
Treatment of Sjögren’s Syndrome with Mesenchymal Stem Cells: A Systematic Review

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Research Strategy
2.2. Selection of Articles
2.3. Information Extraction
3. Results and Discussion
3.1. Search Results
Study Characteristics
- Umbilical MSCs, umbilical cord MSCs (UMSCs, UCMSCs);
- Bone MSCs, bone marrow MSCs (BMSCs, BMMSCs);
- Dental pulp stem cells (DPSCs);
- Stem cells from exfoliated deciduous teeth (SHED);
- Murine embryonic MSCs (ME-MSCs);
- Olfactory ecto-MSCs, olfactory ecto-MSC-derived exosomes (OE-MSCs, OE-MSCs-Exos);
- Induced pluripotent stem cells (iPSC-MSCs);
- Salivary gland MSCs (SGMSCs).
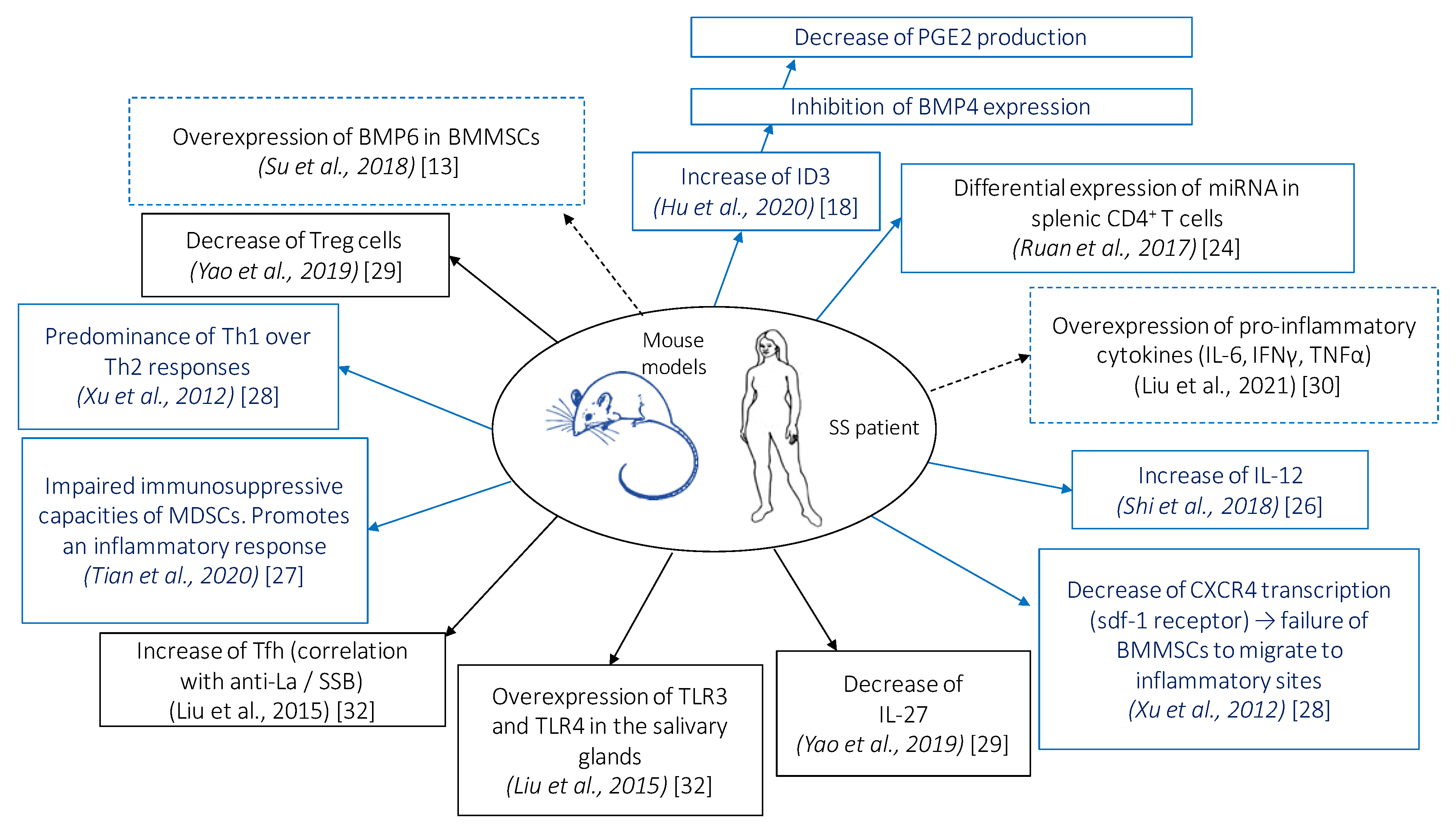
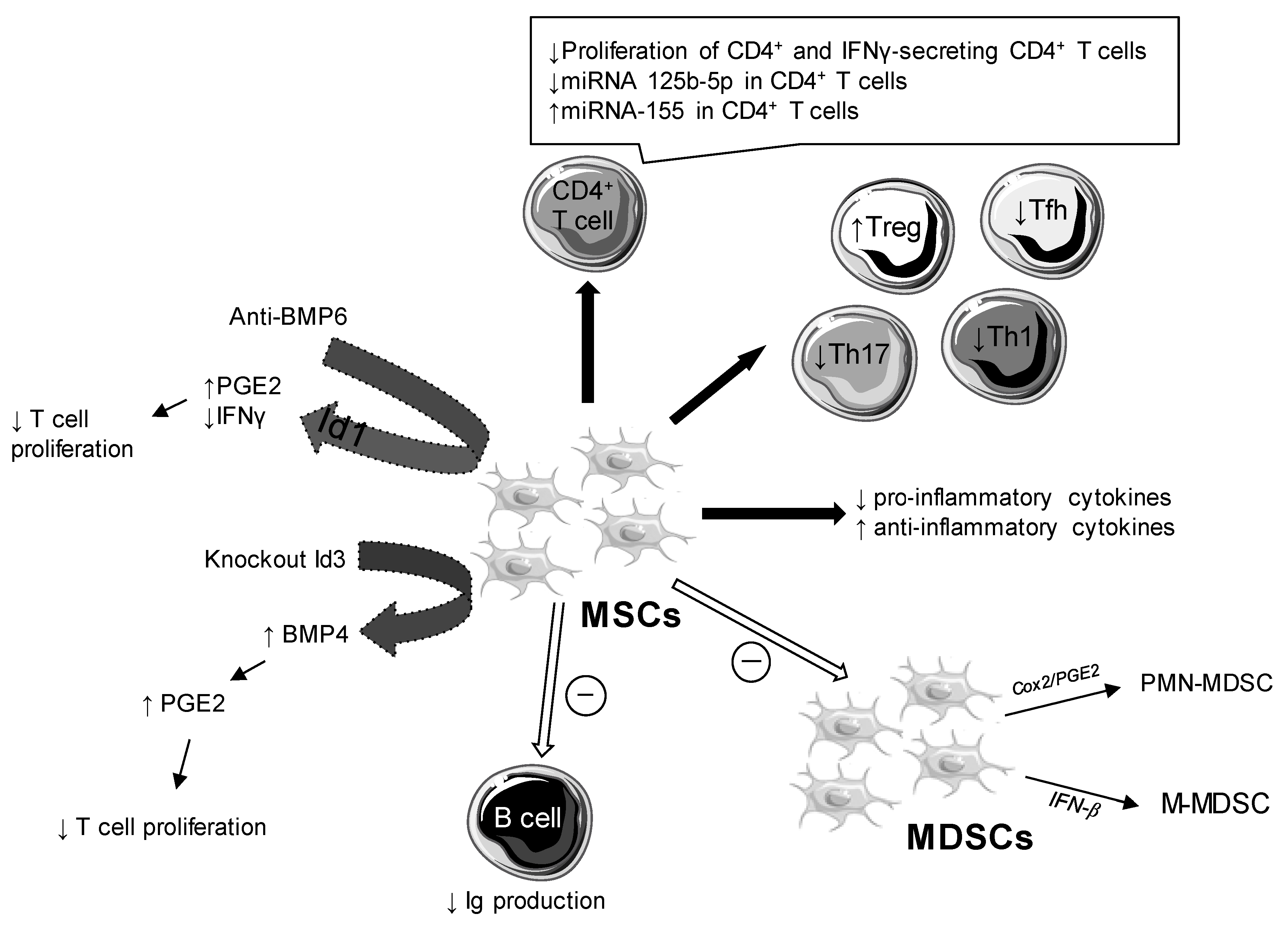
3.2. Cellular and Molecular Changes
3.2.1. Role of Bone Morphogenic Proteins
3.2.2. Impact on Pro- and Anti-Inflammatory Cytokines
3.2.3. Role of IL-12
3.2.4. Other Cytokines
3.2.5. Impact on Apoptotic Cells
3.2.6. Impact on Treg, Th1, Th2, Th17, and Tfh Cell Responses
3.2.7. Impact on B Cell Response
3.2.8. Anti-SSA/Ro and Anti-SSB/La Antibodies
3.3. Impact on Salivary Flow
3.4. Follow Up and Risk of Transplant
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Le Gall, M.; Cornec, D.; Pers, J.O.; Saraux, A.; Jousse-Joulin, S.; Cochener, B.; Roguedas-Contios, A.M.; Devauchelle-Pensec, V.; Boisramé, S. A prospective evaluation of dental and periodontal status in patients with suspected Sjögren’s syndrome. Jt. Bone Spine 2016, 83, 235–236. [Google Scholar] [CrossRef] [PubMed]
- Baer, A.N.; Walitt, B. Update on Sjögren Syndrome and Other Causes of Sicca in Older Adults. Rheum. Dis. Clin. N. Am. 2018, 44, 419–436. [Google Scholar] [CrossRef] [PubMed]
- Kittridge, A.; Routhouska, S.B.; Korman, N.J. Dermatologic Manifestations of Sjögren Syndrome. J. Cutan. Med. Surg. 2011, 15, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Mielle, J.; Tison, A.; Cornec, D.; Le Pottier, L.; Daien, C.; Pers, J.O. B cells in Sjögren’s syndrome: From pathophysiology to therapeutic target. Rheumatology 2021, 60, 2545–2560. [Google Scholar] [CrossRef]
- Maehara, T.; Moriyama, M.; Hayashida, J.N.; Tanaka, A.; Shinozaki, S.; Kubo, Y.; Matsumura, S. Selective localization of T helper subsets in labial salivary glands from primary Sjögren’s syndrome patients. Clin. Exp. Immunol. 2012, 169, 89–99. [Google Scholar] [CrossRef]
- Depinoy, T.; Saraux, A.; Pers, J.O.; Boisramé, S.; Cornec, D.; Marhadour, T.; Guellec, D.; Devauchelle-Pensec, V.; Bressollette, B.; Jousse-Joulin, S. Salivary Glands and Periodontal Changes in a Population of Sjögren’s and Sicca Syndrome Treated by Pilocarpine: A Pilot Study. Rheumatol. Ther. 2021, 8, 219–231. [Google Scholar] [CrossRef]
- Chen, W.; Yu, Y.; Ma, J.; Olsen, N.; Lin, J. Mesenchymal Stem Cells in Primary Sjögren’s Syndrome: Prospective and Challenges. Stem Cells Int. 2018, 2018, 4357865. [Google Scholar] [CrossRef]
- Jensen, D.H.; Oliveri, R.S.; Kølle, S.-F.T.; Fischer-Nielsen, A.; Specht, L.; Bardow, A.; von Buchwald, C. Mesenchymal stem cell therapy for salivary gland dysfunction and xerostomia: A systematic review of preclinical studies. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2014, 117, 335–342.e1. [Google Scholar] [CrossRef]
- Srivastava, A.; Makarenkova, H.P. Innate Immunity and Biological Therapies for the Treatment of Sjögren’s Syndrome. Int. J. Mol. Sci. 2020, 21, 9172. [Google Scholar] [CrossRef]
- Peltzer, J.; Aletti, M.; Frescaline, N.; Busson, E.; Lataillade, J.-J.; Martinaud, C. Mesenchymal Stromal Cells Based Therapy in Systemic Sclerosis: Rational and Challenges. Front. Immunol. 2018, 9, 2013. [Google Scholar] [CrossRef]
- Maria, A.T.; Maumus, M.; Le Quellec, A.; Jorgensen, C.; Noël, D.; Guilpain, P. Adipose-Derived Mesenchymal Stem Cells in Autoimmune Disorders: State of the Art and Perspectives for Systemic Sclerosis. Clin. Rev. Allergy Immunol. 2017, 52, 234–259. [Google Scholar] [CrossRef]
- Xu, J.; Su, Y.; Hu, L.; Cain, A.; Gu, Y.; Liu, B.; Wu, R.; Wang, S.; Wang, H. Effect of Bone Morphogenetic Protein 6 on Immunomodulatory Functions of Salivary Gland-Derived Mesenchymal Stem Cells in Sjögren’s Syndrome. Stem Cells Dev. 2018, 27, 1540–1548. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Gu, Y.; Wu, R.; Wang, H. Bone Morphogenetic Protein 6 Inhibits the Immunomodulatory Property of BMMSCs via Id1 in Sjögren’s Syndrome. Stem Cells Int. 2018, 2018, 9837035. [Google Scholar] [CrossRef] [PubMed]
- Abughanam, G.; Elkashty, O.A.; Liu, Y.; Bakkar, M.O.; Tran, S.D. Mesenchymal Stem Cells Extract (MSCsE)-Based Therapy Alleviates Xerostomia and Keratoconjunctivitis Sicca in Sjogren’s Syndrome-Like Disease. Int. J. Mol. Sci. 2021, 22, 894. [Google Scholar] [CrossRef]
- Du, Z.H.; Ding, C.; Zhang, Q.; Zhang, Y.; Ge, X.Y.; Li, S.L.; Yu, G.Y. Stem cells from exfoliated deciduous teeth alleviate hyposalivation caused by Sjögren syndrome. Oral Dis. 2019, 25, 1530–1544. [Google Scholar] [CrossRef] [PubMed]
- Gong, B.; Zheng, L.; Huang, W.; Pu, J.; Pan, S.; Liang, Y.; Wu, Z.; Tang, J. Murine embryonic mesenchymal stem cells attenuated xerostomia in Sjögren-like mice via improving salivary gland epithelial cell structure and secretory function. Int. J. Clin. Exp. Pathol. 2020, 13, 954–963. [Google Scholar] [PubMed]
- Hai, B.; Shigemoto-Kuroda, T.; Zhao, Q.; Lee, R.H.; Liu, F. Inhibitory Effects of iPSC-MSCs and Their Extracellular Vesicles on the Onset of Sialadenitis in a Mouse Model of Sjögren’s Syndrome. Stem Cells Int. 2018, 2018, 2092315. [Google Scholar] [CrossRef]
- Hu, L.; Xu, J.; Wu, T.; Fan, Z.; Sun, L.; Liu, Y.; Li, Y.; Zhang, C.; Wang, J.; Ding, Y.; et al. Depletion of ID3 enhances mesenchymal stem cells therapy by targeting BMP4 in Sjögren’s syndrome. Cell Death Dis. 2020, 11, 172. [Google Scholar] [CrossRef]
- Khalili, S.; Liu, Y.; Kornete, M.; Roescher, N.; Kodama, S.; Peterson, A.; Piccirillo, C.A.; Tran, S.D. Mesenchymal stromal cells improve salivary function and reduce lymphocytic infiltrates in mice with Sjögren’s-like disease. PLoS ONE 2012, 7, e38615. [Google Scholar] [CrossRef]
- Khalili, S.; Liu, Y.; Sumita, Y.; Maria, O.M.; Blank, D.; Key, S.; Mezey, E.; Tran, S.D. Bone marrow cells are a source of undifferentiated cells to prevent Sjögren’s syndrome and to preserve salivary glands function in the non-obese diabetic mice. Int. J. Biochem. Cell Biol. 2010, 42, 1893–1899. [Google Scholar] [CrossRef][Green Version]
- Matsumura-Kawashima, M.; Ogata, K.; Moriyama, M.; Murakami, Y.; Kawado, T.; Nakamura, S. Secreted factors from dental pulp stem cells improve Sjögren’s syndrome via regulatory T cell-mediated immunosuppression. Stem Cell Res. Ther. 2021, 12, 182. [Google Scholar] [CrossRef] [PubMed]
- Ogata, K.; Matsumura-Kawashima, M.; Moriyama, M.; Kawado, T.; Nakamura, S. Dental pulp-derived stem cell-conditioned media attenuates secondary Sjögren’s syndrome via suppression of inflammatory cytokines in the submandibular glands. Regen. Ther. 2021, 16, 73–80. [Google Scholar] [CrossRef]
- Qi, J.; Tang, X.; Li, W.; Chen, W.; Yao, G.; Sun, L. Mesenchymal stem cells inhibited the differentiation of MDSCs via COX2/PGE2 in experimental sialadenitis. Stem Cell Res. Ther. 2020, 11, 325. [Google Scholar] [CrossRef] [PubMed]
- Ruan, G.; Zheng, L.; Huang, J.; Huang, W.-X.; Gong, B.; Fang, X.; Zhang, X.; Tang, J. Effect of mesenchymal stem cells on Sjögren-like mice and the microRNA expression profiles of splenic CD4+ T cells. Exp. Ther. Med. 2017, 13, 2828–2838. [Google Scholar] [CrossRef]
- Rui, K.; Hong, Y.; Zhu, Q.; Shi, X.; Xiao, F.; Fu, H.; Yin, Q.; Xing, Y.; Wu, X.; Kong, X.; et al. Olfactory ecto-mesenchymal stem cell-derived exosomes ameliorate murine Sjögren’s syndrome by modulating the function of myeloid-derived suppressor cells. Cell. Mol. Immunol. 2021, 18, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Shi, B.; Qi, J.; Yao, G.; Feng, R.; Zhang, Z.; Wang, D.; Chen, C.; Tang, X.; Lu, L.; Chen, W.; et al. Mesenchymal stem cell transplantation ameliorates Sjögren’s syndrome via suppressing IL-12 production by dendritic cells. Stem Cell Res. Ther. 2018, 9, 308. [Google Scholar] [CrossRef]
- Tian, J.; Hong, Y.; Zhu, Q.; Zhou, H.; Zhang, Y.; Shen, Z.; Guo, H.; Zhang, Y.; Ai, X.; Zhao, F.; et al. Mesenchymal Stem Cell Enhances the Function of MDSCs in Experimental Sjögren Syndrome. Front. Immunol. 2020, 11, 604607. [Google Scholar] [CrossRef]
- Xu, J.; Wang, D.; Liu, D.; Fan, Z.; Zhang, H.; Liu, O.; Ding, G.; Gao, R.; Zhang, C.; Ding, Y.; et al. Allogeneic mesenchymal stem cell treatment alleviates experimental and clinical Sjögren syndrome. Blood 2012, 120, 3142–3151. [Google Scholar] [CrossRef]
- Yao, G.; Qi, J.; Liang, J.; Shi, B.; Chen, W.; Li, W.; Chen, W.; Hou, Y.; Sun, L. Mesenchymal stem cell transplantation alleviates experimental Sjögren’s syndrome through IFN-β/IL-27 signaling axis. Theranostics 2019, 9, 8253–8265. [Google Scholar] [CrossRef]
- Liu, Y.; Li, C.; Wang, S.; Guo, J.; Guo, J.; Fu, J.; Ren, L.; An, Y.; He, J.; Li, Z. Human umbilical cord mesenchymal stem cells confer potent immunosuppressive effects in Sjögren’s syndrome by inducing regulatory T cells. Mod. Rheumatol. 2021, 31, 186–196. [Google Scholar] [CrossRef]
- Alunno, A.; Montanucci, P.; Bistoni, O.; Basta, G.; Caterbi, S.; Pescara, T.; Pennoni, I.; Bini, V.; Bartoloni, E.; Gerli, R.; et al. In vitro immunomodulatory effects of microencapsulated umbilical cord Wharton jelly-derived mesenchymal stem cells in primary Sjögren’s syndrome. Rheumatology 2015, 54, 163–168. [Google Scholar] [CrossRef]
- Liu, R.; Su, D.; Zhou, M.; Feng, X.; Li, X.; Sun, L. Umbilical cord mesenchymal stem cells inhibit the differentiation of circulating T follicular helper cells in patients with primary Sjögren’s syndrome through the secretion of indoleamine 2,3-dioxygenase. Rheumatology 2015, 54, 332–342. [Google Scholar] [CrossRef]
- Gong, B.; Zheng, L.; Lu, Z.; Huang, J.; Pu, J.; Pan, S.; Zhang, M.; Liu, J.; Tang, J. Mesenchymal stem cells negatively regulate CD4+ T cell activation in patients with primary Sjögren syndrome through the miRNA-125b and miRNA-155 TCR pathway. Mol. Med. Rep. 2021, 23, 43. [Google Scholar] [PubMed]
- Youinou, P.; Pers, J.O. Disturbance of cytokine networks in Sjögren’s syndrome. Arthritis Res. Ther. 2011, 13, 227. [Google Scholar] [CrossRef]
- Qi, J.; Li, D.; Shi, G.; Zhang, X.; Pan, Y.; Dou, H.; Wang, T.; Yao, J.; Hou, Y. Interleukin-12 exacerbates Sjögren’s syndrome through induction of myeloid-derived suppressor cells. Mol. Med. Rep. 2019, 20, 1131–1138. [Google Scholar]
- Szodoray, P.; Alex, P.; Brun, J.G.; Centola, M.; Jonsson, R. Circulating cytokines in primary Sjögren’s syndrome determined by a multiplex cytokine array system. Scand. J. Immunol. 2004, 59, 592–599. [Google Scholar] [CrossRef]
- Teng, M.W.L.; Bowman, E.P.; McElwee, J.J.; Smyth, M.; Casanova, J.-L.; Cooper, A.; Cua, D.J. IL-12 and IL-23 cytokines: From discovery to targeted therapies for immune-mediated inflammatory diseases. Nat. Med. 2015, 21, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Pers, J.O.; Lahiri, A.; Tobón, G.J.; Youinou, P. Pathophysiological cytokine network in primary Sjögren’s syndrome. Presse Medicale 2012, 41 Pt 2, e467–e474. [Google Scholar] [CrossRef]
- Shiboski, C.H.; Shiboski, S.C.; Seror, R.; Criswell, L.A.; Labetoulle, M.; Lietman, T.M.; Rasmussen, A.; Scofield, H.; Vitali, V.; Bowman, S.J.; et al. 2016 American College of Rheumatology/European League Against Rheumatism Classification Criteria for Primary Sjögren’s Syndrome: A Consensus and Data-Driven Methodology Involving Three International Patient Cohorts. Arthritis Rheumatol. 2017, 69, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Tran, S.D.; Sumita, Y.; Khalili, S. Bone marrow-derived cells: A potential approach for the treatment of xerostomia. Int. J. Biochem. Cell Biol. 2011, 43, 5–9. [Google Scholar] [CrossRef]
- Liang, J.; Zhang, H.; Kong, W.; Deng, W.; Wang, D.; Feng, X.; Zhao, C.; Hua, B.; Wang, H.; Sun, H. Safety analysis in patients with autoimmune disease receiving allogeneic mesenchymal stem cells infusion: A long-term retrospective study. Stem Cell Res. Ther. 2018, 9, 312. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Chen, S.; Yang, P.; Cao, H.; Li, L. The role of mesenchymal stem cells in hematopoietic stem cell transplantation: Prevention and treatment of graft-versus-host disease. Stem Cell Res. Ther. 2019, 10, 182. [Google Scholar] [CrossRef]
- Tessier, L.; Bienzle, D.; Williams, L.B.; Koch, T.G. Phenotypic and Immunomodulatory Properties of Equine Cord Blood-Derived Mesenchymal Stromal Cells. PLoS ONE 2015, 10, e0122954. [Google Scholar] [CrossRef] [PubMed]
- Son, M.Y.; Lee, M.O.; Jeon, H.; Seol, B.; Kim, J.H.; Chang, J.S.; Cho, Y.S. Generation and characterization of integration-free induced pluripotent stem cells from patients with autoimmune disease. Exp. Mol. Med. 2016, 48, e232. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Type of MSC | Mouse Model | Administration | Follow Up | Major Effects of MSC Treatment | Ref. | |
|---|---|---|---|---|---|---|
| Increase | Decrease | |||||
| OE-MSCs BMMSCs | C57BL/6 mice with experimental SS induced by immunization with SG proteins | 2 IV injections (day 18 and 25 after immunization) | Day 42 after immunization | - Salivary flow rate | - Histological score in SG - Anti-M3R and anti-SSA/Ro autoantibody levels in serum (only with BMMSCs) -Production of inflammatory cytokines (IFN-γ and IL-17) | [25] |
| BMMSCs | Day 35 and Week 15 after immunization | - Histological score in SG - Anti-M3R and anti-nuclear autoantibody levels in serum (only for BMMSCs) - Production of inflammatory cytokines (IFNγ and IL-17) | [27] | |||
| DPSCs BMMSCs | MRL/MpJ-faslpr/faslpr (MRL/lpr) | 4 IV injections of conditioned-media MSC (twice a week) | Week 2 after last injection | - Salivary flow rate - Production of anti-inflammatory cytokines (TFGβ1, IL-10 and IL-13) | - Focus score of SG - Epithelial cell apoptosis in SG | [22] |
| DPSCs BMMSCs | NOD mice | 4 IV injections of conditioned-media MSC (twice a week) | Week 2 after last injection | - Salivary flow rate - Production of anti-inflammatory cytokines (TFGβ1, IL-10, and IL-13) | - Focus score of SG (only with DPSCs) - Epithelial cell apoptosis in SG | [21] |
| BMMSCs | NOD/Ltj mice (Cdh23ahl) | IV injections at early stage of SS (week 6 = prevention group) or at developed stage (week 16 = treatment group) | Week 2 after injection | - Salivary flow rate in 2 groups - Production of anti-inflammatory cytokines (TFGβ1, IL-10, and IL-13) in treatment group | - Anti-nucleic, anti-α-fodrine and anti-SSA/Ro autoantibody levels in serum in treatment group - Production of inflammatory cytokines (IFN-γ, IL-6, and IL-17) in treatment group | [28] |
| Id3-deficient BMMSCs | NOD/ShiLtj mice | IV injection of Id3-deficient BMMSCs or WT BMMSCs | Week 2 after injection | - Salivary flow rate, better with Id3-deficient BMMSCs | / | [18] |
| ME-MSCs | NOD/Ltj mice | 4 IV injections of MSC (twice a week) | Week 2 after last injection | - Salivary flow rate | - Epithelial cell apoptosis in SG | [16] |
| BMMSCs Extract | NOD mice | 4 IV injections (once a week) | Baseline Week 4, 8, 12, and 16 after last injection | - Salivary flow rate - Tear flow rate - Production of anti-inflammatory cytokines (IL-10) | - Anti-SSA/Ro autoantibody levels in serum | [14] |
| SHED | NOD mice | 2 IV injections (once a week) | Baseline Week 1 Week 2 | - Stimulated salivary flow rate at week 2 | - Focus score of SG at week 2 - Epithelial cell apoptosis in SG | [15] |
| iPSC-MSCs BMMSCs | NOD/ShiLtj mice | 2 IV injections (once a week) | Week 3 after last injection | / | - Anti-SSA/Ro and anti-SSB/La autoantibody levels in serum | [17] |
| hUCMSCs | NOD mice | IV injection | Week 4 | - Salivary flow rate | / | [23] |
| UCMSCs | NOD/Ltj mice | IV injection | Day 28 after injection | - Salivary flow rate | / | [26] |
| BMSCs | NOD/Ltj mice | 4 IV injections (twice a week) | Week 4 | - Salivary flow rate -Production of anti-inflammatory cytokines (TFGβ1 and IL-10) | - Focus score of SG - Production of inflammatory cytokines (IFNγ) | [24] |
| BMSCs CD45-/TER119- | NOD mice | 4 IV injections (twice a week) | Baseline Week 10 Week 14 | - Salivary flow rate | - Lymphocytic infiltrate in SG | [19] |
| BMSCs | NOD mice | 12 IV injections (twice a week) | Baseline Week 2, 12, 34, 38, 44, and 52 | - Salivary flow rate (week 12, 34, and 52) | / | [20] |
| UCMSCs | NOD mice | 5 Intraperitoneal injections (once a day) | Week 8 or Week 12 | - Salivary flow rate - Production of anti-inflammatory cytokines (IL-10) | - Anti-α-fodrine and anti-SSA/Ro autoantibody levels in serum - Histological score in SG - Production of inflammatory cytokines (IL-6) | [30] |
| UCMSCs | NOD/Ltj mice | IV injection | Week 4 after injection | - Salivary flow rate | / | [29] |
| Type of MSC | In Vivo/In Vitro Study | Follow Up | Major Effects of MSC | Ref. | |
|---|---|---|---|---|---|
| Increase | Decrease | ||||
| UCMSCs | In vivo IV injection (10 patients) | Day 7 | - IL-12 levels in serum | / | [26] |
| UCMSCs | In vivo IV injection (24 patients) | Baseline Week 2 Month 1 Month 3 Month 6 Month 12 | - Unstimulated and stimulated salivary flow rate for most of 11 patients | - SSDAI score for all 24 patients and VAS score for most of patients - SSA/Ro autoantibodies for 7 patients - SSB/La autoantibodies for 6 patients | [28] |
| UCMSCs | In vivo IV injection (38 patients) | Week 1 | - IL-27 and TGFβ levels in serum - Tregs among PBMCs | - IL-17-expressing T cells among PBMCs | [29] |
| In vitro Coculture with monocyte-derived DCs | Unknown | - IL-27 production of DCs | / | ||
| UCMSCs | In vitro Coculture with PBMC from SS patients or from HC | Day 4 | / | - Expansion and differentiation of Tfh from PBMC from SS patients | [32] |
| UCMSCs free or encapsulated | In vitro Coculture with PBMC from SS patients or from HC | Day 4 | - Tregs among PBMCs (with encapsulated UCMSCs) | - Proliferation of T cells from SS patients (only with encapsulated UCMSCs) | [31] |
| UCMSCs | In vitro Coculture with CD4+ T cells from blood of SS patients or of HC | Day 3 | - IL-6 production in supernatants - Upregulation of 13 miRNA whose miR-115–5p having impact on TCR signaling pathway | - Proliferation of T cells from SS patients - IFNγ production in supernatants | [33] |
| BMMSCs | In vitro Coculture of PBMC from HC with BMMSCs from HC or SS patients | Day 4 | - T cell proliferation with BMMSCs from SS patients compared to BMMSCs from HC | / | [28] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chihaby, N.; Orliaguet, M.; Le Pottier, L.; Pers, J.-O.; Boisramé, S. Treatment of Sjögren’s Syndrome with Mesenchymal Stem Cells: A Systematic Review. Int. J. Mol. Sci. 2021, 22, 10474. https://doi.org/10.3390/ijms221910474
Chihaby N, Orliaguet M, Le Pottier L, Pers J-O, Boisramé S. Treatment of Sjögren’s Syndrome with Mesenchymal Stem Cells: A Systematic Review. International Journal of Molecular Sciences. 2021; 22(19):10474. https://doi.org/10.3390/ijms221910474
Chicago/Turabian StyleChihaby, Najwa, Marie Orliaguet, Laëtitia Le Pottier, Jacques-Olivier Pers, and Sylvie Boisramé. 2021. "Treatment of Sjögren’s Syndrome with Mesenchymal Stem Cells: A Systematic Review" International Journal of Molecular Sciences 22, no. 19: 10474. https://doi.org/10.3390/ijms221910474
APA StyleChihaby, N., Orliaguet, M., Le Pottier, L., Pers, J.-O., & Boisramé, S. (2021). Treatment of Sjögren’s Syndrome with Mesenchymal Stem Cells: A Systematic Review. International Journal of Molecular Sciences, 22(19), 10474. https://doi.org/10.3390/ijms221910474