
Duration of Social Isolation Affects Production of Nitric Oxide in the Rat Brain

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
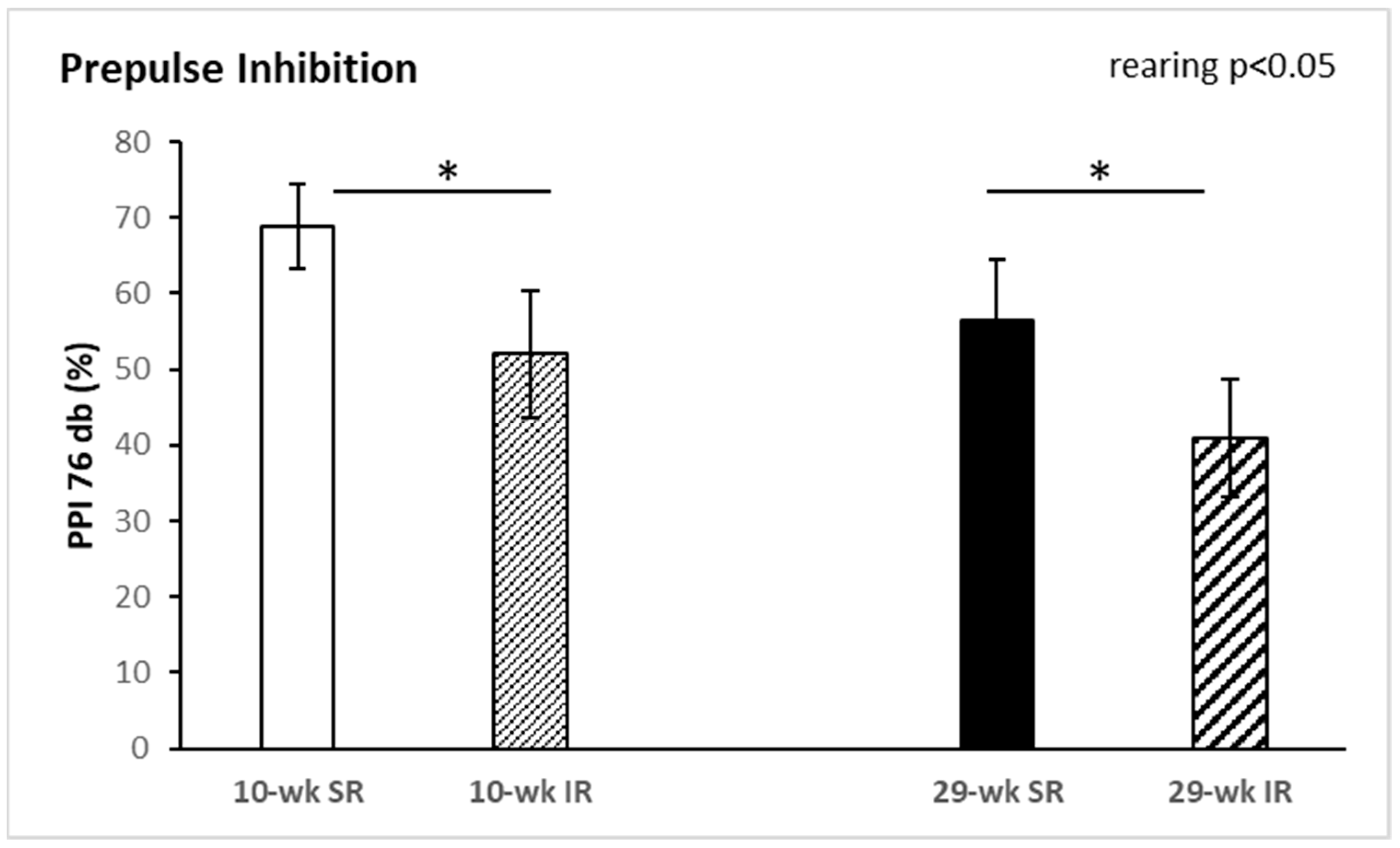
2.1. Behavioral Tests
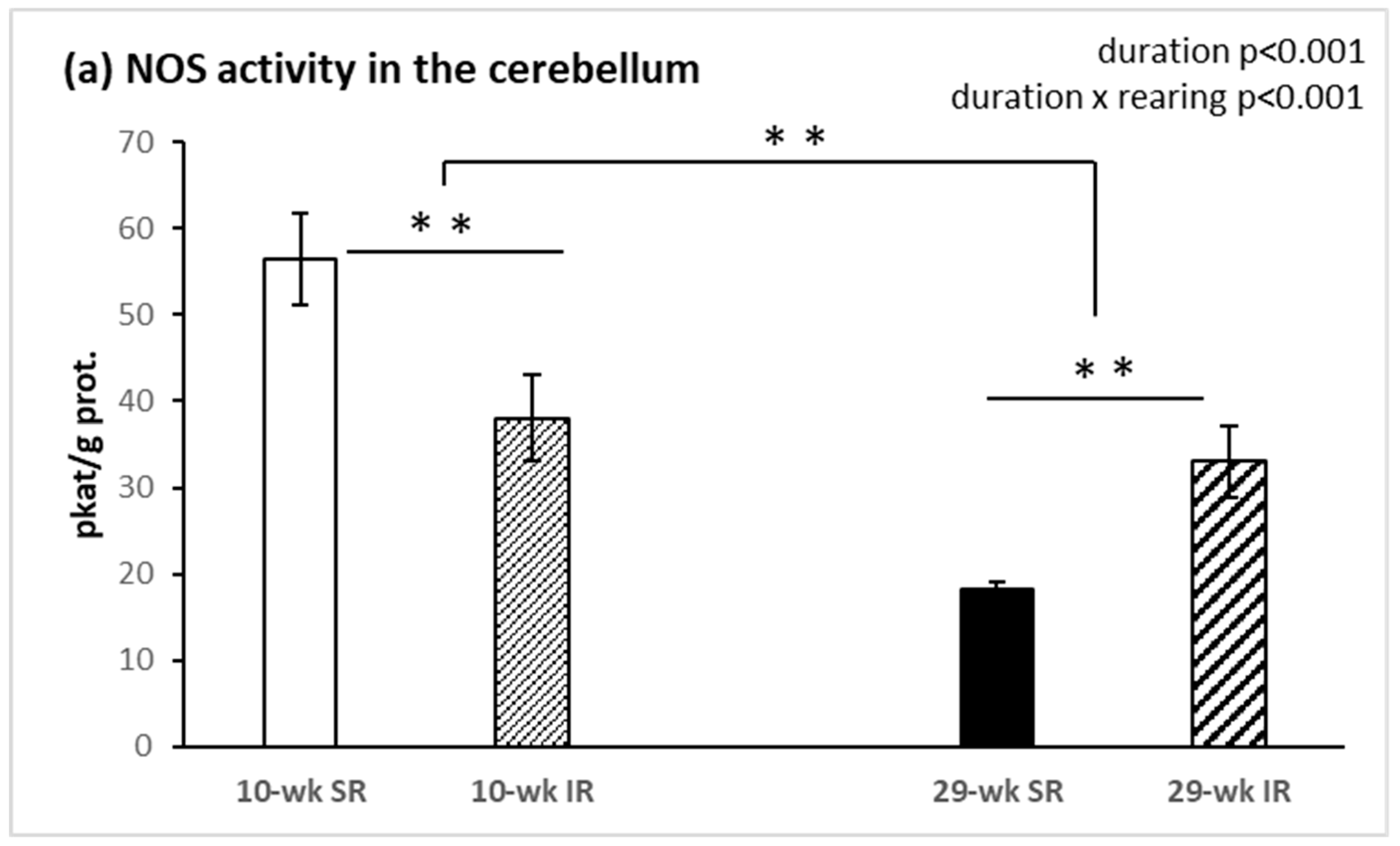
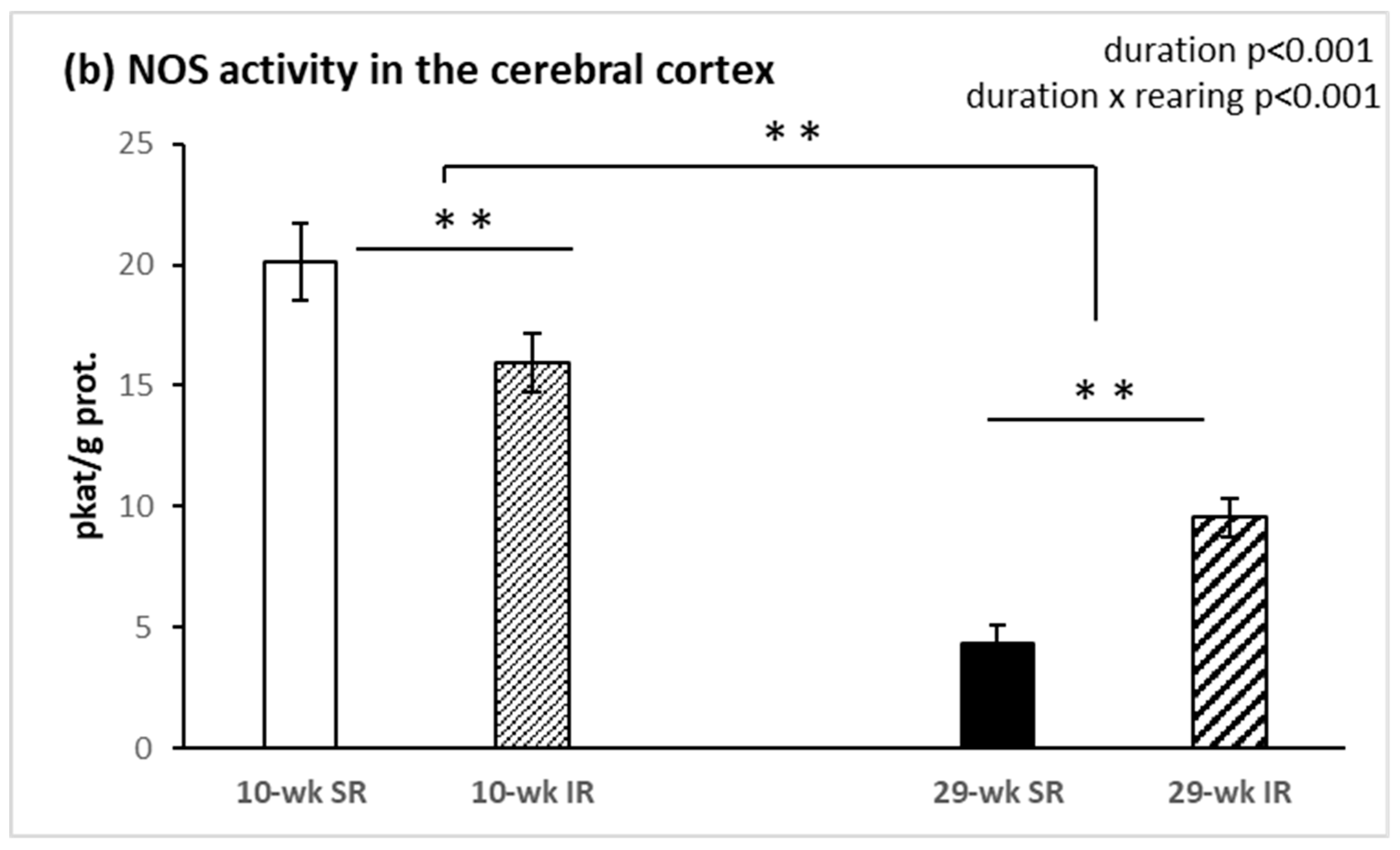
2.2. Total Activity of NOS
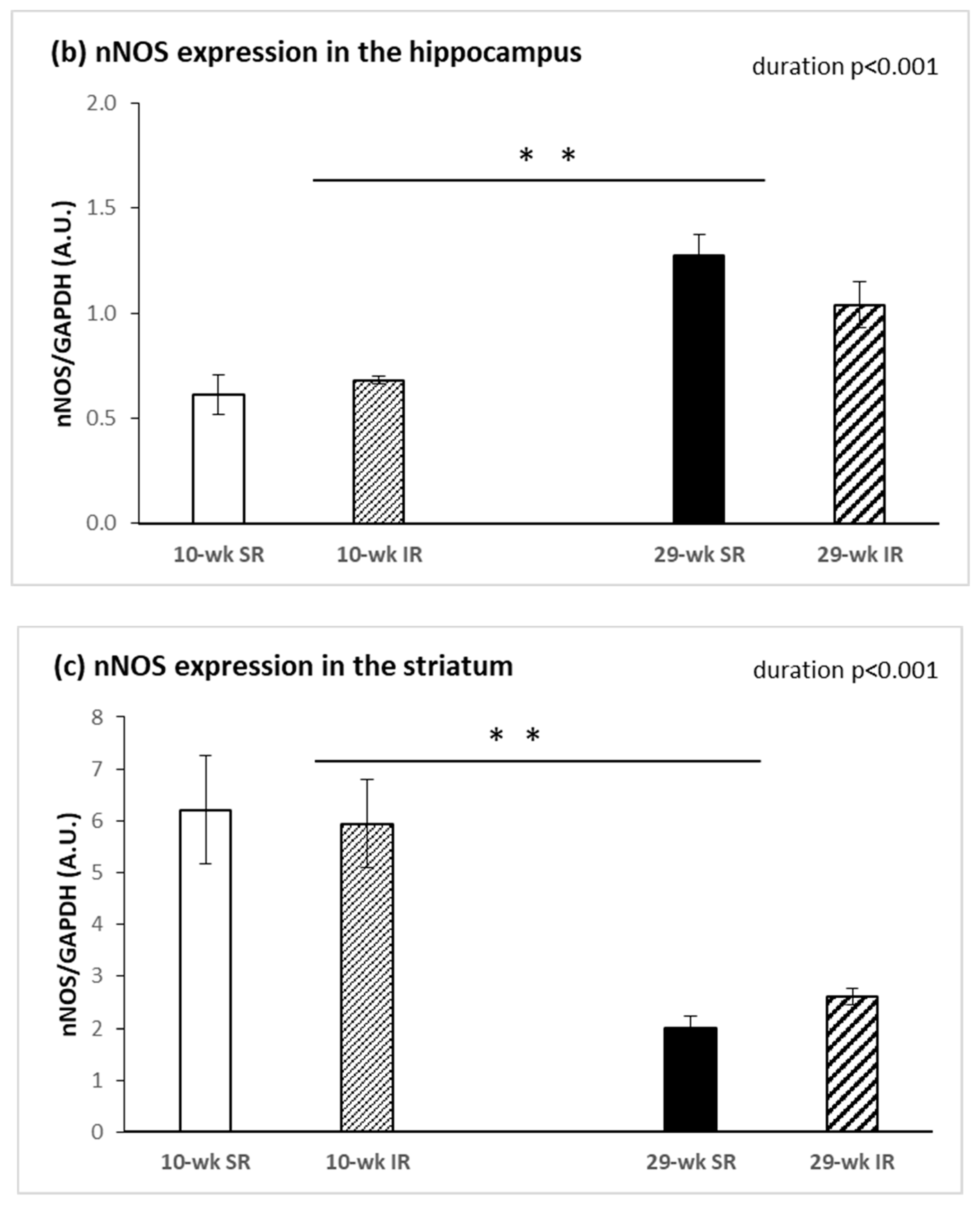
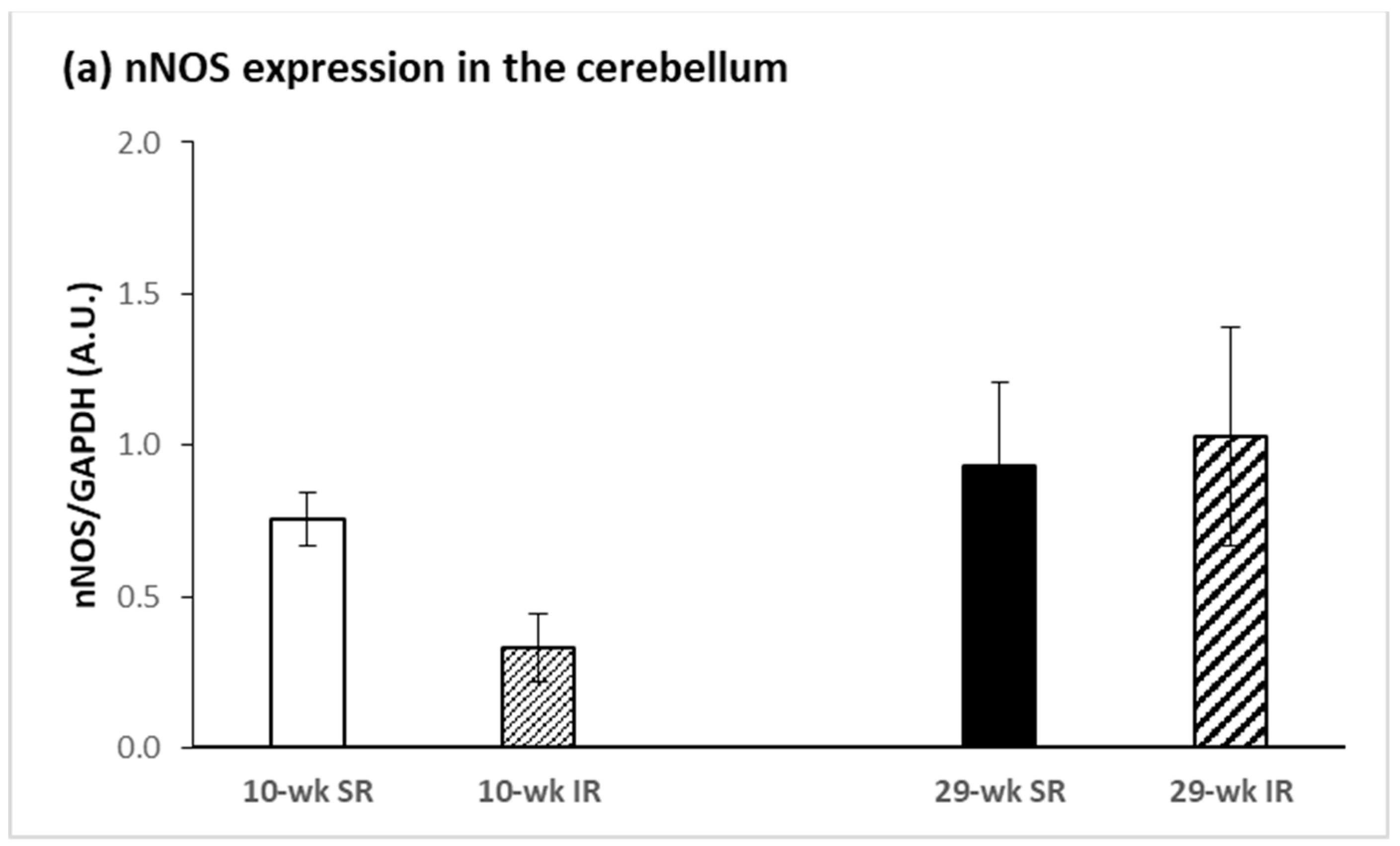
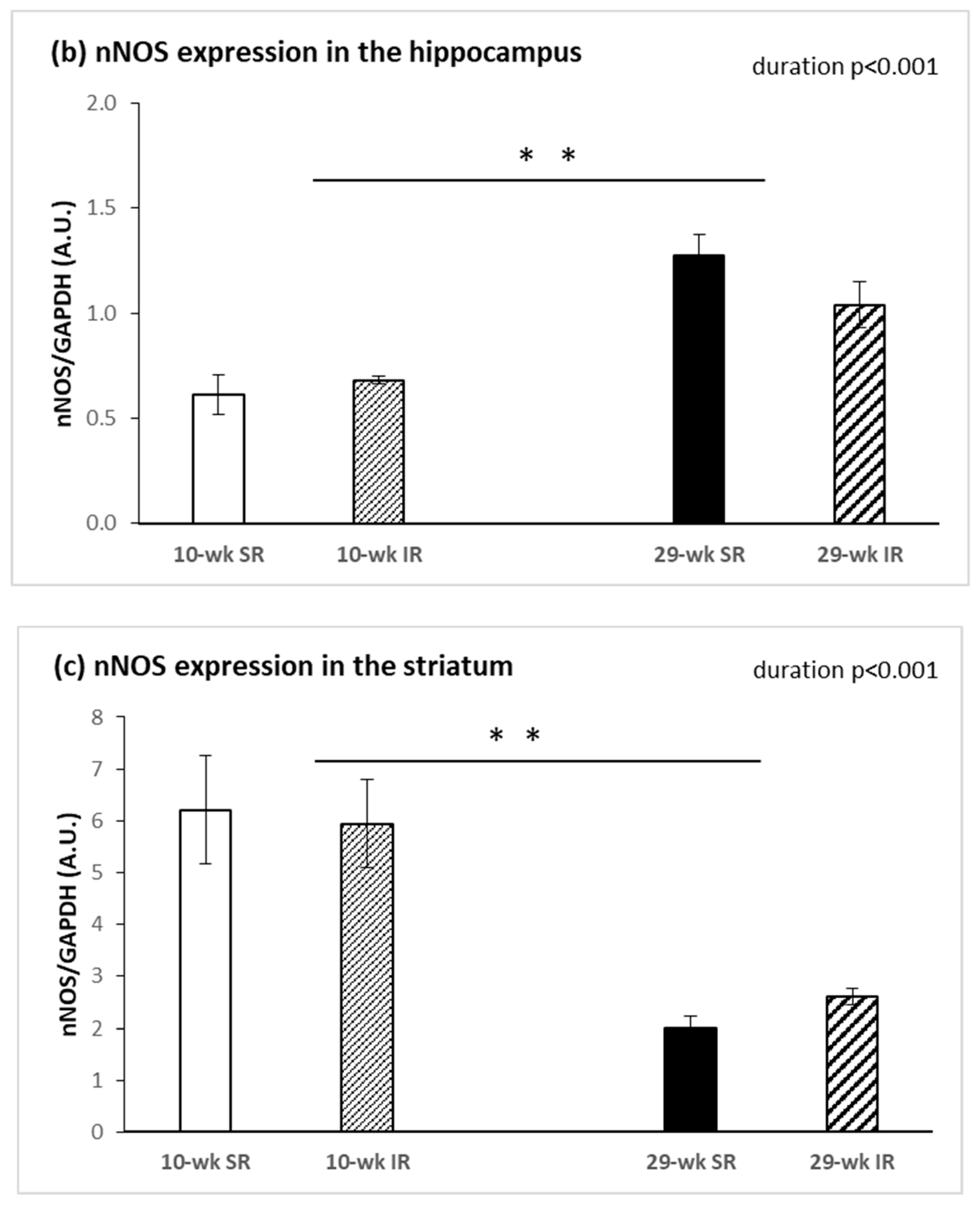
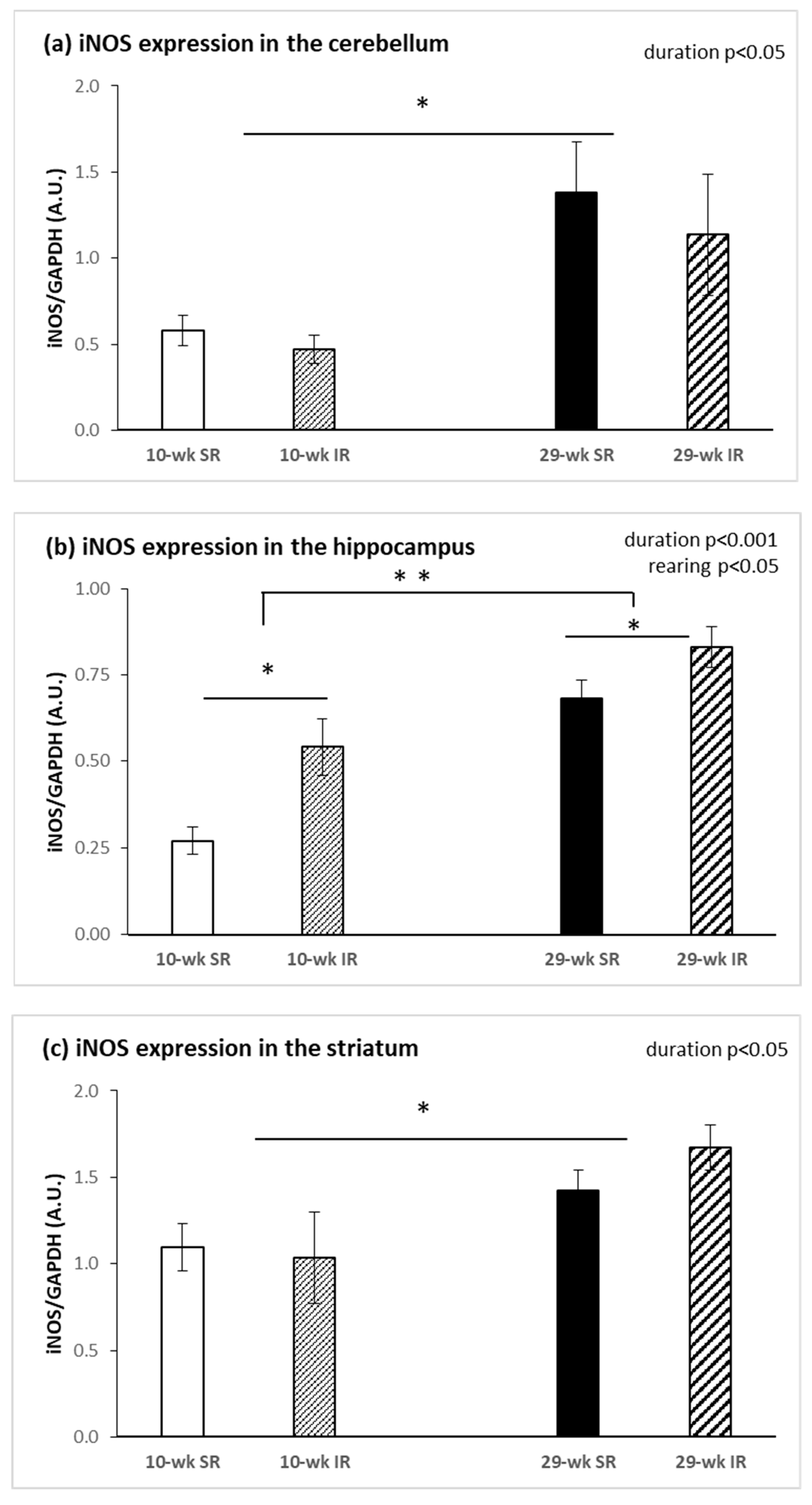
2.3. Protein Expression of nNOS and iNOS
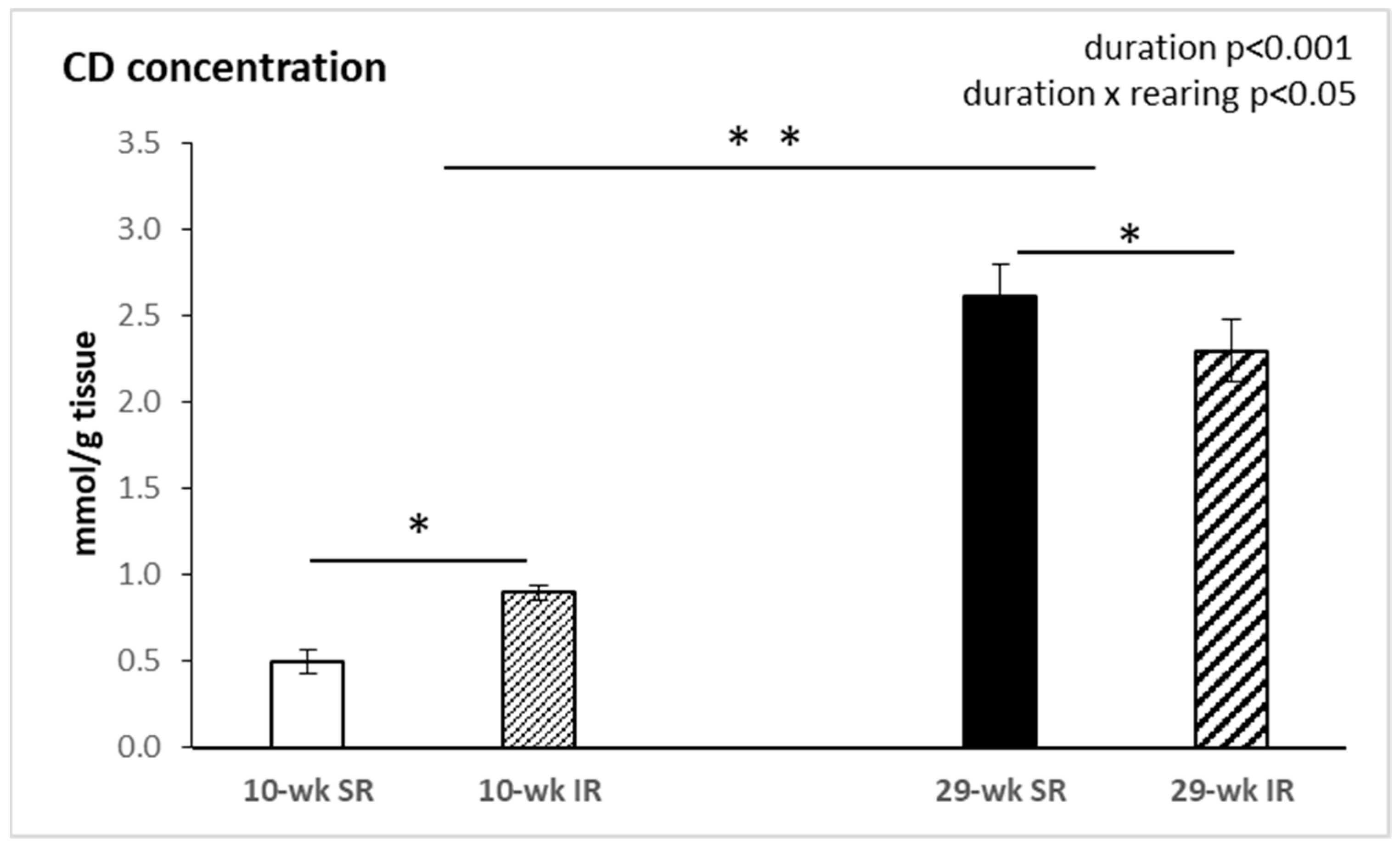
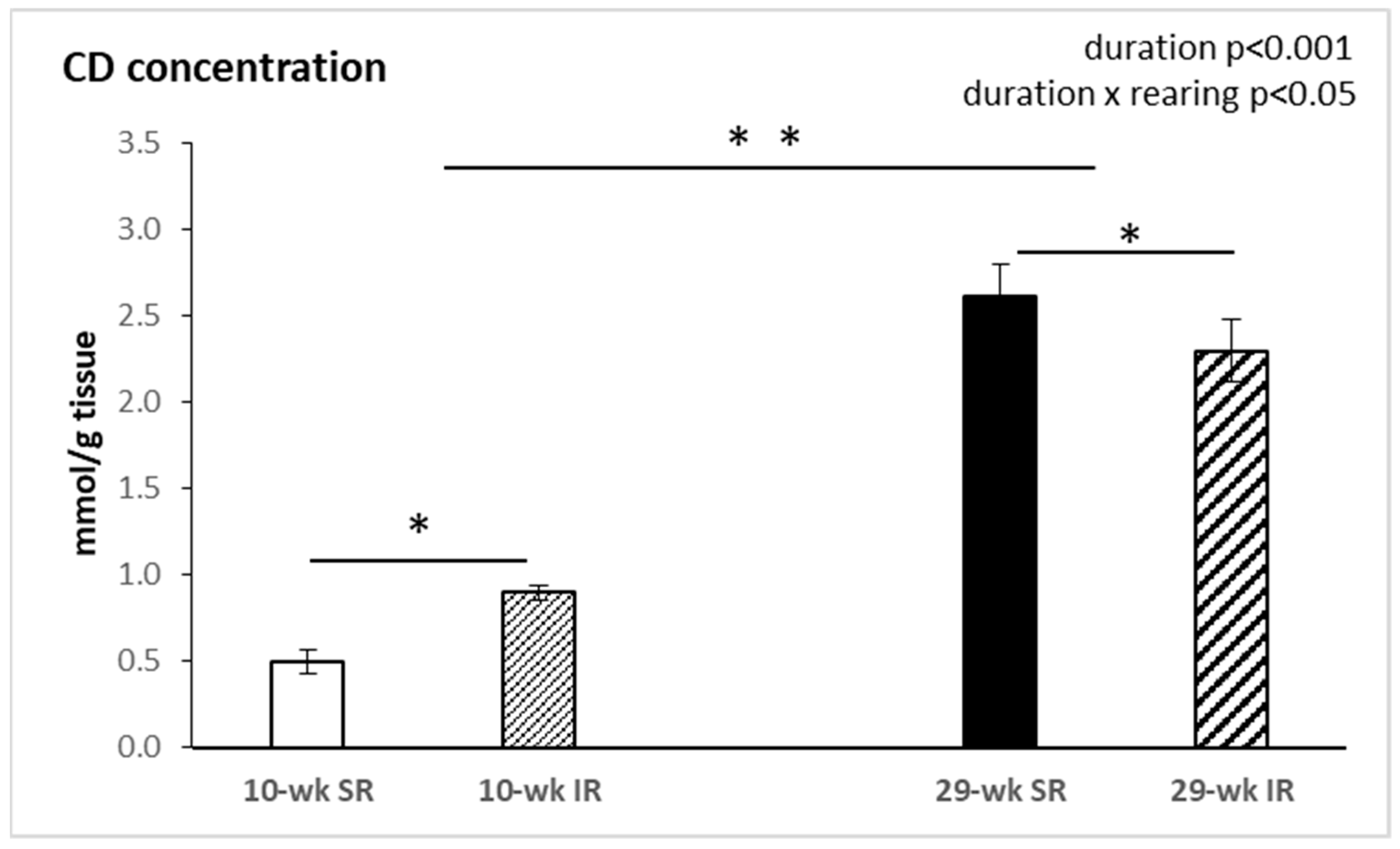
2.4. Conjugated Dienes Concentration
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Social Isolation
4.3. Behavioral Testing
4.3.1. Open-Field Test
4.3.2. Prepulse Inhibition of Startle (PPI) Paradigm
4.4. Nitric Oxide Synthase Activity Assay
4.5. Western Blot Analysis
4.6. Concentration of Conjugated Dienes
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Veenema, A.H.; Reber, S.O.; Selch, S.; Obermeier, F.; Neumann, I.D. Early Life Stress Enhances the Vulnerability to Chronic Psychosocial Stress and Experimental Colitis in Adult Mice. Endocrinology 2008, 149, 2727–2736. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.; Umemori, H. The best-laid plans go oft awry: Synaptogenic growth factor signaling in neuropsychiatric disease. Front. Synaptic Neurosci. 2014, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Mumtaz, F.; Khan, M.I.; Zubair, M.; Dehpour, A.R. Neurobiology and consequences of social isolation stress in animal model—A comprehensive review. Biomed. Pharmacother. 2018, 105, 1205–1222. [Google Scholar] [CrossRef]
- Schubert, M.; Porkess, M.; Dashdorj, N.; Fone, K.; Auer, D. Effects of social isolation rearing on the limbic brain: A combined behavioral and magnetic resonance imaging volumetry study in rats. Neuroscience 2009, 159, 21–30. [Google Scholar] [CrossRef]
- Cirulli, F.; Berry, A.; Bonsignore, L.T.; Capone, F.; D’Andrea, I.; Aloe, L.; Branchi, I.; Alleva, E. Early life influences on emotional reactivity: Evidence that social enrichment has greater effects than handling on anxiety-like behaviors, neuroendocrine responses to stress and central BDNF levels. Neurosci. Biobehav. Rev. 2010, 34, 808–820. [Google Scholar] [CrossRef]
- Malkesman, O.; Maayan, R.; Weizman, A.; Weller, A. Aggressive behavior and HPA axis hormones after social isolation in adult rats of two different genetic animal models for depression. Behav. Brain Res. 2006, 175, 408–414. [Google Scholar] [CrossRef]
- Ko, C.-Y.; Liu, Y.-P. Disruptions of sensorimotor gating, cytokines, glycemia, monoamines, and genes in both sexes of rats reared in social isolation can be ameliorated by oral chronic quetiapine administration. Brain Behav. Immun. 2016, 51, 119–130. [Google Scholar] [CrossRef]
- Han, X.; Wang, W.; Shao, F.; Li, N. Isolation rearing alters social behaviors and monoamine neurotransmission in the medial prefrontal cortex and nucleus accumbens of adult rats. Brain Res. 2011, 1385, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Yan, G.; Xuan, Y.; Peng, H.; Huang, Q.-J.; Wu, R.; Xu, H. Chronic social isolation decreases glutamate and glutamine levels and induces oxidative stress in the rat hippocampus. Behav. Brain Res. 2015, 282, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Murínová, J.; Hlaváčová, N.; Chmelová, M.; Riečanský, I. The Evidence for Altered BDNF Expression in the Brain of Rats Reared or Housed in Social Isolation: A Systematic Review. Front. Behav. Neurosci. 2017, 11, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toth, M.; Mikics, E.; Tulogdi, A.; Aliczki, M.; Haller, J. Post-weaning social isolation induces abnormal forms of aggression in conjunction with increased glucocorticoid and autonomic stress responses. Horm. Behav. 2011, 60, 28–36. [Google Scholar] [CrossRef]
- Amiri, S.; Haj-Mirzaian, A.; Rahimi-Balaei, M.; Razmi, A.; Kordjazy, N.; Shirzadian, A.; Mehr, S.E.; Sianati, H.; Dehpour, A.R. Co-occurrence of anxiety and depressive-like behaviors following adolescent social isolation in male mice; possible role of nitrergic system. Physiol. Behav. 2015, 145, 38–44. [Google Scholar] [CrossRef]
- Brenman, J.E.; Bredt, D.S. Synaptic signaling by nitric oxide. Curr. Opin. Neurobiol. 1997, 7, 374–378. [Google Scholar] [CrossRef]
- Nikonenko, I.; Boda, B.; Steen, S.; Knott, G.W.; Welker, E.; Muller, D. PSD-95 promotes synaptogenesis and multiinnervated spine formation through nitric oxide signaling. J. Cell Biol. 2008, 183, 1115–1127. [Google Scholar] [CrossRef] [PubMed]
- Nott, A.; Nitarska, J.; Veenvliet, J.V.; Schacke, S.; Derijck, A.A.H.A.; Sirko, P.; Muchardt, C.; Pasterkamp, J.; Smidt, M.; Riccio, A. S-nitrosylation of HDAC2 regulates the expression of the chromatin-remodeling factor Brm during radial neuron migration. Proc. Natl. Acad. Sci. USA 2013, 110, 3113–3118. [Google Scholar] [CrossRef] [Green Version]
- Huang, P.L.; Dawson, T.M.; Bredt, D.S.; Snyder, S.H.; Fishman, M.C. Targeted disruption of the neuronal nitric oxide synthase gene. Cell 1993, 75, 1273–1286. [Google Scholar] [CrossRef]
- Hardingham, N.; Dachtler, J.; Fox, K. The role of nitric oxide in pre-synaptic plasticity and homeostasis. Front. Cell. Neurosci. 2013, 7, 190. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, H.-G.; Bogerts, B.; Keilhoff, G. The many faces of nitric oxide in schizophrenia. A review. Schizophr. Res. 2005, 78, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Steinert, J.R.; Chernova, T.; Forsythe, I. Nitric Oxide Signaling in Brain Function, Dysfunction, and Dementia. Neuroscientist 2010, 16, 435–452. [Google Scholar] [CrossRef]
- Joca, S.R.L.; Sartim, A.G.; Roncalho, A.L.; Diniz, C.F.; Wegener, G. Nitric oxide signalling and antidepressant action revisited. Cell Tissue Res. 2019, 377, 45–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigo, J.; Alonso, D.; Fernández, A.P.; Serrano, J.; Richart, A.; López, J.C.; Santacana, M.; Martínez-Murillo, R.; Bentura, M.L.; Ghiglione, M.; et al. Neuronal and inducible nitric oxide synthase expression and protein nitration in rat cerebellum after oxygen and glucose deprivation. Brain Res. 2001, 909, 20–45. [Google Scholar] [CrossRef]
- Sochocka, M.; Diniz, B.; Leszek, J. Inflammatory Response in the CNS: Friend or Foe? Mol. Neurobiol. 2017, 54, 8071–8089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.-G.; Hua, Y.; Hu, M.; Luo, C.-X.; Han, X.; Zhu, X.-J.; Wang, B.; Xu, J.; Zhu, D.-Y. Neuronal nitric oxide synthase contributes to chronic stress-induced depression by suppressing hippocampal neurogenesis. J. Neurochem. 2007, 103, 1843–1854. [Google Scholar] [CrossRef] [PubMed]
- Lauer, M.; Johannes, S.; Fritzen, S.; Senitz, D.; Riederer, P.; Reif, A. Morphological Abnormalities in Nitric-Oxide-Synthase-Positive Striatal Interneurons of Schizophrenic Patients. Neuropsychobiology 2005, 52, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Nasyrova, R.F.; Ivashchenko, D.; Ivanov, M.V.; Neznanov, N.G. Role of nitric oxide and related molecules in schizophrenia pathogenesis: Biochemical, genetic and clinical aspects. Front. Physiol. 2015, 6, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakshi, V.P.; Geyer, M.A. Ontogeny of Isolation Rearing-Induced Deficits in Sensorimotor Gating in Rats. Physiol. Behav. 1999, 67, 385–392. [Google Scholar] [CrossRef]
- Chmelova, M.; Balagova, L.; Marko, M.; Vrankova, S.; Cebova, M.; Jezova, D.; Riečanský, I.; Hlavacova, N. Behavioral alterations induced by post-weaning isolation rearing of rats are accompanied by reduced VGF/BDNF/TrkB signaling in the hippocampus. Neurochem. Int. 2019, 129, 104473. [Google Scholar] [CrossRef] [PubMed]
- Klamer, D.; Engel, J.A.; Svensson, L. The neuronal selective nitric oxide synthase inhibitor, Nω-propyl-l-arginine, blocks the effects of phencyclidine on prepulse inhibition and locomotor activity in mice. Eur. J. Pharmacol. 2004, 503, 103–107. [Google Scholar] [CrossRef]
- Fejgin, K.; Pålsson, E.; Wass, C.; Svensson, L.; Klamer, D. Nitric Oxide Signaling in the Medial Prefrontal Cortex is Involved in the Biochemical and Behavioral Effects of Phencyclidine. Neuropsychopharmacology 2008, 33, 1874–1883. [Google Scholar] [CrossRef] [Green Version]
- Issy, A.C.; Pedrazzi, J.F.C.; Yoneyama, B.H.; Del-Bel, E.A. Critical role of nitric oxide in the modulation of prepulse inhibition in Swiss mice. Psychopharmacology 2014, 231, 663–672. [Google Scholar] [CrossRef]
- Ledo, A.; Frade, J.; Barbosa, R.M.; Laranjinha, J. Nitric oxide in brain: Diffusion, targets and concentration dynamics in hippocampal subregions. Mol. Asp. Med. 2004, 25, 75e89. [Google Scholar] [CrossRef] [Green Version]
- Moncada, S.; Bolanos, J.P. Nitric oxide, cell bioenergetics and neurodegeneration. J. Neurochem. 2006, 97, 1676–1689. [Google Scholar] [CrossRef] [PubMed]
- Issy, A.; Lazzarini, M.; Szawka, R.; Carolino, R.; Anselmo-Franci, J.; Del Bel, E. Nitric oxide synthase inhibitors improve prepulse inhibition responses of Wistar rats. Behav. Brain Res. 2011, 217, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, N.R.; Geyer, M.A. Clozapine and haloperidol in an animal model of sensorimotor gating deficits in schizophrenia. Pharmacol. Biochem. Behav. 1993, 44, 741–744. [Google Scholar] [CrossRef]
- Braff, D.L.; Geyer, M.A.; Swerdlow, N.R. Human studies of prepulse inhibition of startle: Normal subjects, patient groups, and pharmacological studies. Psychopharmacology 2001, 156, 234–258. [Google Scholar] [CrossRef] [PubMed]
- Khoja, S.; Asatryan, L.; Jakowec, M.W.; Davies, D.L. Dopamine Receptor Blockade Attenuates Purinergic P2X4 Receptor-Mediated Prepulse Inhibition Deficits and Underlying Molecular Mechanisms. Front. Cell. Neurosci. 2019, 13, 331. [Google Scholar] [CrossRef] [Green Version]
- Ralph, R.J.; Caine, S.B. Dopamine D1 and D2 Agonist Effects on Prepulse Inhibition and Locomotion: Comparison of Sprague-Dawley Rats to Swiss-Webster, 129X1/SvJ, C57BL/6J, and DBA/2J Mice. J. Pharmacol. Exp. Ther. 2005, 312, 733–741. [Google Scholar] [CrossRef]
- Komeima, K.; Hayashi, Y.; Naito, Y.; Watanabe, Y. Inhibition of Neuronal Nitric-oxide Synthase by Calcium/Calmodulin-dependent Protein Kinase IIα through Ser847 Phosphorylation in NG108-15 Neuronal Cells. J. Biol. Chem. 2000, 275, 28139–28143. [Google Scholar] [CrossRef] [Green Version]
- Tanda, K.; Nishi, A.; Matsuo, N.; Nakanishi, K.; Yamasaki, N.; Sugimoto, T.; Toyama, K.; Takao, K.; Miyakawa, T. Abnormal social behavior, hyperactivity, impaired remote spatial memory, and increased D1-mediated dopaminergic signaling in neuronal nitric oxide synthase knockout mice. Mol. Brain 2009, 2, 19. [Google Scholar] [CrossRef] [Green Version]
- Zlatković, J.; Filipović, D. Chronic social isolation induces NF-κB activation and upregulation of iNOS protein expression in rat prefrontal cortex. Neurochem. Int. 2013, 63, 172–179. [Google Scholar] [CrossRef]
- Zlatković, J.; Bernardi, R.E.; Filipović, D. Protective effect of Hsp70i against chronic social isolation stress in the rat hippocampus. J. Neural Transm. 2014, 121, 3–14. [Google Scholar] [CrossRef]
- Madrigal, J.; Bueno, B.G.; Caso, J.; Perez-Nievas, B.G.; Leza, J.C. Stress-Induced Oxidative Changes in Brain. CNS Neurol. Disord. Drug Targets 2006, 5, 561–568. [Google Scholar] [CrossRef]
- Zlatković, J.; Filipović, D. Bax and B-cell-lymphoma 2 mediate proapoptotic signaling following chronic isolation stress in rat brain. Neuroscience 2012, 223, 238–245. [Google Scholar] [CrossRef]
- Filipović, D.; Zlatković, J.; Inta, D.; Bjelobaba, I.; Stojiljkovic, M.; Gass, P. Chronic isolation stress predisposes the frontal cortex but not the hippocampus to the potentially detrimental release of cytochrome c from mitochondria and the activation of caspase-3. J. Neurosci. Res. 2011, 89, 1461–1470. [Google Scholar] [CrossRef]
- Jiang, B.; Liang, P.; Deng, G.; Tu, Z.; Liu, M.; Xiao, X. Increased stability of Bcl-2 in HSP70-mediated protection against apoptosis induced by oxidative stress. Cell Stress Chaperon 2011, 16, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Didelot, C.; Schmitt, E.; Brunet, M.; Maingret, L.; Parcellier, A.; Garrido, C. Heat Shock Proteins: Endogenous Modulators of Apoptotic Cell Death. Handb. Exp. Pharmacol. 2006, 26, 171–198. [Google Scholar] [CrossRef]
- Kumar, Y.; Tatu, U. Stress protein flux during recovery from simulated ischemia: Induced heat shock protein 70 confers cytoprotection by suppressing JNK activation and inhibiting apoptotic cell death. Proteomics 2003, 3, 513–526. [Google Scholar] [CrossRef] [PubMed]
- Kleppisch, T.; Feil, R. cGMP Signalling in the Mammalian Brain: Role in Synaptic Plasticity and Behaviour. Handb. Exp. Pharmacol. 2009, 191, 549–579. [Google Scholar] [CrossRef]
- Contestabile, A. Role of Nitric Oxide in Cerebellar Development and Function: Focus on Granule Neurons. Cerebellum 2012, 11, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Keinanen, K.; Wisden, W.; Sommer, B.; Werner, P.; Herb, A.; Verdoorn, T.; Sakmann, B.; Seeburg, P. A family of AMPA-selective glutamate receptors. Science 1990, 249, 556–560. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhu, Z.-A. Nitric oxide show its survival role by NO-PKC pathway through cGMP-dependent or independent on the culture of cerebella granular neurons. Neurosci. Lett. 2014, 583, 165–169. [Google Scholar] [CrossRef]
- Sammut, S.; Park, D.J.; West, A.R. Frontal cortical afferents facilitate striatal nitric oxide transmission in vivo via a NMDA receptor and neuronal NOS-dependent mechanism. J. Neurochem. 2007, 103, 1145–1156. [Google Scholar] [CrossRef]
- Rossetti, Z.L.; Crespi, F. Inhibition of nitric oxide release in vivo by ethanol. Alcohol. Clin. Exp. Res. 2004, 28, 1746–1751. [Google Scholar] [CrossRef]
- Hoque, K.E.; Indorkar, R.P.; Sammut, S.; West, A.R. Impact of dopamine–glutamate interactions on striatal neuronal nitric oxide synthase activity. Psychopharmacology 2010, 207, 571–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munhoz, C.; Bueno, B.G.; Madrigal, J.; Lepsch, L.; Scavone, C.; Leza, J. Stress-induced neuroinflammation: Mechanisms and new pharmacological targets. Braz. J. Med. Biol. Res. 2008, 41, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Shal, B.; Khan, A.; Khan, A.; Ullah, R.; Ali, G.; Islam, S.; Haq, I.; Ali, H.; Seo, E.-K.; Khan, S. Alleviation of Memory Deficit by Bergenin via the Regulation of Reelin and Nrf-2/NF-κB Pathway in Transgenic Mouse Model. Int. J. Mol. Sci. 2021, 22, 6603. [Google Scholar] [CrossRef] [PubMed]
- Perluigi, M.; Coccia, R.; Butterfield, D.A. 4-Hydroxy-2-Nonenal, a Reactive Product of Lipid Peroxidation, and Neurodegenerative Diseases: A Toxic Combination Illuminated by Redox Proteomics Studies. Antioxid. Redox Signal. 2012, 17, 1590–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.; Shal, B.; Khan, A.; Ullah, R.; Baig, M.; Haq, I.U.; Seo, E.; Khan, S. Suppression of TRPV1/TRPM8/P2Y Nociceptors by Withametelin via Downregulating MAPK Signaling in Mouse Model of Vincristine-Induced Neuropathic Pain. Int. J. Mol. Sci. 2021, 22, 6084. [Google Scholar] [CrossRef] [PubMed]
- Herken, H.; Uz, E.; Özyurt, H.; Söğüt, S.; Virit, O.; Akyol, O. Evidence that the activities of erythrocyte free radical scavenging enzymes and the products of lipid peroxidation are increased in different forms of schizophrenia. Mol. Psychiatry 2001, 6, 66–73. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Tan, Y.L.; Cao, L.Y.; Wu, G.Y.; Xu, Q.; Shen, Y.; Zhou, D.F. Antioxidant enzymes and lipid peroxidation in different forms of schizophrenia treated with typical and atypical antipsychotics. Schizophr. Res. 2006, 81, 291–300. [Google Scholar] [CrossRef]
- Möller, M.; Du Preez, J.L.; Viljoen, F.P.; Berk, M.; Harvey, B.H. N-acetyl cysteine reverses social isolation rearing induced changes in cortico-striatal monoamines in rats. Metab. Brain Dis. 2013, 28, 687–696. [Google Scholar] [CrossRef]
- Meza, C.A.; La Favor, J.D.; Kim, D.-H.; Hickner, R.C. Endothelial Dysfunction: Is There a Hyperglycemia-Induced Imbalance of NOX and NOS? Int. J. Mol. Sci. 2019, 20, 3775. [Google Scholar] [CrossRef] [Green Version]
- Hlavacova, N.; Jezova, D. Chronic treatment with the mineralocorticoid hormone aldosterone results in increased anxiety-like behavior. Horm. Behav. 2008, 54, 90–97. [Google Scholar] [CrossRef]
- Geyer, M.A.; Swerdlow, N.R. Measurement of Startle Response, Prepulse Inhibition, and Habituation. Curr. Protoc. Neurosci. 2001, 8, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Bredt, D.S.; Snyder, S.H. Isolation of nitric oxide synthetase, a calmodulin-requiring enzyme. Proc. Natl. Acad. Sci. USA 1990, 87, 682–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pecháňová, O.; Bernátová, I.; Pelouch, V.; Šimko, F. Protein Remodelling of the Heart in NO-deficient Hypertension: The Effect of Captopril. J. Mol. Cell. Cardiol. 1997, 29, 3365–3374. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vrankova, S.; Galandakova, Z.; Benko, J.; Cebova, M.; Riecansky, I.; Pechanova, O. Duration of Social Isolation Affects Production of Nitric Oxide in the Rat Brain. Int. J. Mol. Sci. 2021, 22, 10340. https://doi.org/10.3390/ijms221910340
Vrankova S, Galandakova Z, Benko J, Cebova M, Riecansky I, Pechanova O. Duration of Social Isolation Affects Production of Nitric Oxide in the Rat Brain. International Journal of Molecular Sciences. 2021; 22(19):10340. https://doi.org/10.3390/ijms221910340
Chicago/Turabian StyleVrankova, Stanislava, Zuzana Galandakova, Jakub Benko, Martina Cebova, Igor Riecansky, and Olga Pechanova. 2021. "Duration of Social Isolation Affects Production of Nitric Oxide in the Rat Brain" International Journal of Molecular Sciences 22, no. 19: 10340. https://doi.org/10.3390/ijms221910340
APA StyleVrankova, S., Galandakova, Z., Benko, J., Cebova, M., Riecansky, I., & Pechanova, O. (2021). Duration of Social Isolation Affects Production of Nitric Oxide in the Rat Brain. International Journal of Molecular Sciences, 22(19), 10340. https://doi.org/10.3390/ijms221910340