
Roles of Non-Canonical Wnt Signalling Pathways in Bone Biology

Abstract
:1. Introduction: The Role of Wnt Signalling Pathways in Bone Biology
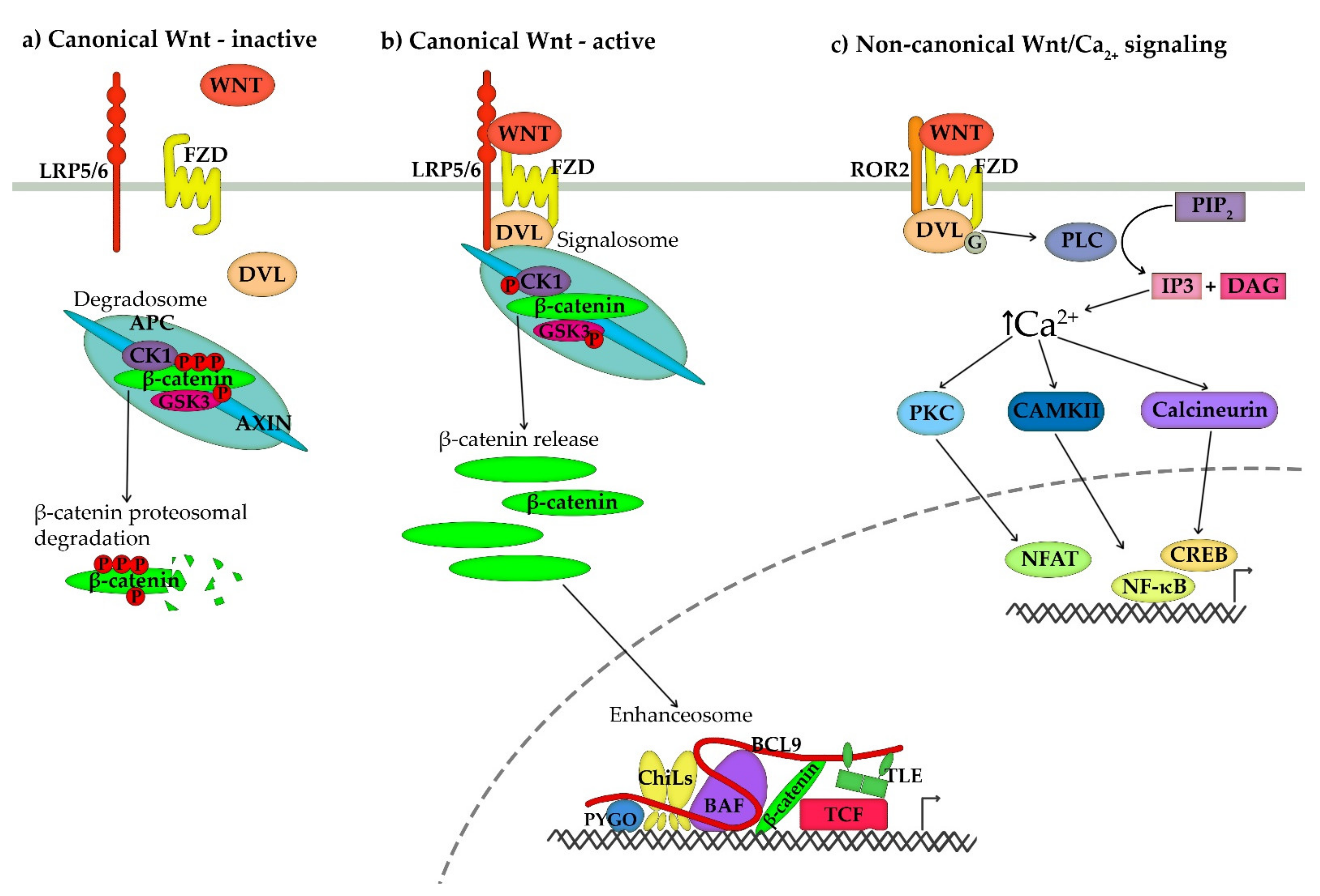
2. Overview of the Canonical and Non-Canonical Wnt Signalling Pathways

{kind=link}
{kind=link}
{kind=link}
| Ligand | Effects of Activation of Non-Canonical | Non-Canonical Signal Transduction | Effects of Activation of Canonical |
|---|---|---|---|
| Wnt1 | N/A | Stem cell maintenance [56] | |
| Promotion of Osteoblastogenesis [56,57] | |||
| Wnt2b | N/A | Osteogenic differentiation [58,59] | |
| Wnt3a | Suppression of osteoclast differentiation | cAMP/PKA pathway [60] | Stem cell maintenance [36,61,62,63] |
| Suppression of chondrocyte differentiation | c-Jun/AP-1 [60] | Suppression of osteoblastogenesis [64,65] | |
| Induction of osteoblastogenesis | PKCδ [66] | Suppression of osteoclastogenesis [60,67] | |
| mTORC1, mTORC2 [68,69] | Suppression of chondrogenesis [70] | ||
| JNK pathway [31] | |||
| Promotion of osteoblastogenesis | RhoA [71] | ||
| Prevention of starvation induced apoptosis | c-Src/ERK1/2 and PI3K/Akt [32] | ||
| Wnt4 | Osteogenic differentiation, bone formation | p38 MAPK [72] | N/A |
| Inhibition of osteoclast differentiation | TAK1-TAB2-NLK [73] | ||
| Wnt5a | Promotion of osteoclastogenesis | DAAM2/Rho/PKN3/c-Src [74] | N/A |
| JNK/c-Jun/Sp1 [19] | |||
| PKC/CAMKIIα/JNK [64] | |||
| Suppression of canonical Wnt signalling | PKC/CAMKIIα/JNK [64] | ||
| TAK1/NLK [75] | |||
| Prevention of starvation-induced apoptosis | c-Src/ERK1/2 and PI3K/Akt [32] | ||
| Promotion of chondrogenesis | CaMK/NFAT [76] | ||
| Inhibition of chondrogenesis | PI3K/Akt/IKK/NF-κB [76] | ||
| Promotion of osteoblastogenesis | RhoA [77] | ||
| JNK/c-Jun [78] | |||
| Wnt5b | Suppression of chondrogenesis | JNK pathway [79] | N/A |
| Wnt7b | Promotion of osteoblastogenesis | mTORC1, mTORC2 [69,80] | N/A |
| Gαq/11/PLCβ/PKCδ [66] | |||
| Wnt10b | Promotion of osteoblastogenesis | mTORC2 [69] | Promotion of osteoblastogenesis [81,82,83,84,85,86] |
| Wnt16 | Suppression of osteoclastogenesis | JNK/c-Jun [86] | Suppression of osteoclastogenesis [87] |
| Promotion of osteogenesis | JNK pathway [65] |
3. Wnt/Ca2+ Non-Canonical Signalling Pathway
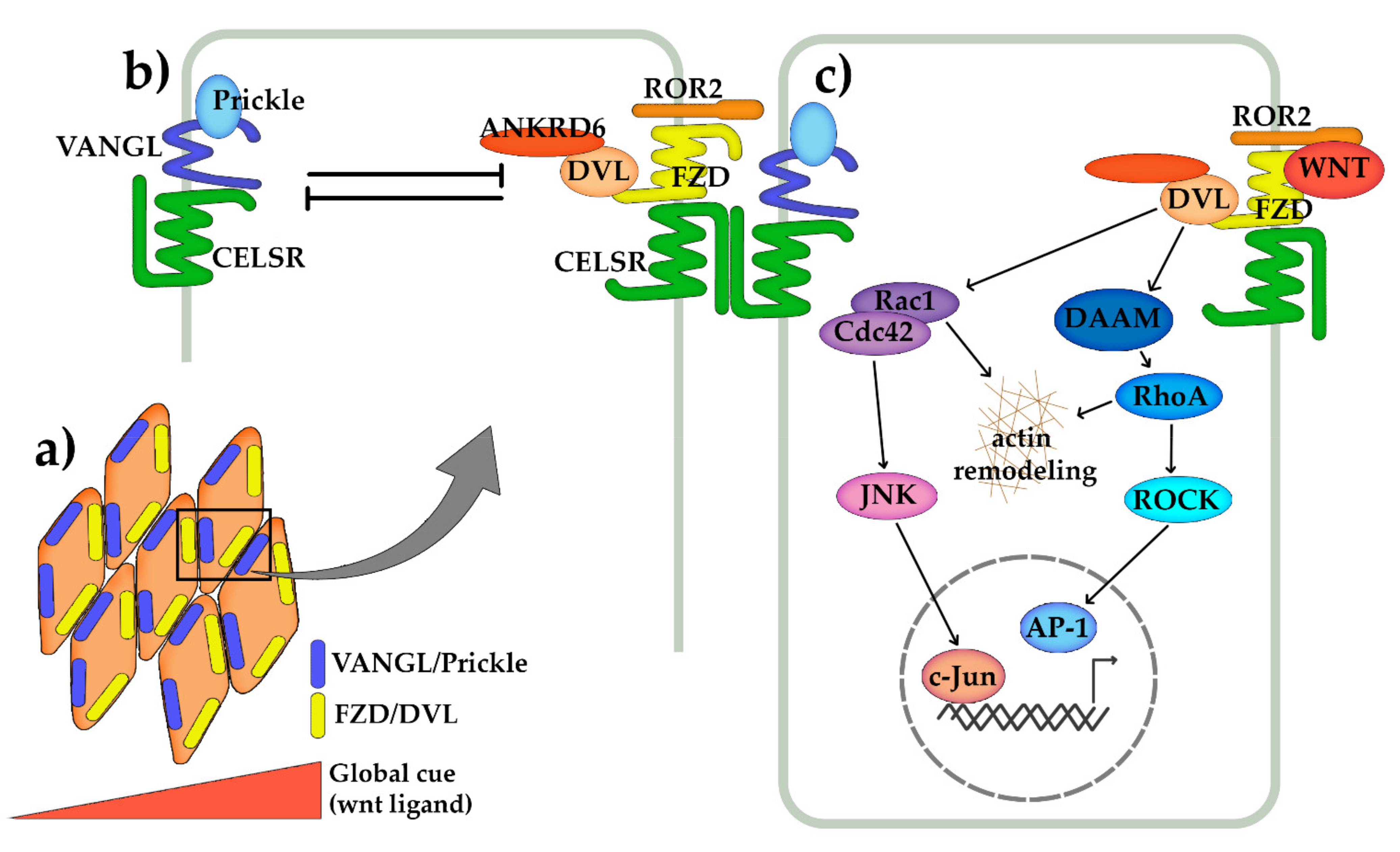
4. Wnt/Planar Cell Polarity (PCP) Pathway
5. Non-Canonical Wnt Crosstalk with Other Signalling Pathways
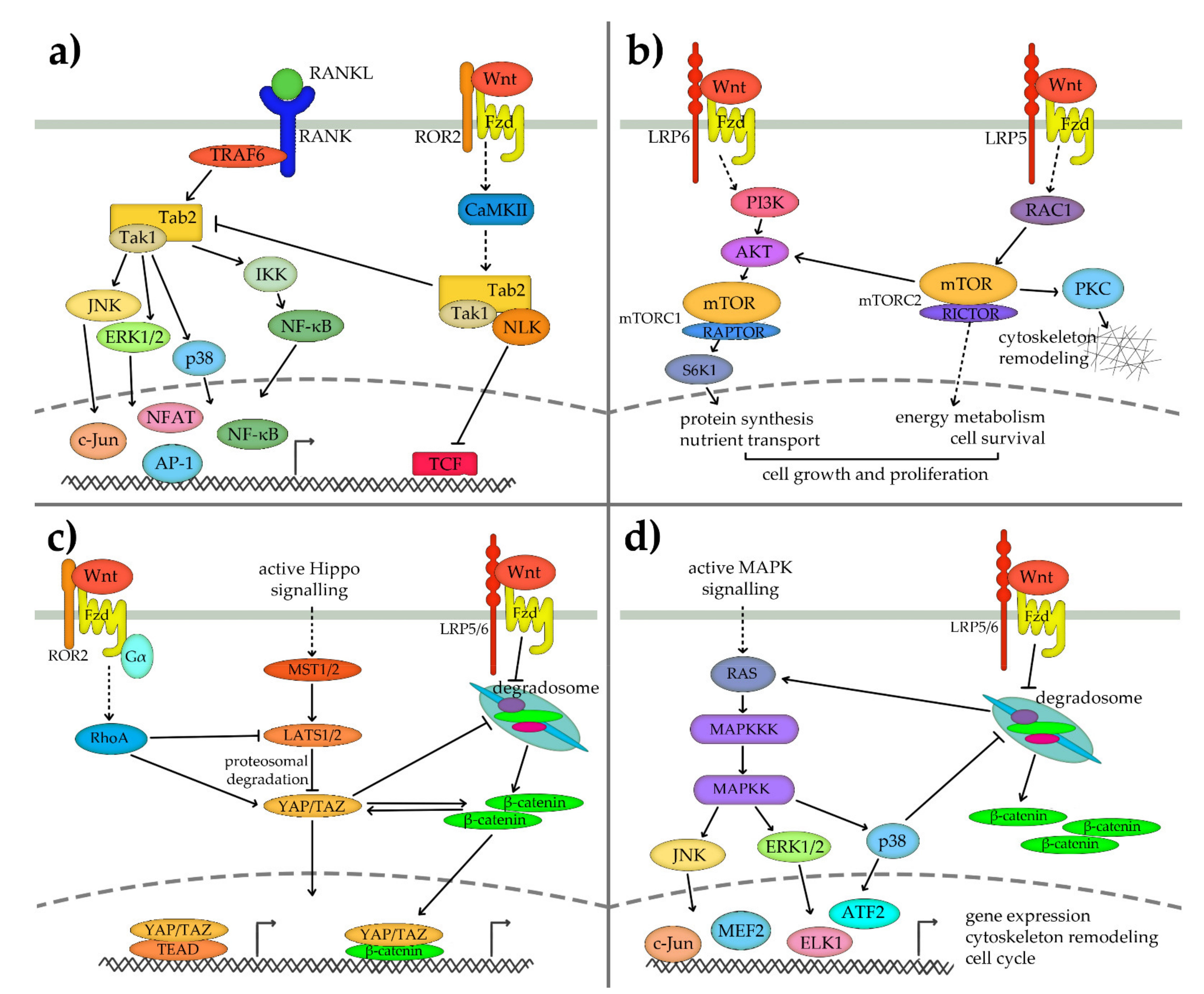
5.1. Non-Canonical Wnt Crosstalk with RANKL/NF-κB Signalling Pathway
5.2. Non-Canonical Wnt Crosstalk with mTOR Pathway
5.3. Non-Canonical Wnt Crosstalk with Hippo Signalling Pathway
5.4. Non-Canonical Wnt Crosstalk with MAPK Pathway
6. Syndromes and Osteoporosis Treatments Related to Non-Canonical Wnt Signalling Pathways
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Logan, C.Y.; Nusse, R. The Wnt Signaling Pathway in Development and Disease. Annu. Rev. Cell Dev. Biol. 2004, 20, 781–810. [Google Scholar] [CrossRef] [Green Version]
- Houschyar, K.S.; Tapking, C.; Borrelli, M.R.; Popp, D.; Duscher, D.; Maan, Z.N.; Chelliah, M.P.; Li, J.; Harati, K.; Wallner, C.; et al. Wnt Pathway in Bone Repair and Regeneration—What Do We Know So Far. Front. Cell Dev. Biol. 2019, 6, 170. [Google Scholar] [CrossRef]
- Zhan, T.; Rindtorff, N.; Boutros, M. Wnt Signaling in Cancer. Oncogene 2017, 36, 1461–1473. [Google Scholar] [CrossRef] [PubMed]
- Aghaizu, N.D.; Jin, H.; Whiting, P.J. Dysregulated Wnt Signalling in the Alzheimer’s Brain. Brain Sci. 2020, 10, 902. [Google Scholar] [CrossRef]
- Abou Ziki, M.D.; Mani, A. The Interplay of Canonical and Noncanonical Wnt Signaling in Metabolic Syndrome. Nutr. Res. 2019, 70, 18–25. [Google Scholar] [CrossRef]
- Chen, J.; Ning, C.; Mu, J.; Li, D.; Ma, Y.; Meng, X. Role of Wnt Signaling Pathways in Type 2 Diabetes Mellitus. Mol. Cell. Biochem. 2021, 476, 2219–2232. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Uehara, S.; Udagawa, N.; Takahashi, N. Regulation of Bone Metabolism by Wnt Signals. J. Biochem. 2016, 159, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, V.; Bryant, H.U.; Macdougald, O.A. Regulation of Bone Mass by Wnt Signaling. J. Clin. Investig. 2006, 116, 1202–1209. [Google Scholar] [CrossRef]
- Gong, Y.; Slee, R.B.; Fukai, N.; Rawadi, G.; Roman-Roman, S.; Reginato, A.M.; Wang, H.; Cundy, T.; Glorieux, F.H.; Lev, D.; et al. LDL Receptor-Related Protein 5 (LRP5) Affects Bone Accrual and Eye Development. Cell 2001, 107, 513–523. [Google Scholar] [CrossRef] [Green Version]
- Huybrechts, Y.; Mortier, G.; Boudin, E.; Van Hul, W. WNT Signaling and Bone: Lessons From Skeletal Dysplasias and Disorders. Front. Endocrinol. 2020, 11, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trajanoska, K.; Rivadeneira, F. The Genetic Architecture of Osteoporosis and Fracture Risk. Bone 2019, 126, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, M.D.; Chen-Plotkin, A.S. The Post-GWAS Era: From Association to Function. Am. J. Hum. Genet. 2018, 102, 717–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, M.; Patel, M.S.; Levasseur, R.; Lobov, I.; Chang, B.H.-J.; Glass, D.A.; Hartmann, C.; Li, L.; Hwang, T.-H.; Brayton, C.F.; et al. Cbfa1-Independent Decrease in Osteoblast Proliferation, Osteopenia, and Persistent Embryonic Eye Vascularization in Mice Deficient in Lrp5, a Wnt Coreceptor. J. Cell Biol. 2002, 157, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Holmen, S.L.; Zylstra, C.R.; Mukherjee, A.; Sigler, R.E.; Faugere, M.-C.; Bouxsein, M.L.; Deng, L.; Clemens, T.L.; Williams, B.O. Essential Role of Beta-Catenin in Postnatal Bone Acquisition. J. Biol. Chem. 2005, 280, 21162–21168. [Google Scholar] [CrossRef] [Green Version]
- Hill, T.P.; Später, D.; Taketo, M.M.; Birchmeier, W.; Hartmann, C. Canonical Wnt/Beta-Catenin Signaling Prevents Osteoblasts from Differentiating into Chondrocytes. Dev. Cell 2005, 8, 727–738. [Google Scholar] [CrossRef] [Green Version]
- Day, T.F.; Guo, X.; Garrett-Beal, L.; Yang, Y. Wnt/Beta-Catenin Signaling in Mesenchymal Progenitors Controls Osteoblast and Chondrocyte Differentiation during Vertebrate Skeletogenesis. Dev. Cell 2005, 8, 739–750. [Google Scholar] [CrossRef] [Green Version]
- Glass, D.A.; Bialek, P.; Ahn, J.D.; Starbuck, M.; Patel, M.S.; Clevers, H.; Taketo, M.M.; Long, F.; McMahon, A.P.; Lang, R.A.; et al. Canonical Wnt Signaling in Differentiated Osteoblasts Controls Osteoclast Differentiation. Dev. Cell 2005, 8, 751–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, I.; Halleux, C.; Keller, H.; Pegurri, M.; Gooi, J.H.; Weber, P.B.; Feng, J.Q.; Bonewald, L.F.; Kneissel, M. Osteocyte Wnt/Beta-Catenin Signaling Is Required for Normal Bone Homeostasis. Mol. Cell. Biol. 2010, 30, 3071–3085. [Google Scholar] [CrossRef] [Green Version]
- Maeda, K.; Kobayashi, Y.; Udagawa, N.; Uehara, S.; Ishihara, A.; Mizoguchi, T.; Kikuchi, Y.; Takada, I.; Kato, S.; Kani, S.; et al. Wnt5a-Ror2 Signaling between Osteoblast-Lineage Cells and Osteoclast Precursors Enhances Osteoclastogenesis. Nat. Med. 2012, 18, 405–412. [Google Scholar] [CrossRef]
- Zeng, X.; Huang, H.; Tamai, K.; Zhang, X.; Harada, Y.; Yokota, C.; Almeida, K.; Wang, J.; Doble, B.; Woodgett, J.; et al. Initiation of Wnt Signaling: Control of Wnt Coreceptor Lrp6 Phosphorylation/Activation via Frizzled, Dishevelled and Axin Functions. Development 2008, 135, 367–375. [Google Scholar] [CrossRef] [Green Version]
- Bienz, M. Signalosome Assembly by Domains Undergoing Dynamic Head-to-Tail Polymerization. Trends Biochem. Sci. 2014, 39, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, M.; Graeb, M.; Mieszczanek, J.; Rutherford, T.J.; Johnson, C.M.; Bienz, M. An Ancient Pygo-Dependent Wnt Enhanceosome Integrated by Chip/LDB-SSDP. eLife 2015, 4, e09073. [Google Scholar] [CrossRef] [PubMed]
- Sekiya, T.; Zaret, K.S. Repression by Groucho/TLE/Grg Proteins: Genomic Site Recruitment Generates Compacted Chromatin in Vitro and Impairs Activator Binding in Vivo. Mol. Cell 2007, 28, 291–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arce, L.; Yokoyama, N.N.; Waterman, M.L. Diversity of LEF/TCF Action in Development and Disease. Oncogene 2006, 25, 7492–7504. [Google Scholar] [CrossRef] [Green Version]
- Turki-Judeh, W.; Courey, A.J. Groucho: A Corepressor with Instructive Roles in Development. Curr. Top. Dev. Biol. 2012, 98, 65–96. [Google Scholar] [CrossRef] [PubMed]
- Veeman, M.T.; Axelrod, J.D.; Moon, R.T. A Second Canon. Functions and Mechanisms of Beta-Catenin-Independent Wnt Signaling. Dev. Cell 2003, 5, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Mikels, A.J.; Nusse, R. Purified Wnt5a Protein Activates or Inhibits Beta-Catenin-TCF Signaling Depending on Receptor Context. PLoS Biol. 2006, 4, e115. [Google Scholar] [CrossRef]
- Van Amerongen, R.; Fuerer, C.; Mizutani, M.; Nusse, R. Wnt5a Can Both Activate and Repress Wnt/β-Catenin Signaling during Mouse Embryonic Development. Dev. Biol. 2012, 369, 101–114. [Google Scholar] [CrossRef] [Green Version]
- Nalesso, G.; Sherwood, J.; Bertrand, J.; Pap, T.; Ramachandran, M.; De Bari, C.; Pitzalis, C.; Dell’Accio, F. WNT-3A Modulates Articular Chondrocyte Phenotype by Activating Both Canonical and Noncanonical Pathways. J Cell Biol. 2011, 193, 551–564. [Google Scholar] [CrossRef] [Green Version]
- Grumolato, L.; Liu, G.; Mong, P.; Mudbhary, R.; Biswas, R.; Arroyave, R.; Vijayakumar, S.; Economides, A.N.; Aaronson, S.A. Canonical and Noncanonical Wnts Use a Common Mechanism to Activate Completely Unrelated Coreceptors. Genes Dev. 2010, 24, 2517–2530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, W.; Chen, L.; Kassem, M. Activation of Non-Canonical Wnt/JNK Pathway by Wnt3a Is Associated with Differentiation Fate Determination of Human Bone Marrow Stromal (Mesenchymal) Stem Cells. Biochem. Biophys. Res. Commun. 2011, 413, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.; Han, L.; Bellido, T.; Manolagas, S.C.; Kousteni, S. Wnt Proteins Prevent Apoptosis of Both Uncommitted Osteoblast Progenitors and Differentiated Osteoblasts by β-Catenin-Dependent and -Independent Signaling Cascades Involving Src/ERK and Phosphatidylinositol 3-Kinase/AKT. J. Biol. Chem. 2005, 280, 41342–41351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weidinger, G.; Moon, R.T. When Wnts Antagonize Wnts. J. Cell Biol. 2003, 162, 753–756. [Google Scholar] [CrossRef] [PubMed]
- Baksh, D.; Boland, G.M.; Tuan, R.S. Cross-Talk between Wnt Signaling Pathways in Human Mesenchymal Stem Cells Leads to Functional Antagonism during Osteogenic Differentiation. J. Cell. Biochem. 2007, 101, 1109–1124. [Google Scholar] [CrossRef]
- Van Amerongen, R.; Mikels, A.; Nusse, R. Alternative Wnt Signaling Is Initiated by Distinct Receptors. Sci. Signal. 2008, 1, re9. [Google Scholar] [CrossRef] [Green Version]
- Quarto, N.; Behr, B.; Longaker, M.T. Opposite Spectrum of Activity of Canonical Wnt Signaling in the Osteogenic Context of Undifferentiated and Differentiated Mesenchymal Cells: Implications for Tissue Engineering. Tissue Eng. Part A 2010, 16, 3185–3197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, J.-C.; Rattner, A.; Smallwood, P.M.; Nathans, J. Biochemical Characterization of Wnt-Frizzled Interactions Using a Soluble, Biologically Active Vertebrate Wnt Protein. Proc. Natl. Acad. Sci. USA 1999, 96, 3546–3551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Nusse, R. Ligand Receptor Interactions in the Wnt Signaling Pathway InDrosophila. J. Biol. Chem. 2002, 277, 41762–41769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rulifson, E.J.; Wu, C.H.; Nusse, R. Pathway Specificity by the Bifunctional Receptor Frizzled Is Determined by Affinity for Wingless. Mol. Cell 2000, 6, 117–126. [Google Scholar] [CrossRef]
- Wang, Y.; Chang, H.; Rattner, A.; Nathans, J. Frizzled Receptors in Development and Disease. Curr. Top. Dev. Biol. 2016, 117, 113–139. [Google Scholar] [CrossRef] [Green Version]
- Kushwaha, P.; Kim, S.; Foxa, G.E.; Michalski, M.N.; Williams, B.O.; Tomlinson, R.E.; Riddle, R.C. Frizzled-4 Is Required for Normal Bone Acquisition despite Compensation by Frizzled-8. J. Cell. Physiol. 2020, 235, 6673–6683. [Google Scholar] [CrossRef]
- Liu, Y.; Bhat, R.A.; Seestaller-Wehr, L.M.; Fukayama, S.; Mangine, A.; Moran, R.A.; Komm, B.S.; Bodine, P.V.N.; Billiard, J. The Orphan Receptor Tyrosine Kinase Ror2 Promotes Osteoblast Differentiation and Enhances Ex Vivo Bone Formation. Mol. Endocrinol. 2007, 21, 376–387. [Google Scholar] [CrossRef] [Green Version]
- Billiard, J.; Way, D.S.; Seestaller-Wehr, L.M.; Moran, R.A.; Mangine, A.; Bodine, P.V.N. The Orphan Receptor Tyrosine Kinase Ror2 Modulates Canonical Wnt Signaling in Osteoblastic Cells. Mol. Endocrinol. 2005, 19, 90–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saldanha, J.; Singh, J.; Mahadevan, D. Identification of a Frizzled-like Cysteine Rich Domain in the Extracellular Region of Developmental Receptor Tyrosine Kinases. Protein Sci. 1998, 7, 1632–1635. [Google Scholar] [CrossRef] [Green Version]
- Semënov, M.; Tamai, K.; He, X. SOST Is a Ligand for LRP5/LRP6 and a Wnt Signaling Inhibitor. J. Biol. Chem. 2005, 280, 26770–26775. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, Y.; Kang, H.; Liu, W.; Liu, P.; Zhang, J.; Harris, S.E.; Wu, D. Sclerostin Binds to LRP5/6 and Antagonizes Canonical Wnt Signaling. J. Biol. Chem. 2005, 280, 19883–19887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, B.; Wu, W.; Li, Y.; Hoppe, D.; Stannek, P.; Glinka, A.; Niehrs, C. LDL-Receptor-Related Protein 6 Is a Receptor for Dickkopf Proteins. Nature 2001, 411, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Bafico, A.; Liu, G.; Yaniv, A.; Gazit, A.; Aaronson, S.A. Novel Mechanism of Wnt Signalling Inhibition Mediated by Dickkopf-1 Interaction with LRP6/Arrow. Nat. Cell Biol. 2001, 3, 683–686. [Google Scholar] [CrossRef] [PubMed]
- Semënov, M.V.; Zhang, X.; He, X. DKK1 Antagonizes Wnt Signaling without Promotion of LRP6 Internalization and Degradation. J. Biol. Chem. 2008, 283, 21427–21432. [Google Scholar] [CrossRef] [Green Version]
- Lin, K.; Wang, S.; Julius, M.A.; Kitajewski, J.; Moos, M.; Luyten, F.P. The Cysteine-Rich Frizzled Domain of Frzb-1 Is Required and Sufficient for Modulation of Wnt Signaling. Proc. Natl. Acad. Sci. USA 1997, 94, 11196–11200. [Google Scholar] [CrossRef] [Green Version]
- Bhat, R.A.; Stauffer, B.; Komm, B.S.; Bodine, P.V.N. Structure–Function Analysis of Secreted Frizzled-Related Protein-1 for Its Wnt Antagonist Function. J. Cell. Biochem. 2007, 102, 1519–1528. [Google Scholar] [CrossRef]
- Cho, S.W.; Yang, J.-Y.; Sun, H.J.; Jung, J.Y.; Her, S.J.; Cho, H.Y.; Choi, H.J.; Kim, S.W.; Kim, S.Y.; Shin, C.S. Wnt Inhibitory Factor (WIF)-1 Inhibits Osteoblastic Differentiation in Mouse Embryonic Mesenchymal Cells. Bone 2009, 44, 1069–1077. [Google Scholar] [CrossRef]
- Mii, Y.; Taira, M. Secreted Frizzled-Related Proteins Enhance the Diffusion of Wnt Ligands and Expand Their Signalling Range. Development 2009, 136, 4083–4088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Üren, A.; Reichsman, F.; Anest, V.; Taylor, W.G.; Muraiso, K.; Bottaro, D.P.; Cumberledge, S.; Rubin, J.S. Secreted Frizzled-Related Protein-1 Binds Directly to Wingless and Is a Biphasic Modulator of Wnt Signaling. J. Biol. Chem. 2000, 275, 4374–4382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baron, R.; Kneissel, M. WNT Signaling in Bone Homeostasis and Disease: From Human Mutations to Treatments. Nat. Med. 2013, 19, 179–192. [Google Scholar] [CrossRef]
- Liu, G.; Vijayakumar, S.; Grumolato, L.; Arroyave, R.; Qiao, H.; Akiri, G.; Aaronson, S.A. Canonical Wnts Function as Potent Regulators of Osteogenesis by Human Mesenchymal Stem Cells. J. Cell Biol. 2009, 185, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Joeng, K.S.; Lee, Y.-C.; Lim, J.; Chen, Y.; Jiang, M.-M.; Munivez, E.; Ambrose, C.; Lee, B.H. Osteocyte-Specific WNT1 Regulates Osteoblast Function during Bone Homeostasis. J. Clin. Investig. 2017, 127, 2678–2688. [Google Scholar] [CrossRef] [Green Version]
- Jia, B.; Wang, Z.; Sun, X.; Chen, J.; Zhao, J.; Qiu, X. Long Noncoding RNA LINC00707 Sponges MiR-370-3p to Promote Osteogenesis of Human Bone Marrow-Derived Mesenchymal Stem Cells through Upregulating WNT2B. Stem Cell Res. 2019, 10, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mak, W.; Shao, X.; Dunstan, C.R.; Seibel, M.J.; Zhou, H. Biphasic Glucocorticoid-Dependent Regulation of Wnt Expression and Its Inhibitors in Mature Osteoblastic Cells. Calcif. Tissue Int. 2009, 85, 538–545. [Google Scholar] [CrossRef]
- Weivoda, M.M.; Ruan, M.; Hachfeld, C.M.; Pederson, L.; Howe, A.; Davey, R.A.; Zajac, J.D.; Kobayashi, Y.; Williams, B.O.; Westendorf, J.J.; et al. Wnt Signaling Inhibits Osteoclast Differentiation by Activating Canonical and Noncanonical CAMP/PKA Pathways. J. Bone Miner. Res. 2016, 31, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Boland, G.M.; Perkins, G.; Hall, D.J.; Tuan, R.S. Wnt 3a Promotes Proliferation and Suppresses Osteogenic Differentiation of Adult Human Mesenchymal Stem Cells. J. Cell. Biochem. 2004, 93, 1210–1230. [Google Scholar] [CrossRef]
- De Boer, J.; Siddappa, R.; Gaspar, C.; van Apeldoorn, A.; Fodde, R.; van Blitterswijk, C. Wnt Signaling Inhibits Osteogenic Differentiation of Human Mesenchymal Stem Cells. Bone 2004, 34, 818–826. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.H.; Kim, Y.J.; Kim, S.J.; Kim, J.H.; Bae, Y.C.; Ba, B.; Jung, J.S. Endogenous Wnt Signaling Promotes Proliferation and Suppresses Osteogenic Differentiation in Human Adipose Derived Stromal Cells. Tissue Eng. 2006, 12, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Keller, K.C.; Ding, H.; Tieu, R.; Sparks, N.R.L.; Ehnes, D.D.; Zur Nieden, N.I. Wnt5a Supports Osteogenic Lineage Decisions in Embryonic Stem Cells. Stem Cells Dev. 2016, 25, 1020–1032. [Google Scholar] [CrossRef]
- Shen, J.; Chen, X.; Jia, H.; Meyers, C.A.; Shrestha, S.; Asatrian, G.; Ding, C.; Tsuei, R.; Zhang, X.; Peault, B.; et al. Effects of WNT3A and WNT16 on the Osteogenic and Adipogenic Differentiation of Perivascular Stem/Stromal Cells. Tissue Eng. Part A 2018, 24, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Tu, X.; Joeng, K.S.; Nakayama, K.I.; Nakayama, K.; Rajagopal, J.; Carroll, T.J.; McMahon, A.P.; Long, F. Noncanonical Wnt Signaling through G Protein-Linked PKCδ Activation Promotes Bone Formation. Dev. Cell 2007, 12, 113–127. [Google Scholar] [CrossRef] [Green Version]
- Hamamura, K.; Chen, A.; Nishimura, A.; Tanjung, N.; Sudo, A.; Yokota, H. Predicting and Validating the Pathway of Wnt3a-Driven Suppression of Osteoclastogenesis. Cell. Signal. 2014, 26, 2358–2369. [Google Scholar] [CrossRef] [Green Version]
- Karner, C.M.; Esen, E.; Okunade, A.L.; Patterson, B.W.; Long, F. Increased Glutamine Catabolism Mediates Bone Anabolism in Response to WNT Signaling. J. Clin. Investig. 2015, 125, 551–562. [Google Scholar] [CrossRef] [Green Version]
- Esen, E.; Chen, J.; Karner, C.M.; Okunade, A.L.; Patterson, B.W.; Long, F. WNT-LRP5 Signaling Induces Warburg Effect through MTORC2 Activation during Osteoblast Differentiation. Cell Metab. 2013, 17, 745–755. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.-G.; Yu, S.-S.; Lee, S.-W.; Chun, J.-S. Wnt-3a Regulates Chondrocyte Differentiation via c-Jun/AP-1 Pathway. FEBS Lett. 2005, 579, 4837–4842. [Google Scholar] [CrossRef] [Green Version]
- Rossol-Allison, J.; Stemmle, L.N.; Swenson-Fields, K.I.; Kelly, P.; Fields, P.E.; McCall, S.J.; Casey, P.J.; Fields, T.A. RHO GTPASE activity modulates WNT3A/β-catenin signaling. Cell. Signal. 2009, 21, 1559–1568. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.; Sonoyama, W.; Wang, Z.; Jin, Q.; Zhang, C.; Krebsbach, P.H.; Giannobile, W.; Shi, S.; Wang, C.-Y. Noncanonical Wnt-4 Signaling Enhances Bone Regeneration of Mesenchymal Stem Cells in Craniofacial Defects through Activation of P38 MAPK. J. Biol. Chem. 2007, 282, 30938–30948. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Chang, J.; Liu, Y.; Li, J.; Kevork, K.; Al-Hezaimi, K.; Graves, D.T.; Park, N.-H.; Wang, C.-Y. Wnt4 Signaling Prevents Skeletal Aging and Inflammation by Inhibiting Nuclear Factor-ΚB. Nat. Med. 2014, 20, 1009–1017. [Google Scholar] [CrossRef] [Green Version]
- Uehara, S.; Udagawa, N.; Mukai, H.; Ishihara, A.; Maeda, K.; Yamashita, T.; Murakami, K.; Nishita, M.; Nakamura, T.; Kato, S.; et al. Protein Kinase N3 Promotes Bone Resorption by Osteoclasts in Response to Wnt5a-Ror2 Signaling. Sci. Signal. 2017, 10, eaan0023. [Google Scholar] [CrossRef] [Green Version]
- Ishitani, T.; Kishida, S.; Hyodo-Miura, J.; Ueno, N.; Yasuda, J.; Waterman, M.; Shibuya, H.; Moon, R.T.; Ninomiya-Tsuji, J.; Matsumoto, K. The TAK1-NLK Mitogen-Activated Protein Kinase Cascade Functions in the Wnt-5a/Ca2+ Pathway To Antagonize Wnt/β-Catenin Signaling. Mol. Cell. Biol. 2003, 23, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Bradley, E.W.; Drissi, M.H. WNT5A Regulates Chondrocyte Differentiation through Differential Use of the CaN/NFAT and IKK/NF-ΚB Pathways. Mol. Endocrinol. 2010, 24, 1581–1593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnsdorf, E.J.; Tummala, P.; Jacobs, C.R. Non-Canonical Wnt Signaling and N-Cadherin Related β-Catenin Signaling Play a Role in Mechanically Induced Osteogenic Cell Fate. PLoS ONE 2009, 4, e5388. [Google Scholar] [CrossRef] [Green Version]
- Bilkovski, R.; Schulte, D.M.; Oberhauser, F.; Gomolka, M.; Udelhoven, M.; Hettich, M.M.; Roth, B.; Heidenreich, A.; Gutschow, C.; Krone, W.; et al. Role of WNT-5a in the Determination of Human Mesenchymal Stem Cells into Preadipocytes. J. Biol. Chem. 2010, 285, 6170–6178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradley, E.W.; Drissi, M.H. Wnt5b Regulates Mesenchymal Cell Aggregation and Chondrocyte Differentiation through the Planar Cell Polarity Pathway. J. Cell. Physiol. 2011, 226, 1683–1693. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tu, X.; Esen, E.; Joeng, K.S.; Lin, C.; Arbeit, J.M.; Rüegg, M.A.; Hall, M.N.; Ma, L.; Long, F. WNT7B Promotes Bone Formation in Part through MTORC1. PLoS Genet. 2014, 10, e1004145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, C.N.; Ouyang, H.; Ma, Y.L.; Zeng, Q.; Gerin, I.; Sousa, K.M.; Lane, T.F.; Krishnan, V.; Hankenson, K.D.; MacDougald, O.A. Wnt10b Increases Postnatal Bone Formation by Enhancing Osteoblast Differentiation. J. Bone Miner. Res. 2007, 22, 1924–1932. [Google Scholar] [CrossRef] [PubMed]
- Bennett, C.N.; Longo, K.A.; Wright, W.S.; Suva, L.J.; Lane, T.F.; Hankenson, K.D.; MacDougald, O.A. Regulation of Osteoblastogenesis and Bone Mass by Wnt10b. Proc. Natl. Acad. Sci. USA 2005, 102, 3324–3329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cawthorn, W.P.; Bree, A.J.; Yao, Y.; Du, B.; Hemati, N.; Martinez-Santibañez, G.; MacDougald, O.A. Wnt6, Wnt10a and Wnt10b Inhibit Adipogenesis and Stimulate Osteoblastogenesis through a β-Catenin-Dependent Mechanism. Bone 2012, 50, 477–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Mak, W.; Zheng, Y.; Dunstan, C.R.; Seibel, M.J. Osteoblasts Directly Control Lineage Commitment of Mesenchymal Progenitor Cells through Wnt Signaling. J. Biol. Chem. 2008, 283, 1936–1945. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Bennett, C.N.; Gerin, I.; Rapp, L.A.; Hankenson, K.D.; MacDougald, O.A. Wnt Signaling Stimulates Osteoblastogenesis of Mesenchymal Precursors by Suppressing CCAAT/Enhancer-Binding Protein α and Peroxisome Proliferator-Activated Receptor γ. J. Biol. Chem. 2007, 282, 14515–14524. [Google Scholar] [CrossRef]
- Movérare-Skrtic, S.; Henning, P.; Liu, X.; Nagano, K.; Saito, H.; Börjesson, A.E.; Sjögren, K.; Windahl, S.H.; Farman, H.; Kindlund, B.; et al. Osteoblast-Derived WNT16 Represses Osteoclastogenesis and Prevents Cortical Bone Fragility Fractures. Nat. Med. 2014, 20, 1279–1288. [Google Scholar] [CrossRef] [Green Version]
- Kühl, M.; Sheldahl, L.C.; Park, M.; Miller, J.R.; Moon, R.T. The Wnt/Ca2+ Pathway: A New Vertebrate Wnt Signaling Pathway Takes Shape. Trends Genet. 2000, 16, 279–283. [Google Scholar] [CrossRef]
- Wang, Q.; Symes, A.J.; Kane, C.A.; Freeman, A.; Nariculam, J.; Munson, P.; Thrasivoulou, C.; Masters, J.R.W.; Ahmed, A. A Novel Role for Wnt/Ca2+ Signaling in Actin Cytoskeleton Remodeling and Cell Motility in Prostate Cancer. PLoS ONE 2010, 5, e10456. [Google Scholar] [CrossRef]
- Kühl, M.; Sheldahl, L.C.; Malbon, C.C.; Moon, R.T. Ca(2+)/Calmodulin-Dependent Protein Kinase II Is Stimulated by Wnt and Frizzled Homologs and Promotes Ventral Cell Fates in Xenopus. J. Biol. Chem. 2000, 275, 12701–12711. [Google Scholar] [CrossRef] [Green Version]
- Freisinger, C.M.; Fisher, R.A.; Slusarski, D.C. Regulator of G Protein Signaling 3 Modulates Wnt5b Calcium Dynamics and Somite Patterning. PLoS Genet. 2010, 6, e1001020. [Google Scholar] [CrossRef]
- Thrasivoulou, C.; Millar, M.; Ahmed, A. Activation of Intracellular Calcium by Multiple Wnt Ligands and Translocation of β-Catenin into the Nucleus: A convergent model OF Wnt/Ca2+ AND Wnt/β-catenin pathways. J. Biol. Chem. 2013, 288, 35651–35659. [Google Scholar] [CrossRef] [Green Version]
- Oishi, I.; Suzuki, H.; Onishi, N.; Takada, R.; Kani, S.; Ohkawara, B.; Koshida, I.; Suzuki, K.; Yamada, G.; Schwabe, G.C.; et al. The Receptor Tyrosine Kinase Ror2 Is Involved in Non-Canonical Wnt5a/JNK Signalling Pathway. Genes Cells 2003, 8, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Blumenthal, A.; Ehlers, S.; Lauber, J.; Buer, J.; Lange, C.; Goldmann, T.; Heine, H.; Brandt, E.; Reiling, N. The Wingless Homolog WNT5A and Its Receptor Frizzled-5 Regulate Inflammatory Responses of Human Mononuclear Cells Induced by Microbial Stimulation. Blood 2006, 108, 965–973. [Google Scholar] [CrossRef] [Green Version]
- Gu, Q.; Tian, H.; Zhang, K.; Chen, D.; Chen, D.; Wang, X.; Zhao, J. Wnt5a/FZD4 Mediates the Mechanical Stretch-Induced Osteogenic Differentiation of Bone Mesenchymal Stem Cells. Cell. Physiol. Biochem. 2018, 48, 215–226. [Google Scholar] [CrossRef]
- Liu, X.; Chen, W.; Zhou, Y.; Tang, K.; Zhang, J. Mechanical Tension Promotes the Osteogenic Differentiation of Rat Tendon-Derived Stem Cells Through the Wnt5a/Wnt5b/JNK Signaling Pathway. Cell. Physiol. Biochem. 2015, 36, 517–530. [Google Scholar] [CrossRef]
- Westendorf, J.J.; Kahler, R.A.; Schroeder, T.M. Wnt Signaling in Osteoblasts and Bone Diseases. Gene 2004, 341, 19–39. [Google Scholar] [CrossRef]
- Slusarski, D.C.; Yang-Snyder, J.; Busa, W.B.; Moon, R.T. Modulation of Embryonic Intracellular Ca2+ Signaling by Wnt-5A. Dev. Biol. 1997, 182, 114–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slusarski, D.C.; Corces, V.G.; Moon, R.T. Interaction of Wnt and a Frizzled Homologue Triggers G-Protein-Linked Phosphatidylinositol Signalling. Nature 1997, 390, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Sheldahl, L.C.; Park, M.; Malbon, C.C.; Moon, R.T. Protein Kinase C Is Differentially Stimulated by Wnt and Frizzled Homologs in AG-Protein-Dependent Manner. Curr. Biol. 1999, 9, 695–698. [Google Scholar] [CrossRef] [Green Version]
- Clapham, D.E. Calcium Signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batoon, L.; Millard, S.M.; Raggatt, L.J.; Pettit, A.R. Osteomacs and Bone Regeneration. Curr. Osteoporos. Rep. 2017, 15, 385–395. [Google Scholar] [CrossRef]
- Sinder, B.P.; Pettit, A.R.; McCauley, L.K. Macrophages: Their Emerging Roles in Bone. J. Bone Min. Res. 2015, 30, 2140–2149. [Google Scholar] [CrossRef] [Green Version]
- Butler, M.T.; Wallingford, J.B. Planar Cell Polarity in Development and Disease. Nat. Rev. Mol. Cell Biol. 2017, 18, 375–388. [Google Scholar] [CrossRef]
- Yang, Y.; Mlodzik, M. Wnt-Frizzled/Planar Cell Polarity Signaling: Cellular Orientation by Facing the Wind (Wnt). Annu. Rev. Cell Dev. Biol. 2015, 31, 623–646. [Google Scholar] [CrossRef] [Green Version]
- Sebbagh, M.; Borg, J.-P. Insight into Planar Cell Polarity. Exp. Cell Res. 2014, 328, 284–295. [Google Scholar] [CrossRef]
- Gao, B.; Song, H.; Bishop, K.; Elliot, G.; Garrett, L.; English, M.; Andre, P.; Robinson, J.; Sood, R.; Minami, Y.; et al. Wnt Signaling Gradients Establish Planar Cell Polarity by Inducing Vangl2 Phosphorylation through Ror2. Dev. Cell 2011, 20, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Andre, P.; Wang, Q.; Wang, N.; Gao, B.; Schilit, A.; Halford, M.M.; Stacker, S.A.; Zhang, X.; Yang, Y. The Wnt Coreceptor Ryk Regulates Wnt/Planar Cell Polarity by Modulating the Degradation of the Core Planar Cell Polarity Component Vangl2. J. Biol. Chem. 2012, 287, 44518–44525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macheda, M.L.; Sun, W.W.; Kugathasan, K.; Hogan, B.M.; Bower, N.I.; Halford, M.M.; Zhang, Y.F.; Jacques, B.E.; Lieschke, G.J.; Dabdoub, A.; et al. The Wnt Receptor Ryk Plays a Role in Mammalian Planar Cell Polarity Signaling. J. Biol. Chem. 2012, 287, 29312–29323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, H.-Y.H.; Susman, M.W.; Bikoff, J.B.; Ryu, Y.K.; Jonas, A.M.; Hu, L.; Kuruvilla, R.; Greenberg, M.E. Wnt5a–Ror–Dishevelled Signaling Constitutes a Core Developmental Pathway That Controls Tissue Morphogenesis. Proc. Natl. Acad. Sci. USA 2012, 109, 4044–4051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Struhl, G.; Casal, J.; Lawrence, P.A. Dissecting the Molecular Bridges That Mediate the Function of Frizzled in Planar Cell Polarity. Development 2012, 139, 3665–3674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.-S.; Antic, D.; Matis, M.; Logan, C.Y.; Povelones, M.; Anderson, G.A.; Nusse, R.; Axelrod, J.D. Asymmetric Homotypic Interactions of the Atypical Cadherin Flamingo Mediate Intercellular Polarity Signaling. Cell 2008, 133, 1093–1105. [Google Scholar] [CrossRef] [Green Version]
- Gao, B. Chapter Eleven—Wnt Regulation of Planar Cell Polarity (PCP). In Planar Cell Polarity During Development; Current Topics in Developmental Biology; Yang, Y., Ed.; Academic Press: Cambridge, MA, USA, 2012; Volume 101, pp. 263–295. [Google Scholar]
- Wang, B.; Sinha, T.; Jiao, K.; Serra, R.; Wang, J. Disruption of PCP Signaling Causes Limb Morphogenesis and Skeletal Defects and May Underlie Robinow Syndrome and Brachydactyly Type B. Hum. Mol. Genet. 2011, 20, 271–285. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Dudley, A.T. Noncanonical Frizzled Signaling Regulates Cell Polarity of Growth Plate Chondrocytes. Development 2009, 136, 1083–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavin, B.J.; McMahon, J.A.; McMahon, A.P. Expression of Multiple Novel Wnt-1/Int-1-Related Genes during Fetal and Adult Mouse Development. Genes Dev. 1990, 4, 2319–2332. [Google Scholar] [CrossRef]
- Yamaguchi, T.P.; Bradley, A.; McMahon, A.P.; Jones, S. A Wnt5a Pathway Underlies Outgrowth of Multiple Structures in the Vertebrate Embryo. Development 1999, 126, 1211–1223. [Google Scholar] [CrossRef] [PubMed]
- Hopyan, S.; Sharpe, J.; Yang, Y. Budding Behaviors: Growth of the Limb as a Model of Morphogenesis. Dev. Dyn. 2011, 240, 1054–1062. [Google Scholar] [CrossRef] [PubMed]
- Galea, G.L.; Meakin, L.B.; Savery, D.; Taipaleenmaki, H.; Delisser, P.; Stein, G.S.; Copp, A.J.; van Wijnen, A.J.; Lanyon, L.E.; Price, J.S. Planar Cell Polarity Aligns Osteoblast Division in Response to Substrate Strain. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2015, 30, 423–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, Y.; Lantz, B.; Cusack, B.J.; Szabo-Rogers, H.L. Prickle1 Regulates Differentiation of Frontal Bone Osteoblasts. Sci. Rep. 2018, 8, 18021. [Google Scholar] [CrossRef] [PubMed]
- Bikkavilli, R.K.; Feigin, M.E.; Malbon, C.C. Gαo Mediates WNT-JNK Signaling through Dishevelled 1 and 3, RhoA Family Members, and MEKK 1 and 4 in Mammalian Cells. J. Cell Sci. 2008, 121, 234–245. [Google Scholar] [CrossRef] [Green Version]
- Ridley, A.J.; Hall, A. The Small GTP-Binding Protein Rho Regulates the Assembly of Focal Adhesions and Actin Stress Fibers in Response to Growth Factors. Cell 1992, 70, 389–399. [Google Scholar] [CrossRef]
- McBeath, R.; Pirone, D.M.; Nelson, C.M.; Bhadriraju, K.; Chen, C.S. Cell Shape, Cytoskeletal Tension, and RhoA Regulate Stem Cell Lineage Commitment. Dev. Cell 2004, 6, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Ridley, A.J.; Paterson, H.F.; Johnston, C.L.; Diekmann, D.; Hall, A. The Small GTP-Binding Protein Rac Regulates Growth Factor-Induced Membrane Ruffling. Cell 1992, 70, 401–410. [Google Scholar] [CrossRef]
- Santos, A.; Bakker, A.D.; de Blieck-Hogervorst, J.M.A.; Klein-Nulend, J. WNT5A Induces Osteogenic Differentiation of Human Adipose Stem Cells via Rho-Associated Kinase Rock. Cytotherapy 2010, 12, 924–932. [Google Scholar] [CrossRef] [PubMed]
- Kassem, M.; Marie, P.J. Senescence-Associated Intrinsic Mechanisms of Osteoblast Dysfunctions. Aging Cell 2011, 10, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Udagawa, N.; Nakamura, I.; Murakami, H.; Saito, S.; Yamasaki, K.; Shibasaki, Y.; Morii, N.; Narumiya, S.; Takahashi, N. The Small GTP-Binding Protein, Rho P21, Is Involved in Bone Resorption by Regulating Cytoskeletal Organization in Osteoclasts. J. Cell Sci. 1995, 108, 2285–2292. [Google Scholar] [CrossRef]
- Chellaiah, M.A.; Soga, N.; Swanson, S.; McAllister, S.; Alvarez, U.; Wang, D.; Dowdy, S.F.; Hruska, K.A. Rho-A Is Critical for Osteoclast Podosome Organization, Motility, and Bone Resorption. J. Biol. Chem. 2000, 275, 11993–12002. [Google Scholar] [CrossRef] [Green Version]
- Ory, S.; Munari-Silem, Y.; Fort, P.; Jurdic, P. Rho and Rac Exert Antagonistic Functions on Spreading of Macrophage-Derived Multinucleated Cells and Are Not Required for Actin Fiber Formation. J. Cell Sci. 2000, 113, 1177–1188. [Google Scholar] [CrossRef]
- Thompson, M.; Nejak-Bowen, K.; Monga, S.P.S. Crosstalk of the Wnt Signaling Pathway. In Targeting the Wnt Pathway in Cancer; Goss, K.H., Kahn, M., Eds.; Springer: New York, NY, USA, 2011; pp. 51–80. ISBN 978-1-4419-8023-6. [Google Scholar]
- Itasaki, N.; Hoppler, S. Crosstalk between Wnt and Bone Morphogenic Protein Signaling: A Turbulent Relationship. Dev. Dyn. 2010, 239, 16–33. [Google Scholar] [CrossRef]
- Nemoto, E.; Ebe, Y.; Kanaya, S.; Tsuchiya, M.; Nakamura, T.; Tamura, M.; Shimauchi, H. Wnt5a Signaling Is a Substantial Constituent in Bone Morphogenetic Protein-2-Mediated Osteoblastogenesis. Biochem. Biophys. Res. Commun. 2012, 422, 627–632. [Google Scholar] [CrossRef]
- Lee, S.; Remark, L.H.; Josephson, A.M.; Leclerc, K.; Lopez, E.M.; Kirby, D.J.; Mehta, D.; Litwa, H.P.; Wong, M.Z.; Shin, S.Y.; et al. Notch-Wnt Signal Crosstalk Regulates Proliferation and Differentiation of Osteoprogenitor Cells during Intramembranous Bone Healing. NPJ Regen. Med. 2021, 6, 29. [Google Scholar] [CrossRef]
- Hu, H.; Hilton, M.J.; Tu, X.; Yu, K.; Ornitz, D.M.; Long, F. Sequential Roles of Hedgehog and Wnt Signaling in Osteoblast Development. Development 2005, 132, 49–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mak, K.K.; Bi, Y.; Wan, C.; Chuang, P.-T.; Clemens, T.; Young, M.; Yang, Y. Hedgehog Signaling in Mature Osteoblasts Regulates Bone Formation and Resorption by Controlling PTHrP and RANKL Expression. Dev. Cell 2008, 14, 674–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boonanantanasarn, K.; Lee, H.-L.; Baek, K.; Woo, K.M.; Ryoo, H.-M.; Baek, J.-H.; Kim, G.-S. EGF Inhibits Wnt/β-Catenin-Induced Osteoblast Differentiation by Promoting β-Catenin Degradation. J. Cell. Biochem. 2015, 116, 2849–2857. [Google Scholar] [CrossRef]
- Wada, T.; Nakashima, T.; Hiroshi, N.; Penninger, J.M. RANKL–RANK Signaling in Osteoclastogenesis and Bone Disease. Trends Mol. Med. 2006, 12, 17–25. [Google Scholar] [CrossRef]
- Boyce, B.F.; Xing, L. Biology of RANK, RANKL, and Osteoprotegerin. Arthritis Res. 2007, 9, S1. [Google Scholar] [CrossRef] [Green Version]
- Kieslinger, M.; Folberth, S.; Dobreva, G.; Dorn, T.; Croci, L.; Erben, R.; Consalez, G.G.; Grosschedl, R. EBF2 Regulates Osteoblast-Dependent Differentiation of Osteoclasts. Dev. Cell 2005, 9, 757–767. [Google Scholar] [CrossRef] [Green Version]
- Boyce, B.F.; Xing, L.; Chen, D. Osteoprotegerin, the Bone Protector, Is a Surprising Target for β-Catenin Signaling. Cell Metab. 2005, 2, 344–345. [Google Scholar] [CrossRef] [Green Version]
- Spencer, G.J.; Utting, J.C.; Etheridge, S.L.; Arnett, T.R.; Genever, P.G. Wnt Signalling in Osteoblasts Regulates Expression of the Receptor Activator of NFkappaB Ligand and Inhibits Osteoclastogenesis in Vitro. J. Cell Sci. 2006, 119, 1283–1296. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Wang, H.; Huang, T.; Wang, J.; Ding, Y.; Li, Z.; Zhang, J.; Li, L. TAB2 Scaffolds TAK1 and NLK in Repressing Canonical Wnt Signaling. J. Biol. Chem. 2010, 285, 13397–13404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxton, R.A.; Sabatini, D.M. MTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, R.C.; Fang, C.; Guan, K.-L. An Emerging Role for TOR Signaling in Mammalian Tissue and Stem Cell Physiology. Development 2011, 138, 3343–3356. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Long, F. MTORC1 Signaling Promotes Osteoblast Differentiation from Preosteoblasts. PLoS ONE 2015, 10, e0130627. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Holguin, N.; Shi, Y.; Silva, M.J.; Long, F. MTORC2 Signaling Promotes Skeletal Growth and Bone Formation in Mice. J. Bone Min. Res. 2015, 30, 369–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Long, F. MTORC1 Signaling Controls Mammalian Skeletal Growth through Stimulation of Protein Synthesis. Development 2014, 141, 2848–2854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.; Liu, L.; Zeng, C.; Cui, Z.-K.; Chen, Y.; Lai, P.; Wang, H.; Shao, Y.; Zhang, H.; Zhang, R.; et al. Activation of MTORC1 in Subchondral Bone Preosteoblasts Promotes Osteoarthritis by Stimulating Bone Sclerosis and Secretion of CXCL12. Bone Res. 2019, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Ji, X.; Lee, W.-C.; Shi, Y.; Li, B.; Abel, E.D.; Jiang, D.; Huang, W.; Long, F. Increased Glycolysis Mediates Wnt7b-Induced Bone Formation. FASEB J. 2019, 33, 7810–7821. [Google Scholar] [CrossRef]
- Lee, W.-C.; Guntur, A.R.; Long, F.; Rosen, C.J. Energy Metabolism of the Osteoblast: Implications for Osteoporosis. Endocr. Rev. 2017, 38, 255–266. [Google Scholar] [CrossRef]
- Yan, B.; Zhang, Z.; Jin, D.; Cai, C.; Jia, C.; Liu, W.; Wang, T.; Li, S.; Zhang, H.; Huang, B.; et al. MTORC1 Regulates PTHrP to Coordinate Chondrocyte Growth, Proliferation and Differentiation. Nat. Commun. 2016, 7, 11151. [Google Scholar] [CrossRef] [Green Version]
- Xiang, X.; Zhao, J.; Xu, G.; Li, Y.; Zhang, W. MTOR and the Differentiation of Mesenchymal Stem Cells. Acta Biochim. Biophys. Sin. 2011, 43, 501–510. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Xu, S.; Li, K.; Tan, K.; Liang, K.; Wang, J.; Shen, J.; Zou, W.; Hu, L.; Cai, D.; et al. MTORC1 Inhibits NF-ΚB/NFATc1 Signaling and Prevents Osteoclast Precursor Differentiation, In Vitro and In Mice. J. Bone Miner. Res. 2017, 32, 1829–1840. [Google Scholar] [CrossRef]
- Singha, U.K.; Jiang, Y.; Yu, S.; Luo, M.; Lu, Y.; Zhang, J.; Xiao, G. Rapamycin Inhibits Osteoblast Proliferation and Differentiation in MC3T3-E1 Cells and Primary Mouse Bone Marrow Stromal Cells. J. Cell. Biochem. 2008, 103, 434–446. [Google Scholar] [CrossRef]
- Lee, K.-W.; Yook, J.-Y.; Son, M.-Y.; Kim, M.-J.; Koo, D.-B.; Han, Y.-M.; Cho, Y.S. Rapamycin Promotes the Osteoblastic Differentiation of Human Embryonic Stem Cells by Blocking the MTOR Pathway and Stimulating the BMP/Smad Pathway. Stem Cells Dev. 2010, 19, 557–568. [Google Scholar] [CrossRef]
- Kichaev, G.; Bhatia, G.; Loh, P.-R.; Gazal, S.; Burch, K.; Freund, M.K.; Schoech, A.; Pasaniuc, B.; Price, A.L. Leveraging Polygenic Functional Enrichment to Improve GWAS Power. Am. J. Hum. Genet. 2019, 104, 65–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemp, J.P.; Morris, J.A.; Medina-Gomez, C.; Forgetta, V.; Warrington, N.M.; Youlten, S.E.; Zheng, J.; Gregson, C.L.; Grundberg, E.; Trajanoska, K.; et al. Identification of 153 New Loci Associated with Heel Bone Mineral Density and Functional Involvement of GPC6 in Osteoporosis. Nat. Genet. 2017, 49, 1468–1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Zhao, M.; Xie, Z.-G.; Liu, J.; Peng, H.-P.; Pei, Y.-F.; Sun, H.-P.; Zhang, L. Twelve New Genomic Loci Associated With Bone Mineral Density. Front. Endocrinol. 2020, 11, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.K. Identification of 613 New Loci Associated with Heel Bone Mineral Density and a Polygenic Risk Score for Bone Mineral Density, Osteoporosis and Fracture. PLoS ONE 2018, 13, e0200785. [Google Scholar] [CrossRef] [Green Version]
- Karner, C.M.; Esen, E.; Chen, J.; Hsu, F.-F.; Turk, J.; Long, F. Wnt Protein Signaling Reduces Nuclear Acetyl-CoA Levels to Suppress Gene Expression during Osteoblast Differentiation. J. Biol. Chem. 2016, 291, 13028–13039. [Google Scholar] [CrossRef] [Green Version]
- Dai, Q.; Xu, Z.; Ma, X.; Niu, N.; Zhou, S.; Xie, F.; Jiang, L.; Wang, J.; Zou, W. MTOR/Raptor Signaling Is Critical for Skeletogenesis in Mice through the Regulation of Runx2 Expression. Cell Death Differ. 2017, 24, 1886–1899. [Google Scholar] [CrossRef]
- Martin, S.K.; Fitter, S.; Dutta, A.K.; Matthews, M.P.; Walkley, C.R.; Hall, M.N.; Ruegg, M.A.; Gronthos, S.; Zannettino, A.C.W. Brief Report: The Differential Roles of MTORC1 and MTORC2 in Mesenchymal Stem Cell Differentiation. Stem Cells 2015, 33, 1359–1365. [Google Scholar] [CrossRef] [Green Version]
- Zeng, H.; Lu, B.; Zamponi, R.; Yang, Z.; Wetzel, K.; Loureiro, J.; Mohammadi, S.; Beibel, M.; Bergling, S.; Reece-Hoyes, J.; et al. MTORC1 Signaling Suppresses Wnt/β-Catenin Signaling through DVL-Dependent Regulation of Wnt Receptor FZD Level. Proc. Natl. Acad. Sci. USA 2018, 115, E10362–E10369. [Google Scholar] [CrossRef] [Green Version]
- Yu, A.; Rual, J.-F.; Tamai, K.; Harada, Y.; Vidal, M.; He, X.; Kirchhausen, T. Association of Dishevelled with the Clathrin AP-2 Adaptor Is Required for Frizzled Endocytosis and Planar Cell Polarity Signaling. Dev. Cell 2007, 12, 129–141. [Google Scholar] [CrossRef] [Green Version]
- Dupont, S.; Morsut, L.; Aragona, M.; Enzo, E.; Giulitti, S.; Cordenonsi, M.; Zanconato, F.; Le Digabel, J.; Forcato, M.; Bicciato, S.; et al. Role of YAP/TAZ in Mechanotransduction. Nature 2011, 474, 179–183. [Google Scholar] [CrossRef]
- Halder, G.; Dupont, S.; Piccolo, S. Transduction of Mechanical and Cytoskeletal Cues by YAP and TAZ. Nat. Rev. Mol. Cell Biol. 2012, 13, 591–600. [Google Scholar] [CrossRef]
- Varelas, X.; Sakuma, R.; Samavarchi-Tehrani, P.; Peerani, R.; Rao, B.M.; Dembowy, J.; Yaffe, M.B.; Zandstra, P.W.; Wrana, J.L. TAZ Controls Smad Nucleocytoplasmic Shuttling and Regulates Human Embryonic Stem-Cell Self-Renewal. Nat. Cell Biol. 2008, 10, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Alarcón, C.; Zaromytidou, A.-I.; Xi, Q.; Gao, S.; Yu, J.; Fujisawa, S.; Barlas, A.; Miller, A.N.; Manova-Todorova, K.; Macias, M.J.; et al. Nuclear CDKs Drive Smad Transcriptional Activation and Turnover in BMP and TGF-Beta Pathways. Cell 2009, 139, 757–769. [Google Scholar] [CrossRef] [Green Version]
- Azzolin, L.; Panciera, T.; Soligo, S.; Enzo, E.; Bicciato, S.; Dupont, S.; Bresolin, S.; Frasson, C.; Basso, G.; Guzzardo, V.; et al. YAP/TAZ Incorporation in the β-Catenin Destruction Complex Orchestrates the Wnt Response. Cell 2014, 158, 157–170. [Google Scholar] [CrossRef] [Green Version]
- Azzolin, L.; Zanconato, F.; Bresolin, S.; Forcato, M.; Basso, G.; Bicciato, S.; Cordenonsi, M.; Piccolo, S. Role of TAZ as Mediator of Wnt Signaling. Cell 2012, 151, 1443–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byun, M.R.; Hwang, J.-H.; Kim, A.R.; Kim, K.M.; Hwang, E.S.; Yaffe, M.B.; Hong, J.-H. Canonical Wnt Signalling Activates TAZ through PP1A during Osteogenic Differentiation. Cell Death Differ. 2014, 21, 854–863. [Google Scholar] [CrossRef] [Green Version]
- Piccolo, S.; Dupont, S.; Cordenonsi, M. The Biology of YAP/TAZ: Hippo Signaling and Beyond. Physiol. Rev. 2014, 94, 1287–1312. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, S.K.; Sullivan, A.J.; Medina, R.; Ito, Y.; van Wijnen, A.J.; Stein, J.L.; Lian, J.B.; Stein, G.S. Tyrosine Phosphorylation Controls Runx2-Mediated Subnuclear Targeting of YAP to Repress Transcription. EMBO J. 2004, 23, 790–799. [Google Scholar] [CrossRef]
- Hong, J.-H.; Hwang, E.S.; McManus, M.T.; Amsterdam, A.; Tian, Y.; Kalmukova, R.; Mueller, E.; Benjamin, T.; Spiegelman, B.M.; Sharp, P.A.; et al. TAZ, a Transcriptional Modulator of Mesenchymal Stem Cell Differentiation. Science 2005, 309, 1074–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, J.-X.; Xiong, L.; Zhao, K.; Zeng, P.; Wang, B.; Tang, F.-L.; Sun, D.; Guo, H.; Yang, X.; Cui, S.; et al. YAP Promotes Osteogenesis and Suppresses Adipogenic Differentiation by Regulating β-Catenin Signaling. Bone Res. 2018, 6, 18. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Han, W.; Qin, A.; Wang, Z.; Xu, J.; Qian, Y. The Emerging Role of Hippo Signaling Pathway in Regulating Osteoclast Formation. J. Cell. Physiol. 2018, 233, 4606–4617. [Google Scholar] [CrossRef]
- Varelas, X.; Miller, B.W.; Sopko, R.; Song, S.; Gregorieff, A.; Fellouse, F.A.; Sakuma, R.; Pawson, T.; Hunziker, W.; McNeill, H.; et al. The Hippo Pathway Regulates Wnt/Beta-Catenin Signaling. Dev. Cell 2010, 18, 579–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, F.; Peng, L.; Li, Z.; Tan, G.; Liang, E.; Chen, S.; Zhao, X.; Zhi, F. YAP Triggers the Wnt/β-Catenin Signalling Pathway and Promotes Enterocyte Self-Renewal, Regeneration and Tumorigenesis after DSS-Induced Injury. Cell Death Dis. 2018, 9, 1–16. [Google Scholar] [CrossRef]
- Park, H.W.; Kim, Y.C.; Yu, B.; Moroishi, T.; Mo, J.-S.; Plouffe, S.W.; Meng, Z.; Lin, K.C.; Yu, F.-X.; Alexander, C.M.; et al. Alternative Wnt Signaling Activates YAP/TAZ. Cell 2015, 162, 780–794. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Carballo, E.; Gámez, B.; Ventura, F. P38 MAPK Signaling in Osteoblast Differentiation. Front. Cell Dev. Biol. 2016, 4, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaiswal, R.K.; Jaiswal, N.; Bruder, S.P.; Mbalaviele, G.; Marshak, D.R.; Pittenger, M.F. Adult Human Mesenchymal Stem Cell Differentiation to the Osteogenic or Adipogenic Lineage Is Regulated by Mitogen-Activated Protein Kinase. J. Biol. Chem. 2000, 275, 9645–9652. [Google Scholar] [CrossRef] [Green Version]
- Gallea, S.; Lallemand, F.; Atfi, A.; Rawadi, G.; Ramez, V.; Spinella-Jaegle, S.; Kawai, S.; Faucheu, C.; Huet, L.; Baron, R.; et al. Activation of Mitogen-Activated Protein Kinase Cascades Is Involved in Regulation of Bone Morphogenetic Protein-2-Induced Osteoblast Differentiation in Pluripotent C2C12 Cells. Bone 2001, 28, 491–498. [Google Scholar] [CrossRef]
- Guicheux, J.; Lemonnier, J.; Ghayor, C.; Suzuki, A.; Palmer, G.; Caverzasio, J. Activation of P38 Mitogen-Activated Protein Kinase and c-Jun-NH2-Terminal Kinase by BMP-2 and Their Implication in the Stimulation of Osteoblastic Cell Differentiation. J. Bone Min. Res. 2003, 18, 2060–2068. [Google Scholar] [CrossRef]
- Suzuki, A.; Palmer, G.; Bonjour, J.P.; Caverzasio, J. Regulation of Alkaline Phosphatase Activity by P38 MAP Kinase in Response to Activation of Gi Protein-Coupled Receptors by Epinephrine in Osteoblast-like Cells. Endocrinology 1999, 140, 3177–3182. [Google Scholar] [CrossRef]
- Yun, M.-S.; Kim, S.-E.; Jeon, S.H.; Lee, J.-S.; Choi, K.-Y. Both ERK and Wnt/Beta-Catenin Pathways Are Involved in Wnt3a-Induced Proliferation. J. Cell Sci. 2005, 118, 313–322. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, A.; Guicheux, J.; Palmer, G.; Miura, Y.; Oiso, Y.; Bonjour, J.P.; Caverzasio, J. Evidence for a Role of P38 MAP Kinase in Expression of Alkaline Phosphatase during Osteoblastic Cell Differentiation. Bone 2002, 30, 91–98. [Google Scholar] [CrossRef]
- Zhang, Y.; Pizzute, T.; Pei, M. A Review of Crosstalk between MAPK and Wnt Signals and Its Impact on Cartilage Regeneration. Cell Tissue Res. 2014, 358, 633–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, W.-J.; Yoon, J.; Park, J.-C.; Lee, S.-H.; Lee, S.-H.; Kaduwal, S.; Kim, H.; Yoon, J.-B.; Choi, K.-Y. Ras Stabilization through Aberrant Activation of Wnt/β-Catenin Signaling Promotes Intestinal Tumorigenesis. Sci. Signal. 2012, 5, ra30. [Google Scholar] [CrossRef] [PubMed]
- Červenka, I.; Wolf, J.; Mašek, J.; Krejci, P.; Wilcox, W.R.; Kozubík, A.; Schulte, G.; Gutkind, J.S.; Bryja, V. Mitogen-Activated Protein Kinases Promote WNT/Beta-Catenin Signaling via Phosphorylation of LRP6. Mol. Cell Biol. 2011, 31, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Haÿ, E.; Nouraud, A.; Marie, P.J. N-Cadherin Negatively Regulates Osteoblast Proliferation and Survival by Antagonizing Wnt, ERK and PI3K/Akt Signalling. PLoS ONE 2009, 4, e8284. [Google Scholar] [CrossRef]
- Koyama, H.; Nakade, O.; Takada, Y.; Kaku, T.; Lau, K.H.W. Melatonin at Pharmacologic Doses Increases Bone Mass by Suppressing Resorption through Down-Regulation of the RANKL-Mediated Osteoclast Formation and Activation. J. Bone Min. Res. 2002, 17, 1219–1229. [Google Scholar] [CrossRef] [PubMed]
- Satomura, K.; Tobiume, S.; Tokuyama, R.; Yamasaki, Y.; Kudoh, K.; Maeda, E.; Nagayama, M. Melatonin at Pharmacological Doses Enhances Human Osteoblastic Differentiation in Vitro and Promotes Mouse Cortical Bone Formation in Vivo. J. Pineal Res. 2007, 42, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Roth, J.A.; Kim, B.G.; Lin, W.L.; Cho, M.I. Melatonin Promotes Osteoblast Differentiation and Bone Formation. J. Biol. Chem. 1999, 274, 22041–22047. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Li, Z.; Wang, J.; Li, Z.; Cui, H.; Dai, G.; Chen, S.; Zhang, M.; Zheng, Z.; Zhan, Z.; et al. Wnt4 Signaling Mediates Protective Effects of Melatonin on New Bone Formation in an Inflammatory Environment. FASEB J. 2019, 33, 10126–10139. [Google Scholar] [CrossRef] [Green Version]
- Radio, N.M.; Doctor, J.S.; Witt-Enderby, P.A. Melatonin Enhances Alkaline Phosphatase Activity in Differentiating Human Adult Mesenchymal Stem Cells Grown in Osteogenic Medium via MT2 Melatonin Receptors and the MEK/ERK (1/2) Signaling Cascade. J. Pineal Res. 2006, 40, 332–342. [Google Scholar] [CrossRef]
- Park, K.-H.; Kang, J.W.; Lee, E.-M.; Kim, J.S.; Rhee, Y.H.; Kim, M.; Jeong, S.J.; Park, Y.G.; Kim, S.H. Melatonin Promotes Osteoblastic Differentiation through the BMP/ERK/Wnt Signaling Pathways. J. Pineal Res. 2011, 51, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Afzal, A.R.; Jeffery, S. One Gene, Two Phenotypes: ROR2 Mutations in Autosomal Recessive Robinow Syndrome and Autosomal Dominant Brachydactyly Type B. Hum. Mutat. 2003, 22, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Schwarzer, W.; Witte, F.; Rajab, A.; Mundlos, S.; Stricker, S. A Gradient of ROR2 Protein Stability and Membrane Localization Confers Brachydactyly Type B or Robinow Syndrome Phenotypes. Hum. Mol. Genet. 2009, 18, 4013–4021. [Google Scholar] [CrossRef] [Green Version]
- White, J.J.; Mazzeu, J.F.; Coban-Akdemir, Z.; Bayram, Y.; Bahrambeigi, V.; Hoischen, A.; van Bon, B.W.M.; Gezdirici, A.; Gulec, E.Y.; Ramond, F.; et al. WNT Signaling Perturbations Underlie the Genetic Heterogeneity of Robinow Syndrome. Am. J. Hum. Genet. 2018, 102, 27–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Person, A.D.; Beiraghi, S.; Sieben, C.M.; Hermanson, S.; Neumann, A.N.; Robu, M.E.; Schleiffarth, J.R.; Billington, C.J.; van Bokhoven, H.; Hoogeboom, J.M.; et al. WNT5A Mutations in Patients with Autosomal Dominant Robinow Syndrome. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2010, 239, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Kiper, P.O.S.; Saito, H.; Gori, F.; Unger, S.; Hesse, E.; Yamana, K.; Kiviranta, R.; Solban, N.; Liu, J.; Brommage, R.; et al. Cortical-Bone Fragility—Insights from SFRP4 Deficiency in Pyle’s Disease. N. Engl. J. Med. 2016, 374, 2553–2562. [Google Scholar] [CrossRef] [PubMed]
- Arboleya, L.; Queiro, R.; Alperi, M.; Lorenzo, J.A.; Ballina, J. Pyle’s Disease: A Human Model of Differentiated Cortical and Trabecular Homeostasis. Reum. Clin. 2020, 16, 56–58. [Google Scholar] [CrossRef]
- Chen, K.; Ng, P.Y.; Chen, R.; Hu, D.; Berry, S.; Baron, R.; Gori, F. Sfrp4 Repression of the Ror2/Jnk Cascade in Osteoclasts Protects Cortical Bone from Excessive Endosteal Resorption. Proc. Natl. Acad. Sci. USA 2019, 116, 14138–14143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodine, P.V.N.; Stauffer, B.; Ponce-de-Leon, H.; Bhat, R.A.; Mangine, A.; Seestaller-Wehr, L.M.; Moran, R.A.; Billiard, J.; Fukayama, S.; Komm, B.S.; et al. A Small Molecule Inhibitor of the Wnt Antagonist Secreted Frizzled-Related Protein-1 Stimulates Bone Formation. Bone 2009, 44, 1063–1068. [Google Scholar] [CrossRef]
- Moore, W.J.; Kern, J.C.; Bhat, R.; Commons, T.J.; Fukayama, S.; Goljer, I.; Krishnamurthy, G.; Magolda, R.L.; Nogle, L.; Pitts, K.; et al. Modulation of Wnt Signaling through Inhibition of Secreted Frizzled-Related Protein I (SFRP-1) with N-Substituted Piperidinyl Diphenylsulfonyl Sulfonamides. J. Med. Chem. 2009, 52, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Moore, W.J.; Kern, J.C.; Bhat, R.; Bodine, P.V.N.; Fukyama, S.; Krishnamurthy, G.; Magolda, R.L.; Pitts, K.; Stauffer, B.; Trybulski, E.J. Modulation of Wnt Signaling through Inhibition of Secreted Frizzled-Related Protein I (SFRP-1) with N-Substituted Piperidinyl Diphenylsulfonyl Sulfonamides: Part II. Bioorg. Med. Chem. 2010, 18, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Bodine, P.V.N.; Zhao, W.; Kharode, Y.P.; Bex, F.J.; Lambert, A.-J.; Goad, M.B.; Gaur, T.; Stein, G.S.; Lian, J.B.; Komm, B.S. The Wnt Antagonist Secreted Frizzled-Related Protein-1 Is a Negative Regulator of Trabecular Bone Formation in Adult Mice. Mol. Endocrinol. 2004, 18, 1222–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bovolenta, P.; Esteve, P.; Ruiz, J.M.; Cisneros, E.; Lopez-Rios, J. Beyond Wnt Inhibition: New Functions of Secreted Frizzled-Related Proteins in Development and Disease. J. Cell Sci. 2008, 121, 737–746. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lojk, J.; Marc, J. Roles of Non-Canonical Wnt Signalling Pathways in Bone Biology. Int. J. Mol. Sci. 2021, 22, 10840. https://doi.org/10.3390/ijms221910840
Lojk J, Marc J. Roles of Non-Canonical Wnt Signalling Pathways in Bone Biology. International Journal of Molecular Sciences. 2021; 22(19):10840. https://doi.org/10.3390/ijms221910840
Chicago/Turabian StyleLojk, Jasna, and Janja Marc. 2021. "Roles of Non-Canonical Wnt Signalling Pathways in Bone Biology" International Journal of Molecular Sciences 22, no. 19: 10840. https://doi.org/10.3390/ijms221910840
APA StyleLojk, J., & Marc, J. (2021). Roles of Non-Canonical Wnt Signalling Pathways in Bone Biology. International Journal of Molecular Sciences, 22(19), 10840. https://doi.org/10.3390/ijms221910840