
Inositols: From Established Knowledge to Novel Approaches


 ,
,  ,
,  ,
,  ,
,  , , ,
, , ,  ,
,  , , , ,
, , , , 
 add
Show full author list
add
Show full author list
Abstract
1. Introduction: An Overview on Inositols
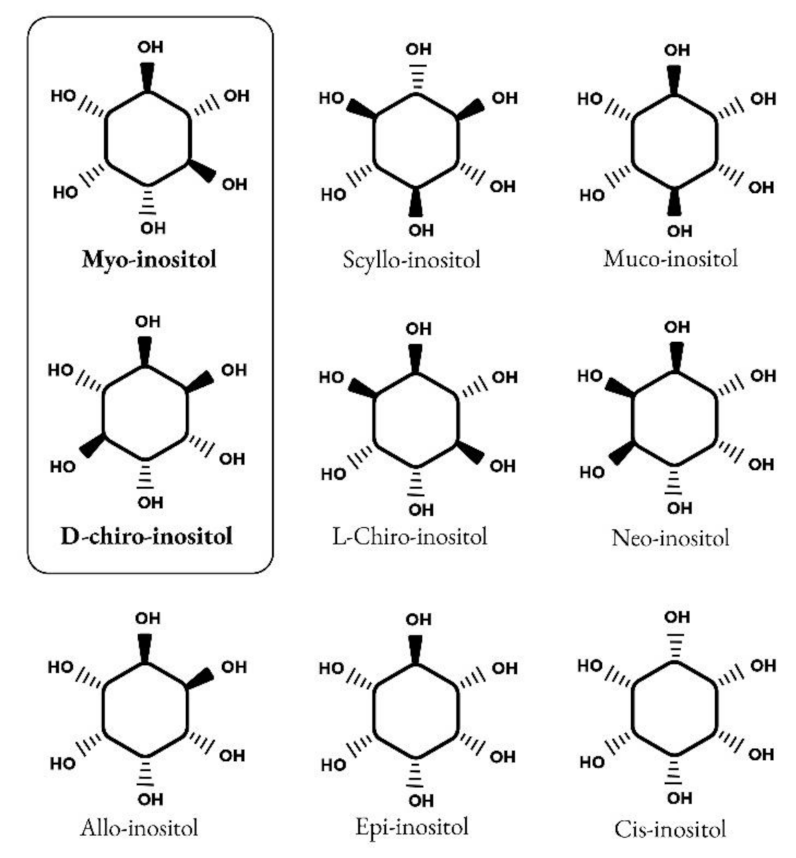
1.1. Inositol Discovery and Biology
1.2. Inositols and Human Reproduction
2. Evidence-Based Effects of Inositols Supplementation on Human Reproduction
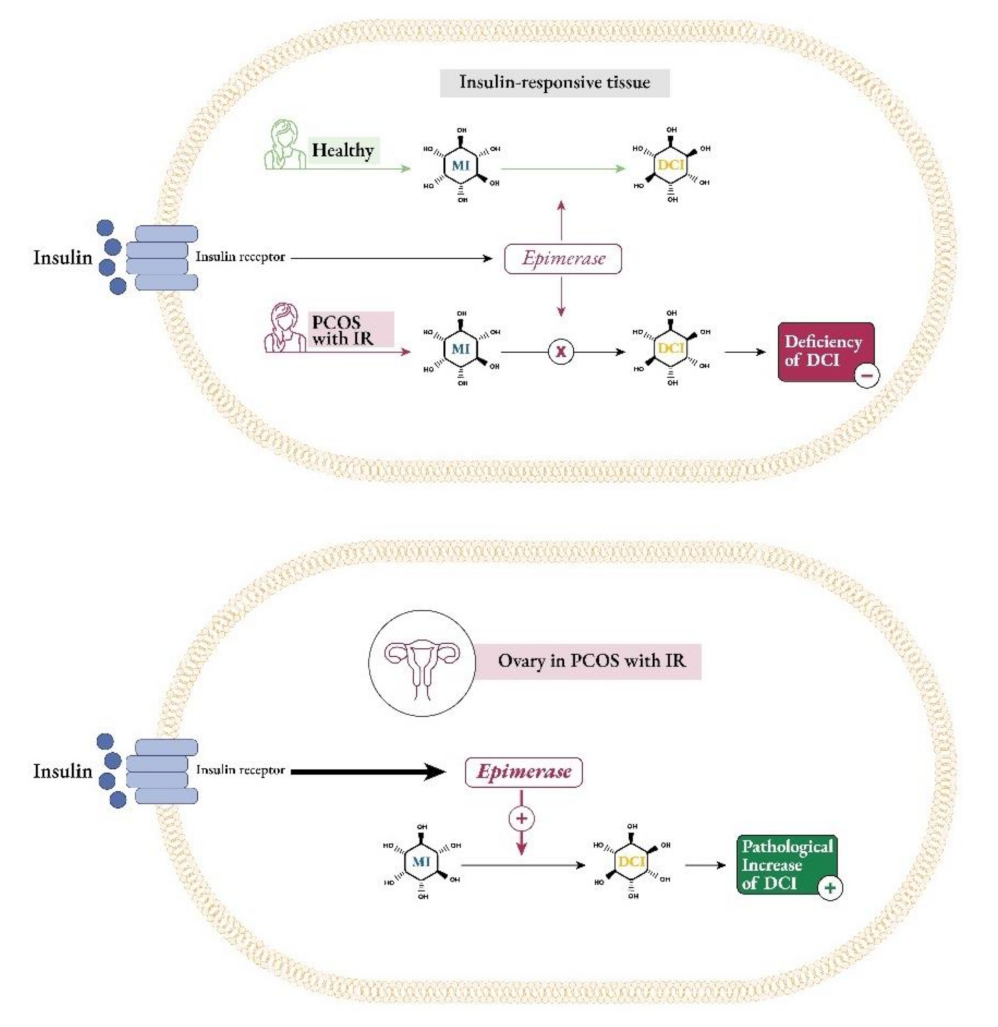
2.1. Inositol Supplementation and Female Reproduction (PCOS Metabolism and Ovulation)
2.2. Pregnancy Development (NTDs in Folic Acid Resistant Women, GDM Prophylaxis)
2.3. Male Reproduction (Impact on Sperm Parameters)
3. Emerging Roles for Inositol Treatments
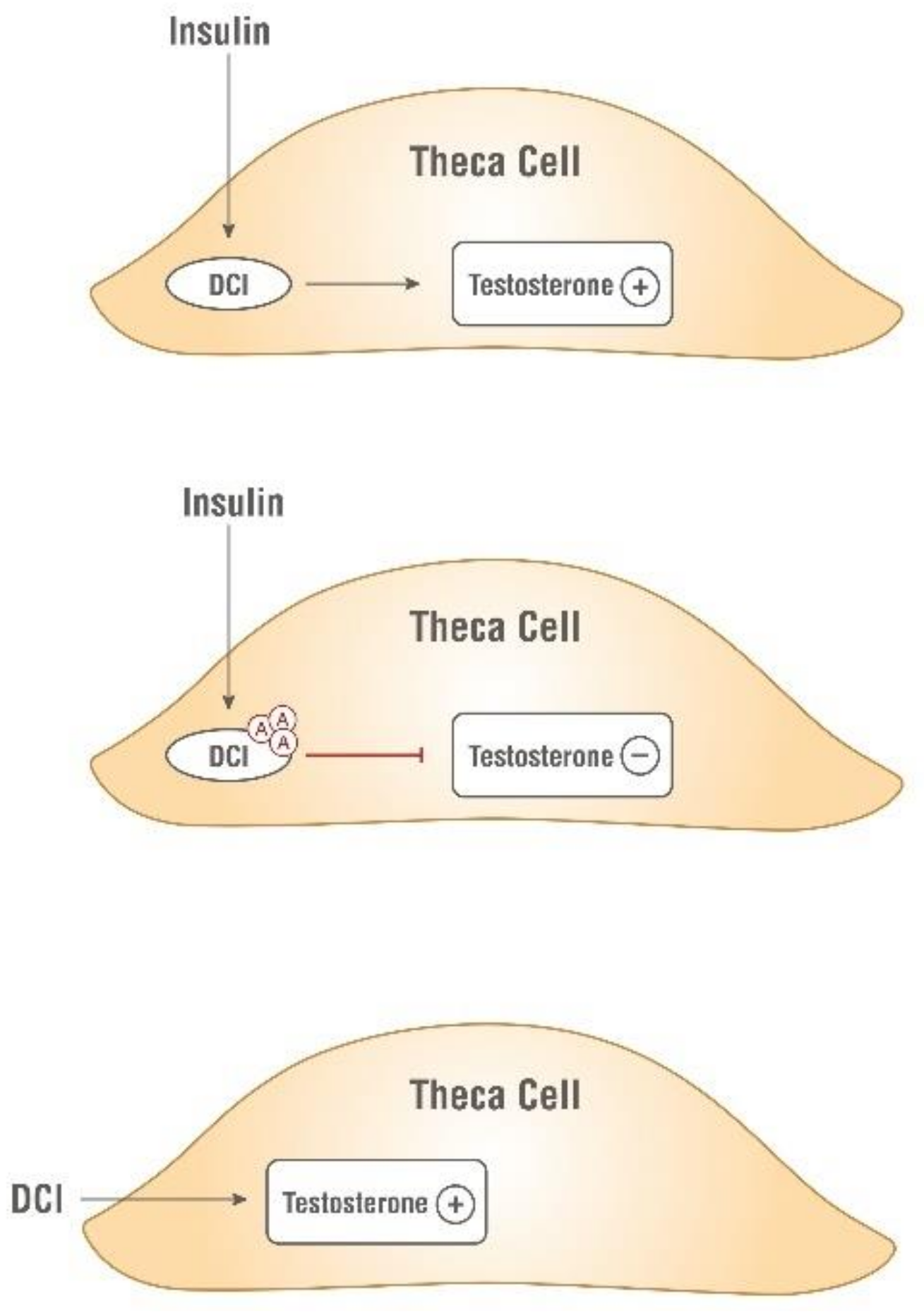
3.1. The Importance of Myo- to D-chiro-Inositol Ratio in Steroidogenesis
3.2. Myo-Inositol Support to Assisted Reproduction Techniques (ART)
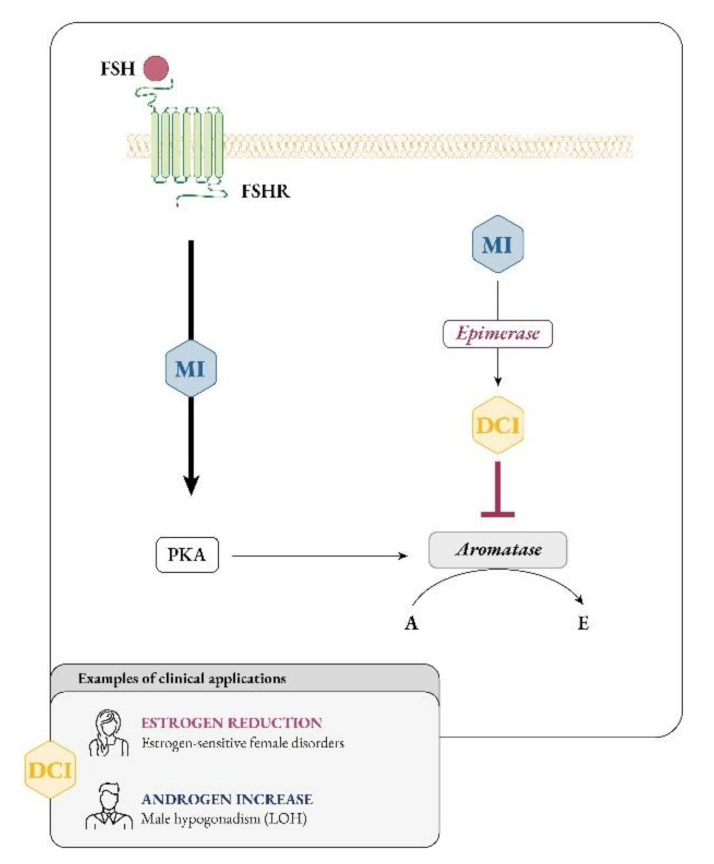
3.3. D-chiro-Inositol Dual Effects (Reduction in Estrogens in Estrogen-Sensitive Female Disorders; Increase in Androgens in Male Hypogonadism)
3.4. D-chiro-Inositol Supplementation: All a Matter of Dose and Time of Administration with a Focus on PCOS
3.5. Enhancing Systemic Absorption of Inositols (Timing and Dose of Administration, Pharmaceutical Techniques)
3.6. The Consequences of Inositol Deficiency Due to Pharmacological Treatments, Malabsorption or Competition with Dietary Glucose
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AEDs | Antiepileptic drugs |
| AIs | Aromatase inhibitors |
| AMH | Anti-Mullerian hormone |
| ART | Assisted Reproductive Technology |
| BBB | Blood brain barrier |
| BMI | Body mass index |
| D-chiro-Ins | D-chiro-inositol |
| E2 | Estradiol |
| EGCG | Epigallocatechin gallate |
| FF | Follicular fluid |
| FSH | Follicle-Stimulating Hormone |
| G6P | Glucose-6-phosphate |
| GDM | Gestational Diabetes Mellitus |
| ICSI | Intracytoplasmic sperm injection |
| IMPA-1 | Inositol monophosphatase-1 |
| IMPase | Inositol monophosphatase-1 |
| InsP2 | Inositol-bisphosphate |
| InsP3 | Inositol-3-phosphate |
| IVF | In Vitro Fertilization |
| LH | Luteinizing Hormone |
| MIPS | myoinositol-phosphate synthase |
| myo-Ins | myo-inositol |
| NTDs | Neural Tube Defects |
| OAT | Oligoasthenoteratozoospermia |
| PCOS | Polycystic Ovary Syndrome |
| SBGH | Sex Binding Globulin Hormone |
| SERMs | Selective Estrogen Receptor Modulators |
| SMIT | Sodium/myo-inositol transporter |
| TRT | Testosterone Replacement Therapy |
| UPA | Ulipristal acetate |
| VA | Valproic acid |
| α-LA | α-lactalbumin |
References
- Scherer. Ueber eine neue, aus dem Muskelfleische gewonnene Zuckerart. Justus Liebigs Annalen Chemie 1850, 73, 322–328. [Google Scholar] [CrossRef]
- Maquenne, L. Préparation, proprietés et constitution se l’inosite. CR Hebd. Séance Acad. Sci. Paris 1887, 104, 225–227. [Google Scholar]
- Wang, W.T.; Safar, J.; Zopf, D. Analysis of inositol by high-performance liquid chromatography. Anal Biochem. 1990, 188, 432–435. [Google Scholar] [CrossRef]
- Murthy, P.P. Structure and nomenclature of inositol phosphates, phosphoinositides, and glycosylphosphatidylinositols. Subcell Biochem. 2006, 39, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.M.; Yamauchi, A.; Uchida, S.; Preston, A.S.; Garcia-Perez, A.; Burg, M.B.; Handler, J.S. Cloning of the cDNa for a Na+/myo-inositol cotransporter, a hypertonicity stress protein. J. Biol. Chem. 1992, 267, 6297–6301. [Google Scholar] [CrossRef]
- Hitomi, K.; Tsukagoshi, N. cDNA sequence for rkST1, a novel member of the sodium ion-dependent glucose cotransporter family. Biochim. Biophys. Acta 1994, 1190, 469–472. [Google Scholar] [CrossRef]
- Garzon, S.; Laganà, A.S.; Monastra, G. Risk of reduced intestinal absorption of myo-inositol caused by D-chiro-inositol or by glucose transporter inhibitors. Expert Opin. Drug Metab. Toxicol. 2019, 15, 697–703. [Google Scholar] [CrossRef]
- Berry, G.T.; Mallee, J.J.; Kwon, H.M.; Rim, J.S.; Mulla, W.R.; Muenke, M.; Spinner, N.B. The human osmoregulatory Na+/myo-inositol cotransporter gene (SLC5A3): Molecular cloning and localization to chromosome 21. Genomics 1995, 25, 507–513. [Google Scholar] [CrossRef]
- Miyakawa, H.; Woo, S.K.; Dahl, S.C.; Handler, J.S.; Kwon, H.M. Tonicity-responsive enhancer binding protein, a rel-like protein that stimulates transcription in response to hypertonicity. Proc. Natl. Acad. Sci. USA 1999, 96, 2538–2542. [Google Scholar] [CrossRef]
- Majunder, A.; Biswas, B. (Eds.) Biology of inositols and phosphoinositides. In Subcellular Biochemistry; Springer: New York, NY, USA, 2006. [Google Scholar]
- Dinicola, S.; Minini, M.; Unfer, V.; Verna, R.; Cucina, A.; Bizzarri, M. Nutritional and Acquired Deficiencies in Inositol Bioavailability. Correlations with Metabolic Disorders. Int. J. Mol. Sci. 2017, 18, 2187. [Google Scholar] [CrossRef]
- Wong, Y.H.; Kalmbach, S.J.; Hartman, B.K.; Sherman, W.R. Immunohistochemical staining and enzyme activity measurements show myo-inositol-1-phosphate synthase to be localized in the vasculature of brain. J. Neurochem. 1987, 48, 1434–1442. [Google Scholar] [CrossRef] [PubMed]
- Loewus, M.W.; Loewus, F.A.; Brillinger, G.U.; Otsuka, H.; Floss, H.G. Stereochemistry of the myo-inositol-1-phosphate synthase reaction. J. Biol. Chem. 1980, 255, 11710–11712. [Google Scholar] [CrossRef]
- Bizzarri, M.; Fuso, A.; Dinicola, S.; Cucina, A.; Bevilacqua, A. Pharmacodynamics and pharmacokinetics of inositol(s) in health and disease. Expert Opin. Drug Metab. Toxicol. 2016, 12, 1181–1196. [Google Scholar] [CrossRef] [PubMed]
- Ijuin, T.; Takenawa, T. Regulation of insulin signaling and glucose transporter 4 (GLUT4) exocytosis by phosphatidylinositol 3,4,5-trisphosphate (PIP3) phosphatase, skeletal muscle, and kidney enriched inositol polyphosphate phosphatase (SKIP). J. Biol. Chem. 2012, 287, 6991–6999. [Google Scholar] [CrossRef]
- Nestler, J.E.; Unfer, V. Reflections on inositol(s) for PCOS therapy: Steps toward success. Gynecol. Endocrinol. 2015, 31, 501–505. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Bizzarri, M. Inositols in Insulin Signaling and Glucose Metabolism. Int. J. Endocrinol. 2018, 2018, 1968450. [Google Scholar] [CrossRef]
- Sun, T.H.; Heimark, D.B.; Nguygen, T.; Nadler, J.L.; Larner, J. Both myo-inositol to chiro-inositol epimerase activities and chiro-inositol to myo-inositol ratios are decreased in tissues of GK type 2 diabetic rats compared to Wistar controls. Biochem. Biophys. Res. Commun. 2002, 293, 1092–1098. [Google Scholar] [CrossRef]
- Fan, C.; Liang, W.; Wei, M.; Gou, X.; Han, S.; Bai, J. Effects of D-Chiro-Inositol on Glucose Metabolism in db/db Mice and the Associated Underlying Mechanisms. Front. Pharmacol. 2020, 11, 354. [Google Scholar] [CrossRef]
- Yap, A.; Nishiumi, S.; Yoshida, K.; Ashida, H. Rat L6 myotubes as an in vitro model system to study GLUT4-dependent glucose uptake stimulated by inositol derivatives. Cytotechnology 2007, 55, 103–108. [Google Scholar] [CrossRef]
- Heimark, D.; McAllister, J.; Larner, J. Decreased myo-inositol to chiro-inositol (M/C) ratios and increased M/C epimerase activity in PCOS theca cells demonstrate increased insulin sensitivity compared to controls. Endocr. J. 2014, 61, 111–117. [Google Scholar] [CrossRef]
- Larner, J.; Huang, L.C.; Tang, G.; Suzuki, S.; Schwartz, C.F.; Romero, G.; Roulidis, Z.; Zeller, K.; Shen, T.Y.; Oswald, A.S.; et al. Insulin mediators: Structure and formation. Cold Spring Harb. Symp. Quant. Biol. 1988, 53 Pt 2, 965–971. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Cruz, H.; Oróstica, L.; Plaza-Parrochia, F.; Torres-Pinto, I.; Romero, C.; Vega, M. The insulin-sensitizing mechanism of myo-inositol is associated with AMPK activation and GLUT-4 expression in human endometrial cells exposed to a PCOS environment. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E237–e248. [Google Scholar] [CrossRef] [PubMed]
- Larner, J.; Romero, G.; Kennington, A.S.; Lilley, K.; Kilgour, E.; Zhang, C.; Heimark, D.; Gamez, G.; Houston, D.B.; Huang, L.C. Duality in the mechanism of action of insulin. Adv. Second Messenger Phosphoprotein Res. 1990, 24, 290–294. [Google Scholar] [PubMed]
- Larner, J.; Price, J.D.; Heimark, D.; Smith, L.; Rule, G.; Piccariello, T.; Fonteles, M.C.; Pontes, C.; Vale, D.; Huang, L. Isolation, structure, synthesis, and bioactivity of a novel putative insulin mediator. A galactosamine chiro-inositol pseudo-disaccharide Mn2+ chelate with insulin-like activity. J. Med. Chem. 2003, 46, 3283–3291. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.C.; Fonteles, M.C.; Houston, D.B.; Zhang, C.; Larner, J. Chiroinositol deficiency and insulin resistance. III. Acute glycogenic and hypoglycemic effects of two inositol phosphoglycan insulin mediators in normal and streptozotocin-diabetic rats in vivo. Endocrinology 1993, 132, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Nestler, J.E.; Romero, G.; Huang, L.C.; Zhang, C.G.; Larner, J. Insulin mediators are the signal transduction system responsible for insulin’s actions on human placental steroidogenesis. Endocrinology 1991, 129, 2951–2956. [Google Scholar] [CrossRef]
- Nestler, J.E.; Jakubowicz, D.J.; de Vargas, A.F.; Brik, C.; Quintero, N.; Medina, F. Insulin stimulates testosterone biosynthesis by human thecal cells from women with polycystic ovary syndrome by activating its own receptor and using inositolglycan mediators as the signal transduction system. J. Clin. Endocrinol. Metab. 1998, 83, 2001–2005. [Google Scholar] [CrossRef]
- Milewska, E.M.; Czyzyk, A.; Meczekalski, B.; Genazzani, A.D. Inositol and human reproduction. From cellular metabolism to clinical use. Gynecol. Endocrinol. 2016, 32, 690–695. [Google Scholar] [CrossRef] [PubMed]
- Ravanos, K.; Monastra, G.; Pavlidou, T.; Goudakou, M.; Prapas, N. Can high levels of D-chiro-inositol in follicular fluid exert detrimental effects on blastocyst quality? Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 5491–5498. [Google Scholar] [CrossRef]
- Sacchi, S.; Marinaro, F.; Tondelli, D.; Lui, J.; Xella, S.; Marsella, T.; Tagliasacchi, D.; Argento, C.; Tirelli, A.; Giulini, S.; et al. Modulation of gonadotrophin induced steroidogenic enzymes in granulosa cells by d-chiroinositol. Reprod. Biol. Endocrinol. 2016, 14, 52. [Google Scholar] [CrossRef]
- Larner, J. D-chiro-inositol—Its functional role in insulin action and its deficit in insulin resistance. Int. J. Exp. Diabetes Res. 2002, 3, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Rotterdam ESHRE/ASRM-Sponsored PCOS consensus workshop group. Revised 2003 consensus on diagnostic criteria and long-term health risks related to polycystic ovary syndrome (PCOS). Hum. Reprod. 2004, 19, 41–47. [Google Scholar] [CrossRef]
- Nestler, J.E. Role of hyperinsulinemia in the pathogenesis of the polycystic ovary syndrome, and its clinical implications. Semin. Reprod. Endocrinol. 1997, 15, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Baillargeon, J.P.; Iuorno, M.J.; Nestler, J.E. Insulin sensitizers for polycystic ovary syndrome. Clin. Obstet. Gynecol. 2003, 46, 325–340. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, M.A.; Deshmukh, H.; Atkin, S.; Sathyapalan, T. A review of therapeutic options for managing the metabolic aspects of polycystic ovary syndrome. Ther. Adv. Endocrinol. Metab. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Facchinetti, F.; Bizzarri, M.; Benvenga, S.; D’Anna, R.; Lanzone, A.; Soulage, C.; Di Renzo, G.C.; Hod, M.; Cavalli, P.; Chiu, T.T.; et al. Results from the International Consensus Conference on Myo-inositol and d-chiro-inositol in Obstetrics and Gynecology: The link between metabolic syndrome and PCOS. Eur. J. Obstet. Gynecol. Reprod. Biol. 2015, 195, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Kennington, A.S.; Hill, C.R.; Craig, J.; Bogardus, C.; Raz, I.; Ortmeyer, H.K.; Hansen, B.C.; Romero, G.; Larner, J. Low urinary chiro-inositol excretion in non-insulin-dependent diabetes mellitus. N. Engl. J. Med. 1990, 323, 373–378. [Google Scholar] [CrossRef]
- Carlomagno, G.; Unfer, V.; Roseff, S. The D-chiro-inositol paradox in the ovary. Fertil. Steril. 2011, 95, 2515–2516. [Google Scholar] [CrossRef]
- Unfer, V.; Carlomagno, G.; Papaleo, E.; Vailati, S.; Candiani, M.; Baillargeon, J.P. Hyperinsulinemia Alters Myoinositol to d-chiroinositol Ratio in the Follicular Fluid of Patients With PCOS. Reprod. Sci. 2014, 21, 854–858. [Google Scholar] [CrossRef]
- Nestler, J.E.; Jakubowicz, D.J.; Reamer, P.; Gunn, R.D.; Allan, G. Ovulatory and metabolic effects of D-chiro-inositol in the polycystic ovary syndrome. N. Engl. J. Med. 1999, 340, 1314–1320. [Google Scholar] [CrossRef]
- Cheang, K.I.; Baillargeon, J.P.; Essah, P.A.; Ostlund, R.E., Jr.; Apridonize, T.; Islam, L.; Nestler, J.E. Insulin-stimulated release of D-chiro-inositol-containing inositolphosphoglycan mediator correlates with insulin sensitivity in women with polycystic ovary syndrome. Metabolism 2008, 57, 1390–1397. [Google Scholar] [CrossRef] [PubMed]
- Unfer, V.; Facchinetti, F.; Orrù, B.; Giordani, B.; Nestler, J. Myo-inositol effects in women with PCOS: A meta-analysis of randomized controlled trials. Endocr. Connect 2017, 6, 647–658. [Google Scholar] [CrossRef]
- Nordio, M.; Basciani, S.; Camajani, E. The 40:1 myo-inositol/D-chiro-inositol plasma ratio is able to restore ovulation in PCOS patients: Comparison with other ratios. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5512–5521. [Google Scholar] [CrossRef] [PubMed]
- Thalamati, S. A comparative study of combination of Myo-inositol and D-chiro-inositol versus Metformin in the management of polycystic ovary syndrome in obese women with infertility. Int. J. Reprod. Contracept. Obstet. Gynecol. 2019, 8, 825–829. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Dragotto, J.; Giuliani, A.; Bizzarri, M. Myo-inositol and D-chiro-inositol (40:1) reverse histological and functional features of polycystic ovary syndrome in a mouse model. J. Cell Physiol. 2019, 234, 9387–9398. [Google Scholar] [CrossRef] [PubMed]
- Benelli, E.; Del Ghianda, S.; Di Cosmo, C.; Tonacchera, M. A Combined Therapy with Myo-Inositol and D-Chiro-Inositol Improves Endocrine Parameters and Insulin Resistance in PCOS Young Overweight Women. Int. J. Endocrinol. 2016, 2016, 3204083. [Google Scholar] [CrossRef] [PubMed]
- Minozzi, M.; Nordio, M.; Pajalich, R. The Combined therapy myo-inositol plus D-Chiro-inositol, in a physiological ratio, reduces the cardiovascular risk by improving the lipid profile in PCOS patients. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 537–540. [Google Scholar] [PubMed]
- Nordio, M.; Proietti, E. The combined therapy with myo-inositol and D-chiro-inositol reduces the risk of metabolic disease in PCOS overweight patients compared to myo-inositol supplementation alone. Eur. Rev. Med. Pharmacol. Sci. 2012, 16, 575–581. [Google Scholar]
- Le Donne, M.; Metro, D.; Alibrandi, A.; Papa, M.; Benvenga, S. Effects of three treatment modalities (diet, myoinositol or myoinositol associated with D-chiro-inositol) on clinical and body composition outcomes in women with polycystic ovary syndrome. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 2293–2301. [Google Scholar] [CrossRef]
- Roseff, S.; Montenegro, M. Inositol Treatment for PCOS Should Be Science-Based and Not Arbitrary. Int. J. Endocrinol. 2020, 2020, 6461254. [Google Scholar] [CrossRef]
- Monastra, G.; Unfer, V.; Harrath, A.H.; Bizzarri, M. Combining treatment with myo-inositol and D-chiro-inositol (40:1) is effective in restoring ovary function and metabolic balance in PCOS patients. Gynecol. Endocrinol. 2017, 33, 1–9. [Google Scholar] [CrossRef]
- Facchinetti, F.; Unfer, V.; Dewailly, D.; Kamenov, Z.A.; Diamanti-Kandarakis, E.; Laganà, A.S.; Nestler, J.E.; Soulage, C.O. Inositols in Polycystic Ovary Syndrome: An Overview on the Advances. Trends Endocrinol. Metab. 2020, 31, 435–447. [Google Scholar] [CrossRef]
- Facchinetti, F.; Appetecchia, M.; Aragona, C.; Bevilacqua, A.; Bezerra Espinola, M.S.; Bizzarri, M.; D’Anna, R.; Dewailly, D.; Diamanti-Kandarakis, E.; Hernández Marín, I.; et al. Experts’ opinion on inositols in treating polycystic ovary syndrome and non-insulin dependent diabetes mellitus: A further help for human reproduction and beyond. Expert Opin. Drug Metab. Toxicol. 2020, 16, 255–274. [Google Scholar] [CrossRef]
- Mendoza, N.; Diaz-Ropero, M.P.; Aragon, M.; Maldonado, V.; Llaneza, P.; Lorente, J.; Mendoza-Tesarik, R.; Maldonado-Lobon, J.; Olivares, M.; Fonolla, J. Comparison of the effect of two combinations of myo-inositol and D-chiro-inositol in women with polycystic ovary syndrome undergoing ICSI: A randomized controlled trial. Gynecol. Endocrinol. 2019, 35, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Bevilacqua, A.; Dragotto, J.; Lucarelli, M.; Di Emidio, G.; Monastra, G.; Tatone, C. High Doses of D-Chiro-Inositol Alone Induce a PCO-Like Syndrome and Other Alterations in Mouse Ovaries. Int. J. Mol. Sci. 2021, 22, 5691. [Google Scholar] [CrossRef] [PubMed]
- Chiefari, E.; Arcidiacono, B.; Foti, D.; Brunetti, A. Gestational diabetes mellitus: An updated overview. J. Endocrinol. Invest. 2017, 40, 899–909. [Google Scholar] [CrossRef] [PubMed]
- Battista, M.C.; Hivert, M.F.; Duval, K.; Baillargeon, J.P. Intergenerational cycle of obesity and diabetes: How can we reduce the burdens of these conditions on the health of future generations? Exp. Diabetes Res. 2011, 2011, 596060. [Google Scholar] [CrossRef]
- Hod, M.; Pretty, M.; Mahmood, T. Joint position statement on universal screening for GDM in Europe by FIGO, EBCOG and EAPM. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 228, 329–330. [Google Scholar] [CrossRef]
- Simmons, D.; Walters, B.N.; Rowan, J.A.; McIntyre, H.D. Metformin therapy and diabetes in pregnancy. Med. J. Aust. 2004, 180, 462–464. [Google Scholar] [CrossRef]
- Hyer, S.; Balani, J.; Shehata, H. Metformin in Pregnancy: Mechanisms and Clinical Applications. Int. J. Mol. Sci. 2018, 19, 1954. [Google Scholar] [CrossRef]
- Simeonova-Krstevska, S.; Bogoev, M.; Bogoeva, K.; Zisovska, E.; Samardziski, I.; Velkoska-Nakova, V.; Livrinova, V.; Todorovska, I.; Sima, A.; Blazevska-Siljanoska, V. Maternal and Neonatal Outcomes in Pregnant Women with Gestational Diabetes Mellitus Treated with Diet, Metformin or Insulin. Open Access Maced. J. Med. Sci. 2018, 6, 803–807. [Google Scholar] [CrossRef]
- Syngelaki, A.; Nicolaides, K.H.; Balani, J.; Hyer, S.; Akolekar, R.; Kotecha, R.; Pastides, A.; Shehata, H. Metformin versus Placebo in Obese Pregnant Women without Diabetes Mellitus. N. Engl. J. Med. 2016, 374, 434–443. [Google Scholar] [CrossRef]
- Chiswick, C.; Reynolds, R.M.; Denison, F.; Drake, A.J.; Forbes, S.; Newby, D.E.; Walker, B.R.; Quenby, S.; Wray, S.; Weeks, A.; et al. Effect of metformin on maternal and fetal outcomes in obese pregnant women (EMPOWaR): A randomised, double-blind, placebo-controlled trial. Lancet Diabetes Endocrinol. 2015, 3, 778–786. [Google Scholar] [CrossRef]
- Begum, M.R.; Khanam, N.N.; Quadir, E.; Ferdous, J.; Begum, M.S.; Khan, F.; Begum, A. Prevention of gestational diabetes mellitus by continuing metformin therapy throughout pregnancy in women with polycystic ovary syndrome. J. Obstet. Gynaecol. Res. 2009, 35, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Glueck, C.J.; Goldenberg, N.; Wang, P.; Loftspring, M.; Sherman, A. Metformin during pregnancy reduces insulin, insulin resistance, insulin secretion, weight, testosterone and development of gestational diabetes: Prospective longitudinal assessment of women with polycystic ovary syndrome from preconception throughout pregnancy. Hum. Reprod. 2004, 19, 510–521. [Google Scholar] [CrossRef] [PubMed]
- Glueck, C.J.; Pranikoff, J.; Aregawi, D.; Wang, P. Prevention of gestational diabetes by metformin plus diet in patients with polycystic ovary syndrome. Fertil. Steril. 2008, 89, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Vanky, E.; Stridsklev, S.; Heimstad, R.; Romundstad, P.; Skogøy, K.; Kleggetveit, O.; Hjelle, S.; von Brandis, P.; Eikeland, T.; Flo, K.; et al. Metformin versus placebo from first trimester to delivery in polycystic ovary syndrome: A randomized, controlled multicenter study. J. Clin. Endocrinol. Metab. 2010, 95, E448–E455. [Google Scholar] [CrossRef]
- Tahir, F.; Majid, Z. Inositol Supplementation in the Prevention of Gestational Diabetes Mellitus. Cureus 2019, 11, e5671. [Google Scholar] [CrossRef]
- Facchinetti, F.; Cavalli, P.; Copp, A.J.; D’Anna, R.; Kandaraki, E.; Greene, N.D.E.; Unfer, V.; Experts Group on Inositol in Basic and Clinical Research. An update on the use of inositols in preventing gestational diabetes mellitus (GDM) and neural tube defects (NTDs). Expert Opin. Drug Metab. Toxicol. 2020, 16, 1187–1198. [Google Scholar] [CrossRef]
- Asimakopoulos, G.; Pergialiotis, V.; Anastasiou, E.; Antsaklis, P.; Theodora, M.; Vogiatzi, E.; Kallergi, A.; Sindos, M.; Loutradis, D.; Daskalakis, G. Effect of dietary myo-inositol supplementation on the insulin resistance and the prevention of gestational diabetes mellitus: Study protocol for a randomized controlled trial. Trials 2020, 21, 633. [Google Scholar] [CrossRef]
- Vitale, S.G.; Corrado, F.; Caruso, S.; Di Benedetto, A.; Giunta, L.; Cianci, A.; D’Anna, R. Myo-inositol supplementation to prevent gestational diabetes in overweight non-obese women: Bioelectrical impedance analysis, metabolic aspects, obstetric and neonatal outcomes - a randomized and open-label, placebo-controlled clinical trial. Int. J. Food Sci. Nutr. 2020, 1–10. [Google Scholar] [CrossRef]
- Gambioli, R.; Forte, G.; Buzzaccarini, G.; Unfer, V.; Laganà, A.S. Myo-Inositol as a Key Supporter of Fertility and Physiological Gestation. Pharmaceuticals 2021, 14, 504. [Google Scholar] [CrossRef]
- D’Anna, R.; Corrado, F.; Loddo, S.; Gullo, G.; Giunta, L.; Di Benedetto, A. Myoinositol plus α-lactalbumin supplementation, insulin resistance and birth outcomes in women with gestational diabetes mellitus: A randomized, controlled study. Sci. Rep. 2021, 11, 8866. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, K.M.; Barton, S.J.; El-Heis, S.; Kenealy, T.; Nield, H.; Baker, P.N.; Chong, Y.S.; Cutfield, W.; Chan, S.Y. Myo-Inositol, Probiotics, and Micronutrient Supplementation from Preconception for Glycemia in Pregnancy: NiPPeR International Multicenter Double-Blind Randomized Controlled Trial. Diabetes Care 2021, 44, 1091–1099. [Google Scholar] [CrossRef] [PubMed]
- Greene, N.D.; Leung, K.Y.; Copp, A.J. Inositol, neural tube closure and the prevention of neural tube defects. Birth Defects Res. 2017, 109, 68–80. [Google Scholar] [CrossRef]
- Cockroft, D.L. Changes with gestational age in the nutritional requirements of postimplantation rat embryos in culture. Teratology 1988, 38, 281–290. [Google Scholar] [CrossRef] [PubMed]
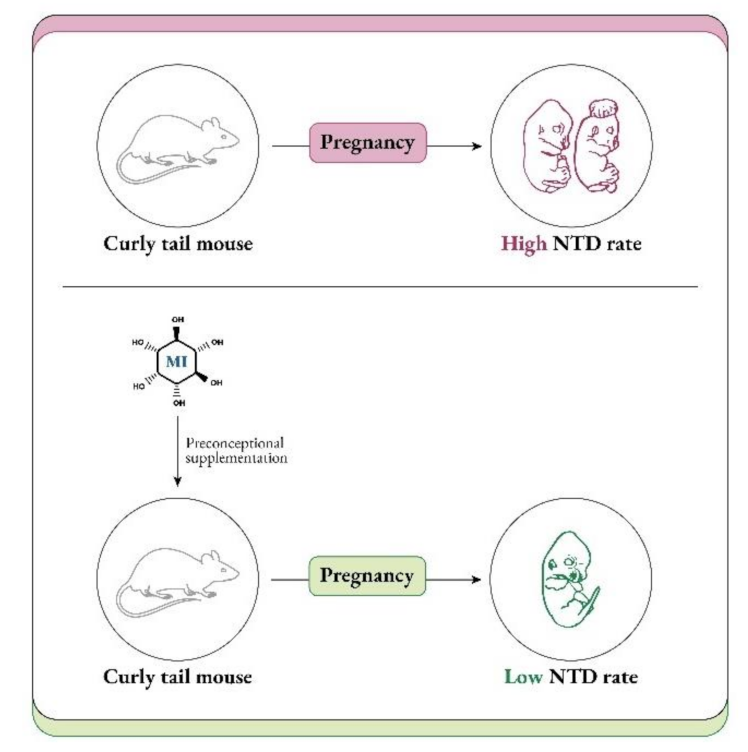
- Cockroft, D.L.; Brook, F.A.; Copp, A.J. Inositol deficiency increases the susceptibility to neural tube defects of genetically predisposed (curly tail) mouse embryos in vitro. Teratology 1992, 45, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Groenen, P.M.; Peer, P.G.; Wevers, R.A.; Swinkels, D.W.; Franke, B.; Mariman, E.C.; Steegers-Theunissen, R.P. Maternal myo-inositol, glucose, and zinc status is associated with the risk of offspring with spina bifida. Am. J. Obstet. Gynecol. 2003, 189, 1713–1719. [Google Scholar] [CrossRef]
- Cindrova-Davies, T.; Jauniaux, E.; Elliot, M.G.; Gong, S.; Burton, G.J.; Charnock-Jones, D.S. RNA-seq reveals conservation of function among the yolk sacs of human, mouse, and chicken. Proc. Natl. Acad. Sci. USA 2017, 114, e4753–e4761. [Google Scholar] [CrossRef]
- Guan, G.; Dai, P.; Shechter, I. cDNA cloning and gene expression analysis of human myo-inositol 1-phosphate synthase. Arch. Biochem. Biophys. 2003, 417, 251–259. [Google Scholar] [CrossRef]
- Groenen, P.M.; Klootwijk, R.; Schijvenaars, M.M.; Straatman, H.; Mariman, E.C.; Franke, B.; Steegers-Theunissen, R.P. Spina bifida and genetic factors related to myo-inositol, glucose, and zinc. Mol. Genet. Metab. 2004, 82, 154–161. [Google Scholar] [CrossRef]
- Jauniaux, E.; Hempstock, J.; Teng, C.; Battaglia, F.C.; Burton, G.J. Polyol concentrations in the fluid compartments of the human conceptus during the first trimester of pregnancy: Maintenance of redox potential in a low oxygen environment. J. Clin. Endocrinol. Metab. 2005, 90, 1171–1175. [Google Scholar] [CrossRef]
- D’Souza, S.W.; Copp, A.J.; Greene, N.D.E.; Glazier, J.D. Maternal Inositol Status and Neural Tube Defects: A Role for the Human Yolk Sac in Embryonic Inositol Delivery? Adv. Nutr. 2021, 12, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Gustavsson, P.; Copp, A.J.; Greene, N.D. Grainyhead genes and mammalian neural tube closure. Birth Defects Res. A Clin. Mol. Teratol. 2008, 82, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Greene, N.D.; Copp, A.J. Inositol prevents folate-resistant neural tube defects in the mouse. Nat. Med. 1997, 3, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Cogram, P.; Hynes, A.; Dunlevy, L.P.; Greene, N.D.; Copp, A.J. Specific isoforms of protein kinase C are essential for prevention of folate-resistant neural tube defects by inositol. Hum. Mol. Genet. 2004, 13, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Cogram, P.; Tesh, S.; Tesh, J.; Wade, A.; Allan, G.; Greene, N.D.; Copp, A.J. D-chiro-inositol is more effective than myo-inositol in preventing folate-resistant mouse neural tube defects. Hum. Reprod. 2002, 17, 2451–2458. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, F.C.; Meschia, G.; Blechner, J.N.; Barron, D.H. The free myo-inositol concentration of adult and fetal tissues of several species. Q. J. Exp. Physiol. Cogn. Med. Sci. 1961, 46, 188–193. [Google Scholar] [CrossRef]
- Di Paolo, G.; De Camilli, P. Phosphoinositides in cell regulation and membrane dynamics. Nature 2006, 443, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Goto, M.P.; Goldman, A.S. Diabetic embryopathy. Curr. Opin. Pediatr. 1994, 6, 486–491. [Google Scholar] [CrossRef]
- Rasmussen, S.A.; Chu, S.Y.; Kim, S.Y.; Schmid, C.H.; Lau, J. Maternal obesity and risk of neural tube defects: A metaanalysis. Am. J. Obstet. Gynecol. 2008, 198, 611–619. [Google Scholar] [CrossRef]
- Hendricks, K.A.; Nuno, O.M.; Suarez, L.; Larsen, R. Effects of hyperinsulinemia and obesity on risk of neural tube defects among Mexican Americans. Epidemiology 2001, 12, 630–635. [Google Scholar] [CrossRef] [PubMed]
- Ray, J.G.; Thompson, M.D.; Vermeulen, M.J.; Meier, C.; Wyatt, P.R.; Wong, P.Y.; Summers, A.M.; Farrell, S.A.; Cole, D.E. Metabolic syndrome features and risk of neural tube defects. BMC Pregnancy Childbirth 2007, 7, 21. [Google Scholar] [CrossRef]
- Shaw, G.M.; Quach, T.; Nelson, V.; Carmichael, S.L.; Schaffer, D.M.; Selvin, S.; Yang, W. Neural tube defects associated with maternal periconceptional dietary intake of simple sugars and glycemic index. Am. J. Clin. Nutr. 2003, 78, 972–978. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yazdy, M.M.; Liu, S.; Mitchell, A.A.; Werler, M.M. Maternal dietary glycemic intake and the risk of neural tube defects. Am. J. Epidemiol. 2010, 171, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Greene, D.A. Sorbitol, myo-inositol and sodium-potassium ATPase in diabetic peripheral nerve. Drugs 1986, 32 (Suppl. 2), 6–14. [Google Scholar] [CrossRef]
- Hod, M.; Star, S.; Passonneau, J.V.; Unterman, T.G.; Freinkel, N. Effect of hyperglycemia on sorbitol and myo-inositol content of cultured rat conceptus: Failure of aldose reductase inhibitors to modify myo-inositol depletion and dysmorphogenesis. Biochem. Biophys. Res. Commun. 1986, 140, 974–980. [Google Scholar] [CrossRef]
- Sussman, I.; Matschinsky, F.M. Diabetes affects sorbitol and myo-inositol levels of neuroectodermal tissue during embryogenesis in rat. Diabetes 1988, 37, 974–981. [Google Scholar] [CrossRef]
- Baker, L.; Piddington, R.; Goldman, A.; Egler, J.; Moehring, J. Myo-inositol and prostaglandins reverse the glucose inhibition of neural tube fusion in cultured mouse embryos. Diabetologia 1990, 33, 593–596. [Google Scholar] [CrossRef]
- Hod, M.; Star, S.; Passonneau, J.; Unterman, T.G.; Freinkel, N. Glucose-induced dysmorphogenesis in the cultured rat conceptus: Prevention by supplementation with myo-inositol. Isr. J. Med. Sci. 1990, 26, 541–544. [Google Scholar]
- Greene, N.D.; Copp, A.J. Neural tube defects. Annu. Rev. Neurosci. 2014, 37, 221–242. [Google Scholar] [CrossRef]
- Berry, R.J.; Li, Z.; Erickson, J.D.; Li, S.; Moore, C.A.; Wang, H.; Mulinare, J.; Zhao, P.; Wong, L.Y.; Gindler, J.; et al. Prevention of neural-tube defects with folic acid in China. China-U.S. Collaborative Project for Neural Tube Defect Prevention. N. Engl. J. Med. 1999, 341, 1485–1490. [Google Scholar] [CrossRef] [PubMed]
- Crider, K.S.; Bailey, L.B.; Berry, R.J. Folic acid food fortification-its history, effect, concerns, and future directions. Nutrients 2011, 3, 370–384. [Google Scholar] [CrossRef]
- MRC Vitamin Study Research Group. Prevention of neural tube defects: Results of the Medical Research Council Vitamin Study. Lancet 1991, 338, 131–137. [Google Scholar]
- Mosley, B.S.; Cleves, M.A.; Siega-Riz, A.M.; Shaw, G.M.; Canfield, M.A.; Waller, D.K.; Werler, M.M.; Hobbs, C.A. Neural tube defects and maternal folate intake among pregnancies conceived after folic acid fortification in the United States. Am. J. Epidemiol. 2009, 169, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Cavalli, P.; Copp, A.J. Inositol and folate resistant neural tube defects. J. Med. Genet. 2002, 39, E5. [Google Scholar] [CrossRef]
- Cavalli, P.; Tonni, G.; Grosso, E.; Poggiani, C. Effects of inositol supplementation in a cohort of mothers at risk of producing an NTD pregnancy. Birth Defects Res. A Clin. Mol. Teratol. 2011, 91, 962–965. [Google Scholar] [CrossRef]
- Greene, N.D.; Leung, K.Y.; Gay, V.; Burren, K.; Mills, K.; Chitty, L.S.; Copp, A.J. Inositol for the prevention of neural tube defects: A pilot randomised controlled trial. Br. J. Nutr. 2016, 115, 974–983. [Google Scholar] [CrossRef]
- Yeung, C.H.; Anapolski, M.; Setiawan, I.; Lang, F.; Cooper, T.G. Effects of putative epididymal osmolytes on sperm volume regulation of fertile and infertile c-ros transgenic Mice. J. Androl. 2004, 25, 216–223. [Google Scholar] [CrossRef]
- De Luca, M.N.; Colone, M.; Gambioli, R.; Stringaro, A.; Unfer, V. Oxidative Stress and Male Fertility: Role of Antioxidants and Inositols. Antioxidants 2021, 10, 1283. [Google Scholar] [CrossRef]
- Condorelli, R.A.; La Vignera, S.; Bellanca, S.; Vicari, E.; Calogero, A.E. Myoinositol: Does it improve sperm mitochondrial function and sperm motility? Urology 2012, 79, 1290–1295. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Heredia, J.; de Mateo, S.; Vidal-Taboada, J.M.; Ballescà, J.L.; Oliva, R. Identification of proteomic differences in asthenozoospermic sperm samples. Hum. Reprod. 2008, 23, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Colone, M.; Marelli, G.; Unfer, V.; Bozzuto, G.; Molinari, A.; Stringaro, A. Inositol activity in oligoasthenoteratospermia—an in vitro study. Eur. Rev. Med. Pharmacol. Sci. 2010, 14, 891–896. [Google Scholar]
- Condorelli, R.A.; Barbagallo, F.; Calogero, A.E.; Cannarella, R.; Crafa, A.; La Vignera, S. D-Chiro-Inositol Improves Sperm Mitochondrial Membrane Potential: In Vitro Evidence. J. Clin. Med. 2020, 9, 1373. [Google Scholar] [CrossRef]
- Artini, P.G.; Casarosa, E.; Carletti, E.; Monteleone, P.; Di Noia, A.; Di Berardino, O.M. In vitro effect of myo-inositol on sperm motility in normal and oligoasthenospermia patients undergoing in vitro fertilization. Gynecol. Endocrinol. 2017, 33, 109–112. [Google Scholar] [CrossRef]
- Rubino, P.; Palini, S.; Chigioni, S.; Carlomagno, G.; Quagliariello, A.; De Stefani, S.; Baglioni, A.; Bulletti, C. Improving fertilization rate in ICSI cycles by adding myoinositol to the semen preparation procedures: A prospective, bicentric, randomized trial on sibling oocytes. J. Assist Reprod. Genet. 2015, 32, 387–394. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gulino, F.A.; Leonardi, E.; Marilli, I.; Musmeci, G.; Vitale, S.G.; Leanza, V.; Palumbo, M.A. Effect of treatment with myo-inositol on semen parameters of patients undergoing an IVF cycle: In vivo study. Gynecol. Endocrinol. 2016, 32, 65–68. [Google Scholar] [CrossRef]
- Palmieri, M.; Papale, P.; Della Ragione, A.; Quaranta, G.; Russo, G.; Russo, S. In Vitro Antioxidant Treatment of Semen Samples in Assisted Reproductive Technology: Effects of Myo-Inositol on Nemaspermic Parameters. Int. J. Endocrinol. 2016, 2016, 2839041. [Google Scholar] [CrossRef]
- Calogero, A.E.; Gullo, G.; La Vignera, S.; Condorelli, R.A.; Vaiarelli, A. Myoinositol improves sperm parameters and serum reproductive hormones in patients with idiopathic infertility: A prospective double-blind randomized placebo-controlled study. Andrology 2015, 3, 491–495. [Google Scholar] [CrossRef]
- Montanino Oliva, M.; Minutolo, E.; Lippa, A.; Iaconianni, P.; Vaiarelli, A. Effect of Myoinositol and Antioxidants on Sperm Quality in Men with Metabolic Syndrome. Int. J. Endocrinol. 2016, 2016, 1674950. [Google Scholar] [CrossRef]
- Montanino Oliva, M.; Buonomo, G.; Carra, M.C.; Lippa, A.; Lisi, F. Myo-inositol impact on sperm motility in vagina and evaluation of its effects on foetal development. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 2704–2709. [Google Scholar] [CrossRef]
- Santoro, M.; Aquila, S.; Russo, G. Sperm performance in oligoasthenoteratozoospermic patients is induced by a nutraceuticals mix, containing mainly myo-inositol. Syst. Biol. Reprod. Med. 2021, 67, 50–63. [Google Scholar] [CrossRef]
- Watson, P.F. The causes of reduced fertility with cryopreserved semen. Anim. Reprod. Sci. 2000, 60–61, 481–492. [Google Scholar] [CrossRef]
- Mohammadi, F.; Varanloo, N.; Heydari Nasrabadi, M.; Vatannejad, A.; Amjadi, F.S.; Javedani Masroor, M.; Bajelan, L.; Mehdizadeh, M.; Aflatoonian, R.; Zandieh, Z. Supplementation of sperm freezing medium with myoinositol improve human sperm parameters and protects it against DNA fragmentation and apoptosis. Cell Tissue Bank 2019, 20, 77–86. [Google Scholar] [CrossRef]
- Stocco, C. Tissue physiology and pathology of aromatase. Steroids 2012, 77, 27–35. [Google Scholar] [CrossRef]
- Erickson, G.F.; Hsueh, A.J.; Quigley, M.E.; Rebar, R.W.; Yen, S.S. Functional studies of aromatase activity in human granulosa cells from normal and polycystic ovaries. J. Clin. Endocrinol. Metab. 1979, 49, 514–519. [Google Scholar] [CrossRef] [PubMed]
- Gambioli, R.; Forte, G.; Aragona, C.; Bevilacqua, A.; Bizzarri, M.; Unfer, V. The use of D-chiro-Inositol in clinical practice. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Monastra, G.; Vucenik, I.; Harrath, A.H.; Alwasel, S.H.; Kamenov, Z.A.; Laganà, A.S.; Monti, N.; Fedeli, V.; Bizzarri, M. PCOS and Inositols: Controversial Results and Necessary Clarifications. Basic Differences Between D-Chiro and Myo-Inositol. Front. Endocrinol. 2021, 12, 660381. [Google Scholar] [CrossRef] [PubMed]
- Bizzarri, M.; Cucina, A.; Dinicola, S.; Harrath, A.H.; Alwasel, S.H.; Unfer, V.; Bevilacqua, A. Does myo-inositol effect on PCOS follicles involve cytoskeleton regulation? Med. Hypotheses 2016, 91, 1–5. [Google Scholar] [CrossRef]
- Unfer, V.; Dinicola, S.; Laganà, A.S.; Bizzarri, M. Altered Ovarian Inositol Ratios May Account for Pathological Steroidogenesis in PCOS. Int. J. Mol. Sci. 2020, 21, 7157. [Google Scholar] [CrossRef]
- Chiu, T.T.; Tam, P.P. A correlation of the outcome of clinical in vitro fertilization with the inositol content and embryotrophic properties of human serum. J. Assist. Reprod. Genet. 1992, 9, 524–530. [Google Scholar] [CrossRef]
- Chiu, T.T.; Rogers, M.S.; Law, E.L.; Briton-Jones, C.M.; Cheung, L.P.; Haines, C.J. Follicular fluid and serum concentrations of myo-inositol in patients undergoing IVF: Relationship with oocyte quality. Hum. Reprod. 2002, 17, 1591–1596. [Google Scholar] [CrossRef] [PubMed]
- Wdowiak, A. Myoinositol Improves Embryo Development in PCOS Patients Undergoing ICSI. Int. J. Endocrinol. 2016, 2016, 6273298. [Google Scholar] [CrossRef] [PubMed]
- Vartanyan, E.V.; Tsaturova, K.A.; Devyatova, E.A.; Mikhaylyukova, A.S.; Levin, V.A.; Petuhova, N.L.; Markin, A.V.; Steptsova, E.M. Improvement in quality of oocytes in polycystic ovarian syndrome in programs of in vitro fertilization. Gynecol. Endocrinol. 2017, 33, 8–11. [Google Scholar] [CrossRef][Green Version]
- Özay, Ö.E.; Özay, A.C.; Çağlıyan, E.; Okyay, R.E.; Gülekli, B. Myo-inositol administration positively effects ovulation induction and intrauterine insemination in patients with polycystic ovary syndrome: A prospective, controlled, randomized trial. Gynecol. Endocrinol. 2017, 33, 524–528. [Google Scholar] [CrossRef]
- Laganà, A.S.; Vitagliano, A.; Noventa, M.; Ambrosini, G.; D’Anna, R. Myo-inositol supplementation reduces the amount of gonadotropins and length of ovarian stimulation in women undergoing IVF: A systematic review and meta-analysis of randomized controlled trials. Arch. Gynecol. Obstet. 2018, 298, 675–684. [Google Scholar] [CrossRef]
- Zheng, X.; Lin, D.; Zhang, Y.; Lin, Y.; Song, J.; Li, S.; Sun, Y. Inositol supplement improves clinical pregnancy rate in infertile women undergoing ovulation induction for ICSI or IVF-ET. Medicine 2017, 96, e8842. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, S.; Eini, F.; Bazarganipour, F.; Taghavi, S.A.; Kutenaee, M.A. The effect of Myo-inositol on fertility rates in poor ovarian responder in women undergoing assisted reproductive technique: A randomized clinical trial. Reprod. Biol. Endocrinol. 2021, 19, 61. [Google Scholar] [CrossRef]
- Laganà, A.S.; Garzon, S.; Unfer, V. New clinical targets of d-chiro-inositol: Rationale and potential applications. Expert Opin. Drug Metab. Toxicol. 2020, 16, 703–710. [Google Scholar] [CrossRef]
- Monastra, G.; Vazquez-Levin, M.; Bezerra Espinola, M.S.; Bilotta, G.; Laganà, A.S.; Unfer, V. D-chiro-inositol, an aromatase down-modulator, increases androgens and reduces estrogens in male volunteers: A pilot study. Basic Clin. Androl. 2021, 31, 13. [Google Scholar] [CrossRef]
- Tsametis, C.P.; Isidori, A.M. Testosterone replacement therapy: For whom, when and how? Metabolism 2018, 86, 69–78. [Google Scholar] [CrossRef]
- Bhasin, S.; Cunningham, G.R.; Hayes, F.J.; Matsumoto, A.M.; Snyder, P.J.; Swerdloff, R.S.; Montori, V.M. Testosterone therapy in men with androgen deficiency syndromes: An Endocrine Society clinical practice guideline. J. Clin. Endocrinol. Metab. 2010, 95, 2536–2559. [Google Scholar] [CrossRef]
- Sukegawa, G.; Tsuji, Y. Risk of Male Infertility Due to Testosterone Replacement Therapy for Late-Onset Hypogonadism (LOH). Hinyokika Kiyo 2020, 66, 407–409. [Google Scholar] [CrossRef]
- Tan, R.B.; Guay, A.T.; Hellstrom, W.J. Clinical Use of Aromatase Inhibitors in Adult Males. Sex Med. Rev. 2014, 2, 79–90. [Google Scholar] [CrossRef]
- Carlomagno, G.; Unfer, V. Inositol safety: Clinical evidences. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 931–936. [Google Scholar]
- Nordio, M.K.P.; Chiefari, A.; Puliani, G. D-Chiro-inositol improves testosterone levels in hypogonadal males: A pilot study. Basic Clin. Androl. 2021. [Google Scholar] [CrossRef]
- Hess, R.A.; Cooke, P.S. Estrogen in the male: A historical perspective. Biol. Reprod. 2018, 99, 27–44. [Google Scholar] [CrossRef] [PubMed]
- Schneider, G.; Kirschner, M.A.; Berkowitz, R.; Ertel, N.H. Increased estrogen production in obese men. J. Clin. Endocrinol. Metab. 1979, 48, 633–638. [Google Scholar] [CrossRef]
- MacDonald, A.A.; Herbison, G.P.; Showell, M.; Farquhar, C.M. The impact of body mass index on semen parameters and reproductive hormones in human males: A systematic review with meta-analysis. Hum. Reprod. Update 2010, 16, 293–311. [Google Scholar] [CrossRef]
- Hammoud, A.; Carrell, D.T.; Meikle, A.W.; Xin, Y.; Hunt, S.C.; Adams, T.D.; Gibson, M. An aromatase polymorphism modulates the relationship between weight and estradiol levels in obese men. Fertil. Steril. 2010, 94, 1734–1738. [Google Scholar] [CrossRef]
- Mah, P.M.; Wittert, G.A. Obesity and testicular function. Mol. Cell Endocrinol. 2010, 316, 180–186. [Google Scholar] [CrossRef]
- Tinelli, A.; Gustapane, S.; D’Oria, O.; Licchelli, M.; Panese, G. Nutraceuticals in fibroid management after ulipristal acetate administration: An observational study on patients’ compliance. Int. J. Gynaecol. Obstet. 2021. [Google Scholar] [CrossRef]
- Stewart, E.A.; Laughlin-Tommaso, S.K.; Catherino, W.H.; Lalitkumar, S.; Gupta, D.; Vollenhoven, B. Uterine fibroids. Nat. Rev. Dis. Primers 2016, 2, 16043. [Google Scholar] [CrossRef]
- Meunier, L.; Meszaros, M.; Pageaux, G.P.; Delay, J.M.; Herrero, A.; Pinzani, V.; Dominique, H.B. Acute liver failure requiring transplantation caused by ulipristal acetate. Clin. Res. Hepatol. Gastroenterol. 2020, 44, e45–e49. [Google Scholar] [CrossRef]
- Brodie, A.; Lu, Q.; Nakamura, J. Aromatase in the normal breast and breast cancer. J. Steroid Biochem. Mol. Biol. 1997, 61, 281–286. [Google Scholar] [CrossRef]
- Imir, A.G.; Lin, Z.; Yin, P.; Deb, S.; Yilmaz, B.; Cetin, M.; Cetin, A.; Bulun, S.E. Aromatase expression in uterine leiomyomata is regulated primarily by proximal promoters I.3/II. J. Clin. Endocrinol. Metab. 2007, 92, 1979–1982. [Google Scholar] [CrossRef]
- Ishikawa, H.; Reierstad, S.; Demura, M.; Rademaker, A.W.; Kasai, T.; Inoue, M.; Usui, H.; Shozu, M.; Bulun, S.E. High aromatase expression in uterine leiomyoma tissues of African-American women. J. Clin. Endocrinol. Metab. 2009, 94, 1752–1756. [Google Scholar] [CrossRef]
- Sumitani, H.; Shozu, M.; Segawa, T.; Murakami, K.; Yang, H.J.; Shimada, K.; Inoue, M. In situ estrogen synthesized by aromatase P450 in uterine leiomyoma cells promotes cell growth probably via an autocrine/intracrine mechanism. Endocrinology 2000, 141, 3852–3861. [Google Scholar] [CrossRef]
- Al-Hendy, A.; Diamond, M.P.; El-Sohemy, A.; Halder, S.K. 1,25-dihydroxyvitamin D3 regulates expression of sex steroid receptors in human uterine fibroid cells. J. Clin. Endocrinol. Metab. 2015, 100, E572–E582. [Google Scholar] [CrossRef]
- Sinai Talaulikar, V. Medical therapy for fibroids: An overview. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 46, 48–56. [Google Scholar] [CrossRef]
- Montanino Oliva, M. D-Chiro-Inositol, vitamin D and epigallocatechin gallate avoid surgery in women with uterine fibroids: 2 case reports. Eur. Med. J. 2021, 9, 95–101. [Google Scholar]
- Roshdy, E.; Rajaratnam, V.; Maitra, S.; Sabry, M.; Allah, A.S.; Al-Hendy, A. Treatment of symptomatic uterine fibroids with green tea extract: A pilot randomized controlled clinical study. Int. J. Womens Health 2013, 5, 477–486. [Google Scholar] [CrossRef]
- Ciavattini, A.; Delli Carpini, G.; Serri, M.; Vignini, A.; Sabbatinelli, J.; Tozzi, A.; Aggiusti, A.; Clemente, N. Hypovitaminosis D and “small burden” uterine fibroids: Opportunity for a vitamin D supplementation. Medicine 2016, 95, e5698. [Google Scholar] [CrossRef]
- Miriello, D.; Galanti, F.; Cignini, P.; Antonaci, D.; Schiavi, M.C.; Rago, R. Uterine fibroids treatment: Do we have new valid alternative? Experiencing the combination of vitamin D plus epigallocatechin gallate in childbearing age affected women. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 2843–2851. [Google Scholar] [CrossRef]
- Shaw, N.D.; Histed, S.N.; Srouji, S.S.; Yang, J.; Lee, H.; Hall, J.E. Estrogen negative feedback on gonadotropin secretion: Evidence for a direct pituitary effect in women. J. Clin. Endocrinol. Metab. 2010, 95, 1955–1961. [Google Scholar] [CrossRef]
- Tulandi, T.; Martin, J.; Al-Fadhli, R.; Kabli, N.; Forman, R.; Hitkari, J.; Librach, C.; Greenblatt, E.; Casper, R.F. Congenital malformations among 911 newborns conceived after infertility treatment with letrozole or clomiphene citrate. Fertil. Steril. 2006, 85, 1761–1765. [Google Scholar] [CrossRef]
- Bezerra Espinola, M.S.; Laganà, A.S.; Bilotta, G.; Gullo, G.; Aragona, C.; Unfer, V. D-chiro-inositol induces ovulation in non-PCOS, non-insulin-resistant young women likely by modulating aromatase expression: Two case reports. Am. J. Case Rep. 2021, 22, e932722. [Google Scholar] [CrossRef]
- Nair, A.; Morsy, M.A.; Jacob, S. Dose translation between laboratory animals and human in preclinical and clinical phases of drug development. Drug Dev. Res. 2018, 79, 373–382. [Google Scholar] [CrossRef]
- Carlomagno, G.; De Grazia, S.; Unfer, V.; Manna, F. Myo-inositol in a new pharmaceutical form: A step forward to a broader clinical use. Expert Opin. Drug Deliv. 2012, 9, 267–271. [Google Scholar] [CrossRef]
- Kamenov, Z.; Gateva, A. Inositols in PCOS. Molecules 2020, 25, 5566. [Google Scholar] [CrossRef]
- Gerli, S.; Papaleo, E.; Ferrari, A.; Di Renzo, G.C. Randomized, double blind placebo-controlled trial: Effects of myo-inositol on ovarian function and metabolic factors in women with PCOS. Eur. Rev. Med. Pharmacol. Sci. 2007, 11, 347–354. [Google Scholar]
- Kamenov, Z.; Kolarov, G.; Gateva, A.; Carlomagno, G.; Genazzani, A.D. Ovulation induction with myo-inositol alone and in combination with clomiphene citrate in polycystic ovarian syndrome patients with insulin resistance. Gynecol. Endocrinol. 2015, 31, 131–135. [Google Scholar] [CrossRef]
- Vita, R.; Fallahi, P.; Antonelli, A.; Benvenga, S. The administration of L-thyroxine as soft gel capsule or liquid solution. Expert Opin. Drug Deliv. 2014, 11, 1103–1111. [Google Scholar] [CrossRef]
- Fallahi, P.; Ferrari, S.M.; Ruffilli, I.; Ragusa, F.; Biricotti, M.; Materazzi, G.; Miccoli, P.; Antonelli, A. Advancements in the treatment of hypothyroidism with L-T4 liquid formulation or soft gel capsule: An update. Expert Opin. Drug Deliv. 2017, 14, 647–655. [Google Scholar] [CrossRef]
- Figgitt, D.P.; Plosker, G.L. Saquinavir soft-gel capsule: An updated review of its use in the management of HIV infection. Drugs 2000, 60, 481–516. [Google Scholar] [CrossRef]
- Proietti, S.; Carlomagno, G.; Dinicola, S.; Bizzarri, M. Soft gel capsules improve melatonin’s bioavailability in humans. Expert Opin. Drug Metab. Toxicol. 2014, 10, 1193–1198. [Google Scholar] [CrossRef]
- Wang, X.; Ai, T.; Meng, X.L.; Zhou, J.; Mao, X.Y. In vitro iron absorption of α-lactalbumin hydrolysate-iron and β-lactoglobulin hydrolysate-iron complexes. J. Dairy Sci. 2014, 97, 2559–2566. [Google Scholar] [CrossRef]
- Delavari, B.; Saboury, A.A.; Atri, M.S.; Ghasemi, A.; Bigdeli, B.; Khammari, A.; Maghami, P.; Moosavi-Movahedi, A.A.; Haertlé, T.; Goliaei, B. Alpha-lactalbumin: A new carrier for vitamin D3 food enrichment. Food Hydrocoll. 2015, 45, 124–131. [Google Scholar] [CrossRef]
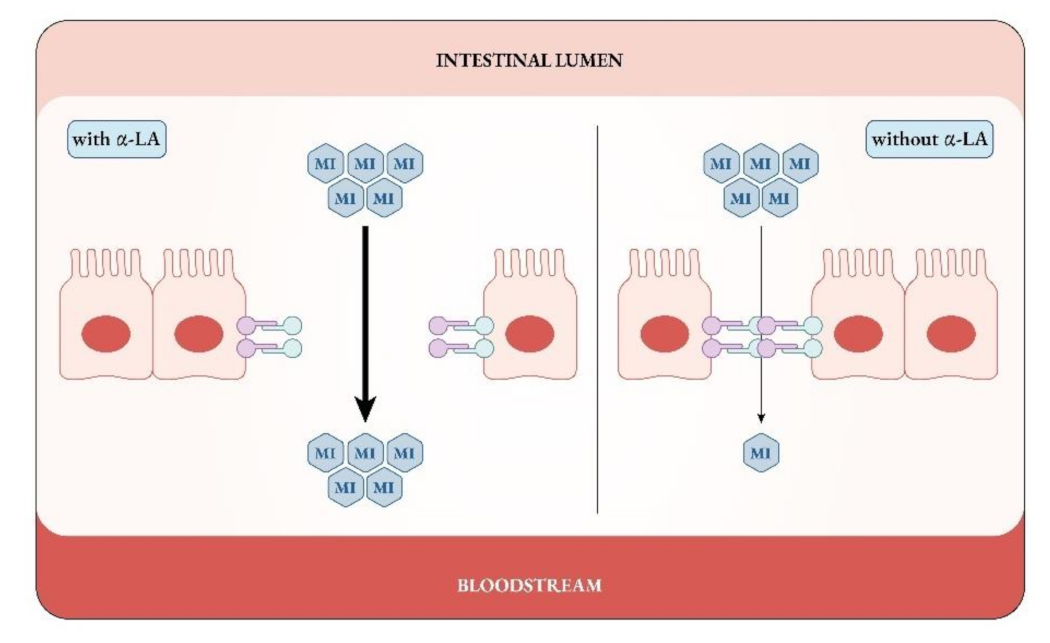
- Monastra, G.; Sambuy, Y.; Ferruzza, S.; Ferrari, D.; Ranaldi, G. Alpha-lactalbumin Effect on Myo-inositol Intestinal Absorption: In vivo and In vitro. Curr. Drug Deliv. 2018, 15, 1305–1311. [Google Scholar] [CrossRef]
- Montanino Oliva, M.; Buonomo, G.; Calcagno, M.; Unfer, V. Effects of myo-inositol plus alpha-lactalbumin in myo-inositol-resistant PCOS women. J. Ovarian Res. 2018, 11, 38. [Google Scholar] [CrossRef]
- Sambuy, Y.; De Angelis, I.; Ranaldi, G.; Scarino, M.L.; Stammati, A.; Zucco, F. The Caco-2 cell line as a model of the intestinal barrier: Influence of cell and culture-related factors on Caco-2 cell functional characteristics. Cell Biol. Toxicol. 2005, 21, 1–26. [Google Scholar] [CrossRef]
- Ranaldi, G.; Ferruzza, S.; Natella, F.; Unfer, V.; Sambuy, Y.; Monastra, G. Enhancement of D-chiro-inositol transport across intestinal cells by alpha-Lactalbumin peptides. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 10143–10154. [Google Scholar] [CrossRef]
- Hernandez Marin, I.; Picconi, O.; Laganà, A.S.; Costabile, L.; Unfer, V. A multicenter clinical study with myo-inositol and alpha-lactalbumin in Mexican and Italian PCOS patients. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 3316–3324. [Google Scholar] [CrossRef]
- Leung, K.Y.; Mills, K.; Burren, K.A.; Copp, A.J.; Greene, N.D. Quantitative analysis of myo-inositol in urine, blood and nutritional supplements by high-performance liquid chromatography tandem mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2011, 879, 2759–2763. [Google Scholar] [CrossRef]
- Ostlund, R.E., Jr.; McGill, J.B.; Herskowitz, I.; Kipnis, D.M.; Santiago, J.V.; Sherman, W.R. D-chiro-inositol metabolism in diabetes mellitus. Proc. Natl. Acad. Sci. USA 1993, 90, 9988–9992. [Google Scholar] [CrossRef]
- Deranieh, R.M.; Greenberg, M.L. Cellular consequences of inositol depletion. Biochem. Soc. Trans. 2009, 37, 1099–1103. [Google Scholar] [CrossRef]
- Murru, A.; Popovic, D.; Pacchiarotti, I.; Hidalgo, D.; León-Caballero, J.; Vieta, E. Management of adverse effects of mood stabilizers. Curr. Psychiatry Rep. 2015, 17, 603. [Google Scholar] [CrossRef]
- Gitlin, M. Lithium side effects and toxicity: Prevalence and management strategies. Int. J. Bipolar Disord. 2016, 4, 27. [Google Scholar] [CrossRef]
- Lepore, E.; Lauretta, R.; Bianchini, M.; Mormando, M.; Di Lorenzo, C.; Unfer, V. Inositols Depletion and Resistance: Principal Mechanisms and Therapeutic Strategies. Int. J. Mol. Sci. 2021, 22, 6796. [Google Scholar] [CrossRef]
- Genton, P.; Bauer, J.; Duncan, S.; Taylor, A.E.; Balen, A.H.; Eberle, A.; Pedersen, B.; Salas-Puig, X.; Sauer, M.V. On the association between valproate and polycystic ovary syndrome. Epilepsia 2001, 42, 295–304. [Google Scholar] [CrossRef]
- Bilo, L.; Meo, R. Polycystic ovary syndrome in women using valproate: A review. Gynecol. Endocrinol. 2008, 24, 562–570. [Google Scholar] [CrossRef]
- Hu, X.; Wang, J.; Dong, W.; Fang, Q.; Hu, L.; Liu, C. A meta-analysis of polycystic ovary syndrome in women taking valproate for epilepsy. Epilepsy Res. 2011, 97, 73–82. [Google Scholar] [CrossRef]
- Viswanathan, L.G.; Satishchandra, P.; Bhimani, B.C.; Reddy, J.Y.; Rama Murthy, B.S.; Subbakrishna, D.K.; Sinha, S. Polycystic ovary syndrome in patients on antiepileptic drugs. Ann. Indian Acad. Neurol. 2016, 19, 339–343. [Google Scholar] [CrossRef]
- Cantelmi, T.; Lambiase, E.; Unfer, V.R.; Gambioli, R.; Unfer, V. Inositol treatment for psychological symptoms in Polycystic Ovary Syndrome women. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 2383–2389. [Google Scholar] [CrossRef]
- Nordio, M.; Basciani, S. Treatment with Myo-Inositol and Selenium Ensures Euthyroidism in Patients with Autoimmune Thyroiditis. Int. J. Endocrinol. 2017, 2017, 2549491. [Google Scholar] [CrossRef]
- Allan, S.J.; Kavanagh, G.M.; Herd, R.M.; Savin, J.A. The effect of inositol supplements on the psoriasis of patients taking lithium: A randomized, placebo-controlled trial. Br. J. Dermatol. 2004, 150, 966–969. [Google Scholar] [CrossRef]
- Kontoangelos, K.; Vaidakis, N.; Zervas, I.; Thomadaki, O.; Christaki, S.; Stavrianeas, N.G.; Papadimitriou, G.N. Administration of inositol to a patient with bipolar disorder and psoriasis: A case report. Cases J. 2010, 3, 69. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physiological Conditions | Insulin Resistance Conditions | |||
|---|---|---|---|---|
| MI (%) | DCI (%) | MI (%) | DCI (%) | |
| Fat | 65 | 35 | 98 | 2 |
| Liver | 70 | 30 | 99.3 | 0.7 |
| Muscle | 74 | 26 | 98.1 | 1.9 |
| Blood | 97 | 3 | 99.6 | 0.4 |
| Kidney | 98 | 2 | 98.2 | 1.8 |
| Intestine | 98 | 2 | 98.2 | 1.8 |
| Spleen | 98.8 | 1.2 | 99 | 1 |
| Heart | 99.5 | 0.5 | 99.3 | 0.7 |
| Brain | 99.5 | 0.5 | 99.2 | 0.8 |
| Follicular fluid | 99.02 | 0.98 | 15 | 85 |
| Ovary (Theca) | 95.24 | 4.76 | 16.67 | 83.33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dinicola, S.; Unfer, V.; Facchinetti, F.; Soulage, C.O.; Greene, N.D.; Bizzarri, M.; Laganà, A.S.; Chan, S.-Y.; Bevilacqua, A.; Pkhaladze, L.; et al. Inositols: From Established Knowledge to Novel Approaches. Int. J. Mol. Sci. 2021, 22, 10575. https://doi.org/10.3390/ijms221910575
Dinicola S, Unfer V, Facchinetti F, Soulage CO, Greene ND, Bizzarri M, Laganà AS, Chan S-Y, Bevilacqua A, Pkhaladze L, et al. Inositols: From Established Knowledge to Novel Approaches. International Journal of Molecular Sciences. 2021; 22(19):10575. https://doi.org/10.3390/ijms221910575
Chicago/Turabian StyleDinicola, Simona, Vittorio Unfer, Fabio Facchinetti, Christophe O. Soulage, Nicholas D. Greene, Mariano Bizzarri, Antonio Simone Laganà, Shiao-Yng Chan, Arturo Bevilacqua, Lali Pkhaladze, and et al. 2021. "Inositols: From Established Knowledge to Novel Approaches" International Journal of Molecular Sciences 22, no. 19: 10575. https://doi.org/10.3390/ijms221910575
APA StyleDinicola, S., Unfer, V., Facchinetti, F., Soulage, C. O., Greene, N. D., Bizzarri, M., Laganà, A. S., Chan, S.-Y., Bevilacqua, A., Pkhaladze, L., Benvenga, S., Stringaro, A., Barbaro, D., Appetecchia, M., Aragona, C., Bezerra Espinola, M. S., Cantelmi, T., Cavalli, P., Chiu, T. T., ... Wdowiak, A. (2021). Inositols: From Established Knowledge to Novel Approaches. International Journal of Molecular Sciences, 22(19), 10575. https://doi.org/10.3390/ijms221910575