
Adaptive Mechanisms of Halophytes and Their Potential in Improving Salinity Tolerance in Plants

 ,
,  , , and
, , and
Abstract
:1. Introduction
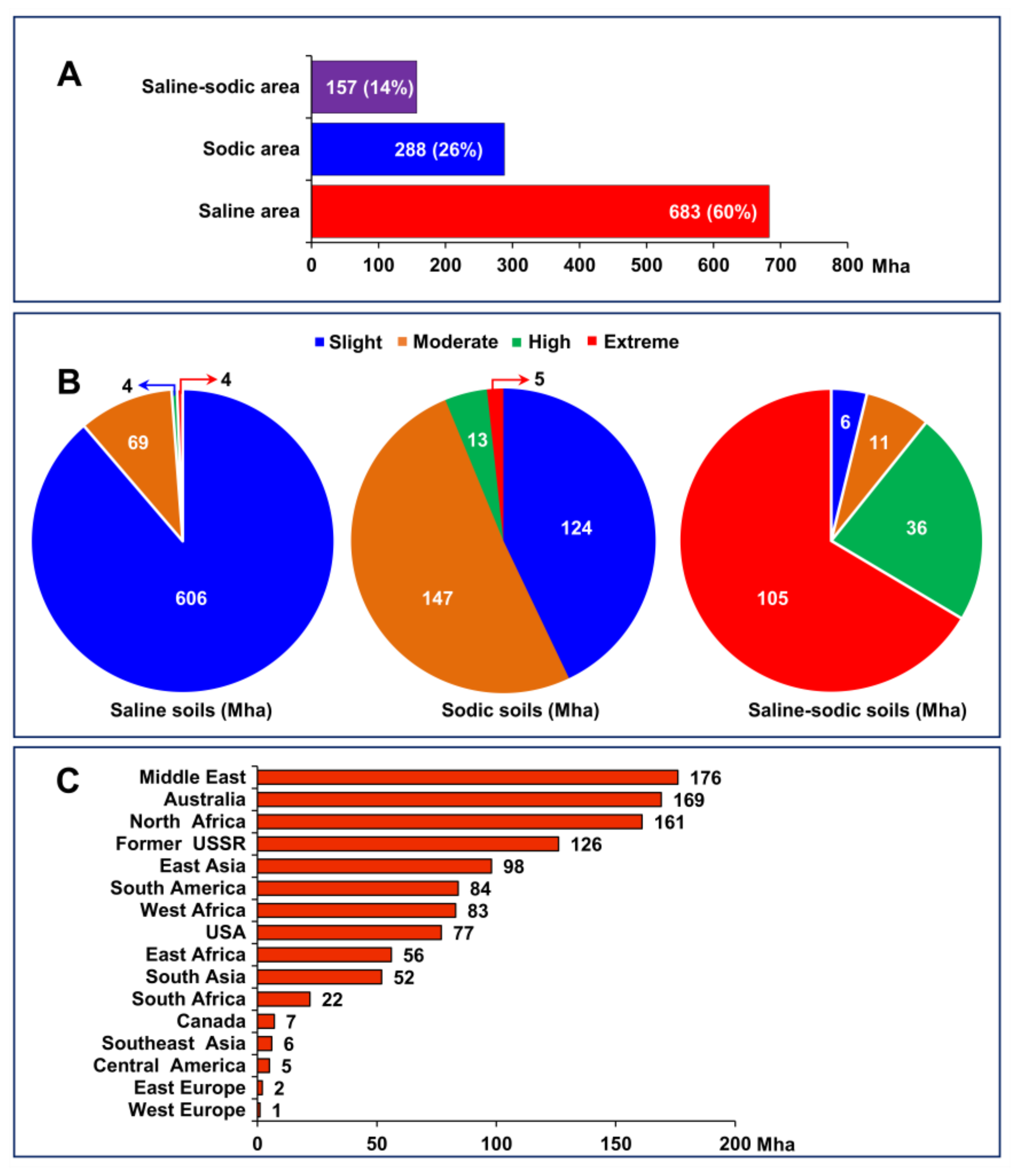
2. Adverse Effects of Salinity on Soil Properties
3. Mechanisms of Salinity-Mediated Adverse Effects on Plants
4. Halophytes and Their Classification
5. Physiological Mechanisms Associated with Halophyte Adaptation to Soil Salinity
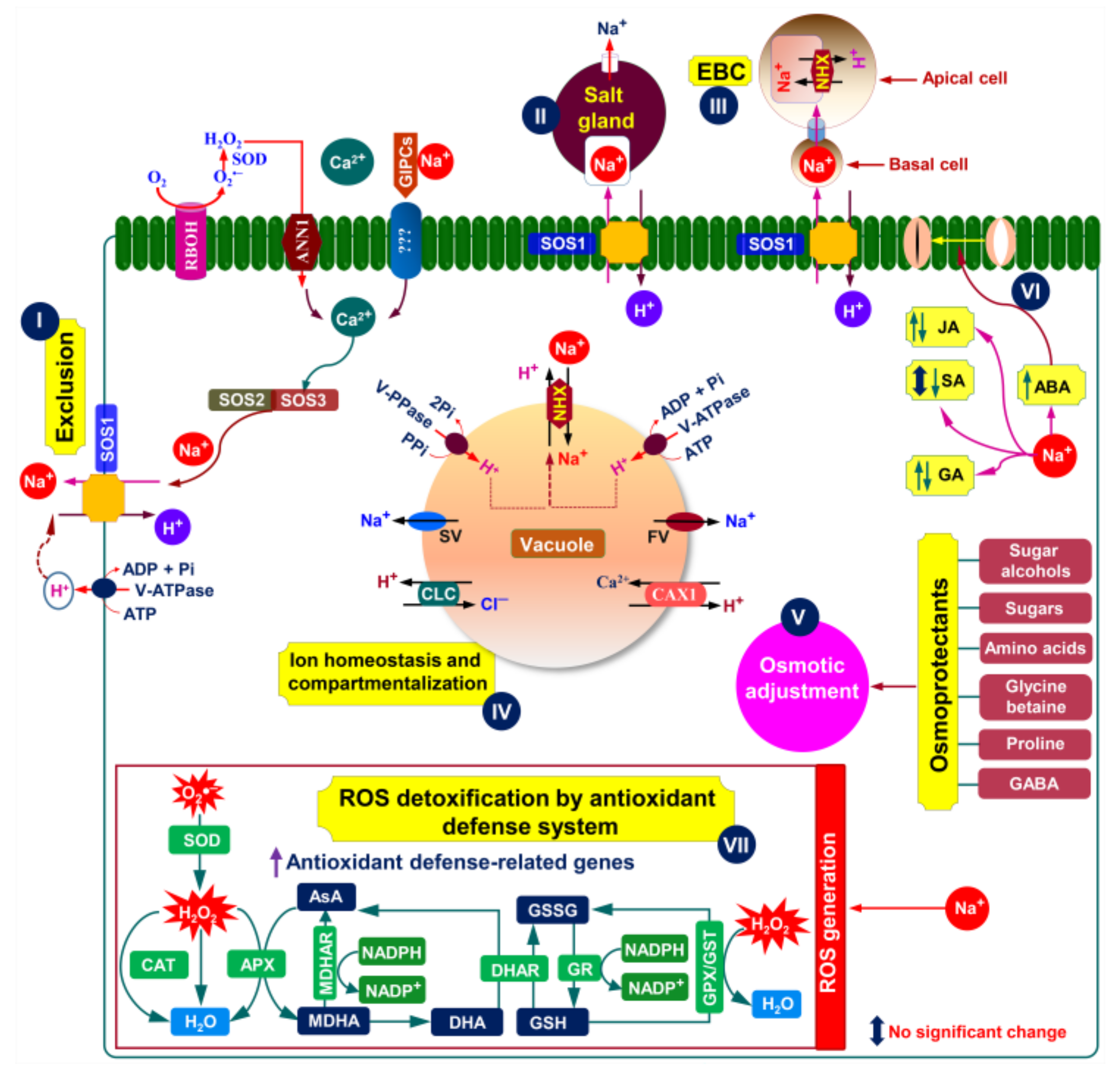
5.1. Salt Exclusion
5.2. Salt Excretion

5.3. Succulence

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl. No. | Species Name | Maximum Salt Tolerance Limit (mM) | Salt Tolerance Mechanisms | References |
|---|---|---|---|---|
| 1 | Haloxylon salicornicum | 400 |
| [88] |
| 2 | Scorzonera hieraciifolia | 600 |
| [89] |
| 3 | Salsola crassa | 300 |
| [90] |
| 4 | Aster tripolium | 600 |
| [91] |
| 5 | Lycium humile | 750 |
| [92] |
| 6 | Limonium bicolor, L. gmelinii, L. otolepis, L. aureum, L. sinuatum, and L. gmelinii | 100, 50, 300, 350, 400, and 420, respectively |
| [66,67,68,69] |
| Tamarix gallica, T. ramosissima, and T. laxa | 300 | |||
| 7 | Chenopodium quinoa and Atriplex centralasiatica | 300–400 |
| [73,78,80,81] |
| 8 | Nitraria sibirica | 200 |
| [93] |
| 9 | Mesembryanthemum crystallinum | 400 |
| [75] |
| 10 | Lobularia maritima | 400 |
| [94] |
| 11 | C. quinoa | 400 |
| [95] |
| 12 | Urochondra setulosa | 500 |
| [96] |
| 13 | Cakile maritima | 400 |
| [97] |
| 14 | Karelinia caspia | 200 |
| [63] |
| 15 | Suaeda salsa | 200 |
| [98] |
| 16 | Hordeum brevisubulatum | 750 |
| [99] |
| 17 | Zoysia macrostachya | 300 |
| [100] |
| 18 | Sarcocornia fruticosa | 60 |
| [85,86,87] |
| Achras sapota | 120 | |||
| S. drummondii | 500 | |||
| 19 | H. salicornicum | 400 |
| [101] |
| 20 | A. tripolium | 300 |
| [102] |
| 21 | Leptochloa fusca | 450 |
| [103] |
| 22 | C. maritima | 400 |
| [104] |
| 23 | Eutrema parvula | 300 |
| [105] |
| 24 | Salvadora persica | 750 |
| [106] |
| 25 | Pongamia pinnata | 500 |
| [107] |
| 26 | A. halimus | 171 |
| [108] |
| 27 | Sesuvium portulacastrum | 120 |
| [109] |
| 28 | Zygophyllum xanthoxylum | 50 |
| [110] |
| 29 | Puccinellia tenuiflora | 150 |
| [111] |
| 30 | Kosteletzkya virginica | 390 |
| [112,113] |
6. Biochemical Mechanisms Associated with Halophyte Adaptation to Salinity
6.1. Ion Homeostasis
6.2. Osmotic Adjustment
6.3. ROS-Detoxification
7. Halophyte-Mediated Phytoremediation of Salinity-Affected Soils

8. Potential of Halophytes as Genetic Resources for the Engineering of Crops with Improved Salt Tolerance
9. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, J.; Shi, H. Physiological and molecular mechanisms of plant salt tolerance. Photosynth. Res. 2013, 115, 1–22. [Google Scholar] [CrossRef]
- Qadir, M.; Quillérou, E.; Nangia, V.; Murtaza, G.; Singh, M.; Thomas, R.J.; Drechsel, P.; Noble, A.D. Economics of salt-induced land degradation and restoration. Nat. Resour. Forum 2014, 38, 282–295. [Google Scholar] [CrossRef]
- Calone, R.; Bregaglio, S.; Sanoubar, R.; Noli, E.; Lambertini, C.; Barbanti, L. Physiological adaptation to water salinity in six wild halophytes suitable for Mediterranean agriculture. Plants 2021, 10, 309. [Google Scholar] [CrossRef]
- Minhas, P.; Qadir, M.; Yadav, R. Groundwater irrigation induced soil sodification and response options. Agric. Water Manag. 2019, 215, 74–85. [Google Scholar] [CrossRef]
- Wang, Y.; Xiao, D.; Li, Y.; Li, X. Soil salinity evolution and its relationship with dynamics of groundwater in the oasis of inland river basins: Case study from the Fubei region of Xinjiang Province, China. Environ. Monit. Assess. 2007, 140, 291–302. [Google Scholar] [CrossRef]
- Omuto, C.T.V.; Vargas, R.R.; El Mobarak, A.M.; Mohamed, N.; Viatkin, K.; Yigini, Y. Mapping of Salt-Affected Soils: Technical Manual; FAO: Rome, Italy, 2020. [Google Scholar]
- Rahman, M.; Haque, A.; Nihad, S.A.I.; Howlader, R.A.; Akand, M.H. Morpho-physiological response of Acacia auriculiformis as influenced by seawater induced salinity stress. For. Syst. 2016, 25, e071. [Google Scholar] [CrossRef] [Green Version]
- Karakas, S.; Bolat, I.; Dikilitas, M. The use of halophytic companion plant (Portulaca oleracea L.) on some growth, fruit, and biochemical parameters of strawberry plants under salt stress. Horticulturae 2021, 7, 63. [Google Scholar] [CrossRef]
- Wicke, B.; Smeets, E.; Dornburg, V.; Vashev, B.; Gaiser, T.; Turkenburg, W.; Faaij, A. The global technical and economic potential of bioenergy from salt-affected soils. Energy Environ. Sci. 2011, 4, 2669–2681. [Google Scholar]
- Rahman, M.; Mostofa, M.G.; Rahman, A.; Islam, R.; Keya, S.S.; Das, A.K.; Miah, G.; Kawser, A.Q.M.R.; Ahsan, S.M.; Hashem, A.; et al. Acetic acid: A cost-effective agent for mitigation of seawater-induced salt toxicity in mung bean. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.-K.; Shabala, S. Mechanisms of plant responses and adaptation to soil salinity. Innovation 2020, 1. [Google Scholar] [CrossRef]
- Zaid, A.; Wani, S.H. Reactive oxygen species generation, scavenging and signaling in plant defense responses. In Bioactive Molecules in Plant Defense; Springer Science and Business Media LLC: Berlin, Germany, 2019; pp. 111–132. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Rahman, M.; Ansary, M.U.; Keya, S.S.; Abdelrahman, M.; Miah, G.; Tran, L.-S.P. Silicon in mitigation of abiotic stress-induced oxidative damage in plants. Crit. Rev. Biotechnol. 2021, 41, 918–934. [Google Scholar] [CrossRef]
- Patel, M.K.; Kumar, M.; Li, W.; Luo, Y.; Burritt, D.J.; Alkan, N.; Tran, L.-S.P. Enhancing salt tolerance of plants: From metabolic reprogramming to exogenous chemical treatments and molecular approaches. Cells 2020, 9, 2492. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Litalien, A.; Zeeb, B. Curing the earth: A review of anthropogenic soil salinization and plant-based strategies for sustainable mitigation. Sci. Total Environ. 2020, 698, 134235. [Google Scholar] [CrossRef]
- Liu, A.; Xiao, Z.; Li, M.-W.; Wong, F.-L.; Yung, W.-S.; Ku, Y.-S.; Wang, Q.; Wang, X.; Xie, M.; Yim, A.K.-Y.; et al. Transcriptomic reprogramming in soybean seedlings under salt stress. Plant Cell Environ. 2018, 42, 98–114. [Google Scholar] [CrossRef] [Green Version]
- Rengasamy, P. Soil chemistry factors confounding crop salinity tolerance—A review. Agronomy 2016, 6, 53. [Google Scholar] [CrossRef] [Green Version]
- Yuan, C.; Feng, S.; Wang, J.; Huo, Z.; Ji, Q. Effects of irrigation water salinity on soil salt content distribution, soil physical properties and water use efficiency of maize for seed production in arid Northwest China. Int. J. Agric. Biol. Eng. 2018, 11, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.; Tian, L.; Chang, C.; Wang, S.; Zhang, J.; Zhou, X.; Li, X.; Tran, L.-S.P.; Tian, C. Grass and maize vegetation systems restore saline-sodic soils in the Songnen Plain of northeast China. Land Degrad. Dev. 2018, 29, 1107–1119. [Google Scholar] [CrossRef]
- Guangming, L.; Xuechen, Z.; Xiuping, W.; Hongbo, S.; Jingsong, Y.; Xiangping, W. Soil enzymes as indicators of saline soil fertility under various soil amendments. Agric. Ecosyst. Environ. 2017, 237, 274–279. [Google Scholar] [CrossRef]
- Zhang, W.-w.; Chong, W.; Rui, X.; Wang, L.-j. Effects of salinity on the soil microbial community and soil fertility. J. Integr. Agric. 2019, 18, 1360–1368. [Google Scholar]
- Tian, L.; Lin, X.; Tian, J.; Ji, L.; Chen, Y.; Tran, L.-S.P.; Tian, C. Research advances of beneficial microbiota associated with crop plants. Int. J. Mol. Sci. 2020, 21, 1792. [Google Scholar] [CrossRef] [Green Version]
- Yan, N.; Marschner, P.; Cao, W.; Zuo, C.; Qin, W. Influence of salinity and water content on soil microorganisms. Int. Soil Water Conserv. Res. 2015, 3, 316–323. [Google Scholar] [CrossRef] [Green Version]
- Rath, K.M.; Rousk, J. Salt effects on the soil microbial decomposer community and their role in organic carbon cycling: A review. Soil Biol. Biochem. 2015, 81, 108–123. [Google Scholar] [CrossRef]
- Xie, X.; Pu, L.; Wang, Q.; Zhu, M.; Xu, Y.; Zhang, M. Response of soil physicochemical properties and enzyme activities to long-term reclamation of coastal saline soil, Eastern China. Sci. Total Environ. 2017, 607–608, 1419–1427. [Google Scholar] [CrossRef]
- Uçarlı, C. Effects of salinity on seed germination and early seedling stage. In Abiotic Stress in Plants; IntechOpen: London, UK, 2020. [Google Scholar]
- El-Keblawy, A.; Aljasmi, M.; Gairola, S.; Mosa, K.A.; Hameed, A. Provenance determines salinity tolerance and germination requirements of the multipurpose tree Prosopis juliflora seeds. Arid Land Res. Manag. 2021, 1–16. [Google Scholar] [CrossRef]
- Miransari, M.; Smith, D. Plant hormones and seed germination. Environ. Exp. Bot. 2014, 99, 110–121. [Google Scholar] [CrossRef]
- Alencar, N.L.M.; Gadelha, C.G.; Gallão, M.I.; Dolder, M.A.H.; Prisco, J.T.; Gomes-Filho, E. Ultrastructural and biochemical changes induced by salt stress in Jatropha curcas seeds during germination and seedling development. Funct. Plant Biol. 2015, 42, 865–874. [Google Scholar] [CrossRef]
- Mwando, E.; Han, Y.; Angessa, T.T.; Zhou, G.; Hill, C.B.; Zhang, X.-Q.; Li, C. Genome- wide association study of salinity tolerance during germination in barley (Hordeum vulgare L.). Front. Plant Sci. 2020, 11, 118. [Google Scholar] [CrossRef] [Green Version]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef]
- Basu, S.; Kumar, A.; Benazir, I.; Kumar, G. Reassessing the role of ion homeostasis for improving salinity tolerance in crop plants. Physiol. Plant. 2020, 171, 502–519. [Google Scholar] [CrossRef]
- Geilfus, C.-M. Chloride in soil: From nutrient to soil pollutant. Environ. Exp. Bot. 2018, 157, 299–309. [Google Scholar] [CrossRef]
- Gururani, M.; Venkatesh, J.; Tran, L.S.P. Regulation of photosynthesis during abiotic stress-induced photoinhibition. Mol. Plant 2015, 8, 1304–1320. [Google Scholar] [CrossRef] [Green Version]
- Miteva, T.; Zhelev, N.Z.; Popova, L. Effect of salinity on the synthesis of ribulose-1,5-bisphosphate carboxylase/oxygenase in barley leaves. J. Plant Physiol. 1992, 140, 46–51. [Google Scholar] [CrossRef]
- Czarnocka, W.; Karpiński, S. Friend or foe? Reactive oxygen species production, scavenging and signaling in plant response to environmental stresses. Free Radic. Biol. Med. 2018, 122, 4–20. [Google Scholar] [CrossRef]
- Geilfus, C.-M. Chloride: From nutrient to toxicant. Plant Cell Physiol. 2018, 59, 877–886. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Plant salt tolerance: Adaptations in halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef] [Green Version]
- Parida, A.K.; Jha, B. Salt tolerance mechanisms in mangroves: A review. Trees 2010, 24, 199–217. [Google Scholar] [CrossRef]
- Von Sengbusch, P. Halophytes Botanik Online; University of Hamburg: Hamburg, Germany, 2003. [Google Scholar]
- Grigore, M.N.; Toma, C.; Boscaiu, M. Dealing with halophytes: An old problem, the same continuous exciting challenge. Analele Stiintifice ale Universitatii” Alexandru Ioan Cuza” din Iasi. Biol. Veg. 2010, 56, 21–32. [Google Scholar]
- Grigore, M.-N.; Toma, C. A proposal for a new halophytes classification, based on integrative anatomy observations. Muz. Olten. Craiova Stud. Com. Şt. Nat. 2010, 26, 45–50. [Google Scholar]
- Breckle, S.-W. Salinity, halophytes and salt affected natural ecosystems. In Salinity: Environment-Plants-Molecules; Springer: Dordrecht, The Netherlands, 2002; pp. 53–77. [Google Scholar]
- Santos, J.; Al-Azzawi, M.; Aronson, J.; Flowers, T.J. eHALOPH a database of salt-tolerant plants: Helping put halophytes to work. Plant Cell Physiol. 2015, 57, e10. [Google Scholar] [CrossRef] [Green Version]
- Breckle, S. How do halophytes overcome salinity. Biol. Salt Toler. Plants 1995, 23, 199–203. [Google Scholar]
- Chen, M.; Yang, Z.; Liu, J.; Zhu, T.; Wei, X.; Fan, H.; Wang, B. Adaptation mechanism of salt excluders under saline conditions and its applications. Int. J. Mol. Sci. 2018, 19, 3668. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.; Yuan, F.; Guo, J.; Han, G.; Wang, C.; Chen, M.; Wang, B. Current understanding of role of vesicular transport in salt secretion by salt glands in recretohalophytes. Int. J. Mol. Sci. 2021, 22, 2203. [Google Scholar] [CrossRef]
- Hussain, S.; Khalid, M.F.; Sohail, M.; Anjum, M.A.; Ejaz, S.; Nafees, M.; Zakir, I.; Ahmad, M.; Ali, S.; Ahmad, S. Role of transporters in accumulating salt ions by halophytes. Approaches Remediat. Inorg. Pollut. 2021, 11–40. [Google Scholar] [CrossRef]
- Lee, Y.; Rubio, M.C.; Alassimone, J.; Geldner, N. A mechanism for localized lignin deposition in the endodermis. Cell 2013, 153, 402–412. [Google Scholar] [CrossRef] [Green Version]
- Naseer, S.; Lee, Y.; Lapierre, C.; Franke, R.; Nawrath, C.; Geldner, N. Casparian strip diffusion barrier in Arabidopsis is made of a lignin polymer without suberin. Proc. Natl. Acad. Sci. USA 2012, 109, 10101–10106. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, P.; Ranathunge, K.; Nayak, S.; Schreiber, L.; Mathew, M.K. Root apoplastic barriers block Na+ transport to shoots in rice (Oryza sativa L.). J. Exp. Bot. 2011, 62, 4215–4228. [Google Scholar] [CrossRef]
- Zhang, S.; Quartararo, A.; Betz, O.K.; Madahhosseini, S.; Heringer, A.S.; Le, T.; Shao, Y.; Caruso, T.; Ferguson, L.; Jernstedt, J. Root vacuolar sequestration and suberization are prominent responses of Pistacia spp. rootstocks during salinity stress. Plant Direct 2021, 5, e00315. [Google Scholar]
- Cui, B.; Liu, R.; Flowers, T.J.; Song, J. Casparian bands and Suberin lamellae: Key targets for breeding salt tolerant crops? Environ. Exp. Bot. 2021, 191, 104600. [Google Scholar]
- Wang, P.; Wang, C.-M.; Gao, L.; Cui, Y.-N.; Yang, H.-L.; De Silva, N.D.G.; Ma, Q.; Bao, A.-K.; Flowers, T.J.; Rowland, O.; et al. Aliphatic suberin confers salt tolerance to Arabidopsis by limiting Na+ influx, K+ efflux and water backflow. Plant Soil 2020, 448, 603–620. [Google Scholar] [CrossRef]
- Hajibagheri, M.A.; Yeo, A.R.; Flowers, T.J. Salt tolerance in Suaeda Maritima (L.) Dum. fine structure and ion concentrations in the apical region of roots. New Phytol. 1985, 99, 331–343. [Google Scholar] [CrossRef]
- Krishnamurthy, P.; Jyothi-Prakash, P.A.; Qin, L.; He, J.; Lin, Q.; Loh, C.-S.; Kumar, P.P. Role of root hydrophobic barriers in salt exclusion of a mangrove plant Avicennia officinalis. Plant Cell Environ. 2014, 37, 1656–1671. [Google Scholar] [CrossRef]
- Liu, Q.; Yang, Y.; Zhang, X.; Liu, R.; Song, J. Chloride allocation in the euhalophyte Suaeda salsa from different habitats in field and controlled saline conditions. Aquat. Bot. 2020, 167, 103292. [Google Scholar] [CrossRef]
- Cheng, H.; Inyang, A.; Li, C.-D.; Fei, J.; Zhou, Y.-W.; Wang, Y.-S. Salt tolerance and exclusion in the mangrove plant Avicennia marina in relation to root apoplastic barriers. Ecotoxicology 2020, 29, 676–683. [Google Scholar] [CrossRef]
- Liu, Q.; Liu, R.; Ma, Y.; Song, J. Physiological and molecular evidence for Na+ and Cl− exclusion in the roots of two Suaeda salsa populations. Aquat. Bot. 2018, 146, 1–7. [Google Scholar] [CrossRef]
- Guo, Q.; Meng, L.; Han, J.; Mao, P.; Tian, X.; Zheng, M.; Mur, L.A. SOS1 is a key systemic regulator of salt secretion and K+/Na+ homeostasis in the recretohalophyte Karelinia caspia. Environ. Exp. Bot. 2020, 177, 104098. [Google Scholar] [CrossRef]
- Roy, S.J.; Negrão, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef]
- Shabala, S.; Bose, J.; Hedrich, R. Salt bladders: Do they matter? Trends Plant Sci. 2014, 19, 687–691. [Google Scholar]
- Leng, B.; Yuan, F.; Dong, X.; Wang, J.; Wang, B. Distribution pattern and salt excretion rate of salt glands in two recretohalophyte species of Limonium (Plumbaginaceae). S. Afr. J. Bot. 2018, 115, 74–80. [Google Scholar] [CrossRef]
- Li, J.; Yuan, F.; Liu, Y.; Zhang, M.; Liu, Y.; Zhao, Y.; Wang, B.; Chen, M. Exogenous melatonin enhances salt secretion from salt glands by upregulating the expression of ion transporter and vesicle transport genes in Limonium bicolor. BMC Plant Biol. 2020, 20, 1–11. [Google Scholar] [CrossRef]
- Mi, P.; Yuan, F.; Guo, J.; Han, G.; Wang, B. Salt glands play a pivotal role in the salt resistance of four recretohalophyte Limonium Mill. species. Plant Biol. 2021. [Google Scholar] [CrossRef]
- Wei, X.; Yan, X.; Yang, Z.; Han, G.; Wang, L.; Yuan, F.; Wang, B. Salt glands of recretohalophyte Tamarix under salinity: Their evolution and adaptation. Ecol. Evol. 2020, 10, 9384–9395. [Google Scholar] [CrossRef]
- Yuan, F.; Leng, B.; Wang, B. Progress in studying salt secretion from the salt glands in recretohalophytes: How do plants secrete salt? Front. Plant Sci. 2016, 7, 977. [Google Scholar]
- Céccoli, G.; Ramos, J.; Pilatti, V.; Dellaferrera, I.; Tivano, J.C.; Taleisnik, E.; Vegetti, A.C. Salt glands in the Poaceae family and their relationship to salinity tolerance. Bot. Rev. 2015, 81, 162–178. [Google Scholar] [CrossRef] [Green Version]
- Arbelet-Bonnin, D.; Ben-Hamed-Louati, I.; Laurenti, P.; Abdelly, C.; Ben-Hamed, K.; Bouteau, F. Cakile maritima, a promising model for halophyte studies and a putative cash crop for saline agriculture. Adv. Agron. 2019, 155, 45–78. [Google Scholar] [CrossRef]
- Kiani-Pouya, A.; Roessner, U.; Jayasinghe, N.S.; Lutz, A.; Rupasinghe, T.; Bazihizina, N.; Bohm, J.; Alharbi, S.; Hedrich, R.; Shabala, S. Epidermal bladder cells confer salinity stress tolerance in the halophyte quinoa and Atriplex species. Plant Cell Environ. 2017, 40, 1900–1915. [Google Scholar] [CrossRef] [Green Version]
- Barkla, B.J.; Rhodes, T.; Tran, K.-N.T.; Wijesinghege, C.; Larkin, J.C.; Dassanayake, M. Making epidermal bladder cells bigger: Developmental- and salinity-induced endopolyploidy in a model halophyte. Plant Physiol. 2018, 177, 615–632. [Google Scholar] [CrossRef]
- Tran, D.Q.; Konishi, A.; Cushman, J.C.; Morokuma, M.; Toyota, M.; Agarie, S. Ion accumulation and expression of ion homeostasis-related genes associated with halophilism, NaCl-promoted growth in a halophyte Mesembryanthemum crystallinum L. Plant Prod. Sci. 2019, 23, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Himabindu, Y.; Chakradhar, T.; Reddy, M.C.; Kanygin, A.; Redding, K.E.; Chandrasekhar, T. Salt-tolerant genes from halophytes are potential key players of salt tolerance in glycophytes. Environ. Exp. Bot. 2016, 124, 39–63. [Google Scholar] [CrossRef] [Green Version]
- Bonales-Alatorre, E.; Shabala, S.; Chen, Z.-H.; Pottosin, I. Reduced tonoplast fast-activating and slow-activating channel activity is essential for conferring salinity tolerance in a facultative halophyte, Quinoa. Plant Physiol. 2013, 162, 940–952. [Google Scholar] [CrossRef] [Green Version]
- Otterbach, S.L.; Khoury, H.; Rupasinghe, T.; Mendis, H.; Kwan, K.H.; Lui, V.; Natera, S.H.A.; Klaiber, I.; Allen, N.M.; Jarvis, D.E.; et al. Characterization of epidermal bladder cells in Chenopodium quinoa. Plant Cell Environ. 2021. [Google Scholar] [CrossRef]
- Roeurn, S.; Hoshino, N.; Soejima, K.-T.; Inoue, Y.; Cushman, J.C.; Agarie, S. MYB and HD-ZIP IV homologs related to trichome formation are involved in epidermal bladder cell development in the halophyte Mesembryanthemum crystallinum L. Plant Prod. Sci. 2017, 20, 72–82. [Google Scholar] [CrossRef]
- Böhm, J.; Messerer, M.; Müller, H.M.; Scholz-Starke, J.; Gradogna, A.; Scherzer, S.; Maierhofer, T.; Bazihizina, N.; Zhang, H.; Stigloher, C.; et al. Understanding the molecular basis of salt sequestration in epidermal bladder cells of Chenopodium quinoa. Curr. Biol. 2018, 28, 3075–3085.e7. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Zhang, X.; Wang, N.; Cui, Y.; Zhang, L.; Fan, S. Transcriptome analysis of salt stress response in halophyte Atriplex centralasiatica leaves. Acta Physiol. Plant. 2019, 42, 3. [Google Scholar] [CrossRef]
- Imamura, T.; Yasui, Y.; Koga, H.; Takagi, H.; Abe, A.; Nishizawa, K.; Mizuno, N.; Ohki, S.; Mizukoshi, H.; Mori, M. A novel WD40-repeat protein involved in formation of epidermal bladder cells in the halophyte quinoa. Commun. Biol. 2020, 3, 1–14. [Google Scholar] [CrossRef]
- Lokhande, V.H.; Suprasanna, P. Prospects of halophytes in understanding and managing abiotic stress tolerance. In Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change; Springer: Berlin, Germany, 2012; pp. 29–56. [Google Scholar]
- Nikalje, G.C.; Srivastava, A.K.; Pandey, G.K.; Suprasanna, P. Halophytes in biosaline agriculture: Mechanism, utilization, and value addition. Land Degrad. Dev. 2017, 29, 1081–1095. [Google Scholar] [CrossRef]
- García-Caparrós, P.; Llanderal, A.; Pestana, M.; Correia, P.J.; Lao, M.T. Nutritional and physiological responses of the dicotyledonous halophyte Sarcocornia fruticosa to salinity. Aust. J. Bot. 2017, 65, 573–581. [Google Scholar] [CrossRef]
- Elnaggar, A.; Mosa, K.A.; El-Keblawy, A.; Tammam, A.; El-Naggar, M. Physiological and biochemical insights for salt stress tolerance in the habitat-indifferent halophyte Salsola drummondii during the vegetative stage. Botany 2020, 98, 1–17. [Google Scholar] [CrossRef]
- Rahman, M.; Mostofa, M.G.; Rahman, A.; Miah, G.; Saha, S.R.; Karim, M.A.; Keya, S.S.; Akter, M.; Islam, M.; Tran, L.-S.P. Insight into salt tolerance mechanisms of the halophyte Achras sapota: An important fruit tree for agriculture in coastal areas. Protoplasma 2018, 256, 181–191. [Google Scholar] [CrossRef]
- Panda, A.; Rangani, J.; Parida, A.K. Unraveling salt responsive metabolites and metabolic pathways using non-targeted metabolomics approach and elucidation of salt tolerance mechanisms in the xero-halophyte Haloxylon salicornicum. Plant Physiol. Biochem. 2020, 158, 284–296. [Google Scholar] [CrossRef]
- Altuntaş, C.; Terzi, R. Concomitant accumulations of ions, osmoprotectants and antioxidant system-related substances provide salt tolerance capability to succulent extreme-halophyte Scorzonera hieraciifolia. Authorea Prepr. 2021. [Google Scholar] [CrossRef]
- Yıldız, M.; Terzi, H. Comparative analysis of salt-induced changes in the root physiology and proteome of the xero-halophyte Salsola crassa. Braz. J. Bot. 2021, 44, 33–42. [Google Scholar] [CrossRef]
- Wiszniewska, A.; Koźmińska, A.; Hanus-Fajerska, E.; Dziurka, K.; Dziurka, M. Insight into phytohormonal modulation of defense mechanisms to salt excess in a halophyte and a glycophyte from Asteraceae family. Plant Soil 2021, 463, 55–76. [Google Scholar] [CrossRef]
- Palchetti, M.V.; Reginato, M.; Llanes, A.; Hornbacher, J.; Papenbrock, J.; Barboza, G.E.; Luna, V.; Cantero, J.J. New insights into the salt tolerance of the extreme halophytic species Lycium humile (Lycieae, Solanaceae). Plant Physiol. Biochem. 2021, 163, 166–177. [Google Scholar] [CrossRef]
- Tang, X.; Zhang, H.; Shabala, S.; Li, H.; Yang, X.; Zhang, H. Tissue tolerance mechanisms conferring salinity tolerance in a halophytic perennial species Nitraria sibirica Pall. Tree Physiol. 2020. [Google Scholar] [CrossRef]
- Ben Hsouna, A.; Ghneim-Herrera, T.; Ben Romdhane, W.; Dabbous, A.; Ben Saad, R.; Brini, F.; Abdelly, C.; Ben Hamed, K. Early effects of salt stress on the physiological and oxidative status of the halophyte Lobularia maritima. Funct. Plant Biol. 2020, 47, 912–924. [Google Scholar] [CrossRef]
- Kiani-Pouya, A.; Rasouli, F.; Shabala, L.; Tahir, A.T.; Zhou, M.; Shabala, S. Understanding the role of root-related traits in salinity tolerance of quinoa accessions with contrasting epidermal bladder cell patterning. Planta 2020, 251, 1–11. [Google Scholar] [CrossRef]
- Mann, A.; Kumar, N.; Kumar, A.; Lata, C.; Kumar, A.; Meena, B.L.; Mishra, D.; Grover, M.; Gaba, S.; Parameswaran, C.; et al. De novo transcriptomic profiling of differentially expressed genes in grass halophyte Urochondra setulosa under high salinity. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Arbelet-Bonnin, D.; Blasselle, C.; Palm, E.R.; Redwan, M.; Ponnaiah, M.; Laurenti, P.; Meimoun, P.; Gilard, F.; Gakière, B.; Mancuso, S.; et al. Metabolism regulation during salt exposure in the halophyte Cakile maritima. Environ. Exp. Bot. 2020, 177, 104075. [Google Scholar] [CrossRef]
- Guo, J.; Lu, C.; Zhao, F.; Gao, S.; Wang, B. Improved reproductive growth of euhalophyte Suaeda salsa under salinity is correlated with altered phytohormone biosynthesis and signal transduction. Funct. Plant Biol. 2020, 47, 170–183. [Google Scholar] [CrossRef]
- Zhang, H.; Xiao, W.; Yu, W.; Jiang, Y.; Li, R. Halophytic Hordeum brevisubulatum HbHAK1 facilitates potassium retention and contributes to salt tolerance. Int. J. Mol. Sci. 2020, 21, 5292. [Google Scholar] [CrossRef]
- Wang, R.; Wang, X.; Liu, K.; Zhang, X.-J.; Zhang, L.-Y.; Fan, S.-J. Transcriptome analysis of halophyte Zoysia macrostachya in response to salinity stress. Plants 2020, 9, 458. [Google Scholar] [CrossRef] [Green Version]
- Panda, A.; Rangani, J.; Parida, A.K. Cross talk between ROS homeostasis and antioxidative machinery contributes to salt tolerance of the xero-halophyte Haloxylon salicornicum. Environ. Exp. Bot. 2019, 166, 103799. [Google Scholar] [CrossRef]
- Wiszniewska, A.; Koźmińska, A.; Hanus-Fajerska, E.; Dziurka, M.; Dziurka, K. Insight into mechanisms of multiple stresses tolerance in a halophyte Aster tripolium subjected to salinity and heavy metal stress. Ecotoxicol. Environ. Saf. 2019, 180, 12–22. [Google Scholar] [CrossRef]
- Mohammadi, F.; Kavousi, H.R.; Mansouri, M. Effects of salt stress on physio-biochemical characters and gene expressions in halophyte grass Leptochloa fusca (L.) Kunth. Acta Physiol. Plant. 2019, 41, 1–10. [Google Scholar] [CrossRef]
- Arbelet-Bonnin, D.; Ben Hamed-Laouti, I.; Laurenti, P.; Abdelly, C.; Ben Hamed, K.; Bouteau, F. Cellular mechanisms to survive salt in the halophyte Cakile maritima. Plant Sci. 2018, 272, 173–178. [Google Scholar] [CrossRef]
- Ali, A.; Khan, I.U.; Jan, M.; Khan, H.A.; Hussain, S.; Nisar, M.; Chung, W.S.; Yun, D.-J. The High-affinity potassium transporter EpHKT1;2 from the extremophile Eutrema parvula mediates salt tolerance. Front. Plant Sci. 2018, 9, 1108. [Google Scholar] [CrossRef] [Green Version]
- Kumari, A.; Parida, A. Metabolomics and network analysis reveal the potential metabolites and biological pathways involved in salinity tolerance of the halophyte Salvadora persica. Environ. Exp. Bot. 2018, 148, 85–99. [Google Scholar] [CrossRef]
- Marriboina, S.; Sengupta, D.; Kumar, S.; Reddy, A.R. Physiological and molecular insights into the high salinity tolerance of Pongamia pinnata (L.) pierre, a potential biofuel tree species. Plant Sci. 2017, 258, 102–111. [Google Scholar] [CrossRef]
- Hamdani, F.; Derridj, A.; Rogers, H.J. Multiple mechanisms mediate growth and survival in young seedlings of two populations of the halophyte Atriplex halimus (L.) subjected to long single-step salinity treatments. Funct. Plant Biol. 2017, 44, 761–773. [Google Scholar] [CrossRef]
- Muchate, N.S.; Nikalje, G.; Rajurkar, N.S.; Suprasanna, P.; Nikam, T.D. Physiological responses of the halophyte Sesuvium portulacastrum to salt stress and their relevance for saline soil bio-reclamation. Flora 2016, 224, 96–105. [Google Scholar] [CrossRef]
- Ma, Q.; Hu, J.; Zhou, X.-R.; Yuan, H.-J.; Kumar, T.; Luan, S.; Wang, S.-M. ZxAKT1 is essential for K+ uptake and K+/Na+ homeostasis in the succulent xerophyte Zygophyllum xanthoxylum. Plant J. 2017, 90, 48–60. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.-D.; Wang, P.; Bao, Z.; Ma, Q.; Duan, L.-J.; Bao, A.-K.; Zhang, J.-L.; Wang, S.-M. SOS1, HKT1;5, and NHX1 synergistically modulate Na+ homeostasis in the halophytic grass Puccinellia tenuiflora. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.; Xia, Y.; Ma, B.L.; Feng, C.; Qin, P. Culture of seashore mallow under different salinity levels using plastic nutrient-rich matrices and transplantation. Agron. J. 2010, 102, 395–402. [Google Scholar] [CrossRef]
- Wang, H.; Tang, X.; Wang, H.; Shao, H.-B. Proline accumulation and metabolism-related genes expression profiles in Kosteletzkya virginica seedlings under salt stress. Front. Plant Sci. 2015, 6, 792. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Li, Z. The importance of Cl− exclusion and vacuolar Cl− sequestration: Revisiting the role of Cl− transport in plant salt tolerance. Front. Plant Sci. 2019, 10, 1418. [Google Scholar] [CrossRef]
- Li, B.; Tester, M.; Gilliham, M. Chloride on the move. Trends Plant Sci. 2017, 22, 236–248. [Google Scholar] [CrossRef]
- Saddhe, A.A.; Jamdade, R.; Gairola, S. Recent advances on cellular signaling paradigm and salt stress responsive genes in halophytes. Handb. Halophytes Mol. Ecosyst. Towards Biosaline Agric. 2020, 1–26. [Google Scholar] [CrossRef]
- Munns, R.; Passioura, J.B.; Colmer, T.D.; Byrt, C.S. Osmotic adjustment and energy limitations to plant growth in saline soil. New Phytol. 2019, 225, 1091–1096. [Google Scholar] [CrossRef] [Green Version]
- Slama, I.; Abdelly, C.; Bouchereau, A.; Flowers, T.; Savouré, A. Diversity, distribution and roles of osmoprotective compounds accumulated in halophytes under abiotic stress. Ann. Bot. 2015, 115, 433–447. [Google Scholar] [CrossRef] [Green Version]
- Castro, B.; Citterico, M.; Kimura, S.; Stevens, D.M.; Wrzaczek, M.; Coaker, G. Stress-induced reactive oxygen species compartmentalization, perception and signalling. Nat. Plants 2021, 7, 403–412. [Google Scholar] [CrossRef]
- Bose, J.; Rodrigo-Moreno, A.; Shabala, S. ROS homeostasis in halophytes in the context of salinity stress tolerance. J. Exp. Bot. 2013, 65, 1241–1257. [Google Scholar] [CrossRef]
- Llanes, A.; Reginato, M.; Devinar, G.; Luna, V. What is known about phytohormones in halophytes? A review. Biologia 2018, 73, 727–742. [Google Scholar] [CrossRef]
- Gupta, A.; Bhardwaj, M.; Tran, L.-S.P. Jasmonic acid at the crossroads of plant immunity and Pseudomonas syringae virulence. Int. J. Mol. Sci. 2020, 21, 7482. [Google Scholar] [CrossRef]
- Reinbothe, C.; Springer, A.; Samól, I.; Reinbothe, S. Plant oxylipins: Role of jasmonic acid during programmed cell death, defence and leaf senescence. FEBS J. 2009, 276, 4666–4681. [Google Scholar] [CrossRef]
- Xia, X.-J.; Zhou, Y.-H.; Shi, K.; Zhou, J.; Foyer, C.H.; Yu, J.-Q. Interplay between reactive oxygen species and hormones in the control of plant development and stress tolerance. J. Exp. Bot. 2015, 66, 2839–2856. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, N. Reclamation of saline and sodic soil through phytoremediation. In Environmental Concerns and Sustainable Development: Volume 2: Biodiversity, Soil and Waste Management; Shukla, V., Kumar, N., Eds.; Springer: Singapore, 2020; pp. 279–306. [Google Scholar]
- Jesus, J.; Danko, A.S.; Fiuza, A.; Borges, M.T. Phytoremediation of salt-affected soils: A review of processes, applicability, and the impact of climate change. Environ. Sci. Pollut. Res. 2015, 22, 6511–6525. [Google Scholar] [CrossRef]
- Qadir, M.; Tubeileh, A.; Akhtar, J.; Larbi, A.; Minhas, P.S.; Khan, M.A. Productivity enhancement of salt-affected environments through crop diversification. Land Degrad. Dev. 2008, 19, 429–453. [Google Scholar] [CrossRef]
- Yazdanpanah, N.; Mahmoodabadi, M. Effect of some amendments on leachate properties of a calcareous saline- sodic soil: A laboratory experiment. Int. Agrophys. 2010, 25, 14738. [Google Scholar]
- Leogrande, R.; Vitti, C. Use of organic amendments to reclaim saline and sodic soils: A review. Arid Land Res. Manag. 2018, 33, 1–21. [Google Scholar] [CrossRef]
- Makni, H.; Ayed, L.; Ben Khedher, M.; Bakhrouf, A. Evaluation of the maturity of organic waste composts. Waste Manag. Res. 2009, 28, 489–495. [Google Scholar] [CrossRef]
- Lakhdar, A.; Scelza, R.; Scotti, R.; A Rao, M.; Jedidi, N.; Gianfreda, L.; Abdelly, C. The effect of compost and sewage sludge on soil biologic activities in salt affected soil. Rev. Cienc. Suelo Nutr. Veg. 2010, 10, 40–47. [Google Scholar] [CrossRef] [Green Version]
- Ouni, Y.; Lakhdar, A.; Scelza, R.; Scotti, R.; Abdelly, C.; Barhoumi, Z.; Rao, M.A. Effects of two composts and two grasses on microbial biomass and biological activity in a salt-affected soil. Ecol. Eng. 2013, 60, 363–369. [Google Scholar] [CrossRef]
- Wang, L.; Wang, X.; Jiang, L.; Zhang, K.; Tanveer, M.; Tian, C.; Zhao, Z. Reclamation of saline soil by planting annual euhalophyte Suaeda salsa with drip irrigation: A three-year field experiment in arid northwestern China. Ecol. Eng. 2020, 159, 106090. [Google Scholar] [CrossRef]
- Liang, J.; Shi, W. Cotton/halophytes intercropping decreases salt accumulation and improves soil physicochemical properties and crop productivity in saline-alkali soils under mulched drip irrigation: A three-year field experiment. Field Crop. Res. 2020, 262, 108027. [Google Scholar] [CrossRef]
- Shaygan, M.; Mulligan, D.; Baumgartl, T. The potential of three halophytes (Tecticornia pergranulata, Sclerolaena longicuspis, and Frankenia serpyllifolia) for the rehabilitation of brine-affected soils. Land Degrad. Dev. 2018, 29, 2002–2014. [Google Scholar] [CrossRef]
- Mann, E.; Rutter, A.; Zeeb, B. Evaluating the efficacy of Atriplex spp. in the phytoextraction of road salt (NaCl) from contaminated soil. Environ. Pollut. 2020, 265, 114963. [Google Scholar] [CrossRef]
- Arevalo-Gardini, E.; Canto, M.; Alegre, J.; Loli, O.; Julca, A.; Baligar, V. Changes in soil physical and chemical properties in long term improved natural and traditional agroforestry management systems of cacao genotypes in Peruvian Amazon. PLoS ONE 2015, 10, e0132147. [Google Scholar] [CrossRef]
- Silva, Y.J.A.B.; Freire, M.G.; Lopes, E.A.P.; Santos, M.A. Atriplex nummularia Lindl. as alternative for improving salt-affected soils conditions in semiarid environments: A field experiment. Chil. J. Agric. Res. 2016, 76, 343–348. [Google Scholar]
- Ashraf, M.Y.; Mahmood, K.; Akhter, J.; Hussain, F.; Arshad, M. Phytoremediation of saline soils for sustainable agricultural productivity. In Plant Adaptation and Phytoremediation; Ashraf, M., Ozturk, M., Ahmad, M.S.A., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 335–355. [Google Scholar]
- Qadir, M.; Noble, A.; Oster, J.D.; Schubert, S.; Ghafoor, A. Driving forces for sodium removal during phytoremediation of calcareous sodic and saline-sodic soils: A review. Soil Use Manag. 2005, 21, 173–180. [Google Scholar] [CrossRef]
- Lastiri-Hernández, M.A.; Álvarez-Bernal, D.; Ochoa-Estrada, S.; Contreras-Ramos, S.M. Potential of Bacopa monnieri (L.) Wettst and Sesuvium verrucosum Raf. as an agronomic management alternative to recover the productivity of saline soils. Int. J. Phytoremediation 2019, 22, 343–352. [Google Scholar] [CrossRef]
- Malik, Z.; Ravindran, K. Bioaccumulation of salts and heavy metals by Suaeda monoica: A salt marsh halophyte from paper mill effluent contaminated soil. Int. J. Sci. Technol. Res. 2020, 9, 7248–7254. [Google Scholar]
- Shabala, S. Learning from halophytes: Physiological basis and strategies to improve abiotic stress tolerance in crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef]
- Barros, N.L.F.; Marques, D.N.; Tadaiesky, L.B.A.; de Souza, C.R.B. Halophytes and other molecular strategies for the generation of salt-tolerant crops. Plant Physiol. Biochem. 2021, 162, 581–591. [Google Scholar] [CrossRef]
- Ye, W.; Wang, T.; Wei, W.; Lou, S.; Lan, F.; Zhu, S.; Li, Q.; Ji, G.; Lin, C.; Wu, X.; et al. The full-length transcriptome of Spartina alterniflora reveals the complexity of high salt tolerance in monocotyledonous halophyte. Plant Cell Physiol. 2020, 61, 882–896. [Google Scholar]
- Guo, R.; Zhao, L.; Zhang, K.; Lu, H.; Bhanbhro, N.; Yang, C. Comparative genomics and transcriptomics of the extreme halophyte Puccinellia tenuiflora provides insights into salinity tolerance differentiation between halophytes and glycophytes. Front. Plant Sci. 2021, 12, 767. [Google Scholar] [CrossRef]
- Aliakbari, M.; Razi, H.; Alemzadeh, A.; Tavakol, E. RNA-seq transcriptome profiling of the halophyte Salicornia persica in response to salinity. J. Plant Growth Regul. 2020, 40, 707–721. [Google Scholar] [CrossRef]
- Han, Z.-J.; Sun, Y.; Zhang, M.; Zhai, J.-T. Transcriptomic profile analysis of the halophyte Suaeda rigida response and tolerance under NaCl stress. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Xiang, D.; Man, L. EhEm1, a novel Em-like protein from Eutrema halophilum, confers tolerance to salt and drought stresses in rice. Mol. Breed. 2018, 38, 17. [Google Scholar] [CrossRef]
- Liu, X.; Cai, S.; Wang, G.; Wang, F.; Dong, F.; Mak, M.; Holford, P.; Ji, J.; Salih, A.; Zhou, M.; et al. Halophytic NHXs confer salt tolerance by altering cytosolic and vacuolar K+ and Na+ in Arabidopsis root cell. Plant Growth Regul. 2017, 82, 333–351. [Google Scholar] [CrossRef]
- Geng, X.; Chen, S.; Yilan, E.; Zhang, W.; Mao, H.; Qiqige, A.; Wang, Y.; Qi, Z.; Lin, X. Overexpression of a tonoplast Na+/H+ antiporter from the halophytic shrub Nitraria sibirica improved salt tolerance and root development in transgenic poplar. Tree Genet. Genomes 2020, 16, 1–14. [Google Scholar] [CrossRef]
- Guo, Q.; Meng, S.; Tao, S.; Feng, J.; Fan, X.; Xu, P.; Xu, Z.; Shen, X. Overexpression of a samphire high-affinity potassium transporter gene SbHKT1 enhances salt tolerance in transgenic cotton. Acta Physiol. Plant. 2020, 42, 36. [Google Scholar] [CrossRef]
- Wang, Y.-Y.; Hsu, P.-K.; Tsay, Y.-F. Uptake, allocation and signaling of nitrate. Trends Plant Sci. 2012, 17, 458–467. [Google Scholar] [CrossRef]
- Yuan, J.-Z.; Cui, Y.-N.; Li, X.-T.; Wang, F.-Z.; He, Z.-H.; Li, X.-Y.; Bao, A.-K.; Wang, S.; Ma, Q. ZxNPF7.3/NRT1.5 from the xerophyte Zygophyllum xanthoxylum modulates salt and drought tolerance by regulating NO3-, Na+ and K+ transport. Environ. Exp. Bot. 2020, 177, 104123. [Google Scholar] [CrossRef]
- Lu, L.; Chen, X.; Zhu, L.; Li, M.; Zhang, J.; Yang, X.; Wang, P.; Lu, Y.; Cheng, T.; Shi, J.; et al. NtCIPK9: A calcineurin B-like protein-interacting protein kinase from the halophyte Nitraria tangutorum, enhances Arabidopsis salt tolerance. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Lu, L.; Chen, X.; Wang, P.; Lu, Y.; Zhang, J.; Yang, X.; Cheng, T.; Shi, J.; Chen, J. CIPK11: A calcineurin B-like protein-interacting protein kinase from Nitraria tangutorum, confers tolerance to salt and drought in Arabidopsis. BMC Plant Biol. 2021, 21, 1–16. [Google Scholar] [CrossRef]
- Udawat, P.; Jha, R.K.; Mishra, A.; Jha, B. Overexpression of a plasma membrane-localized SbSRP-like protein enhances salinity and osmotic stress tolerance in transgenic tobacco. Front. Plant Sci. 2017, 8, 582. [Google Scholar] [CrossRef] [Green Version]
- Kumari, J.; Udawat, P.; Dubey, A.K.; Haque, I.; Rathore, M.S.; Jha, B. Overexpression of SbSI-1, A nuclear protein from Salicornia brachiata confers drought and salt stress tolerance and maintains photosynthetic efficiency in transgenic tobacco. Front. Plant Sci. 2017, 8, 1215. [Google Scholar] [CrossRef] [Green Version]
- Shukla, P.S.; Gupta, K.; Agarwal, P.; Jha, B.; Agarwal, P.K. Overexpression of a novel SbMYB15 from Salicornia brachiata confers salinity and dehydration tolerance by reduced oxidative damage and improved photosynthesis in transgenic tobacco. Planta 2015, 242, 1291–1308. [Google Scholar] [CrossRef]
- Gupta, K.; Jha, B.; Agarwal, P.K. A dehydration-responsive element binding (DREB) transcription factor from the succulent halophyte Salicornia brachiata enhances abiotic stress tolerance in transgenic tobacco. Mar. Biotechnol. 2014, 16, 657–673. [Google Scholar] [CrossRef]
- Kumari, A.; Jha, B. Engineering of a novel gene from a halophyte: Potential for agriculture in degraded coastal saline soil. Land Degrad. Dev. 2018, 30, 595–607. [Google Scholar] [CrossRef]
- Agarwal, P.; Baraiya, B.M.; Joshi, P.S.; Patel, M.; Parida, A.K.; Agarwal, P.K. AlRab7 from Aeluropus lagopoides ameliorates ion toxicity in transgenic tobacco by regulating hormone signaling and reactive oxygen species homeostasis. Physiol. Plant. 2021. [Google Scholar] [CrossRef]
- Dabbous, A.; Ben Saad, R.; Brini, F.; Farhat-Khemekhem, A.; Zorrig, W.; Abdely, C.; Ben Hamed, K. Over-expression of a subunit E1 of a vacuolar H+-ATPase gene (Lm VHA-E1) cloned from the halophyte Lobularia maritima improves the tolerance of Arabidopsis thaliana to salt and osmotic stresses. Environ. Exp. Bot. 2017, 137, 128–141. [Google Scholar] [CrossRef]
- Yang, G.; Yu, L.; Zhang, K.; Zhao, Y.; Guo, Y.; Gao, C. A ThDREB gene from Tamarix hispida improved the salt and drought tolerance of transgenic tobacco and T. hispida. Plant Physiol. Biochem. 2017, 113, 187–197. [Google Scholar] [CrossRef]
- Guo, Q.; Tian, X.; Mao, P.; Meng, L. Overexpression of Iris lactea tonoplast Na+/H+ antiporter gene IlNHX confers improved salt tolerance in tobacco. Biol. Plant. 2020, 64, 50–57. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, R.; Mukherjee, A.; Bandyopadhyay, S.; Mukherjee, S.; Sengupta, S.; Ray, S.; Majumder, A.L. Selective manipulation of the inositol metabolic pathway for induction of salt-tolerance in indica rice variety. Sci. Rep. 2019, 9, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Chen, S.; Xie, L.; Lu, Z.; Liu, M.; Han, X.; Qiao, G.; Jiang, J.; Zhuo, R.; Qiu, W.; et al. Overexpression of cysteine protease gene from Salix matsudana enhances salt tolerance in transgenic Arabidopsis. Environ. Exp. Bot. 2018, 147, 53–62. [Google Scholar] [CrossRef]
- Li, N.; Wang, X.; Ma, B.; Du, C.; Zheng, L.; Wang, Y. Expression of a Na+/H+ antiporter RtNHX1 from a recretohalophyte Reaumuria trigyna improved salt tolerance of transgenic Arabidopsis thaliana. J. Plant Physiol. 2017, 218, 109–120. [Google Scholar] [CrossRef]
- Sui, N.; Tian, S.; Wang, W.; Wang, M.; Fan, H. Overexpression of glycerol-3-phosphate acyltransferase from Suaeda salsa improves salt tolerance in Arabidopsis. Front. Plant Sci. 2017, 8, 1337. [Google Scholar] [CrossRef]
- Chang, W.; Liu, X.; Zhu, J.; Fan, W.; Zhang, Z. An aquaporin gene from halophyte Sesuvium portulacastrum, SpAQP1, increases salt tolerance in transgenic tobacco. Plant Cell Rep. 2015, 35, 385–395. [Google Scholar] [CrossRef]
- Chen, Y.; Li, L.; Zong, J.; Chen, J.; Guo, H.; Guo, A.; Liu, J. Heterologous expression of the halophyte Zoysia matrella H+-pyrophosphatase gene improved salt tolerance in Arabidopsis thaliana. Plant Physiol. Biochem. 2015, 91, 49–55. [Google Scholar] [CrossRef]
| Sl. No. | Halophytic Plants | Isolated Genes | Over-Expressed In | Improved Features Contributed to Salt Tolerance | References |
|---|---|---|---|---|---|
| 1 | Nitraria tangutorum | NtCIPK11 | Arabidopsis thaliana | Seed germination rate, root length, plant growth and expression of proline biosynthesis-associated gene (e.g., P5CS1, P5CS2, P5CR, and ProDH1) | [156] |
| 2 | Aeluropus lagopoides | AlRab7 | Nicotiana tabacum | Seed germination rate, phytohormone (indole acetic acid) accumulation, ion homeostasis status, gas-exchange features and antioxidant enzyme activities | [162] |
| 3 | N. sibirica | NsNHX1 | Populus alba | Root system, chlorophyll content, RWC, proline level, Na+ compartmentalization and activities of antioxidant enzymes | [151] |
| 4 | Salicornia bigelovii | SbHKT1 | Gossypium hirsutum | Seed germination rate, biomass accumulation, optimal K+/Na+ ratio and activities of antioxidant enzymes | [152] |
| 5 | N. tangutorum | NtCIPK11 | A. thaliana | Seed germination rate, root length, and expression of K+ transporter gene (AtHKT1) | [155] |
| 6 | Zygophyllum xanthoxylum | ZxNRT1.5 | A. thaliana | Transportation and uptake of K+ from roots to shoots | [154] |
| 7 | Iris lactea | IlNHX | N. tabacum | Higher vacuolar H+-ATPase (V-ATPase) activity for compartmentalization of Na+ into the vacuoles | [165] |
| 8 | S. brachiata | SbRPC5L | N. tabacum | Photosynthetic rate, ion homeostasis, membrane stability and overexpression of antioxidant-related genes (e.g., NtAPX, NtPOX, and NtSOD) | [161] |
| 9 | Porteresia coarctata | PcINO1 and PcIMT1 | Oryza sativa | Seed germination rate, growth and biomass, photosynthetic activity and inositol level | [166] |
| 10 | Salix matsudana | SmCP | A. thaliana | Seed germination rate, photosynthetic pigment levels, antioxidant enzyme activities and root’s ion homeostasis | [167] |
| 11 | Eutrema halophilum | EhEm1 | O. sativa | Germination rate, levels of chlorophyll and proline, and activities of POD and lactate dehydrogenase | [149] |
| 12 | S. brachiata | SbSRP | N. tabacum | RWC, photosynthetic rate, and accumulations of compatible solutes (e.g., proline, sugars, free amino acids, starch, and polyphenols) | [157] |
| 13 | Lobulariamaritima | LmVHA-E1 | A. thaliana | Expression of stress-related genes (e.g., AtNHX, AtP5CS, AtCAT, AtSOD, AtPOD, and AtLEA) | [163] |
| 14 | Tamarix hispida | ThDREB | N. tabacum | Germination rate, root length, plant biomass, chlorophyll content, and activities of SOD and POD | [164] |
| 15 | S. brachiata | SbSI-1 | N. tabacum | RWC, photosynthetic rate, membrane stability, K+/Na+ ratio, polyphenol content, and activities of antioxidant enzymes (e.g., SOD, CAT, and APX) | [158] |
| 16 | Puccinellia tenuiflora and S. europaea | PutNHX1 and SeNHX1 | A. thaliana | Na+ sequestration in the vacuoles, and K+ retention in the cytosol and vacuoles of root cells | [150] |
| 17 | Reaumuria trigyna | RtNHX1 | A. thaliana | Seed germination rate, plant biomass, root length, photosynthetic pigments, RWC, K+/Na+ ratio, proline level, and activities of antioxidant enzymes (e.g., POD and CAT) | [168] |
| 18 | Suaeda salsa | SsGPAT | A. thaliana | Improved content of unsaturated fatty acid helps to alleviate the photoinhibition of photosystem (PSI)-I and PS-II | [169] |
| 19 | Sesuvium portulacastrum | SpAQP1 | N. tabacum | Seed germination rate, root length, and activities antioxidant enzymes (e.g., SOD, POD, and CAT) | [170] |
| 20 | Zoysia matrella | ZmVP1 | A. thaliana | Sequestration of Na+ into vacuoles, K+ assimilation, and activities of antioxidant enzymes (e.g., SOD, POD, and APX) | [171] |
| 21 | S. brachiata | SbMYB15 | N. tabacum | Membrane stability, stomatal conductance, water use efficiency, photosynthetic rate, K+/Na+ ratio, and expression of stress-responsive genes (e.g., LEA5, ERD10, LTP1, HSF2, ADC1, PLC3, and P5CS) | [159] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, M.M.; Mostofa, M.G.; Keya, S.S.; Siddiqui, M.N.; Ansary, M.M.U.; Das, A.K.; Rahman, M.A.; Tran, L.S.-P. Adaptive Mechanisms of Halophytes and Their Potential in Improving Salinity Tolerance in Plants. Int. J. Mol. Sci. 2021, 22, 10733. https://doi.org/10.3390/ijms221910733
Rahman MM, Mostofa MG, Keya SS, Siddiqui MN, Ansary MMU, Das AK, Rahman MA, Tran LS-P. Adaptive Mechanisms of Halophytes and Their Potential in Improving Salinity Tolerance in Plants. International Journal of Molecular Sciences. 2021; 22(19):10733. https://doi.org/10.3390/ijms221910733
Chicago/Turabian StyleRahman, Md. Mezanur, Mohammad Golam Mostofa, Sanjida Sultana Keya, Md. Nurealam Siddiqui, Md. Mesbah Uddin Ansary, Ashim Kumar Das, Md. Abiar Rahman, and Lam Son-Phan Tran. 2021. "Adaptive Mechanisms of Halophytes and Their Potential in Improving Salinity Tolerance in Plants" International Journal of Molecular Sciences 22, no. 19: 10733. https://doi.org/10.3390/ijms221910733
APA StyleRahman, M. M., Mostofa, M. G., Keya, S. S., Siddiqui, M. N., Ansary, M. M. U., Das, A. K., Rahman, M. A., & Tran, L. S.-P. (2021). Adaptive Mechanisms of Halophytes and Their Potential in Improving Salinity Tolerance in Plants. International Journal of Molecular Sciences, 22(19), 10733. https://doi.org/10.3390/ijms221910733