
Identification of Binding Proteins for TSC22D1 Family Proteins Using Mass Spectrometry

 , ,
, ,
Abstract
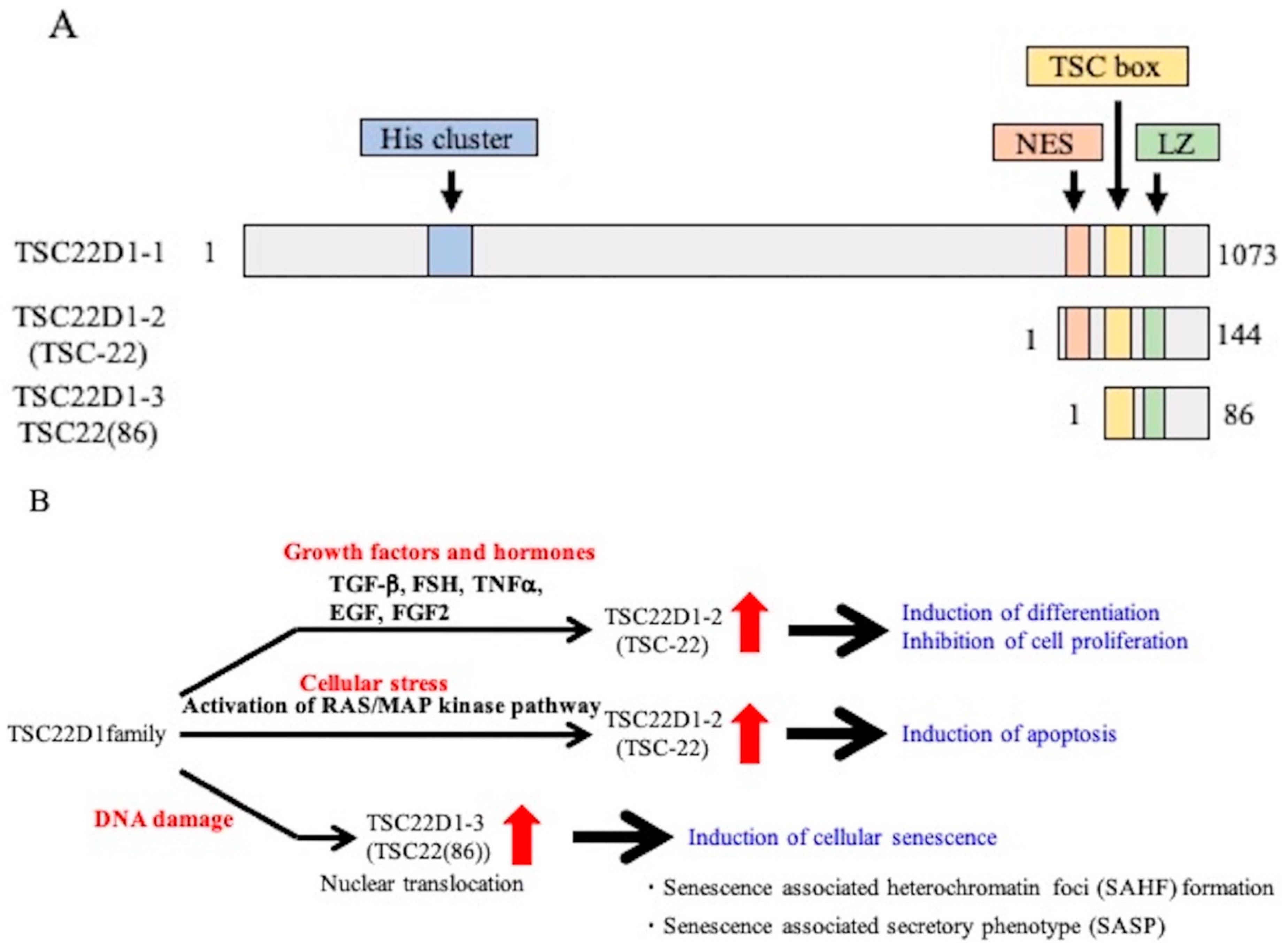
1. Introduction
2. Results
2.1. Identification of TSC-22 (TSC22D1-2)-Binding Protein by In Vitro GST Pull Down
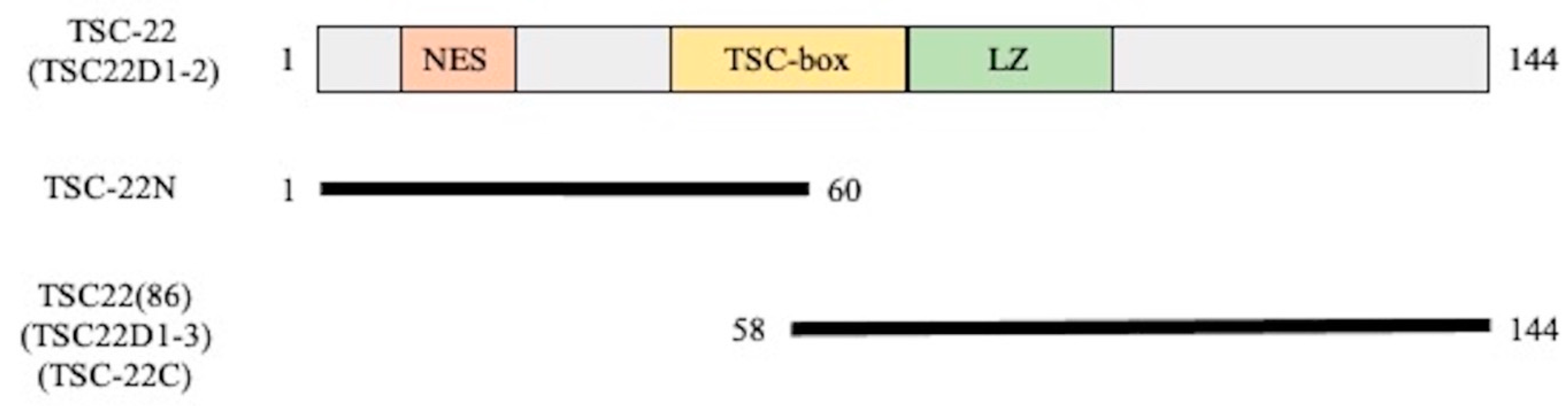
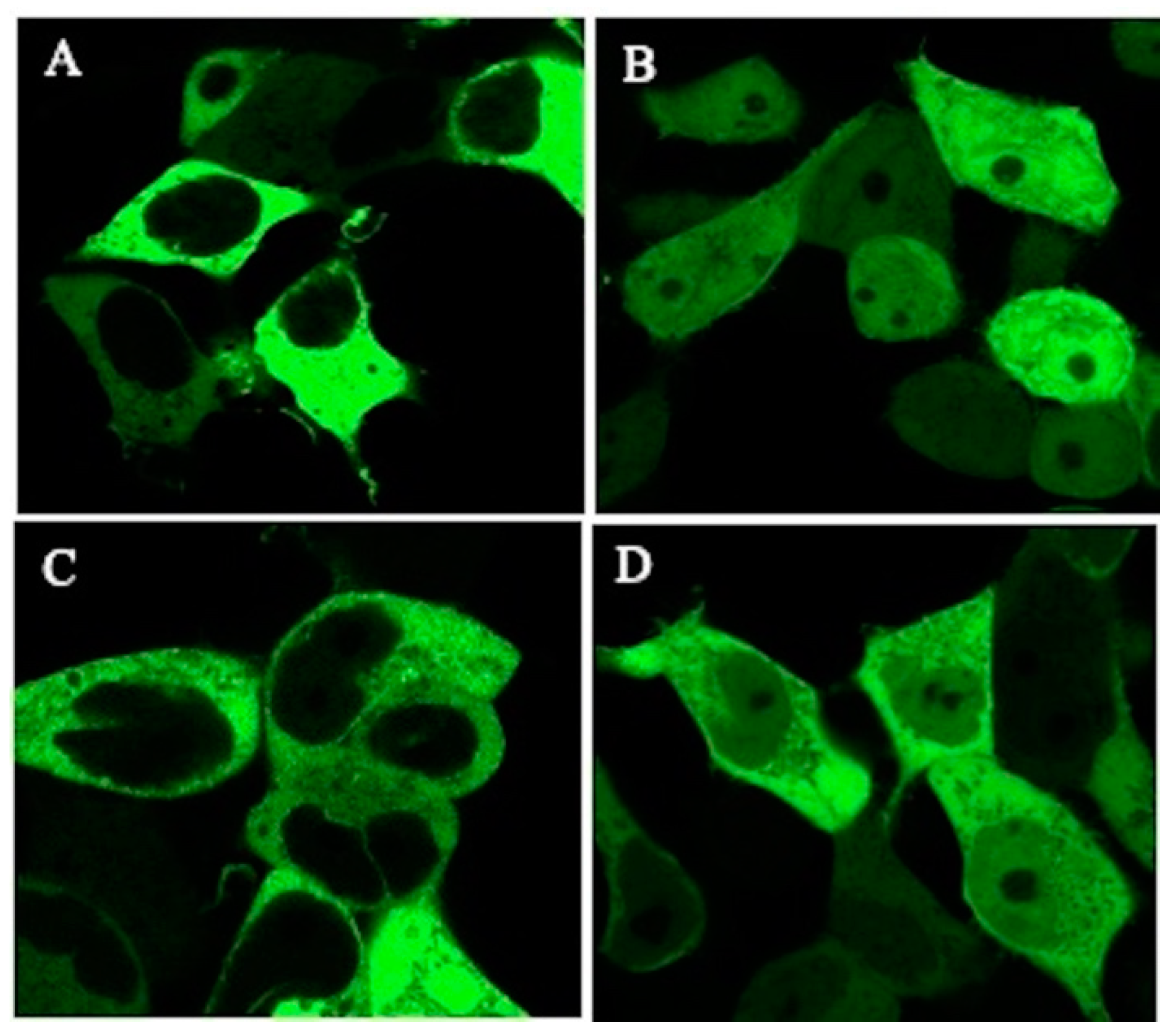
2.2. Intracellular Localization of Green Fluorescent Protein (GFP)-TSC-22 Fusion Proteins and Alterations of Location with DNA Damage
2.3. Intracellular Localization of Endogenous TSC22D1 Proteins
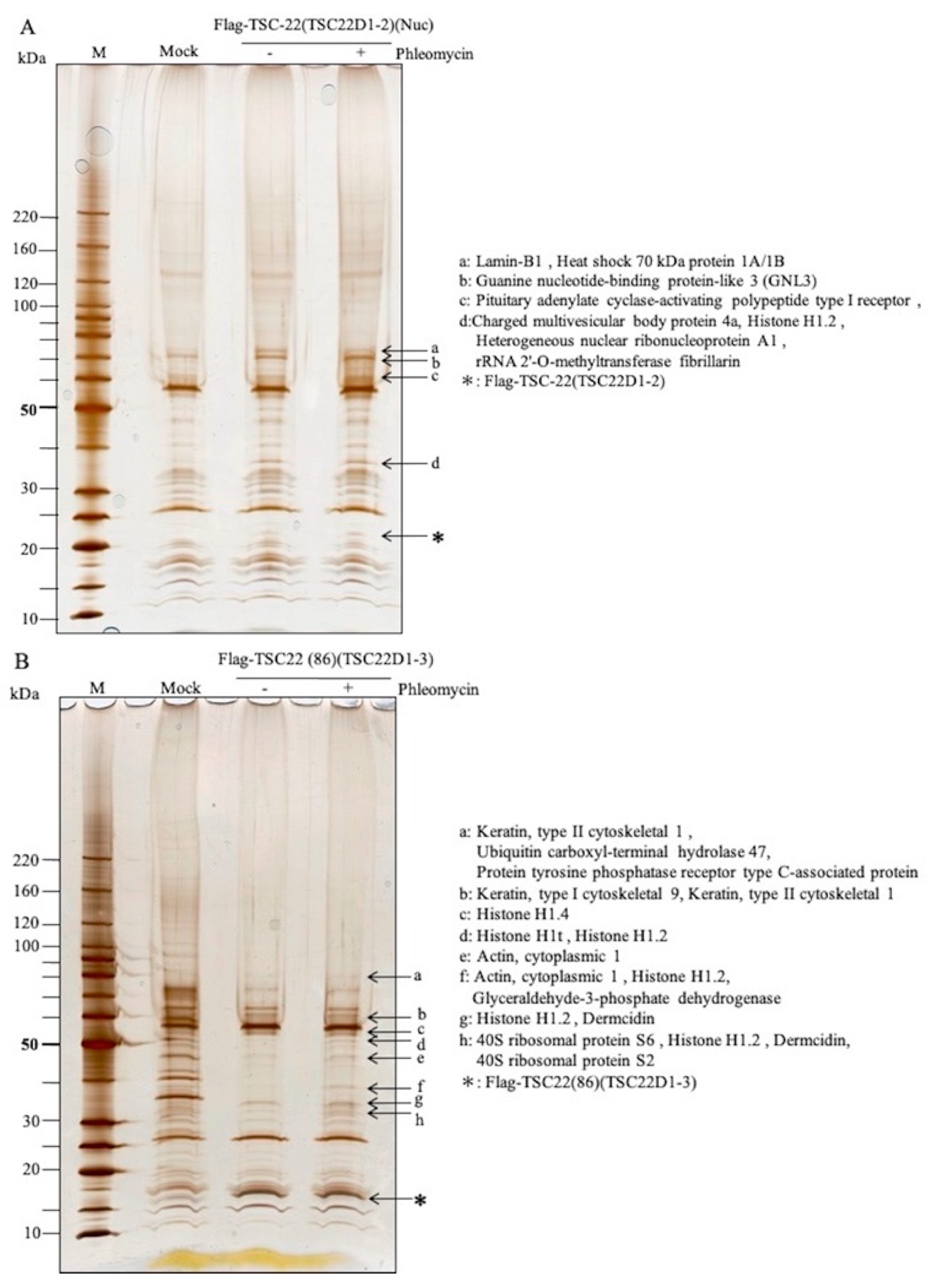
2.4. Identification of Intracellular TSC-22-Binding Proteins before and after DNA Damage
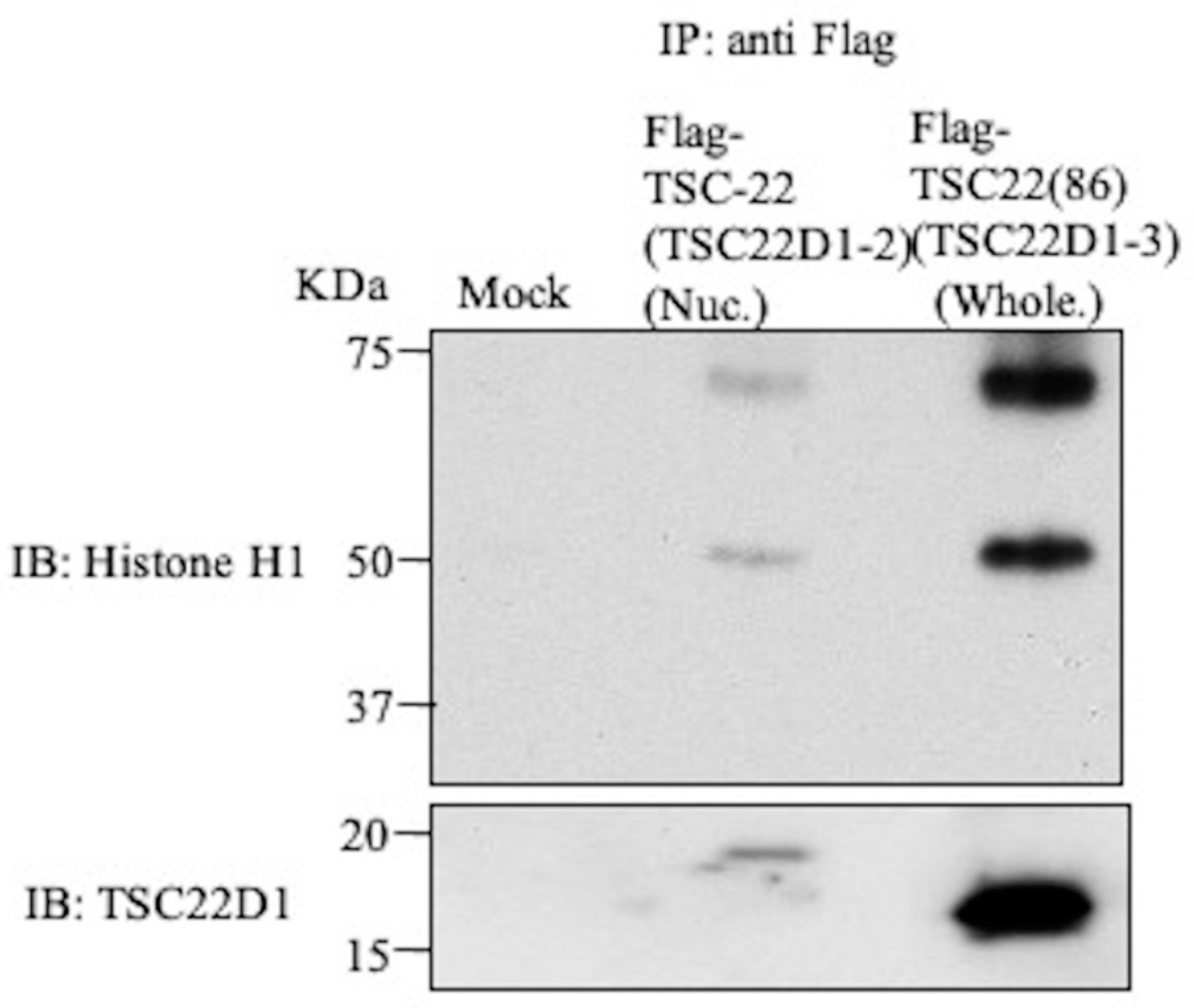
2.5. Intracellular Binding of TSC-22 (TSC22D1-2) with Histone H1 or GNL3
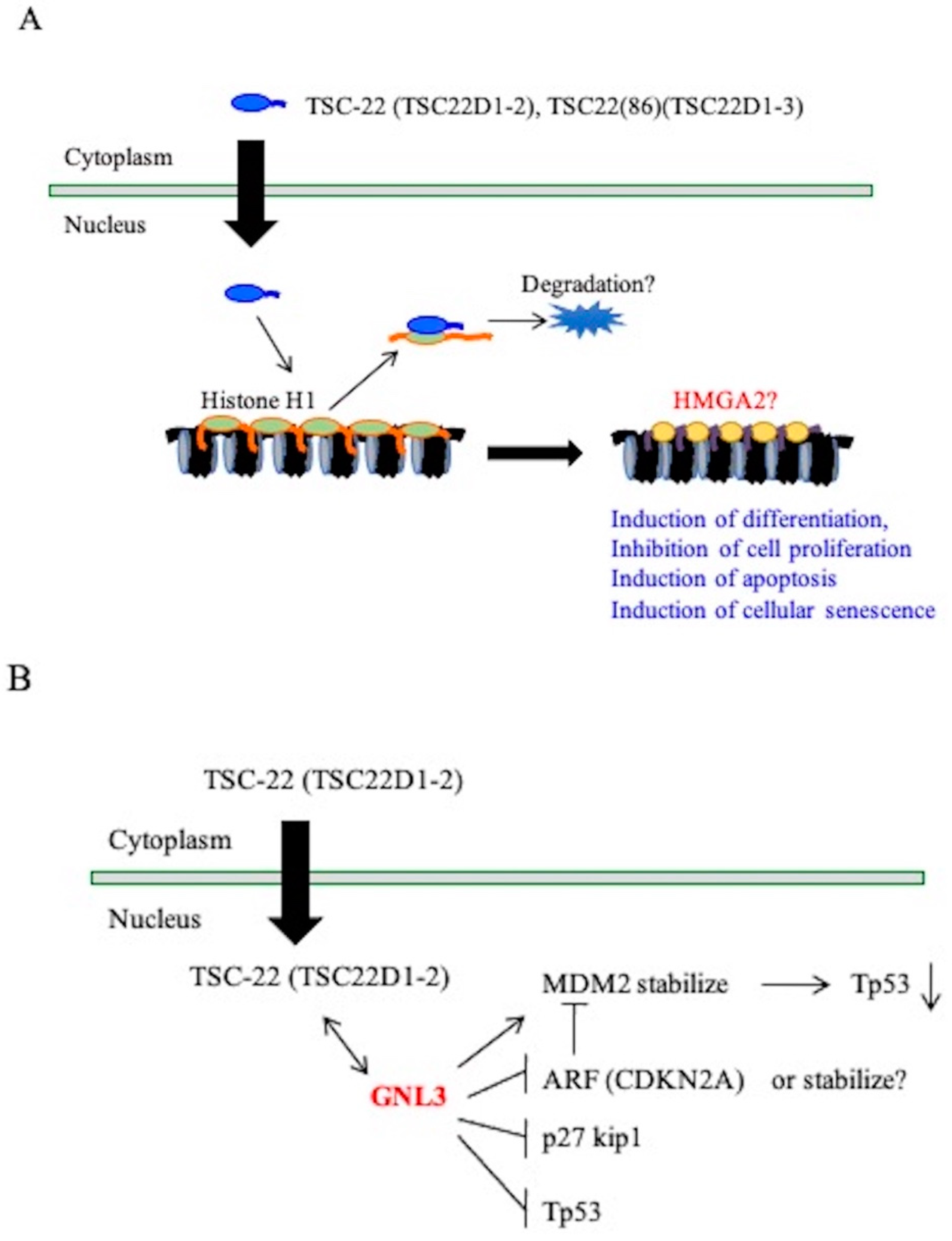
3. Discussion
4. Material and Methods
4.1. Cell Lines and Cell Culture
4.2. Construction of the Expression Vector
4.2.1. pEGFP-TSC-22 Fusion Protein Expression Vector
4.2.2. Flag-TSC-22 and Flag-TSC22(86) Fusion Protein Expression Vectors
4.3. Determination of the TSC-22 (TSC22D1-2)-Binding Protein by In Vitro GST Pull Down
4.4. Mass Spectrometry
4.5. Transfection
4.6. Intracellular Localization of the TSC-22 Fusion Protein
4.7. Localization of Endogenous TSC-22
4.8. Identification of Binding Proteins before and after DNA Damage
4.9. Immunoprecipitation-Western Blotting
4.10. Western Blotting Using Mitochondrial and Nuclear Fractions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fukumoto, C.; Ogisawa, S.; Tani, M.; Hyodo, T.; Kamimura, R.; Sawatani, Y.; Hasegawa, T.; Komiyama, Y.; Fujita, A.; Haruyama, Y.; et al. Clinical characteristics, Treatment methods and Prognosis of Oral Squamous Cell Carcinoma in Patients Aged 85 Years or Older. BMC Geriatr. 2020, 20, 487. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, C.; Sawatani, Y.; Komiyama, Y.; Shimura, M.; Hasegawa, T.; Izumi, S.; Fujita, A.; Wakui, T.; Kawamata, H. Effectiveness of cetuximab as preemptive postsurgical therapy for oral squamous cell carcinoma patients with major risk: A single-center retrospective cohort study. Investig. New Drugs 2021, 39, 846–852. [Google Scholar] [CrossRef]
- Tomita, A.; Kiyoi, H.; Naoe, T. Mechanisms of action and resistance to all-trans retinoic acid (ATRA) and arsenic trioxide (As2O3) in acute promyelocytic leukemia. Int. J. Hematol. 2013, 97, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Stahl, M.; Tallman, M.S. Acute promyelocytic leukemia (APL): Remaining challenges towards a cure for all. Leuk Lymphoma 2019, 60, 3107–3115. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.E. Differentiation, differentiation/gene therapy and cancer. Pharmacol. Ther. 1997, 73, 51–65. [Google Scholar] [CrossRef]
- Xu, W.P.; Zhang, X.; Xie, W.F. Differentiation therapy for solid tumors. J. Dig. Dis. 2014, 15, 159–165. [Google Scholar] [CrossRef]
- De Thé, H. Differentiation therapy revisited. Nat. Rev. Cancer 2018, 18, 117–127. [Google Scholar] [CrossRef]
- Kawamata, H.; Nakashiro, K.; Uchida, D.; Hino, S.; Omotehara, F.; Yoshida, H.; Sato, M. Induction of TSC-22 by treatment with a new anti-cancer drug, vesnarinone, in a human salivary gland cancer cell. Br. J. Cancer 1998, 77, 71–78. [Google Scholar] [CrossRef][Green Version]
- Nakashiro, K.; Kawamata, H.; Hino, S.; Uchida, D.; Miwa, Y.; Hamano, H.; Omotehara, F.; Yoshida, H.; Sato, M. Down-regulation of TSC-22 (Transforming Growth Factor β-stimulated Clone 22) Markedly Enhances the Growth of a Human Salivary Gland Cancer Cell Line in Vitro and in Vivo. Cancer Res. 1998, 58, 549–555. [Google Scholar]
- Uchida, D.; Kawamata, H.; Omotehara, F.; Miwa, Y.; Hino, S.; Begum, N.M.; Yoshida, H.; Sato, M. Over-Expression of TSC-22 (TGF-β Stimulated Clone-22) Markedly Enhances 5-Fluorouracil-Induced Apoptosis in a Human Salivary Gland Cancer Cell Line. Lab. Investig. 2000, 80, 955–963. [Google Scholar] [CrossRef][Green Version]
- Hino, S.; Kawamata, H.; Uchida, D.; Omotehara, F.; Miwa, Y.; Begum, N.M.; Yoshida, H.; Fujimori, T.; Sato, M. Nuclear Translocation of TSC-22 (TGF-β-Stimulated Clone-22) Concomitant with Apoptosis: TSC-22 as a Putative Transcriptional Regulator. Biochem. Biophys Res. Commun. 2000, 278, 659–664. [Google Scholar] [CrossRef]
- Hino, S.; Kawamata, H.; Omotehara, F.; Uchida, D.; Miwa, Y.; Begum, N.M.; Yoshida, H.; Sato, M.; Fujimori, T. Cytoplasmic TSC-22 (Transforming Growth Factor-β- Stimulated Clone-22) Markedly Enhances the Radiation Sensitivity of Salivary Gland Cancer Cells. Biochem. Biophys Res. Commun. 2002, 292, 957–963. [Google Scholar] [CrossRef]
- Uchida, D.; Omotehara, F.; Nakashiro, K.; Tateishi, Y.; Hino, S.; Begum, N.M.; Fujimori, T.; Kawamata, H. Posttranscriptional regulation of TSC-22 (TGF-β-stimulated clone-22) gene by TGF-β1. Biochem. Biophys Res. Commun. 2003, 305, 846–854. [Google Scholar] [CrossRef]
- Uchida, D.; Kawamata, H.; Omotehara, F.; Miwa, Y.; Horiuchi, H.; Furihata, T.; Tachibana, M.; Fujimori, T. Overexpression of TSC-22 (transforming growth factor- β-stimulated clone-22) causes marked obesity, splenic abnormality and B cell lymphoma in transgenic mice. Oncotarget 2016, 7, 14310–14323. [Google Scholar] [CrossRef] [PubMed]
- Pépin, A.; Biola-Vidamment, A.; de Laté, P.L.; Espinasse, M.A.; Godot, V.; Pallardy, M. Les protéines de la famille TSC-22D. Med. Sci. 2015, 31, 75–83. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fiol, D.F.; Mak, S.K.; Kültz, D. Specific TSC22 domain transcripts are hypertonically induced and alternatively spliced to protect mouse kidney cells during osmotic stress. FEBS J. 2007, 274, 9–24. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 24 April 2021).
- Shibanuma, M.; Kuroki, T.; Nose, K. Isolation of a gene encoding a putative leucine zipper structure that is induced by transforming growth factor beta 1 and other growth factors. J. Biol. Chem. 1992, 267, 10219–10224. [Google Scholar] [CrossRef]
- Khoury, C.M.; Yang, Z.; Li, X.Y.; Vignali, M.; Fields, S.; Greenwood, M.T.A. TSC22-like motif defines a novel antiapoptotic protein family. FEMS Yeast Res. 2008, 8, 540–563. [Google Scholar] [CrossRef]
- Hömig-Hölzel, C.; van Doorn, R.; Vogel, C.; Germann, M.; Cecchini, M.G.; Verdegaal, E.; Peeper, D.S. Antagonistic TSC22D1 variants control BRAF (E600)-induced senescence. EMBO J. 2011, 30, 1753–1765. [Google Scholar] [CrossRef] [PubMed]
- Soundararajan, R.; Wang, J.; Melters, D.; Pearce, D. Differential Activities of Glucocorticoid-induced Leucine Zipper Protein Isoforms. J. Biol Chem. 2007, 282, 36303–36313. [Google Scholar] [CrossRef] [PubMed]
- D’Adamio, F.; Zollo, O.; Moraca, R.; Ayroldi, E.; Bruscoli, S.; Bartoli, A.; Cannarile, L.; Migliorati, G.; Riccardi, C. A New Dexamethasone-Induced Gene of the Leucine Zipper Family Protects T Lymphocytes from TCR/CD3-Activated Cell Death. Immunity 1997, 7, 803–812. [Google Scholar] [CrossRef]
- Bruscoli, S.; Donato, V.; Velardi, E.; Di Sante, M.; Migliorati, G.; Donato, R.; Riccardi, C. Glucocorticoid-induced Leucine Zipper (GILZ) and Long GILZ Inhibit Myogenic Differentiation and Mediate Anti-myogenic Effects of Glucocorticoids. J. Biol Chem. 2010, 285, 10385–10396. [Google Scholar] [CrossRef] [PubMed]
- Kester, H.A.; Blanchetot, C.; den Hertog, J.; van der Saag, P.T.; van der Burg, B. Transforming Growth Factor-β-stimulated Clone-22 Is a Member of a Family of Leucine Zipper Proteins That Can Homo- and Heterodimerize and Has Transcriptional Repressor Activity. J. Biol. Chem. 1999, 274, 27439–27447. [Google Scholar] [CrossRef] [PubMed]
- Liang, F.; Li, Q.; Li, X.; Li, Z.; Gong, Z.; Deng, H.; Xiang, B.; Zhou, M.; Li, X.; Li, G.; et al. TSC22D2 interacts with PKM2 and inhibits cell growth in colorectal cancer. Int. J. Oncol. 2016, 49, 1046–1056. [Google Scholar] [CrossRef] [PubMed]
- Venter, J.C.; Adams, M.D.; Myers, E.W.; Li, P.W.; Mural, R.J.; Sutton, G.G.; Smith, H.O.; Yandell, M.; Evans, C.A.; Holt, R.A.; et al. The Sequence of the Human Genome. Science 2001, 291, 1304–1351. [Google Scholar] [CrossRef]
- Kanda, A.; Hirose, I.; Noda, K.; Murata, M.; Ishida, S. Glucocorticoid-transactivated TSC22D3 attenuates hypoxia- and diabetes-induced Müller glial galectin-1 expression via HIF- 1α destabilization. J. Cell Mol. Med. 2020, 24, 4589–4599. [Google Scholar] [CrossRef]
- Sugawara, F.; Yamada, Y.; Kuroe, A.; Someya, Y.; Kubota, A.; Ihara, Y.; Takahashi, K.; Seino, Y. Human TSC-22 gene: No association with type 2 diabetes. Intern. Med. 2001, 40, 993–997. [Google Scholar] [CrossRef][Green Version]
- Vargas, D.M.; De Bastiani, M.A.; Zimmer, E.R.; Klamt, F. Alzheimer’s disease master regulators analysis: Search for potential molecular targets and drug repositioning candidates. Alzheimers Res. Ther. 2018, 10, 1–12. [Google Scholar] [CrossRef]
- Koike, M.; Ninomiya, Y.; Koike, A. Characterization of Ninjurin and TSC22 induction after X-irradiation of normal human skin cells. J. Dermatol. 2008, 35, 6–17. [Google Scholar] [CrossRef]
- Yoon, C.H.; Rho, S.B.; Kim, S.T.; Kho, S.; Park, J.; Jang, I.S.; Woo, S.; Kim, S.S.; Lee, J.H.; Lee, S.H. Crucial role of TSC-22 in preventing the proteasomal degradation of p53 in cervical cancer. PLoS ONE 2012, 7, e42006. [Google Scholar] [CrossRef]
- Hashiguchi, A.; Hitachi, K.; Inui, M.; Okabayashi, K.; Asashima, M. TSC-box is essential for the nuclear localization and antiproliferative effect of XTSC-22. Dev. Growth Differ. 2007, 49, 197–204. [Google Scholar] [CrossRef]
- Zheng, L.; Suzuki, H.; Nakajo, Y.; Nakano, A.; Kato, M. Regulation of c-MYC transcriptional activity by transforming growth factor-beta 1-stimulated clone 22. Cancer Sci. 2018, 109, 395–402. [Google Scholar] [CrossRef]
- Sprenger, C.C.; Haugk, K.; Sun, S.; Coleman, I.; Nelson, P.S.; Vessella, R.L.; Ludwig, D.L.; Wu, J.D.; Plymate, S.R. Transforming Growth Factor-{beta}-Stimulated Clone-22 Is an Androgen-Regulated Gene That Enhances Apoptosis in Prostate Cancer following Insulin-Like Growth Factor-I Receptor Inhibition. Clin. Cancer Res. 2009, 15, 7634–7641. [Google Scholar] [CrossRef]
- Cho, M.-J.; Lee, J.-Y.; Shin, M.-G.; Kim, H.-J.; Choi, Y.-J.; Rho, S.B.; Kim, B.-R.; Jang, I.S.; Lee, S.-H. TSC-22 inhibits CSF-1R function and induces apoptosis in cervical cancer. Oncotarget 2017, 8, 97990–98003. [Google Scholar] [CrossRef]
- Pépin, A.; Espinasse, M.A.; de Laté, P.L.; Szely, N.; Pallardy, M.; Biola-Vidamment, A. TSC-22 Promotes Interleukin-2-Deprivation Induced Apoptosis in T-Lymphocytes. J. Cell Biochem. 2016, 117, 1855–1868. [Google Scholar] [CrossRef] [PubMed]
- Huser, C.A.; Pringle, M.A.; Heath, V.J.; Bell, A.K.; Kendrick, H.; Smalley, M.J.; Crighton, D.; Ryan, K.M.; A Gusterson, B.A.; Stein, T. TSC-22D1 isoforms have opposing roles in mammary epithelial cell survival. Cell Death Differ. 2010, 17, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Kitaura, J.; Enomoto, Y.; Lu, Y.; Nishimura, K.; Isobe, M.; Ozaki, K.; Komeno, Y.; Nakahara, F.; Oki, T.; et al. Transforming growth factor-β-stimulated clone-22 is a negative-feedback regulator of Ras ⁄ Raf signaling: Implications for tumorigenesis. Cancer Sci. 2012, 103, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Dragotto, J.; Canterini, S.; Del Porto, P.; Bevllacqua, A.; Fiorenza, M.T. The interplay between TGF-β-stimulated TSC22 domain family proteins regulates cell-cycle dynamics in medulloblastoma cells. J. Cell Physiol. 2019, 234, 18349–18360. [Google Scholar] [CrossRef]
- Bayona-Feliu, A.; Casas-Lamesa, A.; Reina, O.; Bernués, J.; Azorín, F. Linker histone H1 prevents R-loop accumulation and genome instability in heterochromatin. Nat. Commun. 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Thorslund, T.; Ripplinger, A.; Hoffmann, S.; Wild, T.; Uckelmann, M.; Villumsen, B.; Narita, T.; Sixma, T.K.; Choudhary, C.; Bekker-Jensen, S.; et al. Histone H1 couples initiation and amplification of ubiquitin signalling after DNA damage. Nature 2015, 527, 389–393. [Google Scholar] [CrossRef]
- Yang, Z.; Sun, J.; Hu, Y.; Wang, F.; Wang, X.; Qiao, H.-H.; Xu, J.; Mao, D.; Ren, X.; Pan, L.-X.; et al. Histone H1 defect in escort cells triggers germline tumor in Drosophila ovary. Dev. Biol. 2017, 424, 40–49. [Google Scholar] [CrossRef]
- Sekeri-Pataryas, K.E.; Sourlingas, T.G. The Differentiation-Associated Linker Histone, H1.0, during the in Vitro Aging and Senescence of Human Diploid Fibroblasts. Ann. N. Y. Acad. Sci. 2007, 1100, 361–367. [Google Scholar] [CrossRef]
- Dai, M.S.; Sun, X.X.; Lu, H. Aberrant Expression of Nucleostemin Activates p53 and Induces Cell Cycle Arrest via Inhibition of MDM2. Mol. Cell Biol. 2008, 28, 4365–4376. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Jia, Y.; Wei, Y.; Liu, S.; Yue, B. Gene expression profiling of NB4 cells following knockdown of nucleostemin using DNA microarrays. Mol. Med. Rep. 2016, 14, 175–183. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Li, T.; LI, L.; Wu, X.; Tian, K.; Wang, Y. The oncogenic role of gnl3 in the progression and metastasis of osteosarcoma. Cancer Manag. Res. 2019, 11, 2179–2188. [Google Scholar] [CrossRef]
- Lin, T.; Lin, T.C.; McGrail, D.J.; Bhupal, P.K.; Ku, Y.H.; Zhang, W.; Meng, L.; Lin, S.Y.; Peng, G.; Tsai, R.Y.L. Nucleostemin Reveals a Dichotomous Nature of Genome Maintenance in Mammary Tumor Progression. Oncogene 2019, 38, 3919–3931. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Zhou, Y.; Chen, J.; Yang, L.; Qiu, Y.; Tu, S.; Zhong, M. Low levels of TSC22 enhance tumorigenesis by inducing cell proliferation in colorectal cancer. Biochem. Biophys. Res. Commun. 2018, 497, 1062–1067. [Google Scholar] [CrossRef]
- Tan, K.; Fujimoto, M.; Takii, R.; Takaki, E.; Hayashida, N.; Nakai, A. Mitochondrial SSBP1 protects cells from proteotoxic stresses by potentiating stress- induced HSF1 transcriptional activity. Nat. Commun. 2015, 6, 6580. [Google Scholar] [CrossRef]
- Yanagawa, T.; Hayashi, Y.; Yoshida, H.; Yura, Y.; Nagamine, S.; Bando, T.; Sato, M. An adenoid squamous carcinoma-forming cell line established from an oral keratinizing squamous cell carcinoma expressing carcinoembryonic antigen. Am. J. Pathol. 1986, 124, 496–509. [Google Scholar]
- Jensen, O.; Ansari, S.; Gebauer, L.; Müller, S.F.; Lowjaga, K.A.A.T.; Geyer, J.; Tzvetkov, M.V.; Brockmöller, J. A double-flp-in method for stable overexpression of two genes. Sci. Rep. 2020, 10, 14018. [Google Scholar] [CrossRef]
- Domanski, M.; LaCava, J. Affinity Purification of the RNA Degradation Complex, the Exosome, from HEK-293 Cells. Bio Protoc. 2017, 7, e2238. [Google Scholar] [CrossRef] [PubMed]
- Kinouchi, M.; Izumi, S.; Nakashiro, K.; Haruyama, Y.; Kobashi, G.; Uchida, D.; Hasegawa, T.; Kawamata, H. Determination of the origin of oral squamous cell carcinoma by microarray analysis: Squamous epithelium or minor salivary gland? Int. J. Cancer 2018, 143, 2551–2560. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Transcript or Protein Name (NCBI Accession Number) | Other Names | Protein Length (Number of Amino Acids) | Reference |
|---|---|---|---|---|
| TSC22D1 | TSC22D1-1(NP_904358.2) | TSC22D1-X1 | 1073 | [15,16,17] |
| TSC22D1-2 (NP_006013.1) | TSC-22/TSC22D1-CRA_a | 144 | [8,9,10,11,12,13,14,15,17,18] | |
| TSC22D1-3 (NP_001230727.1) | TSC22 (86) | 86 | [17,19] | |
| TSC22D1-4 (NP_001230728.1) | 570 | [17,19,20] | ||
| TSC22D1-X2 (XP_016876299.1) | 1048 | [17] | ||
| TSC22D1-X3 (XP_005266640.1) | 1028 | [17] | ||
| TSC22D1-X4 (XP_016876300.1) | 985 | [17] | ||
| TSC22D2 | TSC22D2-1 (NP_055594.1) | TSC22D2-CRA_b | 780 | [15,16,17,19] |
| TSC22D2-2 (NP_001290193.1) | 756 | [17,25] | ||
| TSC22D2-X1 (XP_011511639.1) | 691 | [17] | ||
| TSC22D2-CRA_a (EAW78837.1) | 690 | [17,26] | ||
| TSC22D2-CRA_c (EAW78840.1) | 753 | [17,26] | ||
| TSC22D2-CRA_d (EAW78841.1) | 124 | [17,26] | ||
| TSC22D3 | TSC22D3-1 (NP_932174.1) | TSC22D2-X1 TSC22D2-CRA_a | 200 | [16,17,19,20] |
| TSC22D3-2 (NP_004080.2) | GILZ/T SC22D2-CRA_c | 134 | [15,17,21,22] | |
| TSC22D3-3 (NP_001015881.1) | 77 | [17,27] | ||
| TSC22D3-CRA_b (EAX02707.1) | 193 | [17,26] | ||
| Long-GILZ (ACJ09091.1) | 234 | [17,22] | ||
| TSC22D4 | TSC22D4-1 (NP_001289972.1) | THG-1 / TSC22D4-a | 395 | [17,23,24] |
| TSC22D4-2 | 310 | [15,19] | ||
| TSC22D4-3 | 195 | [15,19] |
| GST-TSC-22 Pull-Down Assay (TYS, Whole Cell Extracts) | In Vivo Flag-TSC-22 Binding Assay(HEK293, Nuclear Extracts) | In Vivo Flag-TSC22 (86) Binding Assay (HEK293, Whole Cell Extracts) | |
|---|---|---|---|
| Chromatin structure | Histone H1.2 (P16403) Lamin-B1 (P20700) | Histone H1.2 (P16403), Histone H1.4 (P10412) Histone H1t (P22492) | |
| Cell growth, Apoptosis, differentiation | Guanine nucleotide-binding protein-like 3 (Q9BVP2) | 40S ribosomal protein S6 (P62753) | |
| Signal transduction | Pituitary adenylate cyclase-activating polypeptide type I receptor (P41586) | ||
| Chaperone | Heat shock 70-kDa protein 1A/1B (P0DMV8/P0DMV9) | Heat shock 70-kDa protein 1A/1B (P0DMV8/P0DMV9) | |
| Cytoskeleton | Keratin, type II cytoskeletal 1 (P04264) Rho-related GTP-binding protein Rho6 (Q92730) Pleckstrin homology domain-containing family G member 2 (Q9H7P9) | Keratin, type II cytoskeletal 1 (P04264) Keratin, type I cytoskeletal 9 (P35527) Actin, cytoplasmic 1 (P60709) | |
| Others | Elongation factor 1-gamma (P26641) Carbonyl reductase [NADPH] 1 (P16152) | Heterogeneous nuclear ribonucleoprotein A1 (P09651) Charged multivesicular body protein 4a (Q9BY43) rRNA 2′-O-methyltransferase fibrillarin (P22087) | Glyceraldehyde-3-phosphate dehydrogenase (P04406) Ubiquitin carboxyl-terminal hydrolase 47 (Q96K76) Dermcidin (P81605) Protein tyrosine phosphatase receptor type C-associated protein (Q14761) 40S ribosomal protein S2 (P15880) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamimura, R.; Uchida, D.; Kanno, S.-i.; Shiraishi, R.; Hyodo, T.; Sawatani, Y.; Shimura, M.; Hasegawa, T.; Tsubura-Okubo, M.; Yaguchi, E.; et al. Identification of Binding Proteins for TSC22D1 Family Proteins Using Mass Spectrometry. Int. J. Mol. Sci. 2021, 22, 10913. https://doi.org/10.3390/ijms222010913
Kamimura R, Uchida D, Kanno S-i, Shiraishi R, Hyodo T, Sawatani Y, Shimura M, Hasegawa T, Tsubura-Okubo M, Yaguchi E, et al. Identification of Binding Proteins for TSC22D1 Family Proteins Using Mass Spectrometry. International Journal of Molecular Sciences. 2021; 22(20):10913. https://doi.org/10.3390/ijms222010913
Chicago/Turabian StyleKamimura, Ryouta, Daisuke Uchida, Shin-ichiro Kanno, Ryo Shiraishi, Toshiki Hyodo, Yuta Sawatani, Michiko Shimura, Tomonori Hasegawa, Maki Tsubura-Okubo, Erika Yaguchi, and et al. 2021. "Identification of Binding Proteins for TSC22D1 Family Proteins Using Mass Spectrometry" International Journal of Molecular Sciences 22, no. 20: 10913. https://doi.org/10.3390/ijms222010913
APA StyleKamimura, R., Uchida, D., Kanno, S.-i., Shiraishi, R., Hyodo, T., Sawatani, Y., Shimura, M., Hasegawa, T., Tsubura-Okubo, M., Yaguchi, E., Komiyama, Y., Fukumoto, C., Izumi, S., Fujita, A., Wakui, T., & Kawamata, H. (2021). Identification of Binding Proteins for TSC22D1 Family Proteins Using Mass Spectrometry. International Journal of Molecular Sciences, 22(20), 10913. https://doi.org/10.3390/ijms222010913