm, G.W.; Coppola, V.; Tessarollo, L.; et al. Neuronal selenoprotein expression is required for interneuron development and prevents seizures and neurodegeneration. FASEB J. 2010, 24, 844–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diano, S.; Liu, Z.W.; Jeong, J.K.; Dietrich, M.O.; Ruan, H.B.; Kim, E.; Suyama, S.; Kelly, K.; Gyengesi, E.; Arbiser, J.L.; et al. Peroxisome proliferation-associated control of reactive oxygen species sets melanocortin tone and feeding in diet-induced obesity. Nat. Med. 2011, 17, 1121–1127. [Google Scholar] [CrossRef] [Green Version]
- Kuo, D.Y.; Chen, P.N.; Yang, S.F.; Chu, S.C.; Chen, C.H.; Kuo, M.H.; Yu, C.H.; Hsieh, Y.S. Role of reactive oxygen species-related enzymes in neuropeptide y and proopiomelanocortin-mediated appetite control: A study using atypical protein kinase C knockdown. Antioxid. Redox Signal. 2011, 15, 2147–2159. [Google Scholar] [CrossRef] [PubMed]
- Ogawa-Wong, A.N.; Hashimoto, A.C.; Ha, H.; Pitts, M.W.; Seale, L.A.; Berry, M.J. Sexual Dimorphism in the Selenocysteine Lyase Knockout Mouse. Nutrients 2018, 10, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seale, L.A.; Gilman, C.L.; Hashimoto, A.C.; Ogawa-Wong, A.N.; Berry, M.J. Diet-induced obesity in the selenocysteine lyase knockout mouse. Antioxid Redox Signal. 2015, 23, 761–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seale, L.A.; Hashimoto, A.C.; Kurokawa, S.; Gilman, C.L.; Seyedali, A.; Bellinger, F.P.; Raman, A.V.; Berry, M.J. Disruption of the selenocysteine lyase-mediated selenium recycling pathway leads to metabolic syndrome in mice. Mol. Cell. Biol. 2012, 32, 4141–4154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romanov, G.A.; Sukhoverov, V.S. Arginine CGA codons as a source of nonsense mutations: A possible role in multivariant gene expression, control of mRNA quality, and aging. Mol. Genet. Genom. 2017, 292, 1013–1026. [Google Scholar] [CrossRef]
- Lu, J.; Zhong, L.; Lonn, M.E.; Burk, R.F.; Hill, K.E.; Holmgren, A. Penultimate selenocysteine residue replaced by cysteine in thioredoxin reductase from selenium-deficient rat liver. FASEB J. 2009, 23, 2394–2402. [Google Scholar] [CrossRef] [Green Version]
- Tobe, R.; Naranjo-Suarez, S.; Everley, R.A.; Carlson, B.A.; Turanov, A.A.; Tsuji, P.A.; Yoo, M.H.; Gygi, S.P.; Gladyshev, V.N.; Hatfield, D.L. High error rates in selenocysteine insertion in mammalian cells treated with the antibiotic doxycycline, chloramphenicol, or geneticin. J. Biol. Chem. 2013, 288, 14709–14715. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.M.; Turanov, A.A.; Carlson, B.A.; Yoo, M.H.; Everley, R.A.; Nandakumar, R.; Sorokina, I.; Gygi, S.P.; Gladyshev, V.N.; Hatfield, D.L. Targeted insertion of cysteine by decoding UGA codons with mammalian selenocysteine machinery. Proc. Natl Acad. Sci. USA 2010, 107, 21430–21434. [Google Scholar] [CrossRef] [Green Version]
- Pitts, M.W.; Kremer, P.M.; Hashimoto, A.C.; Torres, D.J.; Byrns, C.N.; Williams, C.S.; Berry, M.J. Competition between the Brain and Testes under Selenium-Compromised Conditions: Insight into Sex Differences in Selenium Metabolism and Risk of Neurodevelopmental Disease. J. Neurosci. 2015, 35, 15326–15338. [Google Scholar] [CrossRef] [Green Version]
- Kremer, P.M.; Torres, D.J.; Hashimoto, A.C.; Berry, M.J. Sex-Specific Metabolic Impairments in a Mouse Model of Disrupted Selenium Utilization. Front. Nutr. 2021, 8, 682700. [Google Scholar] [CrossRef]
- Seale, L.A.; Ogawa-Wong, A.N.; Berry, M.J. Sexual Dimorphism in Selenium Metabolism and Selenoproteins. Free Radic. Biol. Med. 2018, 127, 198–205. [Google Scholar] [CrossRef]
- Oyola, M.G.; Handa, R.J. Hypothalamic-pituitary-adrenal and hypothalamic-pituitary-gonadal axes: Sex differences in regulation of stress responsivity. Stress 2017, 20, 476–494. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.A.; Yoo, S.; Pak, T.; Salvatierra, J.; Velarde, E.; Aja, S.; Blackshaw, S. Dietary and sex-specific factors regulate hypothalamic neurogenesis in young adult mice. Front. Neurosci. 2014, 8, 157. [Google Scholar] [CrossRef] [Green Version]
- Bless, E.P.; Reddy, T.; Acharya, K.D.; Beltz, B.S.; Tetel, M.J. Oestradiol and diet modulate energy homeostasis and hypothalamic neurogenesis in the adult female mouse. J. Neuroendocrinol. 2014, 26, 805–816. [Google Scholar] [CrossRef] [Green Version]
- Bless, E.P.; Yang, J.; Acharya, K.D.; Nettles, S.A.; Vassoler, F.M.; Byrnes, E.M.; Tetel, M.J. Adult Neurogenesis in the Female Mouse Hypothalamus: Estradiol and High-Fat Diet Alter the Generation of Newborn Neurons Expressing Estrogen Receptor alpha. eNeuro 2016, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olofsson, L.E.; Pierce, A.A.; Xu, A.W. Functional requirement of AgRP and NPY neurons in ovarian cycle-dependent regulation of food intake. Proc. Natl Acad. Sci. USA 2009, 106, 15932–15937. [Google Scholar] [CrossRef] [Green Version]
- De Souza, F.S.; Nasif, S.; Lopez-Leal, R.; Levi, D.H.; Low, M.J.; Rubinsten, M. The estrogen receptor alpha colocalizes with proopiomelanocortin in hypothalamic neurons and binds to a conserved motif present in the neuron-specific enhancer nPE2. Eur. J. Pharmacol. 2011, 660, 181–187. [Google Scholar] [CrossRef] [Green Version]
- Yulyaningsih, E.; Rudenko, I.A.; Valdearcos, M.; Dahlen, E.; Vagena, E.; Chan, A.; Alvarez-Buylla, A.; Vaisse, C.; Koliwad, S.K.; Xu, A.W. Acute Lesioning and Rapid Repair of Hypothalamic Neurons outside the Blood-Brain Barrier. Cell. Rep. 2017, 19, 2257–2271. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.N.; Jung, Y.S.; Kwon, H.J.; Seong, J.K.; Granneman, J.G.; Lee, Y.H. Sex differences in sympathetic innervation and browning of white adipose tissue of mice. Biol. Sex Differ. 2016, 7, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toth, M.J.; Gardner, A.W.; Arciero, P.J.; Calles-Escandon, J.; Poehlman, E.T. Gender differences in fat oxidation and sympathetic nervous system activity at rest and during submaximal exercise in older individuals. Clin. Sci. 1998, 95, 59–66. [Google Scholar] [CrossRef]
- Herz, C.T.; Kulterer, O.C.; Prager, M.; Marculescu, R.; Langer, F.B.; Prager, G.; Kautzky-Willer, A.; Haug, A.R.; Kiefer, F.W. Sex differences in brown adipose tissue activity and cold-induced thermogenesis. Mol. Cell. Endocrinol. 2021, 534, 111365. [Google Scholar] [CrossRef]
- Tong, Q.; Ye, C.P.; Jones, J.E.; Elmquist, J.K.; Lowell, B.B. Synaptic release of GABA by AgRP neurons is required for normal regulation of energy balance. Nat. Neurosci. 2008, 11, 998–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumaraswamy, E.; Carlson, B.A.; Morgan, F.; Miyoshi, K.; Robinson, G.W.; Su, D.; Wang, S.; Southon, E.; Tessarollo, L.; Lee, B.J.; et al. Selective removal of the selenocysteine tRNA [Ser]Sec gene (Trsp) in mouse mammary epithelium. Mol. Cell. Biol. 2003, 23, 1477–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hormone Type and Name | Agrp-Cre | TrspAgrpKO | p Value | |
|---|---|---|---|---|
| Females: | ||||
| Metabolic hormones | ||||
| Leptin (ng/mL) | 32.78 ± 1.58 (n = 7) | 31.54 ± 2.18 (n = 9) | 0.67 | |
| Insulin (ng/mL) | 0.54 ± 0.11 (n = 7) | 0.32 ± 0.04 (n = 8) | 0.06 | |
| Pituitary hormones | ||||
| ACTH (pg/mL) | 20.06 ± 4.69 (n = 7) | 21.75 ± 4.11 (n = 8) | 0.79 | |
| FSH (ng/mL) | 1.41 ± 0.54 (n = 6) | 0.39 ± 0.11 (n = 8) | 0.05 | |
| GH (ng/mL) | 1.09 ± 0.43 (n = 6) | 1.97 ± 0.74 (n = 9) | 0.39 | |
| LH (pg/mL) | 125.65 ± 43.95 (n = 7) | 248.90 ± 76.29 (n = 8) | 0.20 | |
| Prolactin (ng/mL) | 9.72 ± 2.10 (n = 7) | 8.36 ± 1.49 (n = 8) | 0.60 | |
| TSH (pg/mL) | 49.40 ± 5.27 (n = 6) | 61.57 ± 25.53 (n = 8) | 0.67 | |
| Males: | ||||
| Metabolic hormones | ||||
| Leptin (ng/mL) | 32.58 ± 2.26 (n = 8) | 28.38 ± 2.15 (n = 9) | 0.20 | |
| Insulin (ng/mL) | 0.93 ± 0.26 (n = 8) | 1.30 ± 0.37 (n = 9) | 0.44 | |
| Pituitary hormones | ||||
| ACTH (pg/mL) | 17.94 ± 3.46 (n = 8) | 22.05 ± 5.53 (n = 9) | 0.55 | |
| FSH (ng/mL) | 3.19 ± 0.67 (n = 8) | 2.77 ± 0.41 (n = 9) | 0.59 | |
| GH (ng/mL) | 0.54 ± 0.09 (n = 6) | 2.14 ± 0.54 (n = 9) | 0.03 | |
| LH (pg/mL) | 392.17 ± 166.81 (n = 8) | 653.28 ± 200.41 (n = 9) | 0.34 | |
| Prolactin (ng/mL) | 1.15 ± 0.33 (n = 8) | 2.07 ± 0.85 (n = 9) | 0.35 | |
| TSH (pg/mL) | 173.10 ± 43.22 (n = 8) | 258.30 ± 53.23 (n = 9) | 0.24 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres, D.J.; Pitts, M.W.; Seale, L.A.; Hashimoto, A.C.; An, K.J.; Hanato, A.N.; Hui, K.W.; Remigio, S.M.A.; Carlson, B.A.; Hatfield, D.L.; et al. Female Mice with Selenocysteine tRNA Deletion in Agrp Neurons Maintain Leptin Sensitivity and Resist Weight Gain While on a High-Fat Diet. Int. J. Mol. Sci. 2021, 22, 11010. https://doi.org/10.3390/ijms222011010
Torres DJ, Pitts MW, Seale LA, Hashimoto AC, An KJ, Hanato AN, Hui KW, Remigio SMA, Carlson BA, Hatfield DL, et al. Female Mice with Selenocysteine tRNA Deletion in Agrp Neurons Maintain Leptin Sensitivity and Resist Weight Gain While on a High-Fat Diet. International Journal of Molecular Sciences. 2021; 22(20):11010. https://doi.org/10.3390/ijms222011010
Chicago/Turabian StyleTorres, Daniel J., Matthew W. Pitts, Lucia A. Seale, Ann C. Hashimoto, Katlyn J. An, Ashley N. Hanato, Katherine W. Hui, Stella Maris A. Remigio, Bradley A. Carlson, Dolph L. Hatfield, and et al. 2021. "Female Mice with Selenocysteine tRNA Deletion in Agrp Neurons Maintain Leptin Sensitivity and Resist Weight Gain While on a High-Fat Diet" International Journal of Molecular Sciences 22, no. 20: 11010. https://doi.org/10.3390/ijms222011010
APA StyleTorres, D. J., Pitts, M. W., Seale, L. A., Hashimoto, A. C., An, K. J., Hanato, A. N., Hui, K. W., Remigio, S. M. A., Carlson, B. A., Hatfield, D. L., & Berry, M. J. (2021). Female Mice with Selenocysteine tRNA Deletion in Agrp Neurons Maintain Leptin Sensitivity and Resist Weight Gain While on a High-Fat Diet. International Journal of Molecular Sciences, 22(20), 11010. https://doi.org/10.3390/ijms222011010