Genetic Variations in Prostaglandin E2 Pathway Identified as Susceptibility Biomarkers for Gastric Cancer in an Intermediate Risk European Country

 ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. Study Population
2.2. Genotype Frequencies and Risk Estimates
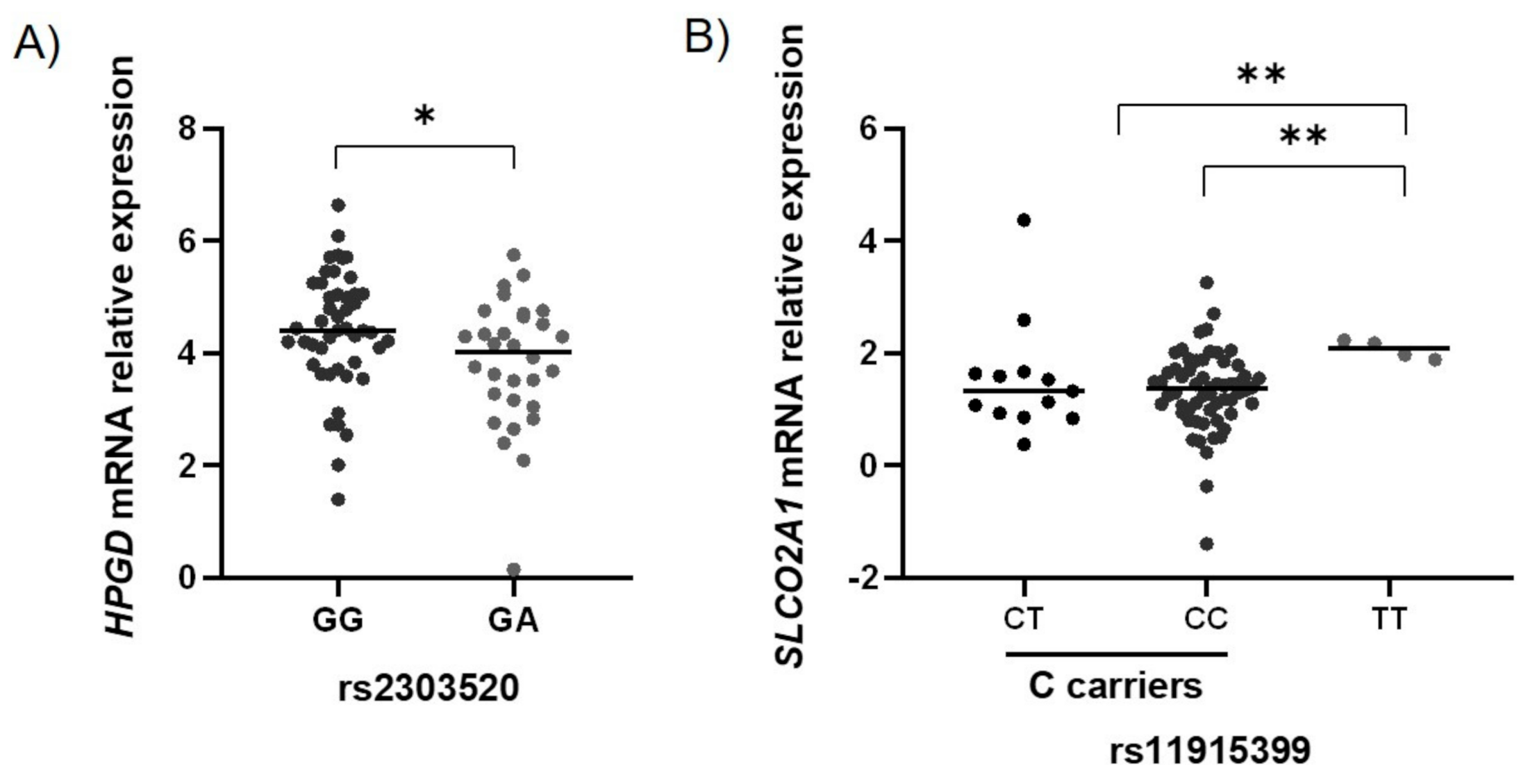
2.3. Functional Characterization of the GC-Associated Biomarkers
2.4. Haplotype Analysis
2.5. Gene-“Environment” Interaction Analysis
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Sample Collection and Processing
4.3. Genetic Polymorphisms Selection and Characterization
4.4. Reverse Transcription Reaction
4.5. Real-Time PCR
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 15-PGDH | 15-hydroxyprostaglandin dehydrogenase |
| ABCC4 | ATP binding cassette subfamily c member 4 |
| B2M | beta-2-microglobulin |
| CASP8 | caspase 8 |
| cDNA | complementary deoxyribonucleic acid |
| CEU | Utah residents with Northern and Western European ancestry |
| CI | confidence interval |
| CT | cycle threshold |
| CVC | cross-variation consistency |
| EM | expectation-maximization algorithm |
| FDR | false discovery rate |
| FFPE | formalin-fixed paraffin-embedded |
| GC | gastric cancer |
| GSTP1 | glutathione s-transferase pi 1 |
| GUSB | glucuronidase beta |
| HPGD | 15-hydroxyprostaglandin dehydrogenase |
| HPRT1 | hypoxanthine phosphoribosyltransferase 1 |
| HWE | Hardy-Weinberg equilibrium |
| IPO8 | importin 8 |
| MDR | multifactor dimensionality reduction |
| mRNA | messenger ribonucleic acid |
| MRP4 | multidrug resistance protein 4 |
| MTX1 | metaxin 1 |
| MUC1 | mucin 1, cell surface-associated |
| OD | optical density |
| PCR | polymerase chain reaction |
| PGE2 | prostaglandin E2 |
| PGT | prostaglandin transporter |
| PKLR | pyruvate kinase L/R |
| PLCE1 | phospholipase C epsilon 1 |
| PPIA | peptidylprolyl isomerase A |
| PRKAA1 | protein kinase AMP-activated catalytic subunit alpha |
| PSCA | prostate stem cell antigen |
| PTGS2 | prostaglandin-endoperoxide synthase 2 |
| RPL29 | ribosomal protein L29 |
| RT | reverse transcription |
| SLCO2A1 | solute carrier organic anion transporter family member 2A1 |
| SPSS | statistical package for the social sciences |
| TGFBR2 | transforming growth factor-beta receptor 2 |
| TNF | tumor necrosis factor-alpha |
References
- Gao, Y.; Zhang, K.; Xi, H.; Cai, A.; Wu, X.; Cui, J.; Li, J.; Qiao, Z.; Wei, B.; Chen, L. Diagnostic and prognostic value of circulating tumor DNA in gastric cancer: A meta-analysis. Oncotarget 2016, 8, 6330–6340. [Google Scholar] [CrossRef] [PubMed]
- Maconi, G.; Manes, G.; Porro, G.-B. Role of symptoms in diagnosis and outcome of gastric cancer. World J. Gastroenterol. 2008, 14, 1149–1155. [Google Scholar] [CrossRef] [PubMed]
- Areia, M.; Spaander, M.C.; Kuipers, E.J.; Dinis-Ribeiro, M. Endoscopic screening for gastric cancer: A cost-utility analysis for countries with an intermediate gastric cancer risk. United Eur. Gastroenterol. J. 2018, 6, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; DuBois, R.N. Eicosanoids and cancer. Nat. Rev. Cancer 2010, 10, 181–193. [Google Scholar] [CrossRef]
- Greenhough, A.; Smartt, H.J.M.; Moore, A.E.; Roberts, H.R.; Williams, A.C.; Paraskeva, C.; Kaidi, A. The COX-2/PGE 2 pathway: Key roles in the hallmarks of cancer and adaptation to the tumour microenvironment. Carcinogenesis 2009, 30, 377–386. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Chen, E.P.; Smyth, E.M. COX-2 and PGE2-dependent immunomodulation in breast cancer. Prostaglandins Other Lipid Mediat. 2011, 96, 14–20. [Google Scholar] [CrossRef]
- Kasper, H.U.; Konze, E.; Dienes, H.P.; Stippel, D.L.; Schirmacher, P.; Kern, M. COX-2 expression and effects of COX-2 inhibition in colorectal carcinomas and their liver metastases. Anticancer Res. 2010, 30, 2017–2023. [Google Scholar]
- Mrena, J.; Wiksten, J.-P.; Kokkola, A.; Nordling, S.; Ristimäki, A.; Haglund, C. COX-2 is associated with proliferation and apoptosis markers and serves as an independent prognostic factor in gastric cancer. Tumor Biol. 2010, 31, 1–7. [Google Scholar] [CrossRef]
- Gomes, R.N.; da Costa, S.F.; Colquhoun, A. Eicosanoids and cancer. Clinics 2018, 73. [Google Scholar] [CrossRef]
- Nakanishi, T.; Tamai, I. Roles of Organic Anion Transporting Polypeptide 2A1 (OATP2A1/SLCO2A1) in Regulating the Pathophysiological Actions of Prostaglandins. AAPS J. 2017, 20, 13. [Google Scholar] [CrossRef] [PubMed]
- Mocellin, S.; Verdi, D.; Pooley, K.A.; Nitti, D. Genetic variation and gastric cancer risk: A field synopsis and meta-analysis. Gut 2015, 64, 1209–1219. [Google Scholar] [CrossRef] [PubMed]
- Lopes, C.; Pereira, C.; Farinha, M.; Medeiros, R.; Dinis-Ribeiro, M. Prostaglandin E2 pathway is dysregulated in gastric adenocarcinoma in a Caucasian population. Int. J. Mol. Sci. 2020, 21, 7680. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; Queiros, S.; Galaghar, A.; Sousa, H.; Pimentel-Nunes, P.; Brandao, C.; Moreira-Dias, L.; Medeiros, R.; Dinis-Ribeiro, M. Genetic variability in key genes in prostaglandin E2 pathway (COX-2, HPGD, ABCC4 and SLCO2A1) and their involvement in colorectal cancer development. PLoS ONE 2014, 9, e92000. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.D.; Silva, C.; Rocha, A.; Nogueira, C.; Castro-Poças, F.; Araujo, A.; Matos, E.; Pereira, C.; Medeiros, R.; Lopes, C. Predictive clinical model of tumor response after chemoradiation in rectal cancer. Oncotarget 2017, 8, 58133–58151. [Google Scholar] [CrossRef]
- Pereira, C.; Queiros, S.; Galaghar, A.; Sousa, H.; Marcos-Pinto, R.; Pimentel-Nunes, P.; Brandao, C.; Medeiros, R.; Dinis-Ribeiro, M. Influence of genetic polymorphisms in prostaglandin E2 pathway (COX-2/HPGD/SLCO2A1/ABCC4) on the risk for colorectal adenoma development and recurrence after polypectomy. Clin. Transl. Gastroenterol. 2016, 7, e191. [Google Scholar] [CrossRef]
- Luo, M.-X.; Long, B.-B.; Li, F.; Zhang, C.; Pan, M.-T.; Huang, Y.-Q.; Chen, B. Roles of Cyclooxygenase-2 gene −765G>C (rs20417) and −1195G>A (rs689466) polymorphisms in gastric cancer: A systematic review and meta-analysis. Gene 2019, 685, 125–135. [Google Scholar] [CrossRef]
- Huang, T.; Shu, Y.; Cai, Y.-D. Genetic differences among ethnic groups. BMC Genom. 2015, 16, 1093. [Google Scholar] [CrossRef]
- Spielman, R.S.; Bastone, L.A.; Burdick, J.T.; Morley, M.; Ewens, W.J.; Cheung, V.G. Common genetic variants account for differences in gene expression among ethnic groups. Nat. Genet. 2007, 39, 226–231. [Google Scholar] [CrossRef]
- Zmigrodzka, M.; Rzepecka, A.; Krzyzowska, M.; Witkowska-Pilaszewicz, O.; Cywinska, A.; Winnicka, A. The cyclooxygenase-2/prostaglandin E2 pathway and its role in the pathogenesis of human and dog hematological malignancies. J. Physiol. Pharmacol. 2018, 69. [Google Scholar] [CrossRef]
- Hashemi Goradel, N.; Najafi, M.; Salehi, E.; Farhood, B.; Mortezaee, K. Cyclooxygenase-2 in cancer: A review. J. Cell. Physiol. 2019, 234, 5683–5699. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Omori, K.; Murata, T. Role of prostaglandins in tumor microenvironment. Cancer Metastasis Rev. 2018, 37, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, M.; Rosenberg, D.W. Multifaceted roles of PGE2 in inflammation and cancer. Semin. Immunopathol. 2013, 35, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.J.; Gagnon-Bartsch, J.A.; Tan, I.B.; Earle, S.; Ruff, L.; Pettinger, K.; Ylstra, B.; van Grieken, N.; Rha, S.Y.; Chung, H.C.; et al. Signatures of tumour immunity distinguish Asian and non-Asian gastric adenocarcinomas. Gut 2015, 64, 1721–1731. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, T.; Tohnai, N. Cyclooxygenase isozymes and their gene structures and expression. Prostaglandins Other Lipid Mediat. 2002, 68–69, 95–114. [Google Scholar] [CrossRef]
- Pereira, C.; Sousa, H.; Silva, J.; Brandao, C.; Elgueta-Karstegl, C.; Farrell, P.J.; Medeiros, R.; Dinis-Ribeiro, M. The -1195G allele increases the transcriptional activity of cyclooxygenase-2 gene (COX-2) in colon cancer cell lines. Mol. Carcinog. 2014, 53 (Suppl. 1), E92–E95. [Google Scholar] [CrossRef]
- Zhang, Y.-C.; Zhao, H.; Chen, C.; Ali, M.A. COX-2 gene rs689466 polymorphism is associated with increased risk of colorectal cancer among Caucasians: A meta-analysis. World J. Surg. Oncol. 2020, 18, 192. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, H.; Liu, T.; Tang, W.; Ma, Z. Cyclooxygenase-2 -1195G>A (rs689466) polymorphism and cancer susceptibility: An updated meta-analysis involving 50,672 subjects. Int. J. Clin. Exp. Med. 2015, 8, 12448–12462. [Google Scholar]
- Zamudio, R.; Pereira, L.; Rocha, C.D.; Berg, D.E.; Muniz-Queiroz, T.; Sant Anna, H.P.; Cabrera, L.; Combe, J.M.; Herrera, P.; Jahuira, M.H.; et al. Population, epidemiological, and functional genetics of gastric cancer candidate genes in Peruvians with predominant Amerindian ancestry. Dig. Dis. Sci. 2016, 61, 107–116. [Google Scholar] [CrossRef]
- Rigau, M.; Juan, D.; Valencia, A.; Rico, D. Intronic CNVs and gene expression variation in human populations. PLoS Genet. 2019, 15, e1007902. [Google Scholar] [CrossRef]
- Lomelin, D.; Jorgenson, E.; Risch, N. Human genetic variation recognizes functional elements in noncoding sequence. Genome Res. 2010, 20, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.D.; Van Steen, K. The search for gene-gene interactions in genome-wide association studies: Challenges in abundance of methods, practical considerations, and biological interpretation. Ann. Transl. Med. 2018, 6, 157. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.-Y.; Braunschweig, T.; Hewitt, S.M. Optimization of recovery of RNA from formalin-fixed, paraffin-embedded tissue. Diagn. Mol. Pathol. 2006, 15, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.-Y.; Braunschweig, T.; Williams, R.; Guerrero, N.; Hoffmann, K.M.; Kwon, M.; Song, Y.K.; Libutti, S.K.; Hewitt, S.M. Factors in tissue handling and processing that impact RNA obtained from formalin-fixed, paraffin-embedded tissue. J. Histochem. Cytochem. 2008, 56, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Antonov, J.; Goldstein, D.R.; Oberli, A.; Baltzer, A.; Pirotta, M.; Fleischmann, A.; Altermatt, H.J.; Jaggi, R. Reliable gene expression measurements from degraded RNA by quantitative real-time PCR depend on short amplicons and a proper normalization. Lab. Investig. 2005, 85, 1040–1050. [Google Scholar] [CrossRef]
- Walter, R.F.H.; Mairinger, F.D.; Wohlschlaeger, J.; Worm, K.; Ting, S.; Vollbrecht, C.; Schmid, K.W.; Hager, T. FFPE tissue as a feasible source for gene expression analysis—A comparison of three reference genes and one tumor marker. Pathol. Res. Pract. 2013, 209, 784–789. [Google Scholar] [CrossRef]
- Mueller, J.C.; Lõhmussaar, E.; Mägi, R.; Remm, M.; Bettecken, T.; Lichtner, P.; Biskup, S.; Illig, T.; Pfeufer, A.; Luedemann, J.; et al. Linkage disequilibrium patterns and tagSNP transferability among European populations. Am. J. Hum. Genet. 2005, 76, 387–398. [Google Scholar] [CrossRef][Green Version]
- Service, S.; Sabatti, C.; Freimer, N. Tag SNPs chosen from HapMap perform well in several population isolates. Genet. Epidemiol. 2007, 31, 189–194. [Google Scholar] [CrossRef]
- Ribas, G.; González-Neira, A.; Salas, A.; Milne, R.L.; Vega, A.; Carracedo, B.; González, E.; Barroso, E.; Fernández, L.P.; Yankilevich, P.; et al. Evaluating HapMap SNP data transferability in a large-scale genotyping project involving 175 cancer-associated genes. Hum. Genet. 2006, 118, 669–679. [Google Scholar] [CrossRef]
- Gu, S.; Pakstis, A.J.; Li, H.; Speed, W.C.; Kidd, J.R.; Kidd, K.K. Significant variation in haplotype block structure but conservation in tagSNP patterns among global populations. Eur. J. Hum. Genet. 2007, 15, 302–312. [Google Scholar] [CrossRef]
- de Bakker, P.I.W.; Burtt, N.P.; Graham, R.R.; Guiducci, C.; Yelensky, R.; Drake, J.A.; Bersaglieri, T.; Penney, K.L.; Butler, J.; Young, S.; et al. Transferability of tag SNPs in genetic association studies in multiple populations. Nat. Genet. 2006, 38, 1298–1303. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.B.; Edge, S.B.; Greene, F.L.; Byrd, D.R.; Brookland, R.K.; Washington, M.K.; Gershenwald, J.E.; Compton, C.C.; Hess, K.R.; Sullivan, D.C.; et al. AJCC Cancer Staging Manual, 8th ed.; Springer: Berlin/Heidelberg, Germany, 2018; p. 1032. [Google Scholar] [CrossRef]
- Pereira, C.; Sousa, H.; Ferreira, P.; Fragoso, M.; Moreira-Dias, L.; Lopes, C.; Medeiros, R.; Dinis-Ribeiro, M. -765G > C COX-2 polymorphism may be a susceptibility marker for gastric adenocarcinoma in patients with atrophy or intestinal metaplasia. World J. Gastroenterol. 2006, 12, 5473–5478. [Google Scholar] [CrossRef] [PubMed]
- Thompson, C.L.; Fink, S.P.; Lutterbaugh, J.D.; Elston, R.C.; Veigl, M.L.; Markowitz, S.D.; Li, L. Genetic variation in 15-hydroxyprostaglandin dehydrogenase and colon cancer susceptibility. PLoS ONE 2013, 8, e64122. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Chen, F.; Liu, X.; Yuan, D.; Zi, Y.; He, X.; He, R. Detection and clinical significance of COX-2 gene SNPs in gastric cancer. Cell Biochem. Biophys. 2015, 72, 657–660. [Google Scholar] [CrossRef] [PubMed]
- Hoeft, B.; Linseisen, J.; Beckmann, L.; Müller-Decker, K.; Canzian, F.; Hüsing, A.; Kaaks, R.; Vogel, U.; Jakobsen, M.U.; Overvad, K.; et al. Polymorphisms in fatty acid metabolism-related genes are associated with colorectal cancer risk. Carcinogenesis 2009, 31, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Sorby, L.A.A.; Andersen, S.N.; Bukholm, I.R.K.; Jacobsen, M.B. Evaluation of suitable reference genes for normalization of real-time reverse transcription PCR analysis in colon cancer. J. Exp. Clin. Cancer Res. 2010, 29. [Google Scholar] [CrossRef]
- Rho, H.W.; Lee, B.C.; Choi, E.S.; Choi, I.J.; Lee, Y.S.; Goh, S.H. Identification of valid reference genes for gene expression studies of human stomach cancer by reverse transcription-qPCR. BMC Cancer 2010, 10, 240. [Google Scholar] [CrossRef]
- Benjamini, Y.; Drai, D.; Elmer, G.; Kafkafi, N.; Golani, I. Controlling the false discovery rate in behavior genetics research. Behav. Brain Res. 2001, 125, 279–284. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Livak, K.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2 (-Delta Delta (T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]


{kind=link}
| Cases (n = 260) | Controls (n = 476) | p Value | ||
|---|---|---|---|---|
| Demographics | ||||
| Age (years) | ||||
| Mean ± SD | 69.87 ± 0.60 | 57.98 ± 0.23 | < 0.001 | |
| Median (min-max) | 70 (50–92) | 58 (50–69) | ||
| Sex, n (%) | ||||
| Male | 152 (58.5) | 312 (65.5) | 0.057 | |
| Female | 108 (41.5) | 164 (34.5) | ||
| Tumor characteristics | ||||
| Tumor location, n (%) | ||||
| Cardia and GEJ | 24 (9.4) | -- | ||
| Fundus and corpus | 41 (16.1) | -- | ||
| Antrum and corpus-antrum transition | 157 (61.6) | -- | ||
| Angularis incisura | 7 (2.7) | |||
| Others * | 26 (10.2) | |||
| Grade, n (%) | ||||
| Well-differentiated | 28 (10.8) | -- | ||
| Moderately differentiated | 157 (60.6) | -- | ||
| Poorly differentiated | 63 (24.3) | -- | ||
| Cannot be assessed | 11 (4.2) | -- | ||
| Stage, n (%) | ||||
| I-II | 145 (56.0) | -- | ||
| III-IV | 114 (44.0) | -- | ||
| Synchronous tumors, n (%) | ||||
| Yes | 6 (2.3) | -- | ||
| No | 254 (97.7) | -- | ||
| SNP | Model | Genotype Frequencies | Univariate Analysis | Multivariate Analysis | Age at Diagnosis | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cases, n (%) | Controls, n (%) | OR | 95% CI | p Value | aOR | 95% CI | p Value | Median (years) | 95% CI | p Value | ||
| PTGS2 | ||||||||||||
| rs689466 | Codominant | |||||||||||
| AA | 121 (61.1) | 322 (68.8) | 1.00 | - | 0.054 | 1.00 | - | 0.021 | 73.00 | 71.25–74.75 | - | |
| AG | 63 (31.8) | 130 (27.8) | 1.29 | 0.89–1.86 | 1.50 | 0.93–2.42 | 73.00 | 68.89–77.11 | 0.21 | |||
| GG | 14 (7.1) | 16 (3.4) | 2.33 | 1.10–4.92 | 3.40 | 1.29–8.97 | 70.00 | 62.49–77.52 | 0.008 | |||
| Dominant | ||||||||||||
| AA | 121 (61.1) | 322 (68.8) | 1.00 | - | 0.056 | 1.00 | - | 0.022 | 73.00 | 71.25–74.75 | - | |
| AG-GG | 77 (38.9) | 146 (31.2) | 1.40 | 0.99–1.98 | 1.69 | 1.08–2.65 | 73.00 | 69.94–76.06 | 0.058 | |||
| Recessive | ||||||||||||
| AA-AG | 184 (92.9) | 452 (96.6) | 1.00 | - | 0.046 | 1.00 | - | 0.027 | 73.00 | 71.48–74.52 | - | |
| GG | 14 (7.1) | 16 (3.4) | 2.15 | 1.03–4.49 | 2.98 | 1.14–7.74 | 70.00 | 62.49–77.52 | 0.011 | |||
| Overdominant | ||||||||||||
| AA-GG | 135 (68.2) | 338 (72.2) | 1.00 | - | 0.30 | 1.00 | - | 0.19 | 73.00 | 71.46–74.54 | - | |
| AG | 63 (31.8) | 130 (27.8) | 1.21 | 0.85–1.74 | 1.37 | 0.86–2.19 | 73.00 | 68.89–77.11 | 0.38 | |||
| Log-additive | - | 1.40 | 1.06–1.86 | 0.021 | 1.66 | 1.15–2.40 | 0.007 | - | ||||
| ABCC4 | ||||||||||||
| rs1678374 | Codominant | |||||||||||
| TT | 90 (40.5) | 161 (33.9) | 1.00 | - | 0.076 | 1.00 | - | 0.063 | 72.00 | 69.48–74.52 | - | |
| TC | 107 (48.2) | 234 (49.3) | 0.82 | 0.58–1.15 | 1.04 | 0.67–1.63 | 72.00 | 69.74–74.26 | 0.47 | |||
| CC | 25 (11.3) | 80 (16.8) | 0.56 | 0.33–0.94 | 0.50 | 0.26–0.97 | 73.00 | 71.00–75.00 | 0.57 | |||
| Dominant | ||||||||||||
| TT | 90 (40.5) | 161 (33.9) | 1.00 | - | 0.09 | 1.00 | - | 0.55 | 72.00 | 69.48–74.52 | - | |
| TC-CC | 132 (59.5) | 314 (66.1) | 0.75 | 0.54–1.04 | 0.88 | 0.58–1.34 | 72.00 | 70.77–73.23 | 0.66 | |||
| rs1678374 | Recessive | |||||||||||
| TT-TC | 197 (88.7) | 395 (83.2) | 1.00 | - | 0.05 | 1.00 | - | 0.019 | 72.00 | 70.38–73.63 | - | |
| CC | 25 (11.3) | 80 (16.8) | 0.63 | 0.39–1.01 | 0.49 | 0.26–0.91 | 73.00 | 71.00–75.00 | 0.39 | |||
| Overdominant | ||||||||||||
| TT-CC | 115 (51.8) | 241 (50.7) | 1.00 | - | 0.79 | 1.00 | - | 0.28 | 72.00 | 70.46–73.54 | - | |
| TC | 107 (48.2) | 234 (49.3) | 0.96 | 0.70–1.32 | 1.25 | 0.83–1.89 | 72.00 | 69.74–74.26 | 0.31 | |||
| Log-additive | - | 0.77 | 0.60–0.97 | 0.027 | 0.78 | 0.58–1.06 | 0.11 | - | ||||
| rs1678405 | Codominant | |||||||||||
| TT | 108 (50.2) | 196 (41.2) | 1.00 | - | 0.052 | 1.00 | - | 0.09 | 72.00 | 70.40–73.60 | - | |
| TC | 91 (42.3) | 226 (47.5) | 0.73 | 0.52–1.02 | 0.81 | 0.52–1.25 | 72.00 | 69.60–74.40 | 0.58 | |||
| CC | 16 (7.4) | 54 (11.3) | 0.54 | 0.29–0.99 | 0.44 | 0.20–0.95 | 74.00 | 67.60–80.40 | 0.68 | |||
| Dominant | ||||||||||||
| TT | 108 (50.2) | 196 (41.2) | 1.00 | - | 0.027 | 1.00 | - | 0.13 | 72.00 | 70.40–73.60 | - | |
| TC-CC | 107 (49.8) | 280 (58.8) | 0.69 | 0.50–0.96 | 0.73 | 0.48–1.10 | 72.00 | 70.18–73.82 | 0.70 | |||
| Recessive | ||||||||||||
| TT-TC | 199 (92.6) | 422 (88.7) | 1.00 | - | 0.11 | 1.00 | - | 0.049 | 72.00 | 70.58–73.42 | - | |
| CC | 16 (7.4) | 54 (11.3) | 0.63 | 0.35–1.13 | 0.49 | 0.23–1.03 | 74.00 | 67.60–80.40 | 0.58 | |||
| Overdominant | ||||||||||||
| TT-CC | 124 (57.7) | 250 (52.5) | 1.00 | - | 0.21 | 1.00 | - | 0.76 | 72.00 | 70.46–73.54 | - | |
| TC | 91 (42.3) | 226 (47.5) | 0.81 | 0.59–1.12 | 0.94 | 0.62–1.42 | 72.00 | 69.60–74.40 | 0.48 | |||
| Log-additive | - | 0.73 | 0.57–0.94 | 0.015 | 0.72 | 0.52–0.99 | 0.041 | - | ||||
| rs1751031 | Codominant | |||||||||||
| AA | 154 (69.4) | 296 (62.3) | 1.00 | - | 0.10 | 1.00 | - | 0.073 | 72.00 | 70.25–73.75 | - | |
| AG | 59 (26.6) | 164 (34.5) | 0.69 | 0.48 | 0.61 | 0.39–0.95 | 72.00 | 69.72–74.28 | 0.66 | |||
| GG | 9 (4.0) | 15 (302) | 1.15 | 0.49–2.70 | 0.57 | 0.17–1.92 | 78.00 | 69.81–86.19 | 0.29 | |||
| rs1751031 | Dominant | |||||||||||
| AA | 154 (69.4) | 296 (62.3) | 1.00 | - | 0.068 | 1.00 | - | 0.022 | 72.00 | 70.25–73.75 | - | |
| AG-GG | 68 (30.6) | 179 (37.7) | 0.73 | 0.52–1.03 | 0.60 | 0.39–0.94 | 72.00 | 70.08–73.93 | 0.46 | |||
| Recessive | ||||||||||||
| AA-AG | 213 (96.0) | 460 (96.8) | 1.00 | - | 0.55 | 1.00 | - | 0.52 | 72.00 | 70.61–73.39 | - | |
| GG | 9 (4.0) | 15 (3.2) | 1.30 | 0.56–3.01 | 0.68 | 0.21–2.24 | 78.00 | 69.81–86.19 | 0.31 | |||
| Overdominant | ||||||||||||
| AA-GG | 163 (73.4) | 311 (65.5) | 1.00 | - | 0.034 | 1.00 | - | 0.036 | 72.00 | 70.28-73.72 | - | |
| AG | 59 (26.6) | 164 (34.5) | 0.69 | 0.48-0.98 | 0.62 | 0.40-0.98 | 72.00 | 69.72-74.28 | 0.79 | |||
| Log-additive | - | 0.81 | 0.61-1.09 | 0.17 | 0.65 | 0.44-0.96 | 0.028 | - | ||||
| HPGD | ||||||||||||
| rs2303520 | Codominant | |||||||||||
| GG | 143 (64.4) | 339 (71.4) | 1.00 | - | 0.037 | 1.00 | - | 0.065 | 72.00 | 70.89–73.11 | - | |
| GA | 76 (34.2) | 122 (25.7) | 1.48 | 1.04–2.09 | 1.61 | 1.02–2.54 | 72.00 | 69.12–74.88 | 0.83 | |||
| AA | 3 (1.4) | 14 (3.0) | 0.51 | 0.14–1.79 | 0.51 | 0.11–2.34 | 69.00 | 64.20–73.80 | 0.61 | |||
| Dominant | ||||||||||||
| GG | 143 (64.4) | 339 (71.4) | 1.00 | - | 0.066 | 1.00 | - | 0.086 | 72.00 | 70.89–73.11 | - | |
| GA-AA | 79 (35.6) | 136 (28.6) | 1.38 | 0.98–1.93 | 1.47 | 0.95–2.29 | 72.00 | 69.05–74.96 | 0.92 | |||
| Recessive | ||||||||||||
| GG-GA | 219 (98.7) | 461 (97.0) | 1.00 | - | 0.18 | 1.00 | - | 0.26 | 72.00 | 70.63–73.37 | - | |
| AA | 3 (1.4) | 14 (3.0) | 0.45 | 0.13–1.59 | 0.45 | 0.10–2.04 | 69.00 | 64.20–73.80 | 0.61 | |||
| Overdominant | ||||||||||||
| GG-AA | 146 (65.8) | 353 (74.3) | 1.00 | - | 0.021 | 1.00 | - | 0.031 | 72.00 | 70.87–73.13 | - | |
| GA | 76 (34.2) | 122 (25.7) | 1.51 | 1.07–2.13 | 1.65 | 1.05–2.59 | 72.00 | 69.12–74.88 | 0.81 | |||
| Log-additive | - | 1.21 | 0.90–1.64 | 0.21 | 1.26 | 0.86–1.84 | 0.24 | - | ||||
| SLCO2A1 | ||||||||||||
| rs10935090 | Codominant | |||||||||||
| CC | 162 (73.0) | 378 (79.6) | 1.00 | - | 0.13 | 1.00 | - | 0.026 | 73.00 | 71.81–74.19 | - | |
| CT | 54 (24.3) | 90 (18.9) | 1.40 | 0.95–2.06 | 0.13 | 1.46 | 0.90–2.39 | 0.026 | 70.00 | 67.56–72.44 | 0.034 | |
| TT | 6 (2.7) | 7 (1.5) | 2.00 | 0.66–6.04 | 4.68 | 1.32–16.6 | 62.00 | 59.61–64.39 | <0.001 | |||
| Dominant | ||||||||||||
| CC | 162 (73.0) | 378 (79.6) | 1.00 | - | 0.054 | 1.00 | - | 0.038 | 73.00 | 71.81–74.19 | - | |
| CT-TT | 60 (27.0) | 97 (20.4) | 1.44 | 1.00–2.09 | 1.65 | 1.03–2.63 | 70.00 | 67.93–72.07 | 0.007 | |||
| Recessive | ||||||||||||
| CC-CT | 216 (97.3) | 468 (98.5) | 1.00 | - | 0.28 | 1.00 | - | 0.026 | 72.00 | 70.87–73.14 | - | |
| TT | 6 (2.7) | 7 (1.5) | 1.86 | 0.62–5.59 | 4.30 | 1.22–15.2 | 62.00 | 59.61–64.39 | <0.001 | |||
| Overdominant | ||||||||||||
| CC-TT | 168 (75.7) | 385 (81.0) | 1.00 | - | 0.11 | 1.00 | - | 0.19 | 73.00 | 71.81–74.19 | - | |
| CT | 54 (24.3) | 90 (18.9) | 1.37 | 0.94–2.02 | 1.39 | 0.86–2.27 | 70.00 | 67.56–72.44 | 0.057 | |||
| Log-additive | - | 1.40 | 1.01–1.95 | 0.044 | 1.69 | 1.12–2.53 | 0.012 | - | ||||
| rs11915399 | Codominant | |||||||||||
| CC | 159 (71.6) | 326 (68.6) | 1.00 | - | 0.72 | 1.00 | - | 0.12 | 72.00 | 70.27–73.73 | - | |
| CT | 57 (25.7) | 135 (28.4) | 0.87 | 0.60–1.24 | 0.61 | 0.38–0.99 | 73.00 | 71.47–74.53 | 0.13 | |||
| TT | 6 (2.7) | 14 (3.0) | 0.88 | 0.33–2.33 | 0.75 | 0.22–2.63 | 74.00 | 59.93–88.07 | 0.52 | |||
| Dominant | ||||||||||||
| CC | 159 (71.6) | 326 (68.6) | 1.00 | - | 0.42 | 1.00 | - | 0.043 | 72.00 | 70.27–73.73 | - | |
| CT-TT | 63 (28.4) | 149 (31.4) | 0.87 | 0.61–1.23 | 0.62 | 0.39–0.99 | 73.00 | 71.53–74.47 | 0.11 | |||
| rs11915399 | Recessive | |||||||||||
| CC-CT | 216 (97.3) | 461 (97.0) | 1.00 | - | 0.86 | 1.00 | - | 0.81 | 72.00 | 70.64–73.37 | - | |
| TT | 6 (2.7) | 14 (3.0) | 0.91 | 0.35–2.41 | 0.86 | 0.25–2.96 | 74.00 | 59.93–88.07 | 0.59 | |||
| Overdominant | ||||||||||||
| CC-TT | 165 (74.3) | 340 (71.6) | 1.00 | - | 0.45 | 1.00 | - | 0.045 | 72.00 | 70.19–73.81 | - | |
| CT | 57 (25.7) | 135 (28.4) | 0.87 | 0.61–1.25 | 0.62 | 0.38–1.00 | 73.00 | 71.47–74.53 | 0.16 | |||
| Log-additive | - | 0.89 | 0.65–1.21 | 0.45 | 0.69 | 0.46–1.03 | 0.065 | - | ||||
| rs9821091 | Codominant | |||||||||||
| GG | 87 (39.2) | 179 (37.7) | 1.00 | - | 0.32 | 1.00 | - | 0.045 | 72.00 | 69.77–74.23 | - | |
| GA | 97 (43.7) | 232 (48.8) | 0.86 | 0.61–1.22 | 0.81 | 0.52–1.28 | 73.00 | 71.43–74.57 | 0.11 | |||
| AA | 38 (17.1) | 64 (13.5) | 1.22 | 0.76–1.97 | 1.75 | 0.95–3.20 | 71.00 | 68.17–73.83 | 0.16 | |||
| Dominant | ||||||||||||
| GG | 87 (39.2) | 179 (37.7) | 1.00 | - | 0.70 | 1.00 | - | 0.96 | 72.00 | 69.77–74.23 | - | |
| GA-AA | 135 (60.8) | 296 (62.3) | 0.94 | 0.68–1.30 | 0.99 | 0.65–1.50 | 72.00 | 70.56–73.44 | 0.42 | |||
| Recessive | ||||||||||||
| GG-GA | 184 (82.9) | 411 (86.5) | 1.00 | - | 0.21 | 1.00 | - | 0.02 | 72.00 | 70.76–73.24 | - | |
| AA | 38 (17.1) | 64 (16.5) | 1.33 | 0.86–1.12 | 1.95 | 1.12–3.40 | 71.00 | 68.17–73.83 | 0.017 | |||
| Overdominant | ||||||||||||
| GG-AA | 125 (56.3) | 243 (51.2) | 1.00 | - | 0.20 | 1.00 | - | 0.085 | 71.00 | 69.06–72.94 | - | |
| GA | 97 (43.7) | 232 (48.8) | 0.81 | 0.59–1.12 | 0.70 | 0.46–1.05 | 73.00 | 71.43–74.57 | 0.018 | |||
| Log-additive | - | 1.05 | 0.83–1.32 | 0.70 | 1.19 | 0.89–1.61 | 0.24 | - | ||||
| Gene/Haplotype | % Cases | % Controls | aOR | 95% CI | p Value |
|---|---|---|---|---|---|
| ABCC4£ | |||||
| T-T-A | 49.09 | 42.16 | 1.00 | Reference | - |
| C-C-A | 13.84 | 16.87 | 0.67 | 0.42–1.09 | 0.11 |
| C-T-A | 13.12 | 12.46 | 0.94 | 0.52–1.69 | 0.83 |
| C-C-G | 7.07 | 9.68 | 0.47 | 0.23–0.93 | 0.032 |
| T-T-G | 7.42 | 7.81 | 0.52 | 0.26–1.06 | 0.074 |
| T-C-A | 6.61 | 8.09 | 0.70 | 0.35–1.43 | 0.33 |
| C-T-G | 1.33 | 2.49 | 0.56 | 0.11–2.74 | 0.47 |
| SLCO2A1¥ | |||||
| C-C-G | 45.37 | 45.31 | 1.00 | Reference | - |
| C-C-A | 26.04 | 27.74 | 0.89 | 0.60–1.32 | 0.57 |
| C-T-G | 9.23 | 11.12 | 0.58 | 0.32–1.07 | 0.084 |
| T-C-A | 6.61 | 4.98 | 2.78 | 1.41–5.48 | 0.0034 |
| T-C-G | 6.44 | 0.48 | 0.91 | 0.44–1.87 | 0.80 |
| CV Accuracy | CV Consistency | aOR | 95% CI | p Value | |
|---|---|---|---|---|---|
| Best model | |||||
| rs689466 | 0.621 | 10/10 | 2.743 | 1.967–3.826 | <0.0001 |
| age, rs1678374 | 0.687 | 5/10 | 4.953 | 3.434–7.143 | <0.0001 |
| age, rs689466, rs1678374 | 0.807 | 8/10 | 17.581 | 11.672–26.482 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopes, C.; Pereira, C.; Farinha, M.; Medeiros, R.; Dinis-Ribeiro, M. Genetic Variations in Prostaglandin E2 Pathway Identified as Susceptibility Biomarkers for Gastric Cancer in an Intermediate Risk European Country. Int. J. Mol. Sci. 2021, 22, 648. https://doi.org/10.3390/ijms22020648
Lopes C, Pereira C, Farinha M, Medeiros R, Dinis-Ribeiro M. Genetic Variations in Prostaglandin E2 Pathway Identified as Susceptibility Biomarkers for Gastric Cancer in an Intermediate Risk European Country. International Journal of Molecular Sciences. 2021; 22(2):648. https://doi.org/10.3390/ijms22020648
Chicago/Turabian StyleLopes, Catarina, Carina Pereira, Mónica Farinha, Rui Medeiros, and Mário Dinis-Ribeiro. 2021. "Genetic Variations in Prostaglandin E2 Pathway Identified as Susceptibility Biomarkers for Gastric Cancer in an Intermediate Risk European Country" International Journal of Molecular Sciences 22, no. 2: 648. https://doi.org/10.3390/ijms22020648
APA StyleLopes, C., Pereira, C., Farinha, M., Medeiros, R., & Dinis-Ribeiro, M. (2021). Genetic Variations in Prostaglandin E2 Pathway Identified as Susceptibility Biomarkers for Gastric Cancer in an Intermediate Risk European Country. International Journal of Molecular Sciences, 22(2), 648. https://doi.org/10.3390/ijms22020648