
Oxytocin, Erectile Function and Sexual Behavior: Last Discoveries and Possible Advances


Abstract
1. Introduction
2. Oxytocin and Erectile Function
2.1. Oxytocin-Induced Penile Erection: Sites of Action in the Brain
2.1.1. PVN
2.1.2. PVN Oxytocinergic Neurons Are Also Activated by Other Neurotransmitters and Neuropeptides Present in the PVN Other Than by Oxytocin
2.1.3. PVN Oxytocinergic Neurons Are Also Inhibited by Neurotransmitters and Neuropeptides Present in the PVN
2.2. Ventral Tegmental Area
2.3. Hippocampus
2.4. Amygdala (Posteromedial Cortical Nucleus)
2.5. Bed Nucleus of the Stria Terminalis
2.6. Spinal Cord
3. Oxytocin and Sexual Behavior
3.1. Oxytocin and Male Sexual Behavior in Laboratory Animals
3.2. Oxytocin and Female Sexual Behavior in Laboratory Animals
4. Oxytocin, Sexual Motivation and Sexual Arousal
5. Oxytocin and Sexual Behavior in Men
6. Oxytocin and Sexual Behavior in Women
7. Oxytocin, Erectile Function and Sexual Behavior: A Synopsis
8. Final Remarks
Funding
Conflicts of Interest
References
- Pierce, J.G.; Gordon, S.; du Vigneaud, V. Further Distribution Studies on the Oxytocic Hormone of the Posterior Lobe of the Pituitary Gland and the Preparation of an Active Crystalline Flavianate. J. Biol. Chem. 1952, 199, 929–940. [Google Scholar] [CrossRef]
- Du Vigneaud, V.; Ressler, C.; Swan, J.M.; Roberts, C.W.; Katsoyannis, P.G. The Synthesis of Oxytocin1. J. Am. Chem. Soc. 1954, 76, 3115–3121. [Google Scholar] [CrossRef]
- Argiolas, A.; Gessa, G.L. Central functions of oxytocin. Neurosci. Biobehav. Rev. 1991, 15, 217–231. [Google Scholar] [CrossRef]
- Richard, P.; Moos, F.; Freund-Mercier, M.J. Central effects of oxytocin. Physiol. Rev. 1991, 71, 331–370. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.S. Oxytocin and sexual behavior. Neurosci. Biobehav. Rev. 1992, 16, 131–144. [Google Scholar] [CrossRef]
- Pedersen, C.A.; Caldwell, J.D.; Jirikowski, G.F.; Insel, T.R. Oxytocin in Maternal, Sexual and Social Behaviors; The New York Academy of Sciences: New York, NY, USA, 1992; Volume 652. [Google Scholar]
- Wagner, C.K.; Clemens, L.G. Neurophysin-containing pathway from the paraventricular nucleus of the hypothalamus to a sexually dimorphic motor nucleus in lumbar spinal cord. J. Comp. Neurol. 1993, 336, 106–116. [Google Scholar] [CrossRef]
- Tang, Y.; Rampin, O.; Calas, A.; Facchinetti, P.; Giuliano, F. Oxytocinergic and serotonergic innervation of identified lumbosacral nuclei controlling penile erection in the male rat. Neuroscience 1997, 82, 241–254. [Google Scholar] [CrossRef]
- Véronneau-Longueville, F.; Rampin, O.; Freund-Mercier, M.-J.; Tang, Y.; Calas, A.; Marson, L.; McKenna, K.; Stoeckel, M.-E.; Benoit, G.; Giuliano, F. Oxytocinergic innervation of autonomic nuclei controlling penile erection in the rat. Neuroscience 1999, 93, 1437–1447. [Google Scholar] [CrossRef]
- Dabrowska, J.; Hazra, R.; Ahern, T.H.; Guo, J.-D.; McDonald, A.J.; Mascagni, F.; Muller, J.F.; Young, L.J.; Rainnie, D.G. Neuroanatomical evidence for reciprocal regulation of the corticotrophin-releasing factor and oxytocin systems in the hypothalamus and the bed nucleus of the stria terminalis of the rat: Implications for balancing stress and affect. Psychoneuroendocrinology 2011, 36, 1312–1326. [Google Scholar] [CrossRef]
- Maejima, S.; Ohishi, N.; Yamaguchi, S.; Tsukahara, S. A neural connection between the central part of the medial preoptic nucleus and the bed nucleus of the stria terminalis to regulate sexual behavior in male rats. Neurosci. Lett. 2015, 606, 66–71. [Google Scholar] [CrossRef]
- Dumais, K.M.; Alonso, A.G.; Bredewold, R.; Veenema, A.H. Role of the oxytocin system in amygdala subregions in the regulation of social interest in male and female rats. Neuroscience 2016, 330, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Dumais, K.M.; Alonso, A.G.; Immormino, M.A.; Bredewold, R.; Veenema, A.H. Involvement of the oxytocin system in the bed nucleus of the stria terminalis in the sex-specific regulation of social recognition. Psychoneuroendocrinology 2016, 64, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Buijs, R.M. Intra- and extrahypothalamic vasopressin and oxytocin pathways in the rat. Cell Tissue Res. 1978, 192, 423–435. [Google Scholar] [CrossRef]
- Sofroniew, M. Morphology of Vasopressin and Oxytocin Neurones and Their Central and Vascular Projections. Prog. Brain Res. 1983, 60, 101–114. [Google Scholar] [CrossRef]
- Witt, D.M. Regulatory Mechanisms of Oxytocin-Mediated Sociosexual Behavior. Ann. N. Y. Acad. Sci. 1997, 807, 287–301. [Google Scholar] [CrossRef]
- Insel, T.R.; Young, L.; Wang, Z. Molecular Aspects of Monogamy. Ann. N. Y. Acad. Sci. 1997, 807, 302–316. [Google Scholar] [CrossRef] [PubMed]
- Argiolas, A.; Melis, M.R. The neuropharmacology of yawning. Eur. J. Pharmacol. 1998, 343, 1–16. [Google Scholar] [CrossRef]
- Gil, M.; Bhatt, R.; Picotte, K.B.; Hull, E.M. Oxytocin in the medial preoptic area facilitates male sexual behavior in the rat. Horm. Behav. 2011, 59, 435–443. [Google Scholar] [CrossRef]
- Gil, M.; Bhatt, R.; Picotte, K.B.; Hull, E.M. Sexual experience increases oxytocin receptor gene expression and protein in the medial preoptic area of the male rat. Psychoneuroendocrinology 2013, 38, 1688–1697. [Google Scholar] [CrossRef]
- Lee, H.-J.; Macbeth, A.H.; Pagani, J.H.; Young, W.S. Oxytocin: The great facilitator of life. Prog. Neurobiol. 2009, 88, 127–151. [Google Scholar] [CrossRef]
- Hung, L.W.; Neuner, S.; Polepalli, J.S.; Beier, K.T.; Wright, M.; Walsh, J.J.; Lewis, E.M.; Luo, L.; Deisseroth, K.; Dölen, G.; et al. Gating of social reward by oxytocin in the ventral tegmental area. Science 2017, 357, 1406–1411. [Google Scholar] [CrossRef] [PubMed]
- Sanna, F.; Bratzu, J.; Argiolas, A.; Melis, M.R. Oxytocin induces penile erection and yawning when injected into the bed nucleus of the stria terminalis: Involvement of glutamic acid, dopamine, and nitric oxide. Horm. Behav. 2017, 96, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Bratzu, J.; Bharatiya, R.; Manca, E.; Cocco, C.; Argiolas, A.; Melis, M.R.; Sanna, F. Oxytocin induces penile erection and yawning when injected into the bed nucleus of the stria terminalis: A microdialysis and immunohistochemical study. Behav. Brain Res. 2019, 375, 112147. [Google Scholar] [CrossRef] [PubMed]
- Born, J.; Lange, T.; Kern, W.; McGregor, G.P.; Bickel, U.; Fehm, H.L. Sniffing neuropeptides: A transnasal approach to the human brain. Nat. Neurosci. 2002, 5, 514–516. [Google Scholar] [CrossRef] [PubMed]
- Churchland, P.S.; Winkielman, P. Modulating social behavior with oxytocin: How does it work? What does it mean? Horm. Behav. 2012, 61, 392–399. [Google Scholar] [CrossRef]
- Leng, G.; Ludwig, M. Intranasal Oxytocin: Myths and Delusions. Biol. Psychiatry 2016, 79, 243–250. [Google Scholar] [CrossRef]
- Insel, T.R. Translating Oxytocin Neuroscience to the Clinic: A National Institute of Mental Health Perspective. Biol. Psychiatry 2016, 79, 153–154. [Google Scholar] [CrossRef] [PubMed]
- Bowen, M.T. Does peripherally administered oxytocin enter the brain? Compelling new evidence in a long-running debate. Pharmacol. Res. 2019, 146, 104325. [Google Scholar] [CrossRef]
- Smith, A.S.; Korgan, A.C.; Young, W.S. Oxytocin delivered nasally or intraperitoneally reaches the brain and plasma of normal and oxytocin knockout mice. Pharmacol. Res. 2019, 146, 104324. [Google Scholar] [CrossRef] [PubMed]
- Baumgartner, T.; Heinrichs, M.; Vonlanthen, A.; Fischbacher, U.; Fehr, E. Oxytocin Shapes the Neural Circuitry of Trust and Trust Adaptation in Humans. Neuron 2008, 58, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Insel, T.R. The Challenge of Translation in Social Neuroscience: A Review of Oxytocin, Vasopressin, and Affiliative Behavior. Neuron 2010, 65, 768–779. [Google Scholar] [CrossRef]
- Hurlemann, R.; Patin, A.; Onur, O.; Cohen, M.; Baumgartner, T.; Metzler, S.; Dziobek, I.; Gallinat, J.; Wagner, M.; Maier, W.; et al. Oxytocin Enhances Amygdala-Dependent, Socially Reinforced Learning and Emotional Empathy in Humans. J. Neurosci. 2010, 30, 4999–5007. [Google Scholar] [CrossRef]
- Graustella, A.J.; MacLeod, C. A critical review of the influence of oxytocin nasal spray on social cognition in humans: Evidence and future directions. Horm. Behav. 2012, 61, 410–418. [Google Scholar] [CrossRef]
- Piva, M.; Chang, S.W. An integrated framework for the role of oxytocin in multistage social decision-making. Am. J. Primatol. 2018, 80, e22735. [Google Scholar] [CrossRef]
- Young, L.J. When too much of a good thing is bad: Chronic oxytocin, development, and social impairments. Biol. Psychiatry 2013, 74, 160–161. [Google Scholar] [CrossRef]
- Quintana, D.S.; Alvares, G.; Hickie, I.; Guastella, A.J. Do delivery routes of intranasally administered oxytocin account for observed effects on social cognition and behavior? A two-level model. Neurosci. Biobehav. Rev. 2015, 49, 182–192. [Google Scholar] [CrossRef]
- Mierop, A.; Mikolajczak, M.; Stahl, C.; Béna, J.; Luminet, O.; Lane, A.; Corneille, O. How Can Intranasal Oxytocin Research Be Trusted? A Systematic Review of the Interactive Effects of Intranasal Oxytocin on Psychosocial Outcomes. Perspect. Psychol. Sci. 2020, 15, 1228–1242. [Google Scholar] [CrossRef] [PubMed]
- Winterton, A.; Westlye, L.T.; Steen, N.E.; Andreassen, O.A.; Quintana, D.S. Improving the precision of intranasal oxytocin research. Nat. Hum. Behav. 2021, 5, 9–18. [Google Scholar] [CrossRef] [PubMed]
- McGregor, I.S.; Bowen, M. Breaking the loop: Oxytocin as a potential treatment for drug addiction. Horm. Behav. 2012, 61, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Yamasue, H.; Yee, J.R.; Hurlemann, R.; Rilling, J.K.; Chen, F.S.; Meyer-Lindenberg, A.; Tost, H. Integrative Approaches Utilizing Oxytocin to Enhance Prosocial Behavior: From Animal and Human Social Behavior to Autistic Social Dysfunction. J. Neurosci. 2012, 32, 14109–14117. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, M.; Matsushita, H.; Tomizawa, K.; Matsui, H. Oxytocin: A therapeutic target for mental disorders. J. Physiol. Sci. 2012, 62, 441–444. [Google Scholar] [CrossRef]
- Modi, M.E.; Young, L.J. The oxytocin system in drug discovery for autism: Animal models and novel therapeutic strategies. Horm. Behav. 2012, 61, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, K.; Feifel, D. Oxytocin in schizophrenia: A review of evidence for its therapeutic effects. Acta Neuropsychiatr. 2012, 24, 130–146. [Google Scholar] [CrossRef]
- Anderberg, U.M.; Uvnäs-Moberg, K. Plasma oxytocin levels in female fibromyalgia syndrome patients. Z. Rheumatol. 2000, 59, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Ettinger, U.; Hurlemann, R.; Chan, R.C.K. Oxytocin and Schizophrenia Spectrum Disorders. Behav. Neurobiol. Psychedelic Drugs 2017, 35, 515–527. [Google Scholar] [CrossRef]
- Yamasue, H.; Domes, G. Oxytocin and Autism Spectrum Disorders. Behav. Neurobiol. Psychedelic Drugs 2017, 35, 449–465. [Google Scholar]
- Le, J.; Zhao, W.; Kou, J.; Fu, M.; Zhang, Y.; Becker, B.; Kendrick, K.M. Oxytocin facilitates socially directed attention. Psychophysiology 2021, 58, e13852. [Google Scholar] [CrossRef] [PubMed]
- De Macedo, L.R.H.; Zuardi, A.W.; Machado-De-Sousa, J.P.; Chagas, M.H.N.; Hallak, J.E. Oxytocin does not improve performance of patients with schizophrenia and healthy volunteers in a facial emotion matching task. Psychiatry Res. 2014, 220, 125–128. [Google Scholar] [CrossRef]
- Mameli, S.; Pisanu, G.M.; Sardo, S.; Marchi, A.; Pili, A.; Carboni, M.; Minerba, L.; Trincas, G.; Carta, M.G.; Melis, M.R.; et al. Oxytocin nasal spray in fibromyalgic patients. Rheumatol. Int. 2014, 34, 1047–1052. [Google Scholar] [CrossRef]
- Agabio, R.; Farci, A.M.G.; Curreli, O.; Deidda, R.; Mercuro, S.; Naitana, R.; Restivo, A.; Tronci, E.; Melis, G.L.G.A.M.R. Oxytocin Nasal Spray in the Treatment of Binge Eating Disorder and Obesity: A Pilot, Randomized, Double-Blind Trial. Clin. Pharmacol. Biopharm. 2016, 5, 155. [Google Scholar] [CrossRef]
- Agabio, R.; Mameli, S.; Sardo, S.; Minerba, L.; Melis, M.R. Oxytocin nasal spray in fibromyalgic patients: Additional information. Rheumatol. Int. 2014, 34, 1335–1336. [Google Scholar] [CrossRef][Green Version]
- Melin, P.; Kihlström, J.E. Influence of Oxytocin on Sexual Behavior in Male Rabbits. Endocrinology 1963, 73, 433–435. [Google Scholar] [CrossRef]
- Lidberg, L. The effect of Synthocinon R on patients suffering from impotence. Pharmacopsychiatry 1972, 5, 187–190. [Google Scholar] [CrossRef]
- Carmichael, M.S.; Humbert, R.; Dixen, J.; Palmisano, G.; Greenleaf, W.; Davidson, J.M. Plasma Oxytocin Increases in the Human Sexual Response. J. Clin. Endocrinol. Metab. 1987, 64, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.R.; Seckl, J.R.; Burton, S.; Checkley, S.A.; Lightman, S.L. Changes in Oxytocin and Vasopressin Secretion During Sexual Activity in Men. J. Clin. Endocrinol. Metab. 1987, 65, 738–741. [Google Scholar] [CrossRef] [PubMed]
- Tindall, J.S. Stimuli that cause the release of oxytocin. In Handbook of Physiology. Section 7, Endocrinology; Geiger, S.R., Knobil, E., Sawyer, W.H., Greef, R., Astwood, E.B., Eds.; American Physiology Society: Washington, DC, USA, 1974; Volume IV, pp. 257–267. [Google Scholar]
- Argiolas, A.; Melis, M.; Gessa, G. Intraventricular oxytoxin induces yawning and penile erection in rats. Eur. J. Pharmacol. 1985, 117, 395–396. [Google Scholar] [CrossRef]
- Argiolas, A.; Melis, M.R.; Gessa, G.L. Oxytocin: An extremely potent inducer of penile erection and yawning in male rats. Eur. J. Pharmacol. 1986, 130, 265–272. [Google Scholar] [CrossRef]
- Arletti, R.; Bazzani, C.; Castelli, M.; Bertolini, A. Oxytocin improves male copulatory performance in rats. Horm. Behav. 1985, 19, 14–20. [Google Scholar] [CrossRef]
- Arletti, R.; Bertolini, A. Oxytocin stimulates lordosis behavior in female rats. Neuropeptides 1985, 6, 247–253. [Google Scholar] [CrossRef]
- Caldwell, J.; Prange, A.; Pedersen, C. Oxytocin facilitates the sexual receptivity of estrogen-treated female rats. Neuropeptides 1986, 7, 175–189. [Google Scholar] [CrossRef]
- Argiolas, A.; Melis, M.; Stancampiano, R.; Gessa, G. Penile erection and yawning induced by oxytocin and related peptides: Structure-activity relationship. Peptides 1989, 10, 559–563. [Google Scholar] [CrossRef]
- Argiolas, A.; Melis, M.R. Neuromodulation of penile erection: An overview of the role of neurotransmitters and neuropeptides. Prog. Neurobiol. 2015, 47, 235–255. [Google Scholar] [CrossRef]
- Argiolas, A.; Melis, M.R. The role of oxytocin and the paraventricular nucleus in the sexual behaviour of male mammals. Physiol. Behav. 2004, 83, 309–317. [Google Scholar] [CrossRef]
- Argiolas, A.; Melis, M.R. Central control of penile erection: Role of the paraventricular nucleus of the hypothalamus. Prog. Neurobiol. 2005, 76, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Argiolas, A.; Melis, M.R. Neuropeptides and central control of sexual behaviour from the past to the present: A review. Prog. Neurobiol. 2013, 108, 80–107. [Google Scholar] [CrossRef]
- Argiolas, A. Neuropeptides and sexual behaviour. Neurosci. Biobehav. Rev. 1999, 23, 1127–1142. [Google Scholar] [CrossRef]
- Gimpl, G.; Fahrenholz, F. The Oxytocin Receptor System: Structure, Function, and Regulation. Physiol. Rev. 2001, 81, 629–683. [Google Scholar] [CrossRef]
- Jurek, B.; Neumann, I.D. The Oxytocin Receptor: From Intracellular Signaling to Behavior. Physiol. Rev. 2018, 98, 1805–1908. [Google Scholar] [CrossRef] [PubMed]
- Melis, M.R.; Mauri, A.; Argiolas, A. Apomorphine- and Oxytocin-Induced Penile Erection and Yawning in Intact and Castrated Male Rats: Effect of Sexual Steroids. Neuroendocrinology 1994, 59, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Sachs, B.D.; Meisel, R.L. The physiology of male sexual behavior. In The Physiology of Reproduction, 1st ed.; Knobil, E., Neil, J., Eds.; Raven Press: New York, NY, USA, 1988; Volume 2, pp. 1393–1485. [Google Scholar]
- Meisel, R.L.; Sachs, B.D. The physiology of male sexual behaviour. In The Physiology of Reproduction, 2nd ed.; Knobil, E., Neil, J., Eds.; Raven Press: New York, NY, USA, 1994; Volume 2, pp. 3–96. [Google Scholar]
- Hull, E.M.; Meisel, R.L.; Sachs, B.D. Male sexual behavior. In Hormones, Brain and Behavior; Pfaff, D.W., Arnold, A.P., Etgen, A.M., Fahrbach, S.E., Rubin, R.T., Eds.; Academic Press: New York, NY, USA, 2002; pp. 3–137. [Google Scholar]
- Argiolas, A. Male erectile dysfunction: Chemical pharmacology of penile erection. Drug Discov. Today Ther. Strat. 2005, 2, 31–36. [Google Scholar] [CrossRef]
- Andersson, K.E.; Wagner, G. Physiology of penile erection. Physiol. Rev. 1995, 75, 191–236. [Google Scholar] [CrossRef] [PubMed]
- McKenna, K.E. Central control of penile erection. Int. J. Impot. Res. 1998, 10, 25–34. [Google Scholar]
- McKenna, K. Some proposals regarding the organization of the central nervous system control of penile erection. Neurosci. Biobehav. Rev. 2000, 24, 535–540. [Google Scholar] [CrossRef]
- Sachs, B.D. Contextual approaches to the physiology and classification of erectile function, erectile dysfunction, and sexual arousal. Neurosci. Biobehav. Rev. 2000, 24, 541–560. [Google Scholar] [CrossRef]
- Giuliano, F.; Rampin, O. Central neural regulation of penile erection. Neurosci. Biobehav. Rev. 2000, 24, 517–533. [Google Scholar] [CrossRef]
- Giuliano, F.; Rampin, O. Neural control of erection. Physiol. Behav. 2004, 83, 189–201. [Google Scholar] [CrossRef]
- Andersson, K.-E. Mechanisms of Penile Erection and Basis for Pharmacological Treatment of Erectile Dysfunction. Pharmacol. Rev. 2011, 63, 811–859. [Google Scholar] [CrossRef] [PubMed]
- Chitaley, K.; Wingard, C.; Webb, R.C.; Branam, H.; Stopper, V.S.; Lewis, R.W.; Mills, T.M. Antagonism of Rho-kinase stimulates rat penile erection via a nitric oxide-independent pathway. Nat. Med. 2001, 7, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Giuliano, F.; Allard, J. Dopamine and sexual function. Int. J. Impotence Res. 2001, 13 (Suppl. S3), 18–28. [Google Scholar] [CrossRef]
- Melis, M.R.; Argiolas, M.R.M.A.A. Central Oxytocinergic Neurotransmission: A Drug Target for the Therapy of Psychogenic Erectile Dysfunction. Curr. Drug Targets 2003, 4, 55–66. [Google Scholar] [CrossRef]
- Heaton, J.P. Central neuropharmacological agents and mechanisms in erectile dysfunction: The role of dopamine. Neurosci. Biobehav. Rev. 2000, 24, 561–569. [Google Scholar] [CrossRef]
- Melis, M.; Argiolas, A.; Gessa, G. Oxytocin-induced penile erection and yawning: Site of action in the brain. Brain Res. 1986, 398, 259–265. [Google Scholar] [CrossRef]
- Melis, M.R.; Melis, T.; Cocco, C.; Succu, S.; Sanna, F.; Pillolla, G.; Boi, A.; Ferri, G.-L.; Argiolas, A. Oxytocin injected into the ventral tegmental area induces penile erection and increases extracellular dopamine in the nucleus accumbens and paraventricular nucleus of the hypothalamus of male rats. Eur. J. Neurosci. 2007, 26, 1026–1035. [Google Scholar] [CrossRef] [PubMed]
- Melis, M.R.; Succu, S.; Sanna, F.; Boi, A.; Argiolas, A. Oxytocin injected into the ventral subiculum or the posteromedial cortical nucleus of the amygdala induces penile erection and increases extracellular dopamine levels in the nucleus accumbens of male rats. Eur. J. Neurosci. 2009, 30, 1349–1357. [Google Scholar] [CrossRef] [PubMed]
- Melis, M.R.; Argiolas, A. Central control of penile erection: A re-visitation of the role of oxytocin and its interaction with dopamine and glutamic acid in male rats. Neurosci. Biobehav. Rev. 2011, 35, 939–955. [Google Scholar] [CrossRef]
- Freund-Mercier, M.J.; Stoeckel, M.E. Somatodendritic autoreceptors on oxytocin neurones. Single Mol. Single Cell Seq. 1995, 395, 185–194. [Google Scholar]
- Freund-Mercier, M.; Richard, P. Excitatory effects of intraventricular injections of oxytocin on the milk ejection reflex in the rat. Neurosci. Lett. 1981, 23, 193–198. [Google Scholar] [CrossRef]
- Freund-Mercier, M.J.; Richard, P. Electrophysiological evidence for facilitatory control of oxytocin neurones by oxytocin during suckling in the rat. J. Physiol. 1984, 352, 447–466. [Google Scholar] [CrossRef] [PubMed]
- Moos, F.; Freund-Mercier, M.J.; Guerné, Y.; Guerné, J.M.; Stoeckel, M.E.; Richard, P. Release of oxytocin and vasopressin by magnocellular nuclei in vitro: Specific facilitatory effect of oxytocin on its own release. J. Endocrinol. 1984, 102, 63–72. [Google Scholar] [CrossRef]
- Yamashita, H.; Okuya, S.; Inenaga, K.; Kasai, M.; Uesugi, S.; Kannan, H.; Kaneko, T. Oxytocin predominantly excites putative oxytocin neurons in the rat supraoptic nucleus in vitro. Brain Res. 1987, 416, 364–368. [Google Scholar] [CrossRef]
- Theodosis, D.T. Oxytocin-immunoreactive terminals synapse on oxytocin neurones in the supraoptic nucleus. Nat. Cell Biol. 1985, 313, 682–684. [Google Scholar] [CrossRef]
- Argiolas, A.; Melis, M.; Mauri, A.; Gessa, G. Paraventricular nucleus lesion prevents yawning and penile erection induced by apomorphine and oxytocin but not by ACTH in rats. Brain Res. 1987, 421, 349–352. [Google Scholar] [CrossRef]
- Liu, Y.-C.; Salamone, J.; Sachs, B. Lesions in Medial Preoptic Area and Bed Nucleus of Stria Terminalis: Differential Effects on Copulatory Behavior and Noncontact Erection in Male Rats. J. Neurosci. 1997, 17, 5245–5253. [Google Scholar] [CrossRef] [PubMed]
- Sachs, B.D. Erection Evoked in Male Rats by Airborne Scent from Estrous Females. Physiol. Behav. 1997, 62, 921–924. [Google Scholar] [CrossRef]
- Edwards, D.A.; Davis, A.B. Deafferentation of the Olfactory Bulbs of Male Rats Reduces Erection to Remote Cues from Females. Physiol. Behav. 1997, 62, 145–149. [Google Scholar] [CrossRef]
- Melis, M.R.; Succu, S.; Mascia, M.S.; Cortis, L.; Argiolas, A. Extra-cellular dopamine increases in the paraventricular nucleus of male rats during sexual activity. Eur. J. Neurosci. 2003, 17, 1266–1272. [Google Scholar] [CrossRef]
- Argiolas, A.; Melis, M.R.; Vargiu, L.; Gessa, G.L. d(CH2)5Tyr(Me)-[Orn8]vasotocin, a potent oxytocin antagonist, antagonizes penile erection and yawning induced by oxytocin and apomorphine, but not by ACTH-(1–24). Eur. J. Pharmacol. 1987, 134, 221–224. [Google Scholar] [CrossRef]
- Melis, M.R.; Spano, M.S.; Succu, S.; Argiolas, A. The oxytocin antagonist d(CH2)5Tyr(Me)2-Orn8-vasotocin reduces non-contact penile erections in male rats. Neurosci. Lett. 1999, 265, 171–174. [Google Scholar] [CrossRef]
- Vincent, S.; Kimura, H. Histochemical mapping of nitric oxide synthase in the rat brain. Neuroscience 1992, 46, 755–784. [Google Scholar] [CrossRef]
- Torres, G.; Lee, S.; Rivier, C. Ontogeny of the Rat Hypothalamic Nitric Oxide Synthase and Colocalization with Neuropeptides. Mol. Cell. Neurosci. 1993, 4, 155–163. [Google Scholar] [CrossRef]
- Melis, M.; Stancampiano, R.; Argiolas, A. Penile erection and yawning induced by paraventricular NMDA injection in male rats are mediated by oxytocin. Pharmacol. Biochem. Behav. 1994, 48, 203–207. [Google Scholar] [CrossRef]
- Melis, M.R.; Succu, S.; Iannucci, U.; Argiolas, A. Oxytocin increases nitric oxide production in the paraventricular nucleus of the hypothalamus of male rats: Correlation with penile erection and yawning. Regul. Pept. 1997, 69, 105–111. [Google Scholar] [CrossRef]
- Bańkowski, K.; Manning, M.; Seto, J.; Haldar, J.; Sawyer, W.H. Design and synthesis of potent in vivo antagonists of oxytocin. Int. J. Pept. Protein Res. 2009, 16, 382–391. [Google Scholar] [CrossRef] [PubMed]
- McCleskey, E.W.; Fox, A.P.; Feldman, D.H.; Cruz, L.J.; Olivera, B.M.; Tsien, R.W.; Yoshikami, D. Omega-conotoxin: Direct and persistent blockade of specific types of calcium channels in neurons but not muscle. Proc. Natl. Acad. Sci. USA 1987, 84, 4327–4331. [Google Scholar] [CrossRef] [PubMed]
- Argiolas, A.; Melis, M.; Stancampiano, R.; Gessa, G. ω-Conotoxin prevents apomorphine- and oxytocin-induced penile erection and yawning in male rats. Pharmacol. Biochem. Behav. 1990, 37, 253–257. [Google Scholar] [CrossRef]
- Succu, S.; Spano, M.S.; Melis, M.R.; Argiolas, A. Different effects of ω-conotoxin on penile erection, yawning and paraventricular nitric oxide in male rats. Eur. J. Pharmacol. 1998, 359, 19–26. [Google Scholar] [CrossRef]
- Argiolas, A. Nitric oxide is a central mediator of penile erection. Neuropharmacology 1994, 33, 1339–1344. [Google Scholar] [CrossRef]
- Melis, M.; Stancampiano, R.; Argiolas, A. Prevention by NG-nitro-L-arginine methyl ester of apomorphine- and oxytocin-induced penile erection and yawning: Site of action in the brain. Pharmacol. Biochem. Behav. 1994, 48, 799–804. [Google Scholar] [CrossRef]
- Melis, M.; Stancampiano, R.; Argiolas, A. Nitric oxide synthase inhibitors prevent N-methyl-d-aspartic acid-induced penile erection and yawning in male rats. Neurosci. Lett. 1994, 179, 9–12. [Google Scholar] [CrossRef]
- Melis, M.; Stancampiano, R.; Lai, C.; Argiolas, A. Nitroglycerin-induced penile erection and yawning in male rats: Mechanism of action in the brain. Brain Res. Bull. 1995, 36, 527–531. [Google Scholar] [CrossRef]
- Melis, M.R.; Argiolas, A. Nitric oxide donors induce penile erection and yawning when injected in the central nervous system of male rats. Eur. J. Pharmacol. 1995, 294, 1–9. [Google Scholar] [CrossRef]
- Melis, M.R.; Succu, S.; Mauri, A.; Argiolas, A. Nitric oxide production is increased in the paraventricular nucleus of the hypothalamus of male rats during non-contact penile erections and copulation. Eur. J. Neurosci. 1998, 10, 1968–1974. [Google Scholar] [CrossRef] [PubMed]
- Benelli, A.; Bertolini, A.; Poggioli, R.; Cavazzuti, E.; Calzà, L.; Giardino, L.; Arletti, R. Nitric oxide is involved in male sexual behavior of rats. Eur. J. Pharmacol. 1995, 294, 505–510. [Google Scholar] [CrossRef]
- Arletti, R.; Calza, L.; Giardino, L.; Benelli, A.; Cavazzuti, E.; Bertolini, A. Sexual impotence is associated with a reduced production of oxytocin and with an increased production of opioid peptides in the paraventricular nucleus of male rats. Neurosci. Lett. 1997, 233, 65–68. [Google Scholar] [CrossRef]
- Baskerville, T.; Douglas, A. Interactions between dopamine and oxytocin in the control of sexual behaviour. Prog. Brain Res. 2008, 170, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Baskerville, T.A.; Allard, J.; Wayman, C.; Douglas, A.J. Dopamine–oxytocin interactions in penile erection. Eur. J. Neurosci. 2009, 30, 2151–2164. [Google Scholar] [CrossRef] [PubMed]
- Sanna, F.; Succu, S.; Hübner, H.; Gmeiner, P.; Argiolas, A.; Melis, M.R. Dopamine D2-like receptor agonists induce penile erection in male rats: Differential role of D2, D3 and D4 receptors in the paraventricular nucleus of the hypothalamus. Behav. Brain Res. 2011, 225, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Sanna, F.; Succu, S.; Melis, M.R.; Argiolas, A. Dopamine agonist-induced penile erection and yawning: Differential role of D2-like receptor subtypes and correlation with nitric oxide production in the paraventricular nucleus of the hypothalamus of male rats. Behav. Brain Res. 2012, 230, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Sanna, F.; Contini, A.; Melis, M.R.; Argiolas, A. Role of dopamine D4 receptors in copulatory behavior: Studies with selective D4 agonists and antagonists in male rats. Pharmacol. Biochem. Behav. 2015, 137, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Sanna, F.; Melis, M.R.; Argiolas, A. Dopamine, erectile function and sexual behavior: Last discoveries and possible advances. In New Developments in Dopamine Research; Hopkins, G., Ed.; Nova Publishers Inc.: New York, NY, USA, 2016; pp. 45–84. [Google Scholar]
- Collins, G.T.; Truccone, A.; Haji-Abdi, F.; Newman, A.H.; Grundt, P.; Rice, K.C.; Husbands, S.M.; Greedy, B.M.; Enguehard-Gueiffier, C.; Gueiffier, A.; et al. Proerectile Effects of Dopamine D2-Like Agonists Are Mediated by the D3 Receptor in Rats and Mice. J. Pharmacol. Exp. Ther. 2009, 329, 210–217. [Google Scholar] [CrossRef]
- Depoortère, R.; Bardin, L.; Rodrigues, M.; Abrial, E.; Aliaga, M.; Newman-Tancredi, A. Penile erection and yawning induced by dopamine D2-like receptor agonists in rats: Influence of strain and contribution of dopamine D2, but not D3 and D4 receptors. Behav. Pharmacol. 2009, 20, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Melis, M.R.; Succu, S.; Iannucci, U.; Argiolas, A. N-Methyl-d-aspartic acid-induced penile erection and yawning: Role of hypothalamic paraventricular nitric oxide. Eur. J. Pharmacol. 1997, 328, 115–123. [Google Scholar] [CrossRef]
- Melis, M.R.; Succu, S.; Spano, M.S.; Argiolas, A. Effect of excitatory amino acid, dopamine, and oxytocin receptor antagonists on noncontact penile erections and paraventricular nitric oxide production in male rats. Behav. Neurosci. 2000, 114, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.-K.; Chang, L.S. Effect of excitatory amino acid receptor agonists on penile erection after administration into paraventricular nucleus of hypothalamus in the rat. Urology 2003, 62, 575–580. [Google Scholar] [CrossRef]
- Zahran, A.R.; Vachon, P.; Courtois, F.; Carrier, S. Increases in intracavernous penile pressure following injections of excitatory amino acid receptor agonists in the hypothalamic paraventricular nucleus of anesthetized rats. J. Urol. 2000, 164, 1793–1797. [Google Scholar] [CrossRef]
- Melis, M.R.; Succu, S.; Spano, M.S.; Locatelli, V.; Torsello, A.; Muller, E.E.; Deghenghi, R.; Argiolas, A. EP 60761 and EP 50885, two hexarelin analogues, induce penile erection in rats. Eur. J. Pharmacol. 2000, 404, 137–143. [Google Scholar] [CrossRef]
- Melis, M.R.; Spano, M.; Succu, S.; Locatelli, V.; Torsello, A.; Muller, E.; Deghenghi, R.; Argiolas, A. EP 60761- and EP 50885-induced penile erection: Structure-activity studies and comparison with apomorphine, oxytocin and N-methyl-D-aspartic acid. Int. J. Impot. Res. 2000, 12, 255–262. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Torsello, A.; Locatelli, V.; Melis, M.R.; Succu, S.; Spano, M.S.; Deghenghi, R.; Müller, E.E.; Argiolas, A. Differential orexigenic effects of hexarelin and its analogs in the rat hypothalamus: Indication for multiple growth hormone secretagogue receptor subtypes. Neuroendocrinology 2000, 72, 327–332. [Google Scholar] [CrossRef]
- Melis, M.R.; Succu, S.; Spano, M.S.; Torsello, A.; Locatelli, V.; Muller, E.E.; Deghenghi, R.; Argiolas, A. Penile erection induced by EP 80661 and other hexarelin peptide analogues: Involvement of paraventricular nitric oxide. Eur. J. Pharmacol. 2001, 411, 305–310. [Google Scholar] [CrossRef]
- Melis, M.R.; Succu, S.; Spano, M.S.; Deghenghi, R.; Argiolas, A. EP 91073 prevents EP 80661-induced penile erection: New evidence for the existence of specific EP peptide receptors mediating penile erection. Neuropharmacology 2001, 41, 254–262. [Google Scholar] [CrossRef]
- Trani, E.; Giorgi, A.; Canu, N.; Amadoro, G.; Rinaldi, A.M.; Halban, P.A.; Ferri, G.L.; Possenti, R.; Schininà†, M.E.; Levi, A. Isolation and characterization of VGF peptides in rat brain. Role of PC1/3 and PC2 in the maturation of VGF precursor. J. Neurochem. 2002, 81, 565–574. [Google Scholar] [CrossRef]
- Succu, S.; Cocco, C.; Mascia, M.S.; Melis, T.; Melis, M.R.; Ferri, G.L.; Possenti, R.; Levi, A.; Argiolas, A. Pro-VGF-derived peptides induce penile erection in male rats: Possible involvement of oxytocin. Eur. J. Neurosci. 2004, 20, 3035–3040. [Google Scholar] [CrossRef] [PubMed]
- Succu, S.; Mascia, M.S.; Melis, T.; Sanna, F.; Melis, M.R.; Possenti, R.; Argiolas, A. Pro-VGF-derived peptides induce penile erection in male rats: Involvement of paraventricular nitric oxide. Neuropharmacology 2005, 49, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, H.; Sasaki, K.; Satomi, Y.; Shimbara, T.; Kageyama, H.; Mondal, M.S.; Toshinai, K.; Date, Y.; González, L.J.; Shioda, S.; et al. Peptidomic Identification and Biological Validation of Neuroendocrine Regulatory Peptide-1 and -2. J. Biol. Chem. 2007, 282, 26354–26360. [Google Scholar] [CrossRef]
- Melis, M.R.; Sanna, F.; Succu, S.; Ferri, G.L.; Argiolas, A. Neuroendocrine regulatory peptide-1 and neuroendocrine regulatory peptide-2 influence differentially feeding and penile erection in male rats: Sites of action in the brain. Regul. Pept. 2012, 177, 46–52. [Google Scholar] [CrossRef]
- Melis, M.R.; Spano, M.S.; Succu, S.; Argiolas, A. Activation of γ-aminobutyric acidA receptors in the paraventricular nucleus of the hypothalamus reduces apomorphine-, N-methyl-d-aspartic acid- and oxytocin-induced penile erection and yawning in male rats. Neurosci. Lett. 2000, 281, 127–130. [Google Scholar] [CrossRef]
- Melis, M.R.; Succu, S.; Mascia, M.S.; Argiolas, A. The activation of γ aminobutyric acidA receptors in the paraventricular nucleus of the hypothalamus reduces non-contact penile erections in male rats. Neurosci. Lett. 2001, 314, 123–126. [Google Scholar] [CrossRef]
- Boudaba, C.; Szabó, K.; Tasker, J.G. Physiological Mapping of Local Inhibitory Inputs to the Hypothalamic Paraventricular Nucleus. J. Neurosci. 1996, 16, 7151–7160. [Google Scholar] [CrossRef]
- Roland, B.L.; Sawchenko, P.E. Local origins of some GABAergic projections to the paraventricular and supraoptic nuclei of the hypothalamus in the rat. J. Comp. Neurol. 1993, 332, 123–143. [Google Scholar] [CrossRef] [PubMed]
- Melis, M.R.; Argiolas, A. Reduction of drug-induced yawning and penile erection and of noncontact erections in male rats by the activation of GABAA receptors in the paraventricular nucleus: Involvement of nitric oxide. Eur. J. Neurosci. 2002, 15, 852–860. [Google Scholar] [CrossRef]
- Pfaus, J.; Gorzalka, B.B. Opioids and sexual behavior. Neurosci. Biobehav. Rev. 1987, 11, 1–34. [Google Scholar] [CrossRef]
- Pfaus, J.G.; Everitt, B.J. The psychopharmacology of sexual behavior. In Psychopharmacology: The Fourth Generation of Progress; Knobil, F.E., Kupfer, D.J., Eds.; Raven Press: New York, NY, USA, 1995; pp. 742–758. [Google Scholar]
- Cushman, P. Sexual behavior in heroin addiction and methadone maintenance. Correlation with plasma luteinizing hormone. N. Y. State J. Med. 1972, 72, 1261–1265. [Google Scholar] [PubMed]
- Melis, M.R.; Stancampiano, R.; Gessa, G.L.; Argiolas, A. Prevention by morphine of apomorphine- and oxytocin-induced penile erection and yawning: Site of action in the brain. Neuropsychopharmacology 1992, 6, 17–21. [Google Scholar]
- Melis, M.R.; Succu, S.; Argiolas, A. Prevention by Morphine of N-Methyl-d-Aspartic Acid-Induced Penile Erection and Yawning: Involvement of Nitric Oxide. Brain Res. Bull. 1997, 44, 689–694. [Google Scholar] [CrossRef]
- Melis, M.R.; Succu, S.; Iannucci, U.; Argiolas, A. Prevention by morphine of apomorphine- and oxytocin-induced penile erection and yawning: Involvement of nitric oxide. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1997, 355, 595–600. [Google Scholar] [CrossRef]
- Rinaldi-Carmona, M.; Barth, F.; Héaulme, M.; Shire, D.; Calandra, B.; Congy, C.; Martinez, S.; Maruani, J.; Néliat, G.; Caput, D.; et al. SR141716A, a potent and selective antagonist of the brain cannabinoid receptor. FEBS Lett. 1994, 350, 240–244. [Google Scholar] [CrossRef]
- Succu, S.; Mascia, M.S.; Melis, T.; Sanna, F.; Boi, A.; Melis, M.R.; Argiolas, A. Morphine reduces penile erection induced by the cannabinoid receptor antagonist SR 141617A in male rats: Role of paraventricular glutamic acid and nitric oxide. Neurosci. Lett. 2006, 404, 1–5. [Google Scholar] [CrossRef]
- Melis, M.R.; Succu, S.; Spano, M.S.; Argiolas, A. Morphine injected into the paraventricular nucleus of the hypothalamus prevents noncontact penile erections and impairs copulation: Involvement of nitric oxide. Eur. J. Neurosci. 1999, 11, 1857–1864. [Google Scholar] [CrossRef]
- Melis, M.R.; Succu, S.; Mascia, M.S.; Argiolas, A. Antagonism of cannabinoid CB1 receptors in the paraventricular nucleus of male rats induces penile erection. Neurosci. Lett. 2004, 359, 17–20. [Google Scholar] [CrossRef]
- Melis, M.R.; Succu, S.; Mascia, M.S.; Sanna, F.; Melis, T.; Castelli, M.P.; Argiolas, A. The cannabinoid receptor antagonist SR-141716A induces penile erection in male rats: Involvement of paraventricular glutamic acid and nitric oxide. Neuropharmacology 2006, 50, 219–228. [Google Scholar] [CrossRef]
- Succu, S.; Mascia, M.S.; Sanna, F.; Melis, T.; Argiolas, A.; Melis, M.R. The cannabinoid CB1 receptor antagonist SR 141716A induces penile erection by increasing extra-cellular glutamic acid in the paraventricular nucleus of male rats. Behav. Brain Res. 2006, 169, 274–281. [Google Scholar] [CrossRef]
- Woodruff, G.N. The interaction between MK-801 and receptors for: Functional consequences. Neuropharmacology 1987, 26, 903–909. [Google Scholar] [CrossRef]
- Castelli, M.P.; Piras, A.; Melis, T.; Succu, S.; Sanna, F.; Melis, M.; Collu, S.; Ennas, M.G.; Diaz, G.; Mackie, K.; et al. Cannabinoid CB1 receptors in the paraventricular nucleus and central control of penile erection: Immunocytochemistry, autoradiography and behavioral studies. Neuroscience 2007, 147, 197–206. [Google Scholar] [CrossRef]
- Freund-Mercier, M.; Stoeckel, M.; Palacios, J.; Pazos, A.; Reichhart, J.; Porte, A.; Richard, P. Pharmacological characteristics and anatomical distribution of [3H]oxytocin-binding sites in the wistar rat brain studied by autoradiography. Neuroscience 1987, 20, 599–614. [Google Scholar] [CrossRef]
- Adan, A.R.; Van Leeuwen, F.W.; Sonnemans, A.M.; Brouns, M.; Hoffman, G.; Verbalis, J.G.; Burbach, J.P. Rat oxytocin receptor in brain, pituitary, mammary gland, and uterus: Partial sequence and immunocytochemical localization. Endocrinology 1995, 136, 4022–4028. [Google Scholar] [CrossRef]
- Vaccari, C.; Lolait, S.J.; Ostrowski, N.L. Comparative Distribution of Vasopressin V1b and Oxytocin Receptor Messenger Ribonucleic Acids in Brain1. Endocrinology 1998, 139, 5015–5033. [Google Scholar] [CrossRef] [PubMed]
- Loup, F.; Tribollet, E.; Dubois-Dauphin, M.; Pizzolato, G.; Dreifuss, J. Localization of oxytocin binding sites in the human brainstem and upper spinal cord: An autoradiographic study. Brain Res. 1989, 500, 223–230. [Google Scholar] [CrossRef]
- Loup, F.; Tribollet, E.; Dubois-Dauphin, M.; Dreifuss, J. Localization of high-affinity binding sites for oxytocin and vasopressin in the human brain. An autoradiographic study. Brain Res. 1991, 555, 220–232. [Google Scholar] [CrossRef]
- Boccia, M.; Petrusz, P.; Suzuki, K.; Marson, L.; Pedersen, C. Immunohistochemical localization of oxytocin receptors in human brain. Neuroscience 2013, 253, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Melis, M.R.; Sanna, F.; Succu, S.; Zarone, P.; Boi, A.; Argiolas, A. The role of oxytocin in the anticipatory and consummatory phases of male rat sexual behaviour. In Handbook of Oxytocin Research: Synthesis, Storage and Release, Actions and Drug Forms; Jastrow, H., Feuerbach, D., Eds.; Nova Publishers Inc.: New York, NY, USA, 2009; pp. 109–125. [Google Scholar]
- Succu, S.; Sanna, F.; Cocco, C.; Melis, T.; Boi, A.; Ferri, G.-L.; Argiolas, A.; Melis, M.R. Oxytocin induces penile erection when injected into the ventral tegmental area of male rats: Role of nitric oxide and cyclic GMP. Eur. J. Neurosci. 2008, 28, 813–821. [Google Scholar] [CrossRef]
- Garthwaite, J.; Southam, E.; Boulton, C.L.; Nielsen, E.B.; Schmidt, K.; Mayer, B. Potent and selective inhibition of nitric ox-ide-sensitive guanylyl cyclase by 1H-[1,2,4]oxadiazolo[4,3-a]quinoxalin-1-one. Mol. Pharmacol. 1995, 48, 184–188. [Google Scholar] [PubMed]
- Sanna, F.; Succu, S.; Boi, A.; Melis, M.R.; Argiolas, A. Phosphodiesterase Type 5 Inhibitors Facilitate Noncontact Erections in Male Rats: Site of Action in the Brain and Mechanism of Action. J. Sex. Med. 2009, 6, 2680–2689. [Google Scholar] [CrossRef] [PubMed]
- Succu, S.; Sanna, F.; Melis, T.; Boi, A.; Argiolas, A.; Melis, M.R. Stimulation of dopamine receptors in the paraventricular nucleus of the hypothalamus of male rats induces penile erection and increases extra-cellular dopamine in the nucleus accumbens: Involvement of central oxytocin. Neuropharmacology 2007, 52, 1034–1043. [Google Scholar] [CrossRef]
- Peris, J.; MacFadyen, K.; Smith, J.A.; de Kloet, A.; Wang, L.; Krause, E.G. Oxytocin receptors are expressed on dopamine and glutamate neurons in the mouse ventral tegmental area that project to nucleus accumbens and other mesolimbic targets. J. Comp. Neurol. 2017, 525, 1094–1108. [Google Scholar] [CrossRef]
- Fanselow, M.S.; Dong, H.-W. Are the Dorsal and Ventral Hippocampus Functionally Distinct Structures? Neuron 2010, 65, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.-K.; Chan, J.Y.; Chang, L.S.; Chen, M.-T.; Chan, S.H. Elicitation of penile erection following activation of the hippocampal formation in the rat. Neurosci. Lett. 1992, 141, 218–222. [Google Scholar] [CrossRef]
- Melis, M.; Argiolas, A.; Stancampiano, R.; Gessa, G. Effect of apomorphine on oxytocin concentrations in different brain areas and plasma of male rats. Eur. J. Pharmacol. 1990, 182, 101–107. [Google Scholar] [CrossRef]
- Furtak, S.C.; Wei, S.-M.; Agster, K.L.; Burwell, R.D. Functional neuroanatomy of the parahippocampal region in the rat: The perirhinal and postrhinal cortices. Hippocampus 2007, 17, 709–722. [Google Scholar] [CrossRef]
- Melis, M.; Stancampiano, R.; Argiolas, A. Hippocampal oxytocin mediates apomorphine-induced penile erection and yawning. Pharmacol. Biochem. Behav. 1992, 42, 61–66. [Google Scholar] [CrossRef]
- Kondo, Y.; Sachs, B. Disparate effects of small medial amygdala lesions on noncontact erection, copulation, and partner preference. Physiol. Behav. 2002, 76, 443–447. [Google Scholar] [CrossRef]
- Boccia, M.L.; Goursaud, A.-P.S.; Bachevalier, J.; Anderson, K.D.; Pedersen, C.A. Peripherally administered non-peptide oxytocin antagonist, L368,899®, accumulates in limbic brain areas: A new pharmacological tool for the study of social motivation in non-human primates. Horm. Behav. 2007, 52, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Melis, M.R.; Succu, S.; Cocco, C.; Caboni, E.; Sanna, F.; Boi, A.; Ferri, G.L.; Argiolas, A. Oxytocin induces penile erection when injected into the ventral subiculum: Role of nitric oxide and glutamic acid. Neuropharmacology 2010, 58, 1153–1160. [Google Scholar] [CrossRef] [PubMed]
- Succu, S.; Sanna, F.; Argiolas, A.; Melis, M.R. Oxytocin injected into the hippocampal ventral subiculum induces penile erection in male rats by increasing glutamatergic neurotransmission in the ventral tegmental area. Neuropharmacology 2011, 61, 181–188. [Google Scholar] [CrossRef]
- Uhl-Bronner, S.; Waltisperger, E.; Martínez-Lorenzana, G.; Condés-Lara, M.; Freund-Mercier, M. Sexually dimorphic expression of oxytocin binding sites in forebrain and spinal cord of the rat. Neuroscience 2005, 135, 147–154. [Google Scholar] [CrossRef]
- Sah, P.; Faber, E.S.L.; De Armentia, M.L.; Power, J. The Amygdaloid Complex: Anatomy and Physiology. Physiol. Rev. 2003, 83, 803–834. [Google Scholar] [CrossRef] [PubMed]
- Kondo, Y.; Sachs, B.; Sakuma, Y. Importance of the medial amygdala in rat penile erection evoked by remote stimuli from estrous females. Behav. Brain Res. 1997, 88, 153–160. [Google Scholar] [CrossRef]
- Dominguez, J.; Riolo, J.V.; Xu, Z.; Hull, E.M. Regulation by the Medial Amygdala of Copulation and Medial Preoptic Dopamine Release. J. Neurosci. 2001, 21, 349–355. [Google Scholar] [CrossRef]
- Ebner, K.; Bosch, O.; Krömer, S.A.; Singewald, N.; Neumann, I.D. Release of Oxytocin in the Rat Central Amygdala Modulates Stress-Coping Behavior and the Release of Excitatory Amino Acids. Neuropsychopharmacology 2004, 30, 223–230. [Google Scholar] [CrossRef]
- Huber, D.; Veinante, P.; Stoop, R. Vasopressin and Oxytocin Excite Distinct Neuronal Populations in the Central Amygdala. Science 2005, 308, 245–248. [Google Scholar] [CrossRef]
- Domes, G.; Heinrichs, M.; Gläscher, J.; Büchel, C.; Braus, D.F.; Herpertz, S.C. Oxytocin Attenuates Amygdala Responses to Emotional Faces Regardless of Valence. Biol. Psychiatry 2007, 62, 1187–1190. [Google Scholar] [CrossRef]
- Petrovic, P.; Kalisch, R.; Singer, T.; Dolan, R. Oxytocin Attenuates Affective Evaluations of Conditioned Faces and Amygdala Activity. J. Neurosci. 2008, 28, 6607–6615. [Google Scholar] [CrossRef]
- Lee, H.-J.; Pagani, J.; Young, W.S. Using transgenic mouse models to study oxytocin’s role in the facilitation of species propagation. Brain Res. 2010, 1364, 216–224. [Google Scholar] [CrossRef]
- Donaldson, Z.; Young, L.J. Oxytocin, Vasopressin, and the Neurogenetics of Sociality. Science 2008, 322, 900–904. [Google Scholar] [CrossRef]
- Preckel, K.; Kanske, P. Amygdala and oxytocin functioning as keys to understanding and treating autism: Commentary on an RDoC based approach. Neurosci. Biobehav. Rev. 2018, 94, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Abram, S.V.; De Coster, L.; Roach, B.J.; Mueller, A.B.; Van Erp, T.G.M.; Calhoun, V.D.; Preda, A.; Lim, K.; Turner, A.J.; Ford, J.M.; et al. Oxytocin Enhances an Amygdala Circuit Associated with Negative Symptoms in Schizophrenia: A Single-Dose, Placebo-Controlled, Crossover, Randomized Control Trial. Schizophr. Bull. 2019, 46, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Han, R.T.; Kim, Y.-B.; Park, E.-H.; Kim, J.Y.; Ryu, C.; Kim, H.Y.; Lee, J.; Pahk, K.; Shanyu, C.; Kim, H.; et al. Long-Term Isolation Elicits Depression and Anxiety-Related Behaviors by Reducing Oxytocin-Induced GABAergic Transmission in Central Amygdala. Front. Mol. Neurosci. 2018, 11, 246. [Google Scholar] [CrossRef]
- Roberto, M.; Kirson, D.; Khom, S. The Role of the Central Amygdala in Alcohol Dependence. Cold Spring Harb. Perspect. Med. 2021, 11, a039339. [Google Scholar] [CrossRef]
- Vinader-Caerols, C.; Collado, P.; Segovia, S.; Guillamon, A. Sex differences in the posteromedial cortical nucleus of the amygdala in the rat. Neuroreport 1998, 9, 2653–2656. [Google Scholar] [CrossRef] [PubMed]
- Vinader-Caerols, C.; Collado, P.; Segovia, S.; Guillamón, A. Estradiol masculinizes the posteromedial cortical nucleus of the amygdala in the rat. Brain Res. Bull. 2000, 53, 269–273. [Google Scholar] [CrossRef]
- Canteras, N.S.; Simerly, R.B.; Swanson, L.W. Organization of projections from the medial nucleus of the amygdala: A PHAL study in the rat. J. Comp. Neurol. 1995, 360, 213–245. [Google Scholar] [CrossRef]
- French, S.; Totterdell, S. Individual nucleus accumbens-projection neurons receive both basolateral amygdala and ventral subicular afferents in rats. Neuroscience 2003, 119, 19–31. [Google Scholar] [CrossRef]
- Kelley, A.; Domesick, V. The distribution of the projection from the hippocampal formation to the nucleus accumbens in the rat: An anterograde and retrograde-horseradish peroxidase study. Neuroscience 1982, 7, 2321–2335. [Google Scholar] [CrossRef]
- Witter, M.P. Connections of the subiculum of the rat: Topography in relation to columnar and laminar organization. Behav. Brain Res. 2006, 174, 251–264. [Google Scholar] [CrossRef]
- Giuliano, F.; Bernabé, J.; McKenna, K.; Longueville, F.; Rampin, O. Spinal proerectile effect of oxytocin in anesthetized rats. Am. J. Physiol. Integr. Comp. Physiol. 2001, 280, R1870–R1877. [Google Scholar] [CrossRef]
- Marson, L.; McKenna, K.E. CNS cell groups involved in the control of the ischiocavernosus and bulbospongiosus muscles: A transneuronal tracing study using pseudorabies virus. J. Comp. Neurol. 1996, 374, 161–179. [Google Scholar] [CrossRef]
- Honda, K.; Yanagimoto, M.; Negoro, H.; Narita, K.; Murata, T.; Higuchi, T. Excitation of oxytocin cells in the hypothalamic supraoptic nucleus by electrical stimulation of the dorsal penile nerve and tactile stimulation of the penis in the rat. Brain Res. Bull. 1999, 48, 309–313. [Google Scholar] [CrossRef]
- Yanagimoto, M.; Honda, K.; Goto, Y.; Negoro, H. Afferents originating from the dorsal penile nerve excite oxytocin cells in the hypothalamic paraventricular nucleus of the rat. Brain Res. 1996, 733, 292–296. [Google Scholar] [CrossRef]
- Rampin, O.; Gougis, S.; Giuliano, F.; Rousseau, J.P. Spinal Fos labeling and penile erection elicited by stimulation of dorsal nerve of the rat penis. Am. J. Physiol. Content 1997, 272, R1425–R1431. [Google Scholar] [CrossRef]
- Kita, I.; Yoshida, Y.; Nishino, S. An activation of parvocellular oxytocinergic neurons in the paraventricular nucleus in oxytocin-induced yawning and penile erection. Neurosci. Res. 2006, 54, 269–275. [Google Scholar] [CrossRef]
- Giuliano, F.; Rampin, O.; Brown, K.; Courtois, F.; Benoit, G.; Jardin, A. Stimulation of the medial preoptic area of the hypothalamus in the rat elicits increases in intracavernous pressure. Neurosci. Lett. 1996, 209, 1–4. [Google Scholar] [CrossRef]
- Giuliano, F.; Bernabé, J.; Brown, K.; Droupy, S.; Benoit, G.; Rampin, O. Erectile response to hypothalamic stimulation in rats: Role of peripheral nerves. Am. J. Physiol. Content 1997, 273, R1990–R1997. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.-K.; Chan, S.H.; Chang, L.S.; Chan, J.Y. Participation of Paraventricular Nucleus of Hypothalamus in Central Regulation of Penile Erection in the Rat. J. Urol. 1997, 158, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Marson, L.; McKenna, K.E. A role for 5-hydroxytryptamine in descending inhibition of spinal sexual reflexes. Exp. Brain Res. 1992, 88, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Marson, L.; List, M.S.; McKenna, K.E. Lesions of the nucleus paragigantocellularis alter ex copula penile reflexes. Brain Res. 1992, 592, 187–192. [Google Scholar] [CrossRef]
- Yells, D.P.; Hendricks, S.E.; Prendergast, M.A. Lesions of the nucleus paragigantocellularis: Effects on mating behavior in male rats. Brain Res. 1992, 596, 73–79. [Google Scholar] [CrossRef]
- Stancampiano, R.; Melis, M.R.; Argiolas, A. Penile erection and yawning induced by 5-HT1C receptor agonists in male rats: Relationship with dopaminergic and oxytocinergic transmission. Eur. J. Pharmacol. 1994, 261, 149–155. [Google Scholar] [CrossRef]
- Kimura, Y.; Naitou, Y.; Wanibuchi, F.; Yamaguchi, T. 5-HT2C receptor activation is a common mechanism on proerectile effects of apomorphine, oxytocin and melanotan-II in rats. Eur. J. Pharmacol. 2008, 589, 157–162. [Google Scholar] [CrossRef]
- Bancila, M.; Giuliano, F.; Rampin, O.; Mailly, P.; Brisorgueil, M.-J.; Calas, A.; Vergé, D. Evidence for a direct projection from the paraventricular nucleus of the hypothalamus to putative serotoninergic neurons of the nucleus paragigantocellularis involved in the control of erection in rats. Eur. J. Neurosci. 2002, 16, 1240–1248. [Google Scholar] [CrossRef]
- De Jong, T.R.; Neumann, I.D. Moderate Role of Oxytocin in the Pro-Ejaculatory Effect of the 5-HT1A Receptor Agonist 8-OH-DPAT. J. Sex. Med. 2015, 12, 17–28. [Google Scholar] [CrossRef]
- Veening, J.; de Jong, T.; Waldinger, M.; Korte, S.M.; Olivier, B. The role of oxytocin in male and female reproductive behavior. Eur. J. Pharmacol. 2015, 753, 209–228. [Google Scholar] [CrossRef]
- Saydoff, A.J.; Rittenhouse, A.P.; Van De Kar, L.D.; Brownfield, M.S. Enhanced serotonergic transmission stimulates oxytocin secretion in conscious male rats. J. Pharmacol. Exp. Ther. 1991, 257, 95–99. [Google Scholar]
- Bagdy, G.; Kalogeras, K.T. Stimulation of 5-HT1A and 5-HT2/5-HT1C receptors induce oxytocin release in the male rat. Brain Res. 1993, 611, 330–332. [Google Scholar] [CrossRef]
- Truitt, W.A. Identification of a Potential Ejaculation Generator in the Spinal Cord. Science 2002, 297, 1566–1569. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, H.; Matsuda, K.-I.; Zuloaga, D.G.; Hongu, H.; Wada, E.; Wada, K.; Jordan, C.L.; Breedlove, S.M.; Kawata, M. Sexually dimorphic gastrin releasing peptide system in the spinal cord controls male reproductive functions. Nat. Neurosci. 2008, 11, 634–636. [Google Scholar] [CrossRef] [PubMed]
- Kozyrev, N.; Coolen, L.M. Activation of galanin and cholecystokinin receptors in the lumbosacral spinal cord is required for ejaculation in male rats. Eur. J. Neurosci. 2017, 45, 846–858. [Google Scholar] [CrossRef] [PubMed]
- Uta, D.; Oti, T.; Sakamoto, T.; Sakamoto, H. In Vivo Electrophysiology of Peptidergic Neurons in Deep Layers of the Lumbar Spinal Cord after Optogenetic Stimulation of Hypothalamic Paraventricular Oxytocin Neurons in Rats. Int. J. Mol. Sci. 2021, 22, 3400. [Google Scholar] [CrossRef] [PubMed]
- Kozyrev, N.; Lehman, M.N.; Coolen, L.M. Activation of Gastrin-releasing Peptide Receptors in the Lumbosacral Spinal Cord is Required for Ejaculation in Male Rats. J. Sex. Med. 2012, 9, 1303–1318. [Google Scholar] [CrossRef]
- Esakamoto, H. Sexually dimorphic nuclei in the spinal cord control male sexual functions. Front. Neurosci. 2014, 8, 184. [Google Scholar] [CrossRef]
- Oti, T.; Satoh, K.; Uta, D.; Nagafuchi, J.; Tateishi, S.; Ueda, R.; Takanami, K.; Young, L.J.; Galione, A.; Morris, J.F.; et al. Oxytocin Influences Male Sexual Activity via Non-synaptic Axonal Release in the Spinal Cord. Curr. Biol. 2021, 31, 103–114. [Google Scholar] [CrossRef]
- Shinghal, R.; Barnes, A.; Mahar, K.M.; Stier, B.; Giancaterino, L.; Condreay, L.D.; Black, L.; McCallum, S.W. Safety and Efficacy of Epelsiban in the Treatment of Men with Premature Ejaculation: A Randomized, Double-Blind, Placebo-Controlled, Fixed-Dose Study. J. Sex. Med. 2013, 10, 2506–2517. [Google Scholar] [CrossRef]
- Clement, P.; Bernabé, J.; Compagnie, S.; Alexandre, L.; McCallum, S.; Giuliano, F. Inhibition of ejaculation by the non-peptide oxytocin receptor antagonist GSK557296: A multi-level site of action. Br. J. Pharmacol. 2013, 169, 1477–1485. [Google Scholar] [CrossRef]
- Althof, S.; Osterloh, I.H.; Muirhead, G.J.; George, K.; Girard, N.; Jennings, W.; Adams, M.; Shockey, G.; Kaminetsky, J.; Miner, M.; et al. The Oxytocin Antagonist Cligosiban Fails to Prolong Intravaginal Ejaculatory Latency in Men with Lifelong Premature Ejaculation: Results of a Randomized, Double-Blind, Placebo-Controlled Phase IIb trial (PEDRIX). J. Sex. Med. 2019, 16, 1188–1198. [Google Scholar] [CrossRef]
- McMahon, C.; Althof, S.; Rosen, R.; Giuliano, F.; Miner, M.; Osterloh, I.H.; Muirhead, G.J.; Harty, B. The Oxytocin Antagonist Cligosiban Prolongs Intravaginal Ejaculatory Latency and Improves Patient-Reported Outcomes in Men with Lifelong Premature Ejaculation: Results of a Randomized, Double-Blind, Placebo-Controlled Proof-of-Concept Trial (PEPIX). J. Sex. Med. 2019, 16, 1178–1187. [Google Scholar] [CrossRef]
- Bitran, D.; Hull, E. Pharmacological analysis of male rat sexual behavior. Neurosci. Biobehav. Rev. 1987, 11, 365–389. [Google Scholar] [CrossRef]
- Sachs, B.D. Conceptual and neural mechanisms of masculine copulatory behavior. In Sex and Behavior: Status and Prospectus; McGill, T.E., Dewsbury, D.A., Sachs, B.D., Eds.; Plenum: New York, NY, USA, 1978; pp. 267–295. [Google Scholar]
- Ågmo, A. Male rat sexual behavior. Brain Res. Protoc. 1997, 1, 203–209. [Google Scholar] [CrossRef]
- Caggiula, A.R.; Herndon, J.G.; Scanlon, R.; Greenstone, D.; Bradshaw, W.; Sharp, D. Dissociation of active from immobility components of sexual behavior in female rats by central 6-hydroxydopamine: Implications for CA involvement in sexual behavior and sensorimotor responsiveness. Brain Res. 1979, 172, 505–520. [Google Scholar] [CrossRef]
- Caggiula, A.R.; Shaw, D.H.; Antelman, S.M.; Edwards, D.J. Interactive effects of brain catecholamines and variations in sexual and non-sexual arousal on copulatory behavior of male rats. Brain Res. 1976, 111, 321–336. [Google Scholar] [CrossRef]
- Burns-Cusato, M.; Scordalakes, E.M.; Rissman, E.F. Of mice and missing data: What we know (and need to learn) about male sexual behavior. Physiol. Behav. 2004, 83, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Okabe, S.; Kitano, K.; Nagasawa, M.; Mogi, K.; Kikusui, T. Testosterone inhibits facilitating effects of parenting experience on parental behavior and the oxytocin neural system in mice. Physiol. Behav. 2013, 118, 159–164. [Google Scholar] [CrossRef]
- Floody, O.R. Processes regulating the initiation and postejaculatory resumption of copulatory behavior in male hamsters. Behav. Process. 2014, 105, 79–84. [Google Scholar] [CrossRef]
- Gray, G.D.; Dewsbury, A.D. A quantitative description of copulatory behavior in prairie voles (Microtus ochrogaster). Brain Behav. Evol. 1973, 8, 426–452. [Google Scholar] [CrossRef] [PubMed]
- Nitsch, F.; Stueckle, S.; Stahl, D.; Zinner, D. Copulation patterns in captive hamadryas baboons: A quantitative analysis. Primates 2011, 52, 373–383. [Google Scholar] [CrossRef]
- Absil, P.; Das, S.; Balthazart, J. Effects of apomorphine on sexual behavior in male quail. Pharmacol. Biochem. Behav. 1994, 47, 77–88. [Google Scholar] [CrossRef]
- Balthazart, J.; Ball, G. Topography in the preoptic region: Differential regulation of appetitive and consummatory male sexual behaviors. Front. Neuroendocr. 2007, 28, 161–178. [Google Scholar] [CrossRef]
- Rauceo, S.; Harding, C.F.; Maldonado, A.; Gaysinkaya, L.; Tulloch, I.; Rodriguez, E. Dopaminergic modulation of reproductive behavior and activity in male zebra finches. Behav. Brain Res. 2008, 187, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Lue, T.F.; Tanagho, E.A. Physiology of Erection and Pharmacological Management of Impotence. J. Urol. 1987, 137, 829–836. [Google Scholar] [CrossRef]
- Burnett, A.; Lowenstein, C.; Bredt, D.; Chang, T.; Snyder, S. Nitric oxide: A physiologic mediator of penile erection. Science 1992, 257, 401–403. [Google Scholar] [CrossRef] [PubMed]
- Rajfer, J.; Aronson, W.J.; Bush, P.A.; Dorey, F.J.; Ignarro, L.J. Nitric Oxide as a Mediator of Relaxation of the Corpus Cavernosum in Response to Nonadrenergic, Noncholinergic Neurotransmission. New Engl. J. Med. 1992, 326, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Lenz, K.M.; McCarthy, M.M. Organized for sex–steroid hormones and the developing hypothalamus. Eur. J. Neurosci. 2010, 32, 2096–2104. [Google Scholar] [CrossRef]
- Snoeren, E.M.S. Female Reproductive Behavior. Curr. Top. Behav. Neurosci. 2018, 43, 1–44. [Google Scholar]
- Jennings, K.J.; De Lecea, L. Neural and Hormonal Control of Sexual Behavior. Endocrinology 2020, 161, 150. [Google Scholar] [CrossRef]
- Bertolini, A.; Gessa, G.L. Behavioral effects of ACTH and MSH peptides. J. Endocrinol. Investig. 1981, 4, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Dornan, W.A.; Malsbury, C.W. Neuropeptides and male sexual behavior. Neurosci. Biobehav. Rev. 1989, 13, 1–15. [Google Scholar] [CrossRef]
- Argiolas, A.; Melis, M.R.; Murgia, S.; Schiöth, H.B. ACTH- and α-MSH-induced grooming, stretching, yawning and penile erection in male rats: Site of action in the brain and role of melanocortin receptors. Brain Res. Bull. 2000, 51, 425–431. [Google Scholar] [CrossRef]
- Paredes, R. Opioids and sexual reward. Pharmacol. Biochem. Behav. 2014, 121, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Stoneham, M.D.; Everitt, B.; Hansen, S.; Lightman, S.L.; Todd, K. Oxytocin and sexual behaviour in the male rat and rabbit. J. Endocrinol. 1985, 107, 97–106. [Google Scholar] [CrossRef]
- Hughes, A.M.; Everitt, B.J.; Lightman, S.L.; Todd, K. Oxytocin in the central nervous system and sexual behaviour in male rats. Brain Res. 1987, 414, 133–137. [Google Scholar] [CrossRef]
- Arletti, R.; Benelli, A.; Bertolini, A. Oxytocin Involvement in Male and Female Sexual Behavior. Ann. N. Y. Acad. Sci. 1992, 652, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Arletti, R.; Benelli, A.; Bertolini, A. Sexual behavior of aging male rats is stimulated by oxytocin. Eur. J. Pharmacol. 1990, 179, 377–381. [Google Scholar] [CrossRef]
- Winslow, J.; Insel, T. Social status in pairs of male squirrel monkeys determines the behavioral response to central oxytocin administration. J. Neurosci. 1991, 11, 2032–2038. [Google Scholar] [CrossRef] [PubMed]
- Argiolas, A.; Collu, M.; Gessa, G.L.; Melis, M.R.; Serra, G. The oxytocin antagonist d(CH2)5Tyr(Me)-Orn8-vasotocin inhibits male copulatory behaviour in rats. Eur. J. Pharmacol. 1988, 149, 389–392. [Google Scholar] [CrossRef]
- Argiolas, A.; Collu, M.; D’Aquila, P.; Gessa, G.; Melis, M.; Serra, G. Apomorphine stimulation of male copulatory behavior is prevented by the oxytocin antagonist d(CH2)5Tyr(Me)-Orn8-vasotocin in rats. Pharmacol. Biochem. Behav. 1989, 33, 81–83. [Google Scholar] [CrossRef]
- Witt, D.M.; Insel, T.R. Increased Fos Expression in Oxytocin Neurons Following Masculine Sexual Behavior. J. Neuroendocr. 1994, 6, 13–18. [Google Scholar] [CrossRef]
- Angioni, L.; Cocco, C.; Ferri, G.-L.; Argiolas, A.; Melis, M.R.; Sanna, F. Involvement of nigral oxytocin in locomotor activity: A behavioral, immunohistochemical and lesion study in male rats. Horm. Behav. 2016, 83, 23–38. [Google Scholar] [CrossRef]
- Nishimori, K.; Young, L.J.; Guo, Q.; Wang, Z.; Insel, T.R.; Matzuk, M.M. Oxytocin is required for nursing but is not essential for parturition or reproductive behavior. Proc. Natl. Acad. Sci. USA 1996, 93, 11699–11704. [Google Scholar] [CrossRef]
- Iii, W.S.Y.; Shepard, E.; Amico, J.; Hennighausen, L.; Wagner, K.-U.; Lamarca, M.E.; McKinney, C.; Ginns, E.I. Deficiency in Mouse Oxytocin Prevents Milk Ejection, but not Fertility or Parturition. J. Neuroendocr. 1996, 8, 847–853. [Google Scholar] [CrossRef]
- Huang, P.L.; Dawson, T.M.; Bredt, D.S.; Snyder, S.H.; Fishman, M.C. Targeted disruption of the neuronal nitric oxide synthase gene. Cell 1993, 75, 1273–1286. [Google Scholar] [CrossRef]
- Gibson, M.J.; Wu, T.; Miller, G.M.; Silverman, A.-J. What Nature’s Knockout Teaches Us about GnRH Activity: Hypogonadal Mice and Neuronal Grafts. Horm. Behav. 1997, 31, 212–220. [Google Scholar] [CrossRef]
- Nishimori, K.; Takayanagi, Y.; Yoshida, M.; Kasahara, Y.; Young, L.J.; Kawamata, M. New aspects of oxytocin receptor function revealed by knockout mice: Sociosexual behaviour and control of energy balance. Prog. Brain Res. 2008, 170, 79–90. [Google Scholar] [CrossRef]
- Lazzari, V.M.; Becker, R.O.; De Azevedo, M.S.; Morris, M.; Rigatto, K.; Almeida, S.; Lucion, A.B.; Giovenardi, M. Oxytocin modulates social interaction but is not essential for sexual behavior in male mice. Behav. Brain Res. 2013, 244, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Benelli, A.; Poggioli, R.; Luppi, P.; Ruini, L.; Bertolini, A.; Arletti, R. Oxytocin enhances, and oxytocin antagonism decreases, sexual receptivity in intact female rats. Neuropeptides 1994, 27, 245–250. [Google Scholar] [CrossRef]
- Pedersen, C.A.; Boccia, M. Oxytocin Maintains as Well as Initiates Female Sexual Behavior: Effects of a Highly Selective Oxytocin Antagonist. Horm. Behav. 2002, 41, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, C.; Boccia, M. Vasopressin interactions with oxytocin in the control of female sexual behavior. Neuroscience 2006, 139, 843–851. [Google Scholar] [CrossRef]
- Gorzalka, B.B.; Lester, G.L. Oxytocin-induced facilitation of lordosis behaviour in rats is progesterone-dependent. Neuropeptides 1987, 10, 55–65. [Google Scholar] [CrossRef]
- Caldwell, J.D.; Jirikowski, G.F.; Greer, E.R.; Stumpf, W.E.; Pedersen, C.A. Ovarian steroids and sexual interaction alter oxytocinergic content and distribution in the basal forebrain. Brain Res. 1988, 446, 236–244. [Google Scholar] [CrossRef]
- Coirini, H.; Schumacher, M.; Flanagan, L.; McEwen, B. Transport of estrogen-induced oxytocin receptors in the ventromedial hypothalamus. J. Neurosci. 1991, 11, 3317–3324. [Google Scholar] [CrossRef]
- Coirini, H.; Johnson, A.E.; Schumacher, M.; McEwen, B.S. Sex Differences in the Regulation of Oxytocin Receptors by Ovarian Steroids in the Ventromedial Hypothalamus of the Rat. Neuroendocrinology 1992, 55, 269–275. [Google Scholar] [CrossRef]
- Johnson, A.E.; Coirini, H.; Insel, T.R.; McEwen, B.S. The Regulation of Oxytocin Receptor Binding in the Ventromedial Hypothalaimic Nucleus by Testosterone and Its Metabolites. Endocrinology 1991, 128, 891–896. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, M.; Coirini, H.; Pfaff, D.; McEwen, B. Behavioral effects of progesterone associated with rapid modulation of oxytocin receptors. Science 1990, 250, 691–694. [Google Scholar] [CrossRef]
- Caldwell, J.; Barakat, A.; Smith, D.; Hruby, V.; Pedersen, C. A uterotonic antagonist blocks the oxytocin-induced facilitation of female sexual receptivity. Brain Res. 1990, 512, 291–296. [Google Scholar] [CrossRef]
- Caldwell, J.D.; Jirikowski, G.F.; Greer, E.R.; Pedersen, C.A. Medial preoptic area oxytocin and female sexual receptivity. Behav. Neurosci. 1989, 103, 655–662. [Google Scholar] [CrossRef]
- McCarthy, M.M.; Kleopoulos, S.P.; Mobbs, C.V.; Pfaff, D.W. Infusion of Antisense Oligo-deoxynucleotides to the Oxytocin Receptor in the Ventromedial Hypothalamus Reduces Estrogen-Induced Sexual Receptivity and Oxytocin Receptor Binding in the Female Rat. Neuroendocrinology 1994, 59, 432–440. [Google Scholar] [CrossRef]
- Bale, T.L.; Davis, A.M.; Auger, A.P.; Dorsa, D.M.; McCarthy, M.M. CNS Region-Specific Oxytocin Receptor Expression: Importance in Regulation of Anxiety and Sex Behavior. J. Neurosci. 2001, 21, 2546–2552. [Google Scholar] [CrossRef] [PubMed]
- Ferri, S.L.; Rohrbach, C.J.; Way, S.E.; Curtis, K.S.; Curtis, J.T.; Flanagan-Cato, L.M. Dendritic arbor of neurons in the hypothalamic ventromedial nucleus in female prairie voles (Microtus ochrogaster). Horm. Behav. 2013, 63, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Griffin, G.D.; Ferri-Kolwicz, S.L.; Reyes, B.A.; Van Bockstaele, E.J.; Flanagan-Cato, L.M. Ovarian hormone-induced reorganization of oxytocin-labeled dendrites and synapses lateral to the hypothalamic ventromedial nucleus in female rats. J. Comp. Neurol. 2010, 518, 4531–4545. [Google Scholar] [CrossRef] [PubMed]
- Frye, C.A.; Walf, A.A. Oxytocin and/or steroid hormone binding globulin infused into the ventral tegmental area modulates progestogen-mediated lordosis. Neuropharmacology 2010, 58, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, L.M.; Pfaus, J.; Pfaff, D.W.; McEwen, B.S. Induction of FOS Immunoreactivity in Oxytocin Neurons after Sexual Activity in Female Rats. Neuroendocrinology 1993, 58, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Mac Cionnaith, C.E.; Lemay, A.; Gomez-Perales, E.L.; Robert, G.; Cernik, R.; Brake, W.G.; Pfaus, J. Fos expression is increased in oxytocin neurons of female rats with a sexually conditioned mate preference for an individual male rat. Horm. Behav. 2019, 117, 104612. [Google Scholar] [CrossRef] [PubMed]
- Nyuyki, K.D.; Waldherr, M.; Baeuml, S.; Neumann, I.D. Yes, I Am Ready Now: Differential Effects of Paced versus Unpaced Mating on Anxiety and Central Oxytocin Release in Female Rats. PLoS ONE 2011, 6, e23599. [Google Scholar] [CrossRef]
- Waldherr, M.; Neumann, I.D. Centrally released oxytocin mediates mating-induced anxiolysis in male rats. Proc. Natl. Acad. Sci. USA 2007, 104, 16681–16684. [Google Scholar] [CrossRef] [PubMed]
- Whitman, D.C.; Albers, H.E. Role of oxytocin in the hypothalamic regulation of sexual receptivity in hamsters. Brain Res. 1995, 680, 73–79. [Google Scholar] [CrossRef]
- Whitman, D.C.; Albers, H.E. Oxytocin immunoreactivity in the hypothalamus of female hamsters. Cell Tissue Res. 1998, 291, 231–237. [Google Scholar] [CrossRef]
- Cushing, B.S.; Carter, C.S. Prior Exposure to Oxytocin Mimics the Effects of Social Contact and Facilitates Sexual Behaviour In Females. J. Neuroendocr. 2001, 11, 765–769. [Google Scholar] [CrossRef] [PubMed]
- Dluzen, D.; Ramírez, V.; Carter, C.; Getz, L. Male vole urine changes luteinizing hormone-releasing hormone and norepinephrine in female olfactory bulb. Science 1981, 212, 573–575. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.; Witt, D.M.; Auksi, T.; Casten, L. Estrogen and the induction of lordosis in female and male prairie voles (Microtus ochrogaster). Horm. Behav. 1987, 21, 65–73. [Google Scholar] [CrossRef]
- Carter, C.; Witt, D.; Manock, S.; Adams, K.; Bahr, J.; Carlstead, K. Hormonal correlates of sexual behavior and ovulation in male-induced and postpartum estrus in female prairie voles. Physiol. Behav. 1989, 46, 941–948. [Google Scholar] [CrossRef]
- Becker, R.O.; Lazzari, V.M.; Menezes, I.C.; Morris, M.; Rigatto, K.; Lucion, A.B.; Rasia-Filho, A.A.; Giovenardi, M. Sexual behavior and dendritic spine density of posterodorsal medial amygdala neurons in oxytocin knockout female mice. Behav. Brain Res. 2013, 256, 95–100. [Google Scholar] [CrossRef]
- Zimmermann-Peruzatto, J.M.; Lazzari, V.M.; Agnes, G.; Becker, R.O.; De Moura, A.C.; Guedes, R.P.; Lucion, A.B.; Almeida, S.; Giovenardi, M. The Impact of Oxytocin Gene Knockout on Sexual Behavior and Gene Expression Related to Neuroendocrine Systems in the Brain of Female Mice. Cell. Mol. Neurobiol. 2017, 37, 803–815. [Google Scholar] [CrossRef] [PubMed]
- Dhungel, S.; Rai, D.; Terada, M.; Orikasa, C.; Nishimori, K.; Sakuma, Y.; Kondo, Y. Oxytocin is indispensable for conspecific-odor preference and controls the initiation of female, but not male, sexual behavior in mice. Neurosci. Res. 2019, 148, 34–41. [Google Scholar] [CrossRef]
- Blitzer, D.; Wells, T.; Hawley, W. Administration of an oxytocin receptor antagonist attenuates sexual motivation in male rats. Horm. Behav. 2017, 94, 33–39. [Google Scholar] [CrossRef]
- Sanna, F.; Argiolas, A.; Melis, M.R. Oxytocin-induced yawning: Sites of action in the brain and interaction with mesolimbic/mesocortical and incertohypothalamic dopaminergic neurons in male rats. Horm. Behav. 2012, 62, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Everitt, B. Sexual motivation: A neural and behavioural analysis of the mechanisms underlying appetitive and copulatory responses of male rats. Neurosci. Biobehav. Rev. 1990, 14, 217–232. [Google Scholar] [CrossRef]
- Everitt, B.J.; Robbins, T. Neural systems of reinforcement for drug addiction: From actions to habits to compulsion. Nat. Neurosci. 2005, 8, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Fibiger, H.C.; Phillips, A.G. Mesocorticolimbic Dopamine Systems and Reward. Ann. N. Y. Acad. Sci. 1988, 537, 206–215. [Google Scholar] [CrossRef]
- Wise, A.R.; Rompre, P.P. Brain Dopamine and Reward. Annu. Rev. Psychol. 1989, 40, 191–225. [Google Scholar] [CrossRef]
- Goto, Y.; Grace, A. Dopaminergic modulation of limbic and cortical drive of nucleus accumbens in goal-directed behavior. Nat. Neurosci. 2005, 8, 805–812. [Google Scholar] [CrossRef]
- Carter, C.S.; Devries, A.C.; Taymans, S.E.; Roberts, R.L.; Williams, J.R.; Getz, L.L. Peptides, Steroids, and Pair Bonding. Ann. N. Y. Acad. Sci. 1997, 807, 260–272. [Google Scholar] [CrossRef]
- Ferguson, J.N.; Young, L.J.; Hearn, E.F.; Matzuk, M.M.; Insel, T.R.; Winslow, J.T. Social amnesia in mice lacking the oxytocin gene. Nat. Genet. 2000, 25, 284–288. [Google Scholar] [CrossRef]
- Ferguson, J.N.; Aldag, J.M.; Insel, T.R.; Young, L.J. Oxytocin in the Medial Amygdala is Essential for Social Recognition in the Mouse. J. Neurosci. 2001, 21, 8278–8285. [Google Scholar] [CrossRef]
- Takayanagi, Y.; Yoshida, M.; Bielsky, I.F.; Ross, H.E.; Kawamata, M.; Onaka, T.; Yanagisawa, T.; Kimura, T.; Matzuk, M.M.; Young, L.J.; et al. Pervasive social deficits, but normal parturition, in oxytocin receptor-deficient mice. Proc. Natl. Acad. Sci. USA 2005, 102, 16096–16101. [Google Scholar] [CrossRef]
- Walch, K.; Eder, R.; Schindler, A.; Feichtinger, W. The Effect of Single-Dose Oxytocin Application on Time to Ejaculation and Seminal Parameters in Men. J. Assist. Reprod. Genet. 2001, 18, 655–659. [Google Scholar] [CrossRef]
- Burri, A.; Heinrichs, M.; Schedlowski, M.; Kruger, T.H. The acute effects of intranasal oxytocin administration on endocrine and sexual function in males. Psychoneuroendocrinology 2008, 33, 591–600. [Google Scholar] [CrossRef]
- Behnia, B.; Heinrichs, M.; Bergmann, W.; Jung, S.; Germann, J.; Schedlowski, M.; Hartmann, U.; Kruger, T.H. Differential effects of intranasal oxytocin on sexual experiences and partner interactions in couples. Horm. Behav. 2014, 65, 308–318. [Google Scholar] [CrossRef]
- MacDonald, K.; Feifel, D. Dramatic Improvement in Sexual Function Induced by Intranasal Oxytocin. J. Sex. Med. 2012, 9, 1407–1410. [Google Scholar] [CrossRef]
- Kruger, T.H.; Deiter, F.; Zhang, Y.; Jung, S.; Schippert, C.; Kahl, K.G.; Heinrichs, M.; Schedlowski, M.; Hartmann, U. Effects of Intranasal Oxytocin Administration on Sexual Functions in Healthy Women. J. Clin. Psychopharmacol. 2018, 38, 239–242. [Google Scholar] [CrossRef]
- Muin, D.A.; Wolzt, M.; Marculescu, R.; Rezaei, S.S.; Salama, M.; Fuchs, C.; Luger, A.; Bragagna, E.; Litschauer, B.; Bayerle-Eder, M. Effect of long-term intranasal oxytocin on sexual dysfunction in premenopausal and postmenopausal women: A randomized trial. Fertil. Steril. 2015, 104, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Snyder, S. Nitric oxide: First in a new class of neurotransmitters. Science 1992, 257, 494–496. [Google Scholar] [CrossRef]
- Alonso, J.R.; Blanco, E.; Sánchez, F.; Arévalo, R.; Aijón, J.; Vazquez, R. Coexistence of NADPH-diaphorase with vasopressin and oxytocin in the hypothalamic magnocellular neurosecretory nuclei of the rat. Cell Tissue Res. 1994, 276, 31–34. [Google Scholar] [CrossRef]
- Melis, M.R.; Argiolas, A. Role op central nitric oxide in the control of penile erection and yawning. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 1997, 21, 899–922. [Google Scholar] [CrossRef]
- Alquicer, G.; Silva-Gómez, A.B.; Peralta, F.; Flores, G. Neonatal ventral hippocampus lesion alters the dopamine content in the limbic regions in postpubertal rats. Int. J. Dev. Neurosci. 2004, 22, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Balfour, M.; Brown, J.; Yu, L.; Coolen, L. Potential contributions of efferents from medial prefrontal cortex to neural activation following sexual behavior in the male rat. Neuroscience 2006, 137, 1259–1276. [Google Scholar] [CrossRef] [PubMed]
- Karreman, M.; Westerink, B.H.C.; Moghaddam, B. Excitatory Amino Acid Receptors in the Ventral Tegmental Area Regulate Dopamine Release in the Ventral Striatum. J. Neurochem. 2002, 67, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Rossetti, Z.L.; Marcangione, C.; Wise, R.A. Increase of extracellular glutamate and expression of Fos-like immunoreactivity in the ventral tegmental area in response to electrical stimulation of the prefrontal cortex. J. Neurochem. 2002, 70, 1503–1512. [Google Scholar] [CrossRef] [PubMed]
- Legault, M.; Wise, R.A. Injections of N-methyl-D-aspartate into the ventral hippocampus increase extracellular dopamine in the ventral tegmental area and nucleus accumbens. Synapse 1999, 31, 241–249. [Google Scholar] [CrossRef]
- Legault, M.; Rompré, P.-P.; Wise, R.A. Chemical Stimulation of the Ventral Hippocampus Elevates Nucleus Accumbens Dopamine by Activating Dopaminergic Neurons of the Ventral Tegmental Area. J. Neurosci. 2000, 20, 1635–1642. [Google Scholar] [CrossRef] [PubMed]
- Floresco, S.B.; Todd, C.L.; Grace, A.A. Glutamatergic Afferents from the Hippocampus to the Nucleus Accumbens Regulate Activity of Ventral Tegmental Area Dopamine Neurons. J. Neurosci. 2001, 21, 4915–4922. [Google Scholar] [CrossRef]
- Daniels, D.; Miselis, R.R.; Flanagan-Cato, L.M. Central Neuronal Circuit Innervating the Lordosis-Producing Muscles Defined by Transneuronal Transport of Pseudorabies Virus. J. Neurosci. 1999, 19, 2823–2833. [Google Scholar] [CrossRef] [PubMed]
- Muin, D.A.; Rezaei, S.S.; Tremmel-Scheinost, M.; Salama, M.; Luger, A.; Wolzt, M.; Husslein, P.W.; Bayerle-Eder, M. Men’s sexual response to female partner’s intranasal oxytocin administration for hypoactive sexual desire disorder: An open prospective cohort study. Fertil. Steril. 2017, 107, 781–787.e3. [Google Scholar] [CrossRef]
- Lalu, K.; Lampelo, S.; Vanha-Perttula, T. Characterization of three aminopeptidases purified from maternal serum. Biochim. Biophys. Acta Protein Struct. Mol. Enzym. 1986, 873, 190–197. [Google Scholar] [CrossRef]
- Burbach, J.P.; Lebouille, J.L. Proteolytic conversion of arginine-vasopressin and oxytocin by brain synaptic membranes. Characterization of formed peptides and mechanisms of proteolysis. J. Biol. Chem. 1983, 258, 1487–1494. [Google Scholar] [CrossRef]
- Stancampiano, R.; Melis, M.; Argiolas, A. Proteolytic conversion of oxytocin by brain synaptic membranes: Role of aminopeptidases and endopeptidases. Peptides 1991, 12, 1119–1125. [Google Scholar] [CrossRef]
- Stancampiano, R.; Argiolas, A. Proteolytic conversion of oxytocin in vivo after microinjection in the rat hippocampus. Peptides 1993, 14, 465–469. [Google Scholar] [CrossRef]
- Manning, M.; Stoev, S.; Chini, B.; Durroux, T.; Mouillac, B.; Guillon, G. Peptide and non-peptide agonists and antagonists for the vasopressin and oxytocin V1a, V1b, V2 and OT receptors: Research tools and potential therapeutic agents. Prog. Brain Res. 2008, 170, 473–512. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, M.; Wiśniewska, H.; Tariga, H.; Ibanez, G.; Collins, J.C.; Wisniewski, K.; Qi, S.; Srinivasan, K.; Hargrove, D.; Lindstrom, B.F. Selective and non-selective OT receptor agonists induce different locomotor behaviors in male rats via central OT receptors and peripheral V1a receptors. Neuropeptides 2018, 70, 64–75. [Google Scholar] [CrossRef]
- Tunstall, B.J.; Kirson, D.; Zallar, L.J.; McConnell, S.A.; Vendruscolo, J.C.M.; Ho, C.P.; Oleata, C.S.; Khom, S.; Manning, M.; Lee, M.R.; et al. Oxytocin blocks enhanced motivation for alcohol in alcohol dependence and blocks alcohol effects on GABAergic transmission in the central amygdala. PLoS Biol. 2019, 17, e2006421. [Google Scholar] [CrossRef] [PubMed]
- Hilfiger, L.; Zhao, Q.; Kerspern, D.; Inquimbert, P.; Andry, V.; Goumon, Y.; Darbon, P.; Hibert, M.; Charlet, A. A Nonpeptide Oxytocin Receptor Agonist for a Durable Relief of Inflammatory Pain. Sci. Rep. 2020, 10, 3017. [Google Scholar] [CrossRef]
- Patel, M.M.; Patel, B.M. Crossing the Blood–Brain Barrier: Recent Advances in Drug Delivery to the Brain. CNS Drugs 2017, 31, 109–133. [Google Scholar] [CrossRef]
- Oppong-Damoah, A.; Zaman, R.U.; D’Souza, M.J.; Murnane, K.S. Nanoparticle encapsulation increases the brain penetrance and duration of action of intranasal oxytocin. Horm. Behav. 2019, 108, 20–29. [Google Scholar] [CrossRef]
- Carter, C.S. Neuroendocrine perspectives on social attachment and love. Psychoneuroendocrinology 1998, 23, 779–818. [Google Scholar] [CrossRef]
- Carter, C.S. The Role of Oxytocin and Vasopressin in Attachment. Psychodyn. Psychiatry 2017, 45, 499–517. [Google Scholar] [CrossRef]
- Veenema, A.; Neumann, I. Central vasopressin and oxytocin release: Regulation of complex social behaviours. Prog. Brain Res. 2008, 170, 261–276. [Google Scholar] [CrossRef] [PubMed]








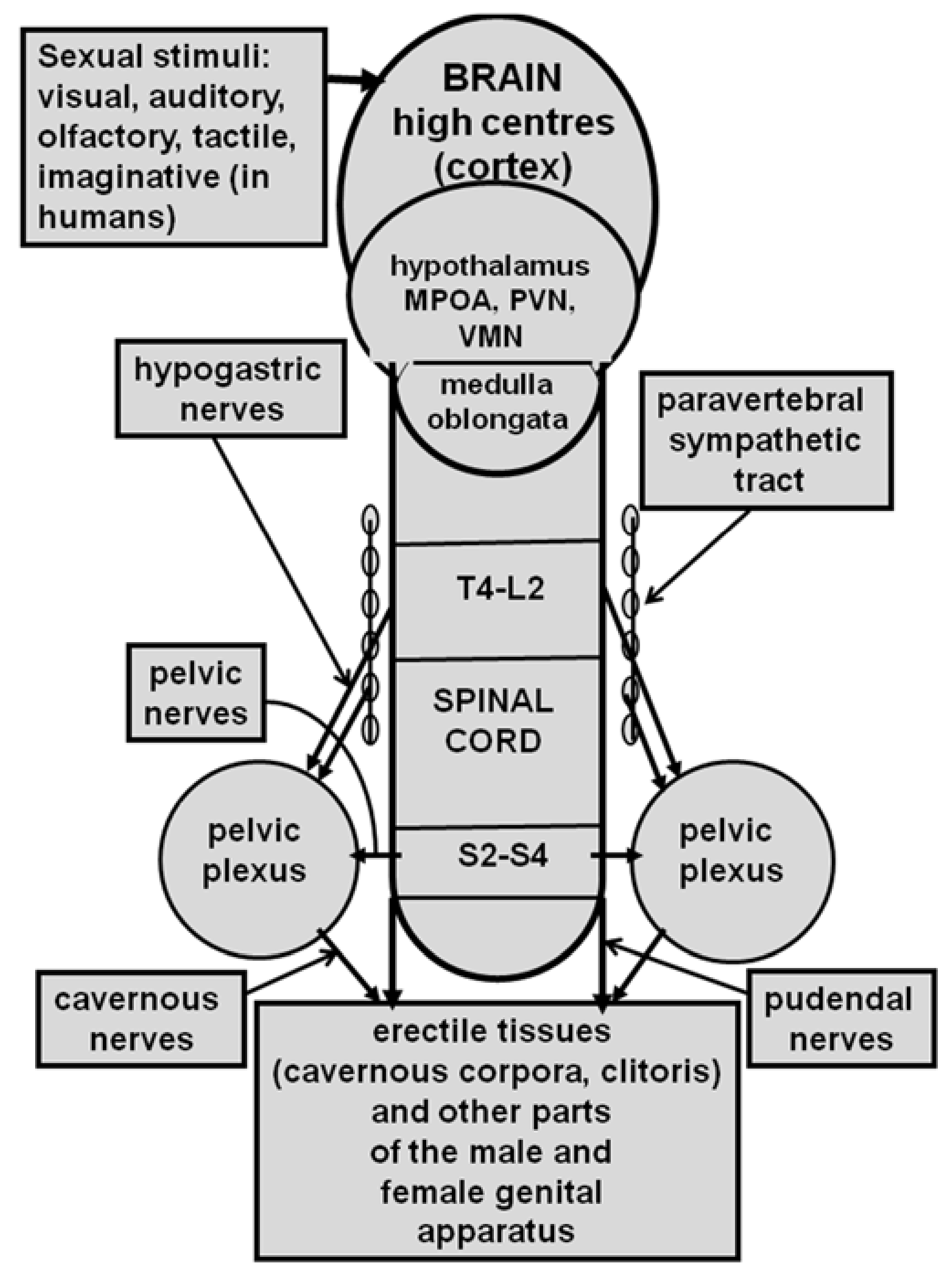{kind=link}
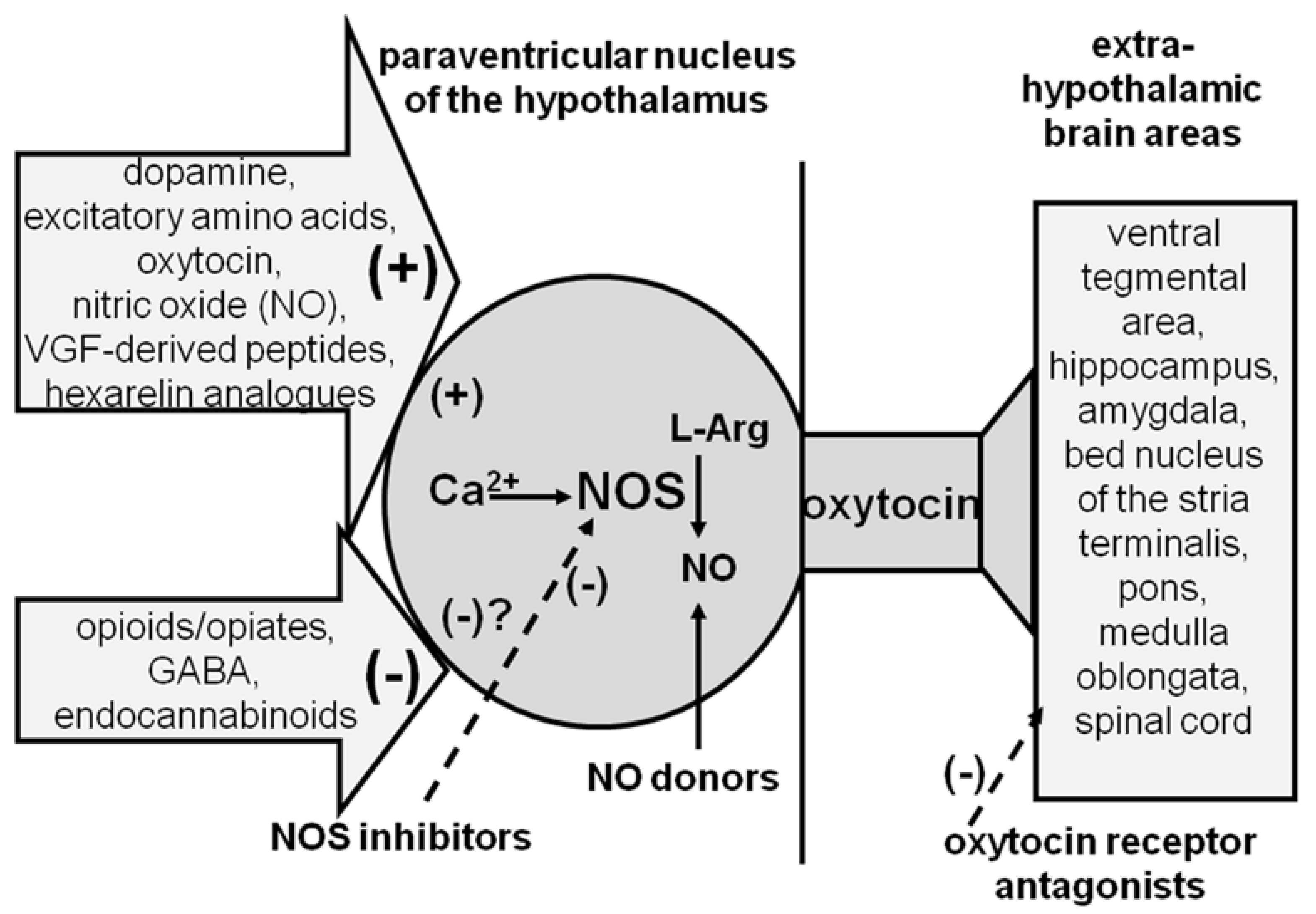{kind=link}
{kind=link}
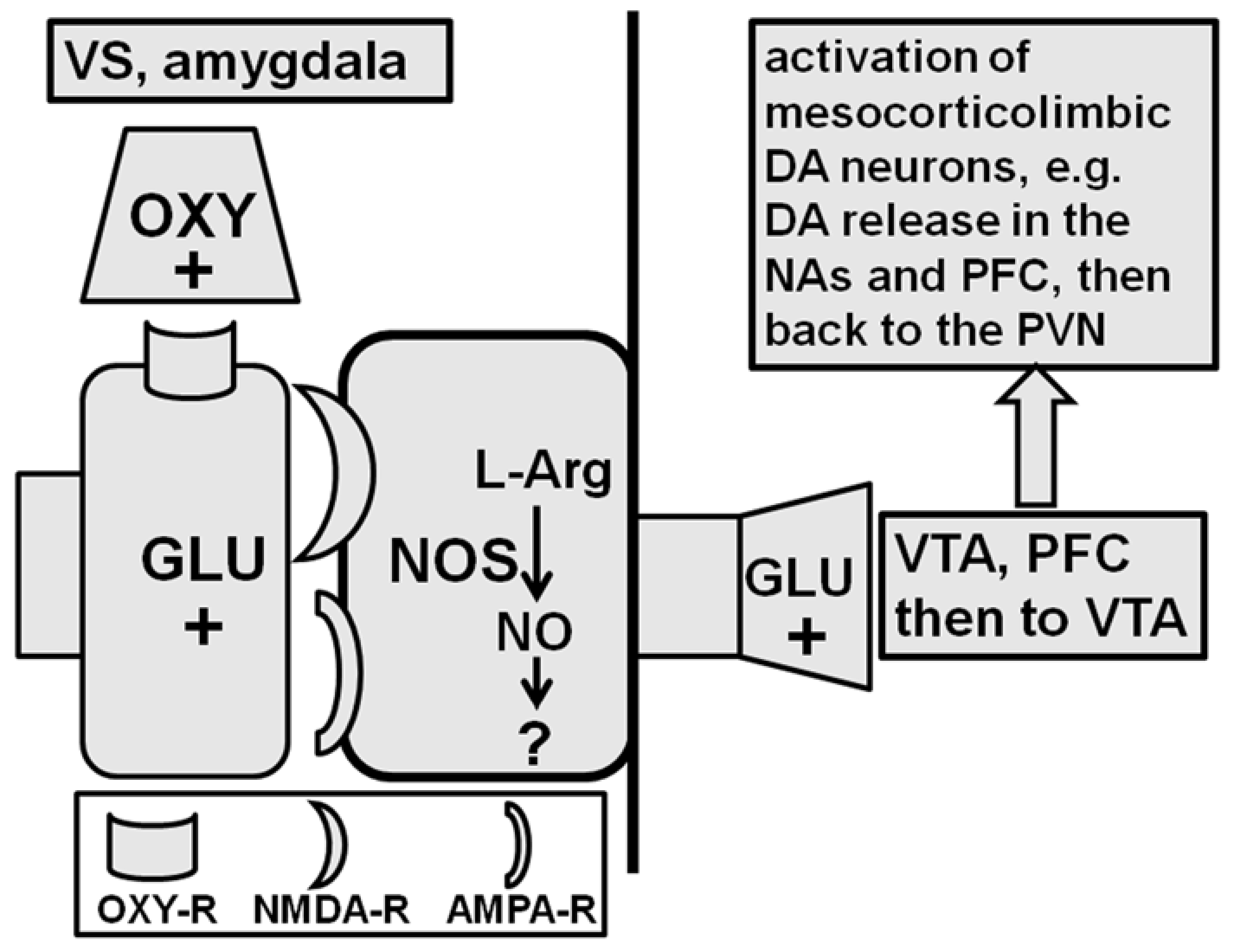{kind=link}
{kind=link}
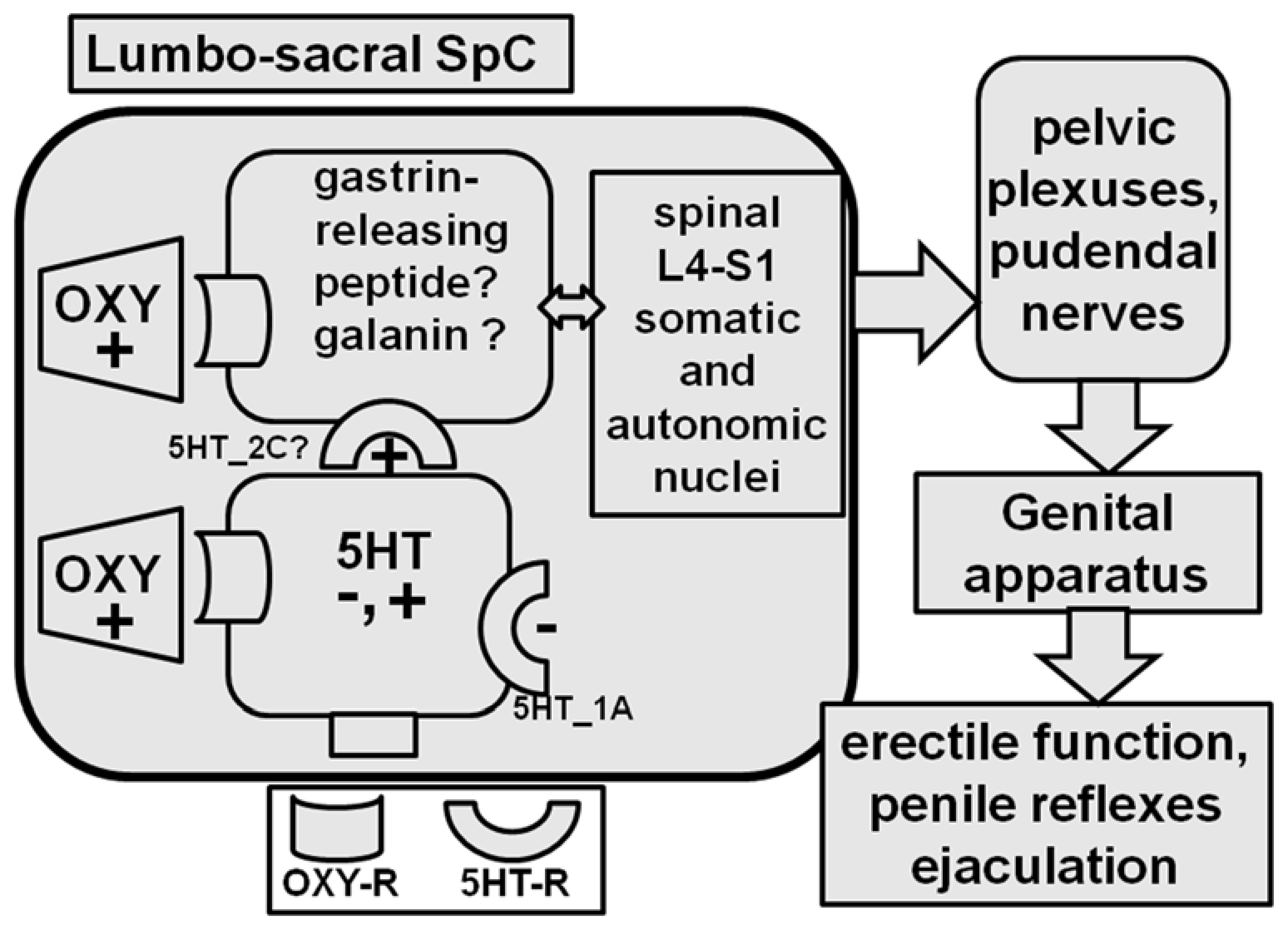{kind=link}
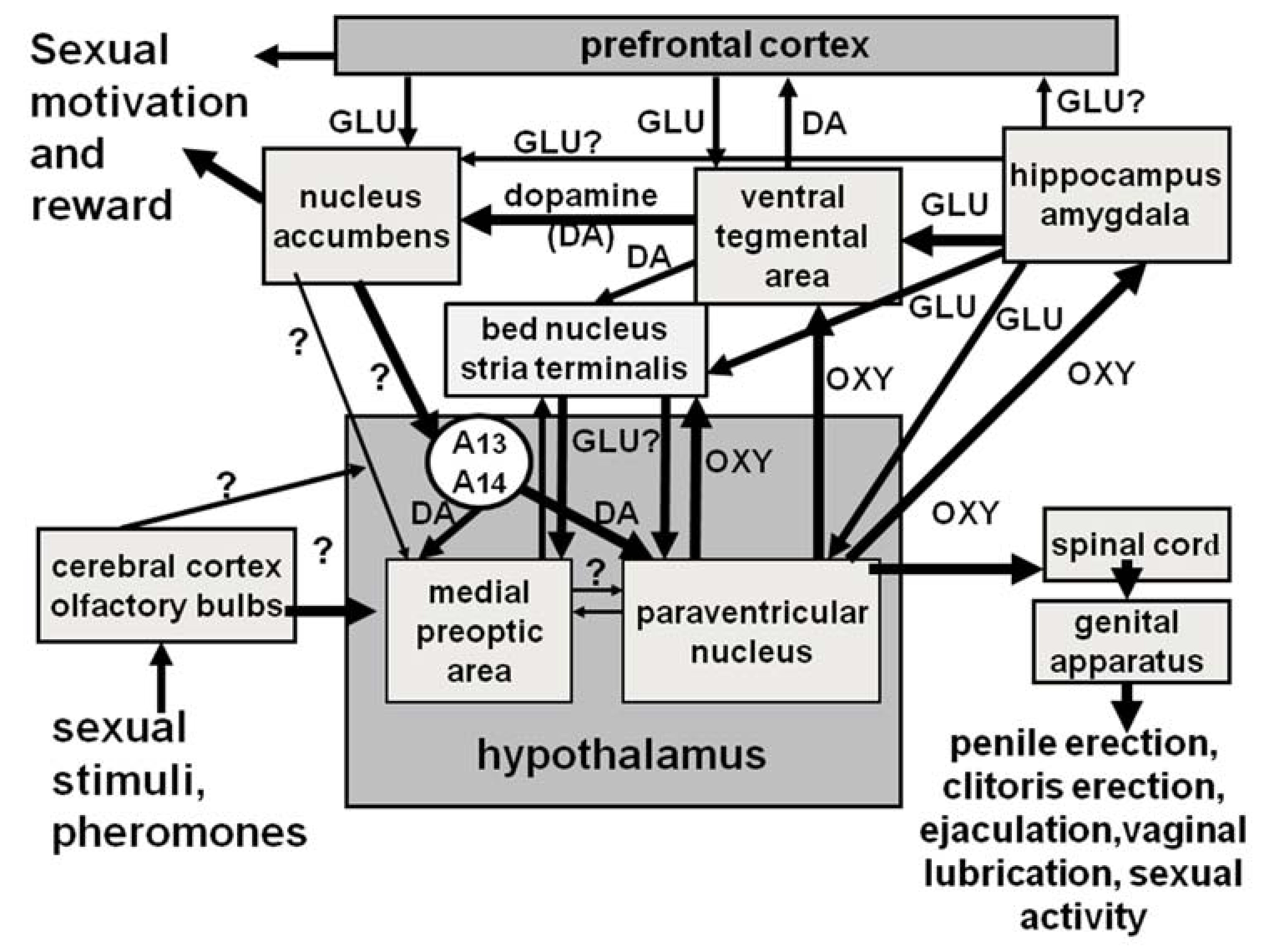{kind=link}
| Peptide, Non-Peptide | Amino Acid Sequence/Chemical Formula | Action on Oxytocin Receptors | Effect on Erectile Function/Sexual Activity |
|---|---|---|---|
| oxytocin | H-Cys-Tyr-Ile-Gln-Asn-Cys-Pro-Leu-GlyNH2 | agonist | facilitatory |
| Arg8-vasopressin | H-Cys-Tyr-Phe-Gln--Asn-Cys-Pro-Arg-GlyNH2 | agonist at 100 times higher doses | none |
| d(CH2)5Tyr(Me)2-Orn8- vasotocin | Mpr-Tyr(Me)-Ile-Gln--Asn-Cys-Pro-Orn-GlyNH2 | antagonist | inhibitory |
| Pen1-Phe(Me)2-Tyr4- Orn8-oxytocin | Pen-Phe(Me)-Ile-Tyr--Asn-Cys-Pro-Orn-GlyNH2 | antagonist | inhibitory |
| d(CH2)5Tyr(Me)2-Arg8- vasopressin | Mpr-Tyr(Me)-Ile-Gln--Asn-Cys-Pro-Arg-GlyNH2 | antagonist at high doses | inhibitory at high doses |
| L-383,899 | 1-((7,7-dimethyl-2(S)-(2(S)-amino-4-(methylsulfonyl) butyramido)bicyclo[2.2.1]-heptan-1(S)-yl)methyl) sulfonyl)-4-(2-methylphenyl)piperazine | antagonist | NA |
| epelsiban (GLK 557296) | (3R,6R)-3-(2,3-dihydro-1H-inden-2-yl)-1-[(1R)-1-(2,6-dimethyl-3- pyridinyl)-2-(4-morpholinyl)-2-oxoethyl]-6-[(1S)-1-methylpropyl]-2,5-piperazinedione | antagonist | NA * |
| cligosiban | 5-{3-[3-(2-chloro-4-fluorophenoxy)azetidin-1-yl]-5-(methoxy methyl)-4H-1,2,4-triazol-4-yl}-2-methoxypyridine | antagonist | NA * |
| Neurotransmitter, Neuropeptide | Receptor | Mechanism of Action |
|---|---|---|
| oxytocin | Uterine type | intracellular Ca2+ increase and NOS activation, no GC-cGMP pathway involvement |
| dopamine | D2 and D4 subtype | intracellular Ca2+ increase and NOS activation, no GC-cGMP pathway involvement |
| glutamic acid | NMDA, some role of AMPA receptors cannot be ruled out | intracellular Ca2+ increase and NOS activation, no GC-cGMP pathway involvement |
| GABA | GABA-A | reduction in intracellular Ca2+ increase and NOS activation |
| opioid peptides | µ subtype | reduction in intracellular Ca2+ increase and NOS activation |
| endocannabinoids | CB1 | reduction in GLU transmission, intracellular Ca2+ increase and NOS activation |
| hexarelin peptides | apparently different from the GHS receptor | intracellular Ca2+ increase and NOS activation, no GC-cGMP pathway involvement |
| C-terminal VGF peptides | unknown | intracellular Ca2+ increase and NOS activation, no GC-cGMP pathway involvement |
| Neurotransmitter, Neuropeptide | Effect on Sexual Function | Brain Areas Involved | Main Interaction |
|---|---|---|---|
| oxytocin | facilitatory | PVN, MPOA, VMN, BNST, VTA, VS, amygdala, nucleus para-giganto cellularis, SpC | activation of PVN oxytocinergic neurons; activation of mesolimbic dopaminergic neurons in the VTA; activation of glutamatergic neurons in the VS, amygdala, BNST; activation of pro-erectile neurons in the thoraco-lumbar SpC; activation of serotoninergic neurons projecting to the SpC; activation of MPOA and VMN leading to lordosis in females |
| dopamine | facilitatory | PVN, BNST, VTA, SN, NAs | activation of PVN oxytocinergic neurons; activation of neurons in the NAs projecting to the PVN; modulation of motor activity at SN level (see [263]). |
| serotonin | Inhibitory, Facilitatory * | SpC | activation/inhibiton* of oxytocinergic neurotransmission in the SpC |
| glutamic acid | facilitatory | PVN, BNST, VTA, VS, MPFC amygdala | activation of PVN oxytocinergic neurons; activation of mesolimbic dopaminergic neurons in the VTA and of glutamatergic neurons in the BNST, VS and amygdala |
| GABA | Inhibitory | PVN, SpC | inhibition of PVN oxytocinergic neurons |
| nitric oxide | facilitatory | PVN, BNST, VTA, VS, amygdala | activation of PVN oxytocinergic neurons; activation of mesolimbic dopaminergic neurons in the VTA; activation of glutamatergic neurons in the VS, amygdala and BNST |
| ACTH-MSH peptides | facilitatory | APVH | NAs (but see [66,67]) |
| opioid peptides | inhibitory | PVN | inhibition of PVN oxytocinergic neurons by stimulation of opioid receptors of the µ subtype |
| hexarelin related peptides | facilitatory | PVN | activation of PVN oxytocinergic neurons, possibly by receptors different from those mediating growth hormone release |
| C-terminal VGF peptides | facilitatory | PVN | activation of PVN oxytocinergic neurons by receptors yet to be identified. |
| endocannabinoids | inhibitory | PVN | inhibition of PVN oxytocinergic neurons secondary to the inhibition of glutamic acid neurotransmission by activation of cannabinoid receptors of the CB1 subtype. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melis, M.R.; Argiolas, A. Oxytocin, Erectile Function and Sexual Behavior: Last Discoveries and Possible Advances. Int. J. Mol. Sci. 2021, 22, 10376. https://doi.org/10.3390/ijms221910376
Melis MR, Argiolas A. Oxytocin, Erectile Function and Sexual Behavior: Last Discoveries and Possible Advances. International Journal of Molecular Sciences. 2021; 22(19):10376. https://doi.org/10.3390/ijms221910376
Chicago/Turabian StyleMelis, Maria Rosaria, and Antonio Argiolas. 2021. "Oxytocin, Erectile Function and Sexual Behavior: Last Discoveries and Possible Advances" International Journal of Molecular Sciences 22, no. 19: 10376. https://doi.org/10.3390/ijms221910376
APA StyleMelis, M. R., & Argiolas, A. (2021). Oxytocin, Erectile Function and Sexual Behavior: Last Discoveries and Possible Advances. International Journal of Molecular Sciences, 22(19), 10376. https://doi.org/10.3390/ijms221910376