
Relationships between Mitochondrial Function, AMPK, and TORC1 Signaling in Lymphoblasts with Premutation Alleles of the FMR1 Gene

 ,
, 

Abstract
:1. Introduction
2. Results
2.1. Participant Cohorts
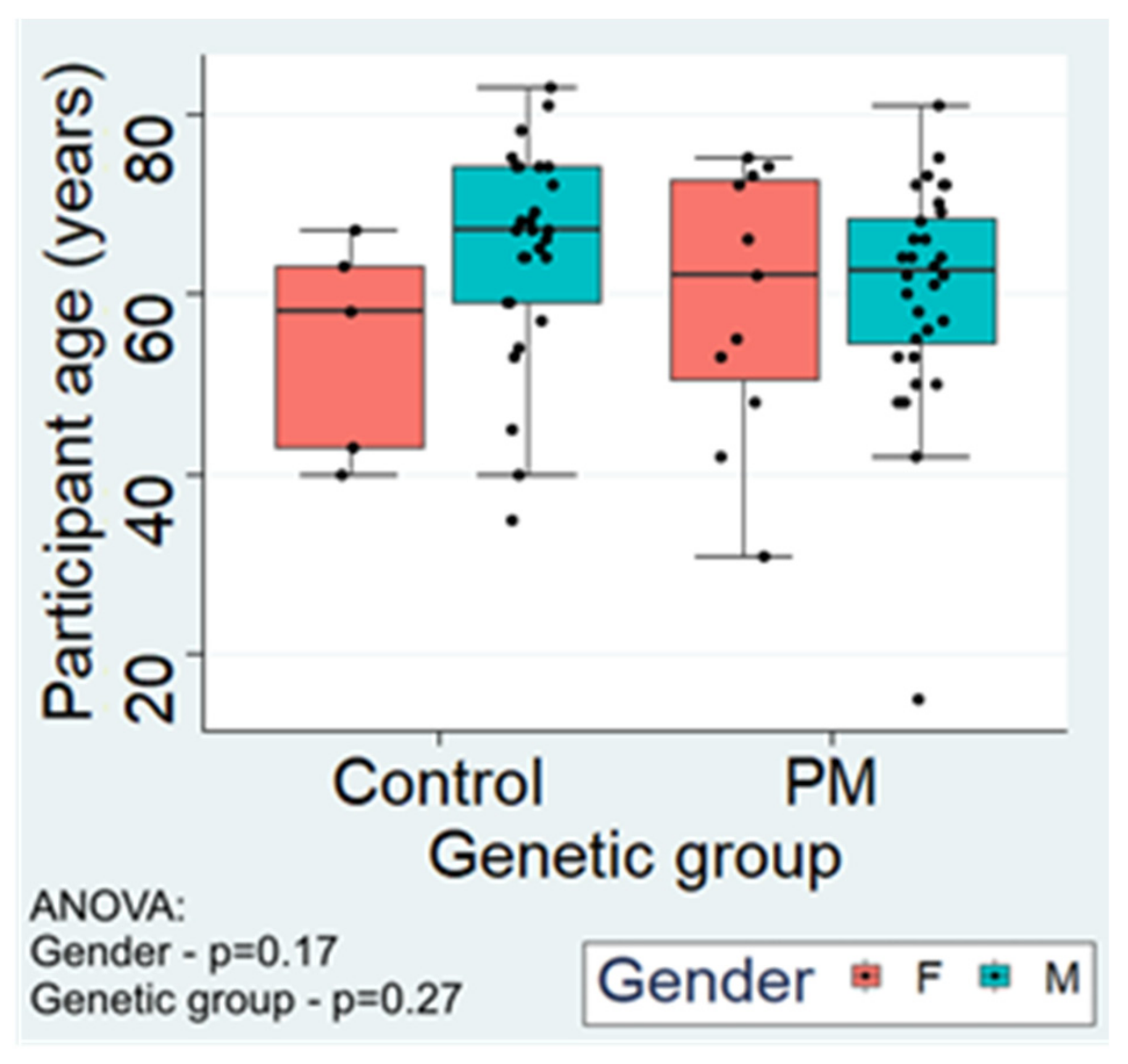
2.1.1. Participant Age and Gender Distribution
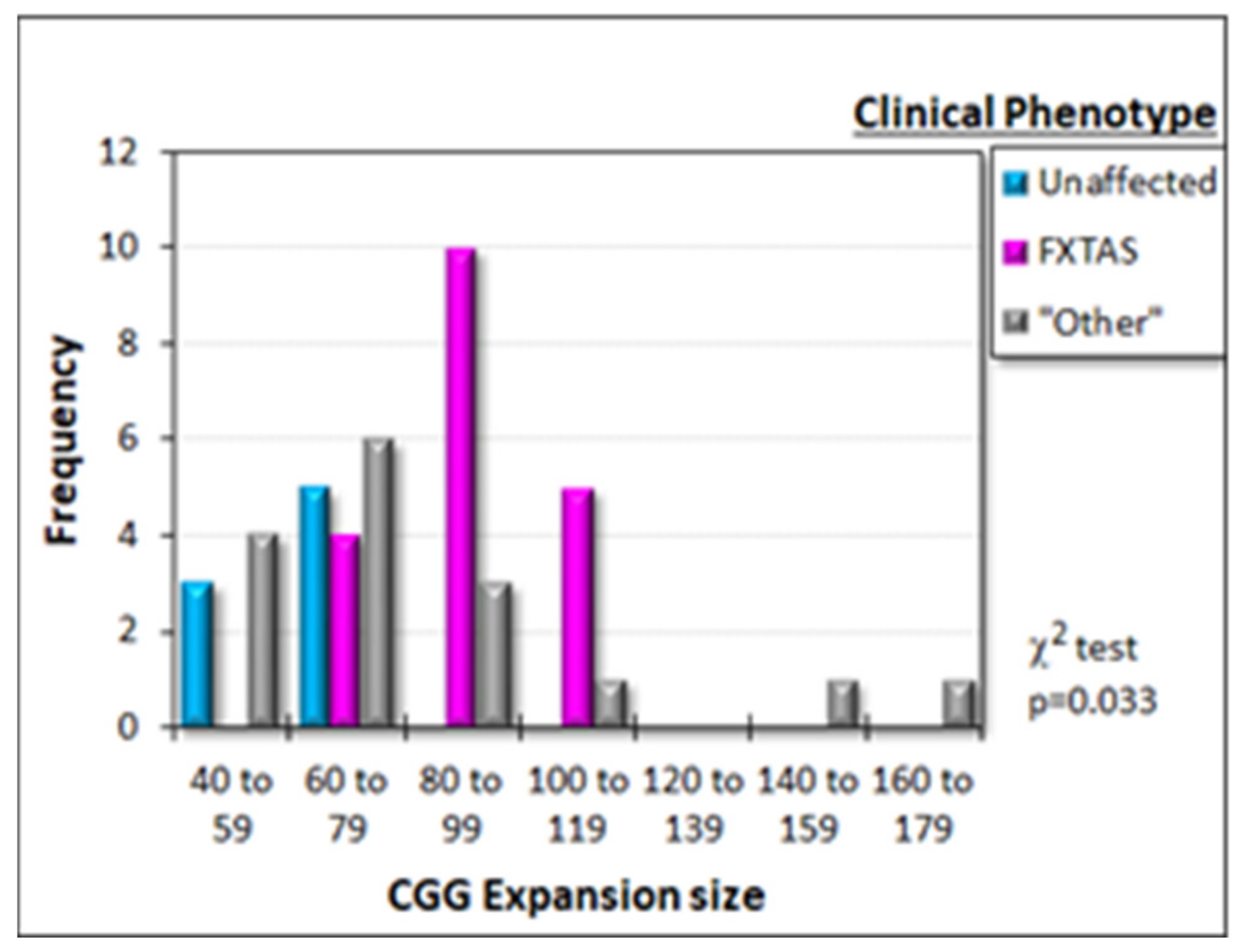
2.1.2. Clinical Phenotypes and CGG Expansion Sizes
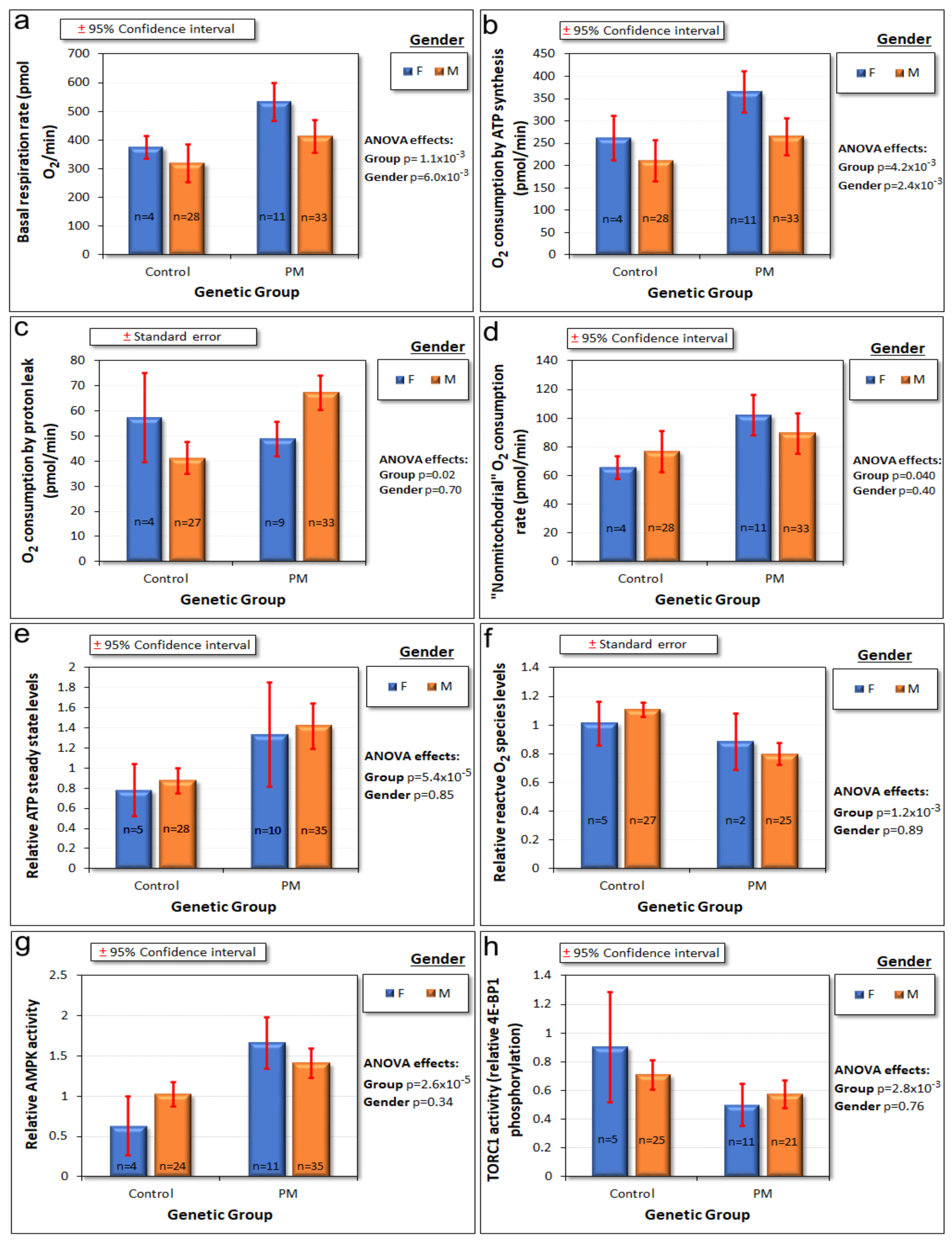
2.2. Mitochondrial Function, ATP Steady State Levels, Redox Balance and Associated Signaling Activities Are Abnormal in PM Lymphoblasts
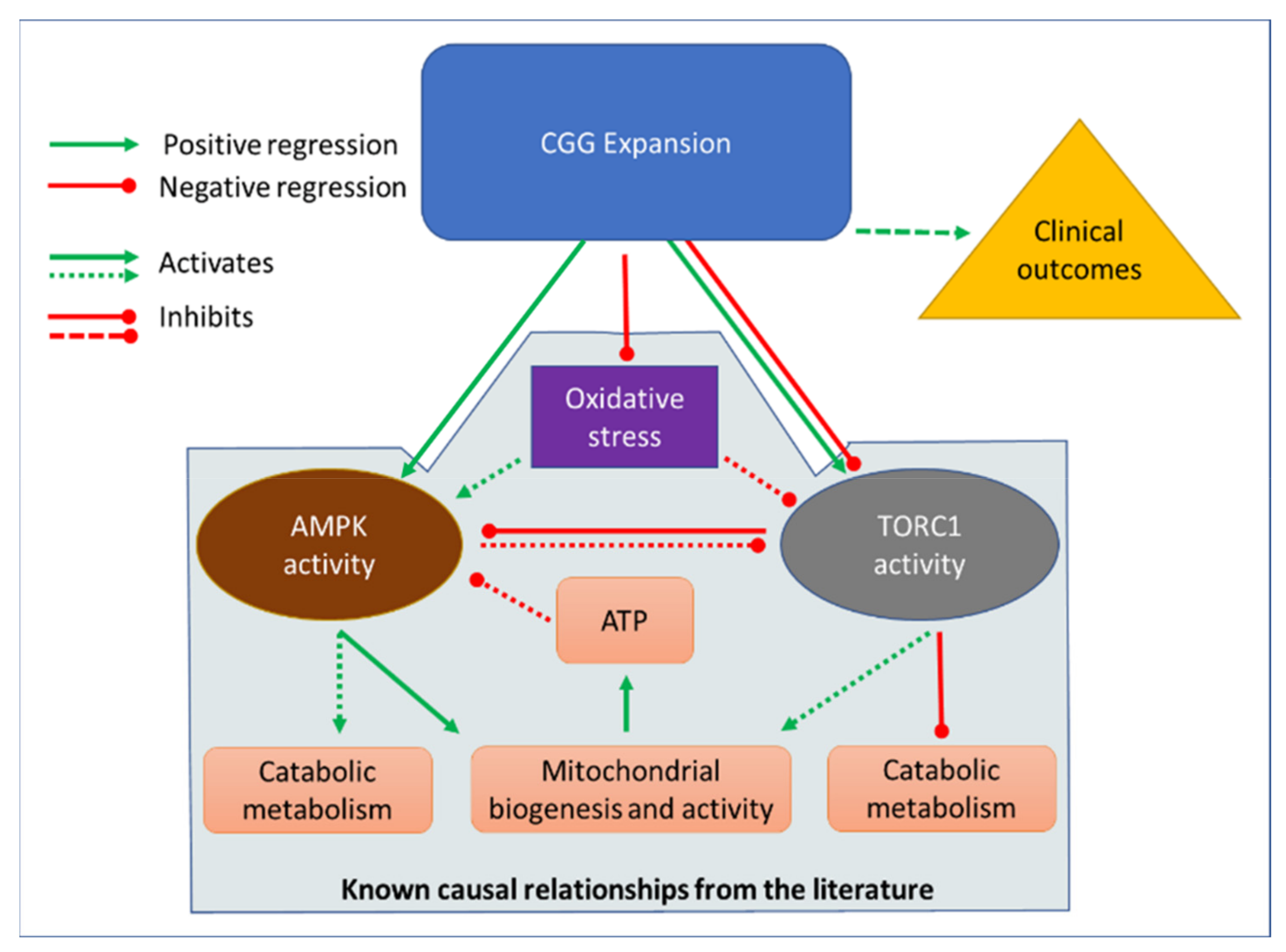
2.3. Multiple Regression Relationships between CGG Repeat Number, Mitochondrial Activity, and Cellular Stress Signaling by AMPK and TORC1
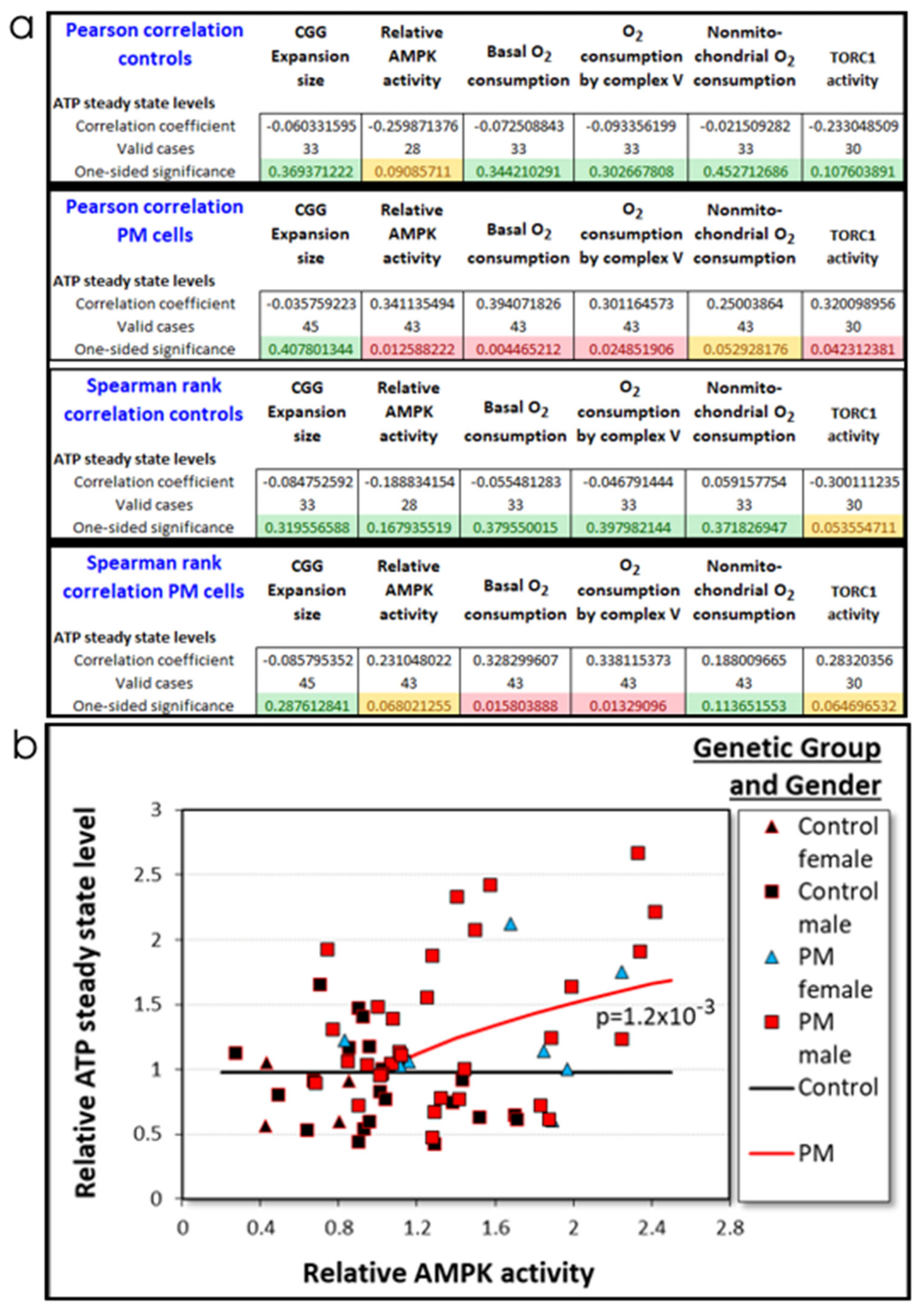
2.3.1. Elevated AMPK Activity Can Explain the Elevated Rates of Mitochondrial Oxidative Phosphorylation in PM Lymphoblasts
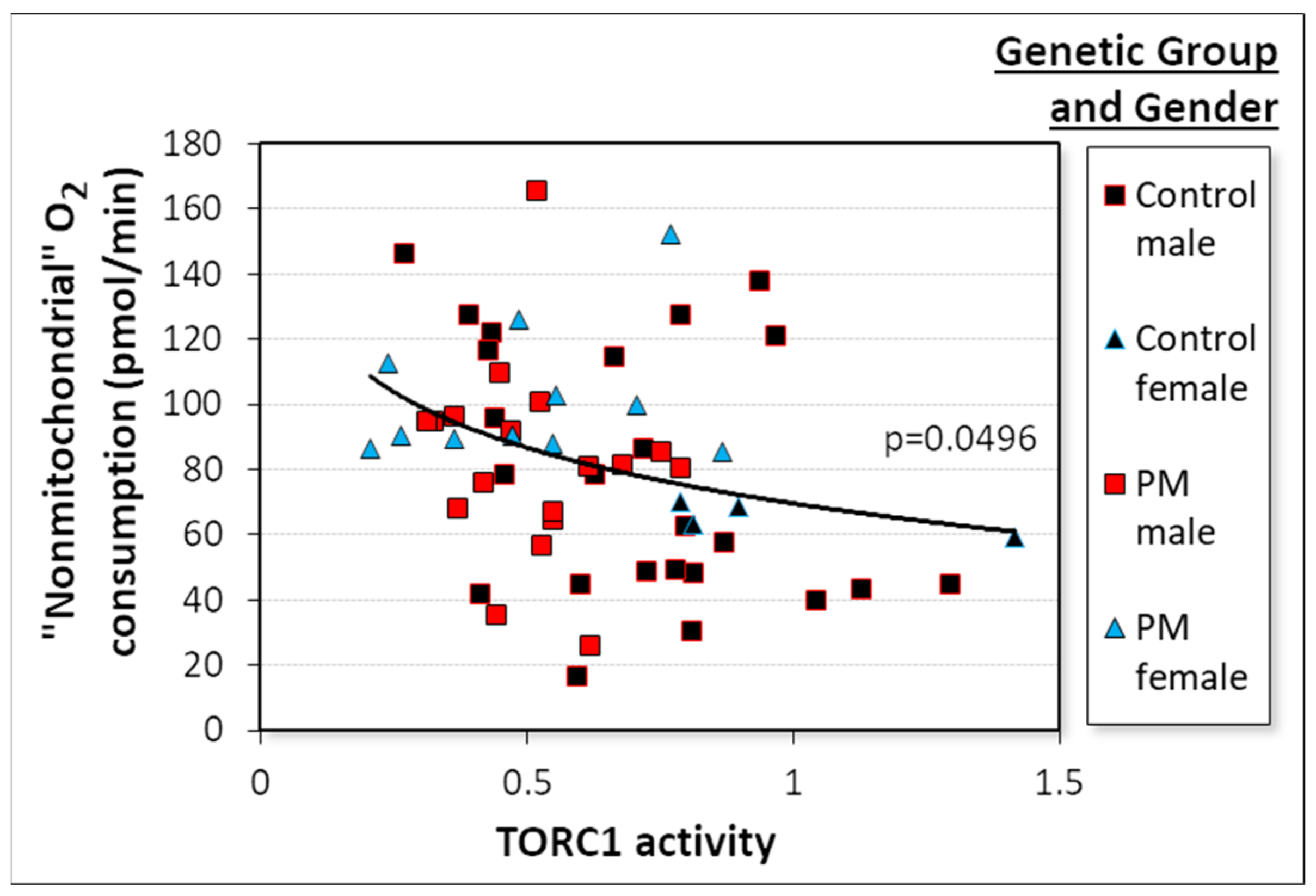
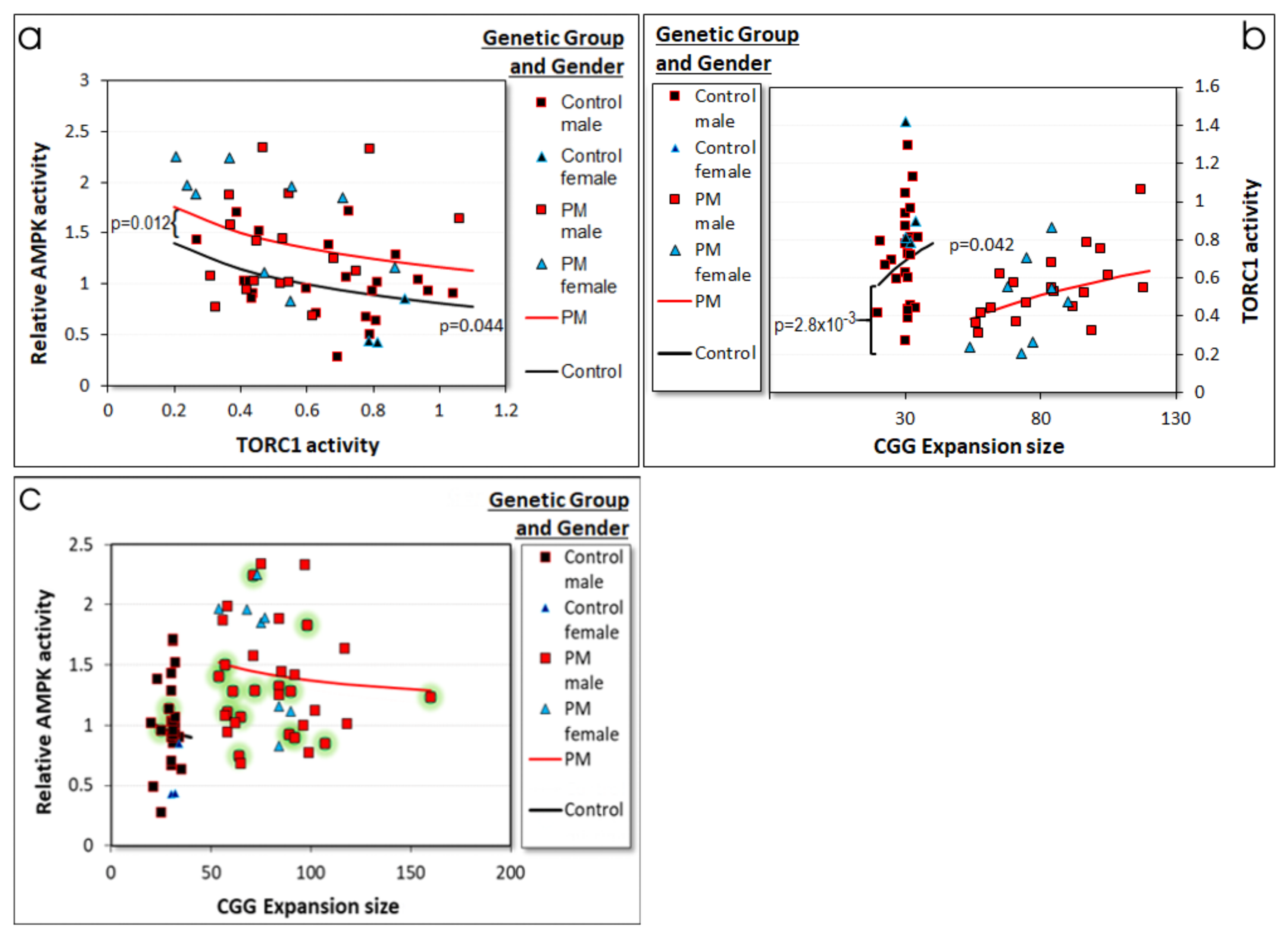
2.3.2. AMPK Activity in Control and PM Lymphoblasts Depends on TORC1 Activity, Which Depends on the CGG Repeat Number
2.3.3. Elevated ATP Steady State Levels in PM Lymphoblasts Depend on AMPK Activity in PM Cells
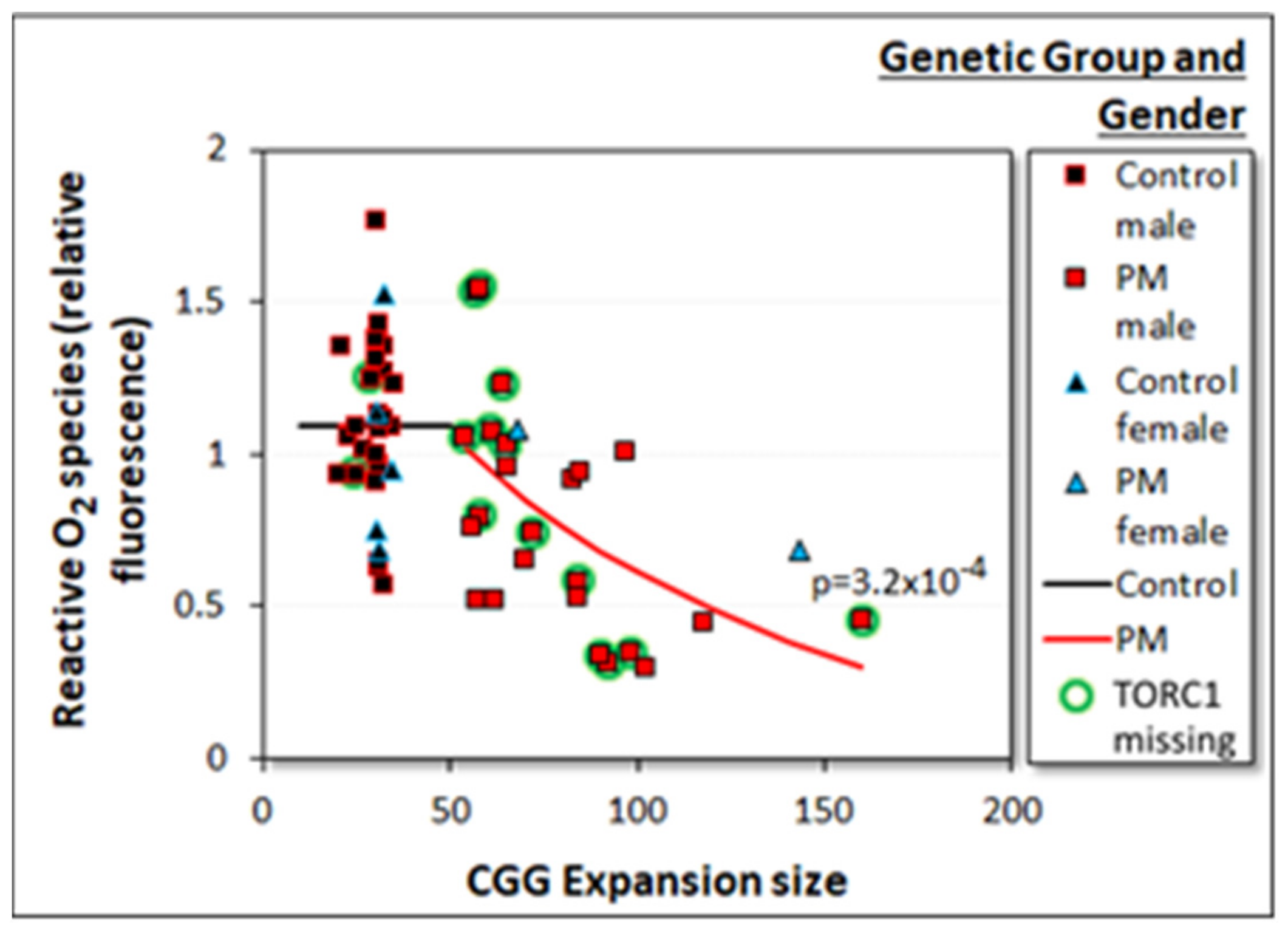
2.3.4. Reactive O2 Species (ROS) Levels Are Reduced in PM Lymphoblasts in a CGG Repeat Number-Dependent Manner
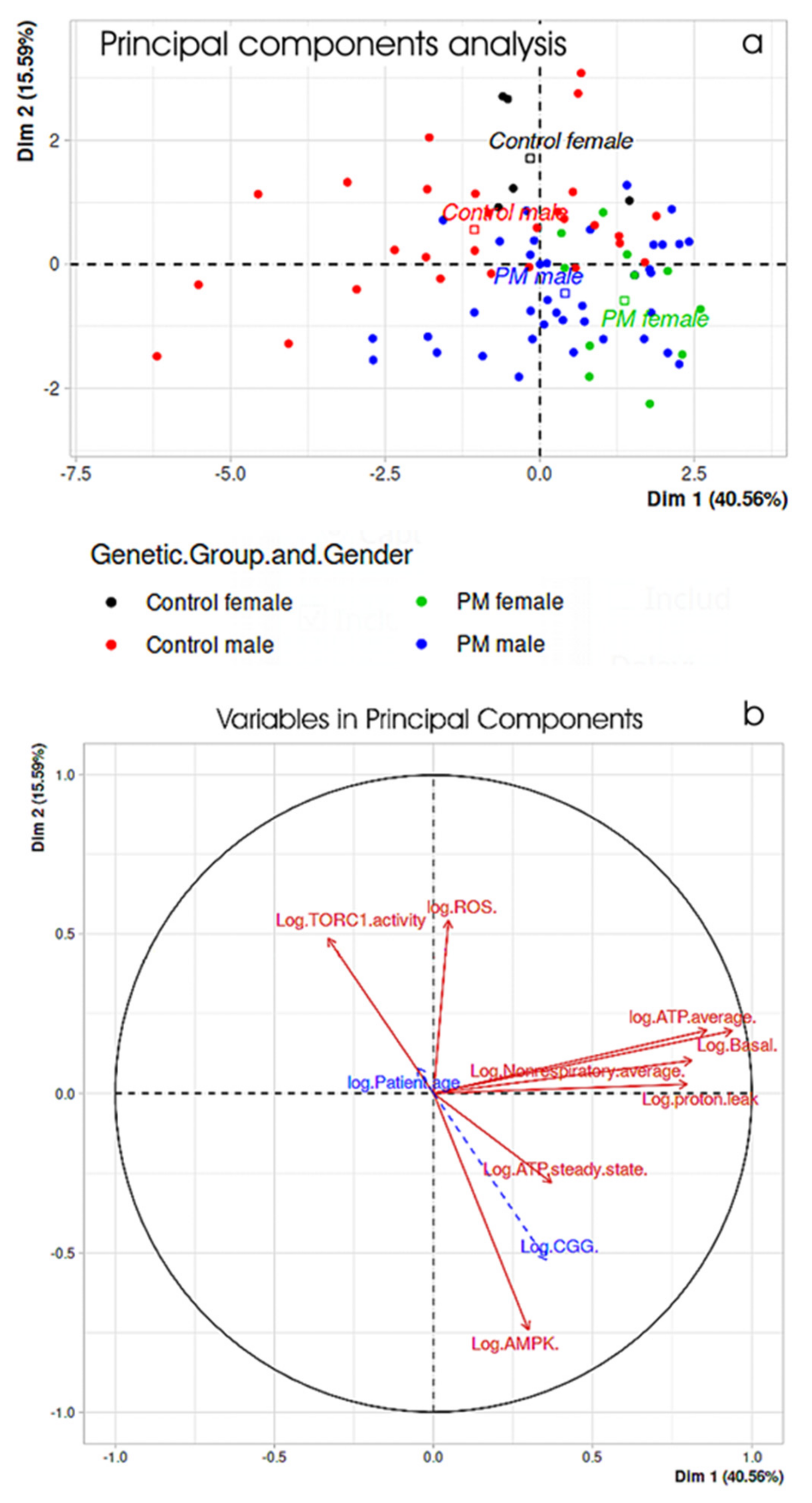
2.4. Principal Components Analysis Confirms the Major Interrelationships Amongst Measures of Mitochondrial Function and Cellular Stress Signaling in PM Lymphoblasts
3. Discussion
4. Materials and Methods
4.1. Sample Description
4.2. Cell culture
4.2.1. PBMC Isolation from Blood and Immortalization
4.2.2. Lymphoblast Culture
4.3. Functional Assays
4.3.1. FMR1 CGG Repeat Number
4.3.2. Mitochondrial Mass and Membrane Potential
4.3.3. Seahorse Respirometry
4.3.4. AMPK Activity
4.3.5. TORC1 Activity
4.3.6. Reactive Oxygen Species (ROS)
4.3.7. ATP Steady State Levels
4.4. Statistical Analysis
4.4.1. Two- and Multi-Sample Tests
4.4.2. Correlation and Linear Regression Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hagerman, R.J.; Leehey, M.; Heinrichs, W.; Tassone, F.; Wilson, R.; Hills, J.; Grigsby, J.; Gage, B. Intention tremor, parkinsonism, and generalized brain atrophy in male carriers of fragile X. Neurology 2001, 57, 127–130. [Google Scholar] [CrossRef]
- Hagerman, P.J.; Hagerman, R.J. Fragile X-associated tremor/ataxia syndrome. Ann. N. Y. Acad. Sci. 2015, 1338, 58–70. [Google Scholar] [CrossRef]
- Cabal-Herrera, A.M.; Tassanakijpanich, N.; Salcedo-Arellano, M.J.; Hagerman, R.J. Fragile X-Associated Tremor/Ataxia Syndrome (FXTAS): Pathophysiology and Clinical Implications. Int. J. Mol. Sci. 2020, 21, 4391. [Google Scholar] [CrossRef] [PubMed]
- Loesch, D.; Hagerman, R. Unstable Mutations in the FMR1 Gene and the Phenotypes. Cancer Biol. Nucl. Envel. 2012, 769, 78–114. [Google Scholar] [CrossRef] [Green Version]
- Loesch, D.Z.; Trost, N.; Bui, M.; Hammersley, E.; Lay, S.T.; Annesley, S.J.; Sanislav, O.; Allan, C.Y.; Tassone, F.; Chen, Z.-P.; et al. The Spectrum of Neurological and White Matter Changes and Premutation Status Categories of Older Male Carriers of the FMR1 Alleles Are Linked to Genetic (CGG and FMR1 mRNA) and Cellular Stress (AMPK) Markers. Front. Genet. 2018, 9, 531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, D.; Tassone, F.; Klepitskaya, O.; Leehey, M. Fragile X-associated tremor ataxia syndrome in FMR1 gray zone allele carriers. Mov. Disord. 2011, 27, 297–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salcedo-Arellano, M.J.; Dufour, B.; McLennan, Y.; Martinez-Cerdeno, V.; Hagerman, R. Fragile X syndrome and associated disorders: Clinical aspects and pathology. Neurobiol. Dis. 2020, 136, 104740. [Google Scholar] [CrossRef] [PubMed]
- Allen, E.G.; Charen, K.; Hipp, H.; Shubeck, L.; Amin, A.; He, W.; Hunter, J.E.; Sherman, S.L. Clustering of comorbid conditions among women who carry an FMR1 premutation. Genet. Med. 2020, 22, 758–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagerman, R.J.; Protic, D.; Rajaratnam, A.; Salcedo, M.; Aydin, E.Y.; Schneider, A. Fragile X-Associated Neuropsychiatric Disorders (FXAND). Front. Psychiatry 2018, 9, 564. [Google Scholar] [CrossRef] [Green Version]
- Martin, E.M.; Zhu, Y.; Kraan, C.M.; Kumar, K.R.; Godler, D.; Field, M. Men with FMR1 premutation alleles of less than 71 CGG repeats have low risk of being affected with fragile X-associated tremor/ataxia syndrome (FXTAS). J. Med. Genet. 2021, 24, 1225–1234. [Google Scholar] [CrossRef]
- Ross-Inta, C.; Omanska-Klusek, A.; Wong, S.; Barrow, C.; Garcia-Arocena, D.; Iwahashi, C.; Berry-Kravis, E.; Hagerman, R.J.; Hagerman, P.J.; Giulivi, C. Evidence of mitochondrial dysfunction in fragile X-associated tremor/ataxia syndrome. Biochem. J. 2010, 429, 545–552. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Mora, M.I.; Rodriguez-Revenga, L.; Madrigal, I.; Guitart-Mampel, M.; Garrabou, G.; Milà, M. Impaired Mitochondrial Function and Dynamics in the Pathogenesis of FXTAS. Mol. Neurobiol. 2017, 54, 6896–6902. [Google Scholar] [CrossRef]
- Napoli, E.; McLennan, Y.A.; Schneider, A.; Tassone, F.; Hagerman, R.J.; Giulivi, C. Characterization of the Metabolic, Clinical and Neuropsychological Phenotype of Female Carriers of the Premutation in the X-Linked FMR1 Gene. Front. Mol. Biosci. 2020, 7, 578640. [Google Scholar] [CrossRef] [PubMed]
- Loesch, D.Z.; Annesley, S.J.; Trost, N.; Bui, M.; Lay, S.T.; Storey, E.; De Piazza, S.W.; Sanislav, O.; Francione, L.M.; Hammersley, E.M.; et al. Novel Blood Biomarkers Are Associated with White Matter Lesions in Fragile X-Associated Tremor/Ataxia Syndrome. Neurodegener. Dis. 2016, 17, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Hindupur, S.K.; González, A.; Hall, M.N. The Opposing Actions of Target of Rapamycin and AMP-Activated Protein Kinase in Cell Growth Control. Cold Spring Harb. Perspect. Biol. 2015, 7, a019141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, N.X.Y.; Kaczmarek, A.; Hoque, A.; Davie, E.; Ngoei, K.R.W.; Morrison, K.R.; Smiles, W.; Forte, G.M.; Wang, T.; Lie, S.; et al. mTORC1 directly inhibits AMPK to promote cell proliferation under nutrient stress. Nat. Metab. 2020, 2, 41–49. [Google Scholar] [CrossRef]
- He, W.; Yadav-Shah, M.; Sherman, S.L.; Allen, E.G. A study of the distributional characteristics of FMR1 transcript levels in 238 individuals. Qual. Life Res. 2004, 114, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Loesch, D.Z.; Kemp, B.E.; Bui, M.Q.; Fisher, P.R.; Allan, C.Y.; Sanislav, O.; Ngoei, K.R.W.; Atkinson, A.; Tassone, F.; Annesley, S.J.; et al. Cellular bioenergetics and AMPK and TORC1 signalling in blood lymphoblasts are biomarkers of clinical status in FMR1 premutation carriers. Front. Psych. 2021. manuscript under minor revision. [Google Scholar]
- Napoli, E.; Schneider, A.; Hagerman, R.; Song, G.; Wong, S.; Tassone, F.; Giulivi, C. Impact of FMR1 Premutation on Neurobehavior and Bioenergetics in Young Monozygotic Twins. Front. Genet. 2018, 9, 338. [Google Scholar] [CrossRef]
- Song, G.; Napoli, E.; Wong, S.; Hagerman, R.; Liu, S.; Tassone, F.; Giulivi, C. Altered Redox Mitochondrial Biology in the Neurodegenerative Disorder Fragile X-Tremor/Ataxia Syndrome: Use of Antioxidants in Precision Medicine. Mol. Med. 2016, 22, 548–559. [Google Scholar] [CrossRef] [Green Version]
- Inoki, K.; Zhu, T.; Guan, K.-L. TSC2 Mediates Cellular Energy Response to Control Cell Growth and Survival. Cell 2003, 115, 577–590. [Google Scholar] [CrossRef] [Green Version]
- Gohel, D.; Berguerand, N.C.; Tassone, F.; Singh, R. The emerging molecular mechanisms for mito-chondrial dysfunctions in FXTAS. Biochim. Et Biophys. Acta-Mol. Basis Dis. 2020, 866, 165918. [Google Scholar]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of metabolism and mitochondrial homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.Y.; Sabatini, D.M. mTOR at the nexus of nutrition, growth, ageing and disease. Nat. Rev. Mol. Cell Biol. 2020, 21, 183–203. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, J.T.; Rodgers, J.T.; Arlow, D.H.; Vazquez, F.; Mootha, V.K.; Puigserver, P. mTOR controls mitochondrial oxidative function through a YY1–PGC-1α transcriptional complex. Nature 2007, 450, 736–740. [Google Scholar] [CrossRef]
- Cardinale, D.A.; Larsen, F.J.; Schiffer, T.A.; Morales-Alamo, D.; Ekblom, B.; Calbet, J.A.L.; Holmberg, H.-C.; Boushel, R. Superior Intrinsic Mitochondrial Respiration in Women Than in Men. Front. Physiol. 2018, 9, 1133. [Google Scholar] [CrossRef]
- Ventura-Clapier, R.; Moulin, M.; Piquereau, J.; Lemaire, C.; Mericskay, M.; Veksler, V.; Garnier, A. Mitochondria: A central target for sex differences in pathologies. Clin. Sci. 2017, 131, 803–822. [Google Scholar] [CrossRef]
- Sharma, A.; Hoeffer, C.A.; Takayasu, Y.; Miyawaki, T.; McBride, S.M.; Klann, E.; Zukin, R.S. Dysregulation of mTOR Signaling in Fragile X Syndrome. J. Neurosci. 2010, 30, 694–702. [Google Scholar] [CrossRef]
- Hoeffer, C.A.; Sanchez, E.; Hagerman, R.J.; Mu, Y.; Nguyen, D.V.; Wong, H.; Whelan, A.M.; Zukin, R.S.; Klann, E.; Tassone, F. Altered mTOR signaling and enhanced CYFIP2 expression levels in subjects with fragile X syndrome. Genes Brain Behav. 2012, 11, 332–341. [Google Scholar] [CrossRef]
- Lu, C.; Lin, L.; Tan, H.; Wu, H.; Sherman, S.L.; Gao, F.; Jin, P.; Chen, D. Fragile X premutation RNA is sufficient to cause primary ovarian insufficiency in mice. Hum. Mol. Genet. 2012, 21, 5039–5047. [Google Scholar] [CrossRef] [Green Version]
- Gantois, I.; Khoutorsky, A.; Popic, J.; Aguilar-Valles, A.; Freemantle, E.; Cao, R.; Sharma, V.; Pooters, T.; Nagpal, A.; Skalecka, A.; et al. Metformin ameliorates core deficits in a mouse model of fragile X syndrome. Nat. Med. 2017, 23, 674–677. [Google Scholar] [CrossRef]
- Sellier, C.; Rau, F.; Liu, Y.; Tassone, F.; Hukema, R.; Gattoni, R.; Schneider, A.; Richard, S.; Willemsen, R.; Elliott, D.; et al. Sam68 sequestration and partial loss of function are associated with splicing alterations in FXTAS patients. EMBO J. 2010, 29, 1248–1261. [Google Scholar] [CrossRef] [Green Version]
- Sellier, C.; Freyermuth, F.; Tabet, R.; Tran, T.; He, F.; Ruffenach, F.; Alunni, V.; Moine, H.; Thibault-Carpentier, C.; Page, A.; et al. Sequestration of DROSHA and DGCR8 by Expanded CGG RNA Repeats Alters MicroRNA Processing in Fragile X-Associated Tremor/Ataxia Syndrome. Cell Rep. 2013, 3, 869–880. [Google Scholar] [CrossRef] [Green Version]
- Krans, A.; Skariah, G.; Zhang, Y.; Bayly, B.; Todd, P.K. Neuropathology of RAN translation proteins in fragile X-associated tremor/ataxia syndrome. Acta Neuropathol. Commun. 2019, 7, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manford, A.G.; Rodríguez-Pérez, F.; Shih, K.Y.; Shi, Z.; Berdan, C.A.; Choe, M.; Titov, D.V.; Nomura, D.K.; Rape, M. A Cellular Mechanism to Detect and Alleviate Reductive Stress. Cell 2020, 183, 46–61.e21. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Loscalzo, J. Metabolic Responses to Reductive Stress. Antioxid. Redox Signal. 2020, 32, 1330–1347. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.Y.; Napoli, E.; Kim, K.; McLennan, Y.A.; Hagerman, R.J.; Giulivi, C. Brain Atrophy and White Matter Damage Linked to Peripheral Bioenergetic Deficits in the Neurodegenerative Disease FXTAS. Int. J. Mol. Sci. 2021, 22, 9171. [Google Scholar] [CrossRef] [PubMed]
- Missailidis, D.; Annesley, S.J.; Allan, C.Y.; Sanislav, O.; Lidbury, B.A.; Lewis, D.P.; Fisher, P.R. An Isolated Complex V Inefficiency and Dysregulated Mitochondrial Function in Immortalized Lymphocytes from ME/CFS Patients. Int. J. Mol. Sci. 2020, 21, 1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Missailidis, D.; Sanislav, O.; Allan, C.Y.; Annesley, S.J.; Fisher, P.R. Cell-Based Blood Biomarkers for Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Int. J. Mol. Sci. 2020, 21, 1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annesley, S.; Lay, S.T.; De Piazza, S.W.; Sanislav, O.; Hammersley, E.; Allan, C.Y.; Francione, L.M.; Bui, M.; Chen, Z.-P.; Ngoei, K.R.W.; et al. Immortalized Parkinson’s Disease lymphocytes have enhanced mitochondrial respiratory activity. Dis. Model. Mech. 2016, 9, 1295–1305. [Google Scholar] [CrossRef] [Green Version]
- Ransohoff, R.M.; Schafer, D.P.; Vincent, A.; Blachère, N.E.; Bar-Or, A. Neuroinflammation: Ways in Which the Immune System Affects the Brain. Neurotherapeutics 2015, 12, 896–909. [Google Scholar] [CrossRef] [Green Version]
- Hernando, H.; Shannon-Lowe, C.; Islam, A.B.; Al-Shahrour, F.; Rodriguez-Ubreva, J.; Cortez, V.C.R.; Javierre, B.M.; Mangas, C.; Fernández, A.F.; Parra, M.; et al. The B cell transcription program mediates hypomethylation and overexpression of key genes in Epstein-Barr virus-associated proliferative conversion. Genome Biol. 2013, 14, R3. [Google Scholar] [CrossRef] [Green Version]
- Schafflick, D.; Wolbert, J.; Heming, M.; Thomas, C.; Hartlehnert, M.; Börsch, A.-L.; Ricci, A.; Mar-tín-Salamanca, S.; Li, X.; Lu, I.-N.; et al. Single-cell profiling of CNS border com-partment leukocytes reveals that B cells and their progenitors reside in non-diseased meninges. Nat. Neurosc. 2021, 24, 1225–1234. [Google Scholar] [CrossRef]
- Hussain, T.; Mulherkar, R. Lymphoblastoid Cell lines: A Continuous in Vitro Source of Cells to Study Carcinogen Sensitivity and DNA Repair. Int. J. Mol. Cell. Med. 2012, 1, 75–87. [Google Scholar] [PubMed]
- Sie, L.; Loong, S.; Tan, E.K. Utility of lymphoblastoid cell lines. J. Neurosci. Res. 2009, 87, 1953–1959. [Google Scholar] [CrossRef] [PubMed]
- Annesley, S.; Fisher, P. Lymphoblastoid Cell Lines as Models to Study Mitochondrial Function in Neurological Disorders. Int. J. Mol. Sci. 2021, 22, 4536. [Google Scholar] [CrossRef] [PubMed]
- Tassone, F.; Pan, R.; Amiri, K.; Taylor, A.K.; Hagerman, P.J. A Rapid Polymerase Chain Reaction-Based Screening Method for Identification of All Expanded Alleles of the Fragile X (FMR1) Gene in Newborn and High-Risk Populations. J. Mol. Diagn. 2008, 10, 43–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pendergrass, W.; Wolf, N.; Poot, M. Efficacy of MitoTracker Green? and CMXrosamine to measure changes in mitochondrial membrane potentials in living cells and tissues. Cytom. A 2004, 61A, 162–169. [Google Scholar] [CrossRef]
- Stapleton, D.; Mitchelhill, K.I.; Gao, G.; Widmer, J.; Michell, B.J.; Teh, T.; House, C.M.; Fernandez, C.S.; Cox, T.; Witters, L.A.; et al. Mammalian AMP-activated Protein Kinase Subfamily. J. Biol. Chem. 1996, 271, 611–614. [Google Scholar] [CrossRef] [Green Version]
- Venables, W.N.; Ripley, B.D. Package MASS. Available online: http://www.r-project.org (accessed on 17 October 2012).
- Fox, J. Using the R Commander: A Point-and-Click Interface for R; Chapman and Hall/CRC Press: London, UK, 2017. [Google Scholar]
- Chan, G.; Leung, J.; Anonymous; Lim, C.; Sun, T.; Stjepa-Nović, D. StatsNotebook, A Graphical User Interface for Data Analysis and Visualisation in R. 2020. Available online: https://statsnotebook.io/ (accessed on 4 June 2021).
- Friedman, J.; Hastie, T.; Tibshirani, R. Regularization Paths for Generalized Linear Models via Co-ordinate Descent. J. Stat. Softw. 2010, 33, 1–22. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spearman Rank Correlations Amongst Mitochondrial Respiratory Activities and ATP Levels | Premutation Lymphoblasts | |||||||
|---|---|---|---|---|---|---|---|---|
| Basal OCR | OCR by ATP Synthesis | Maximum OCR | Complex I OCR | “Nonmitochondrial” OCR | “Proton Leak” OCR | ATP Steady State Levels | ||
| Control lymphoblasts | Basal OCR | |||||||
| Correlation coefficient | 0.896 | 0.700 | 0.699 | 0.771 | 0.505 | 0.328 | ||
| Valid cases | 44 | 44 | 44 | 44 | 42 | 43 | ||
| One-sided significance | 1.09 × 10−16 | 6.31 × 10−8 | 6.47 × 10−8 | 4.61 × 10−10 | 3.23 × 10−4 | 1.58 × 10−2 | ||
| OCR by ATP synthesis | ||||||||
| Correlation coefficient | 0.925 | 0.641 | 0.656 | 0.524 | 0.336 | 0.338 | ||
| Valid cases | 33 | 44 | 44 | 44 | 42 | 43 | ||
| One-sided significance | 7.46 × 10−15 | 1.37 × 10−6 | 6.75 × 10−7 | 1.30 × 10−4 | 1.47 × 10−2 | 1.33 × 10−2 | ||
| Maximum OCR | ||||||||
| Correlation coefficient | 0.890 | 0.785 | 0.989 | 0.592 | 0.401 | 0.279 | ||
| Valid cases | 33 | 33 | 44 | 44 | 42 | 43 | ||
| One-sided significance | 2.11 × 10−12 | 3.09 × 10−8 | 1.86 × 10−36 | 1.14 × 10−5 | 4.27 × 10−3 | 3.51 × 10−2 | ||
| Complex I OCR | ||||||||
| Correlation coefficient | 0.876 | 0.789 | 0.995 | 0.570 | 0.411 | 0.297 | ||
| Valid cases | 33 | 33 | 33 | 44 | 42 | 43 | ||
| One-sided significance | 1.23 × 10−11 | 2.38 × 10−8 | 1.94 × 10−32 | 2.65 × 10−5 | 3.41 × 10−3 | 2.63 × 10−2 | ||
| “Nonmitochondrial” OCR | ||||||||
| Correlation coefficient | 0.817 | 0.633 | 0.812 | 0.777 | 0.418 | 0.188 | ||
| Valid cases | 33 | 33 | 33 | 33 | 42 | 43 | ||
| One-sided significance | 3.45 × 10−9 | 3.90 × 10−5 | 4.80 × 10−9 | 5.14 × 10−8 | 2.92 × 10−3 | 1.14 × 10−1 | ||
| “Proton leak” OCR | ||||||||
| Correlation coefficient | 0.688 | 0.521 | 0.700 | 0.697 | 0.651 | −0.014 | ||
| Valid cases | 31 | 31 | 31 | 31 | 31 | 31 | ||
| One-sided significance | 9.35 × 10−6 | 1.33 × 10−3 | 5.75 × 10−6 | 6.56 × 10−6 | 3.68 × 10−5 | 4.70 × 10−1 | ||
| ATP steady state levels | ||||||||
| Correlation coefficient | −0.055 | −0.047 | 0.054 | 0.082 | 0.059 | 0.097 | ||
| Valid cases | 33 | 33 | 33 | 33 | 33 | 41 | ||
| One-sided significance | 3.80 × 10−1 | 3.98 × 10−1 | 3.83 × 10−1 | 3.26 × 10−1 | 3.72 × 10−1 | 2.74 × 10−1 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fisher, P.R.; Allan, C.Y.; Sanislav, O.; Atkinson, A.; Ngoei, K.R.W.; Kemp, B.E.; Storey, E.; Loesch, D.Z.; Annesley, S.J. Relationships between Mitochondrial Function, AMPK, and TORC1 Signaling in Lymphoblasts with Premutation Alleles of the FMR1 Gene. Int. J. Mol. Sci. 2021, 22, 10393. https://doi.org/10.3390/ijms221910393
Fisher PR, Allan CY, Sanislav O, Atkinson A, Ngoei KRW, Kemp BE, Storey E, Loesch DZ, Annesley SJ. Relationships between Mitochondrial Function, AMPK, and TORC1 Signaling in Lymphoblasts with Premutation Alleles of the FMR1 Gene. International Journal of Molecular Sciences. 2021; 22(19):10393. https://doi.org/10.3390/ijms221910393
Chicago/Turabian StyleFisher, Paul R., Claire Y. Allan, Oana Sanislav, Anna Atkinson, Kevin R. W. Ngoei, Bruce E. Kemp, Elsdon Storey, Danuta Z. Loesch, and Sarah J. Annesley. 2021. "Relationships between Mitochondrial Function, AMPK, and TORC1 Signaling in Lymphoblasts with Premutation Alleles of the FMR1 Gene" International Journal of Molecular Sciences 22, no. 19: 10393. https://doi.org/10.3390/ijms221910393
APA StyleFisher, P. R., Allan, C. Y., Sanislav, O., Atkinson, A., Ngoei, K. R. W., Kemp, B. E., Storey, E., Loesch, D. Z., & Annesley, S. J. (2021). Relationships between Mitochondrial Function, AMPK, and TORC1 Signaling in Lymphoblasts with Premutation Alleles of the FMR1 Gene. International Journal of Molecular Sciences, 22(19), 10393. https://doi.org/10.3390/ijms221910393