
The E3 Ubiquitin Ligase NEDD4-1 Mediates Temozolomide-Resistant Glioblastoma through PTEN Attenuation and Redox Imbalance in Nrf2–HO-1 Axis

 , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
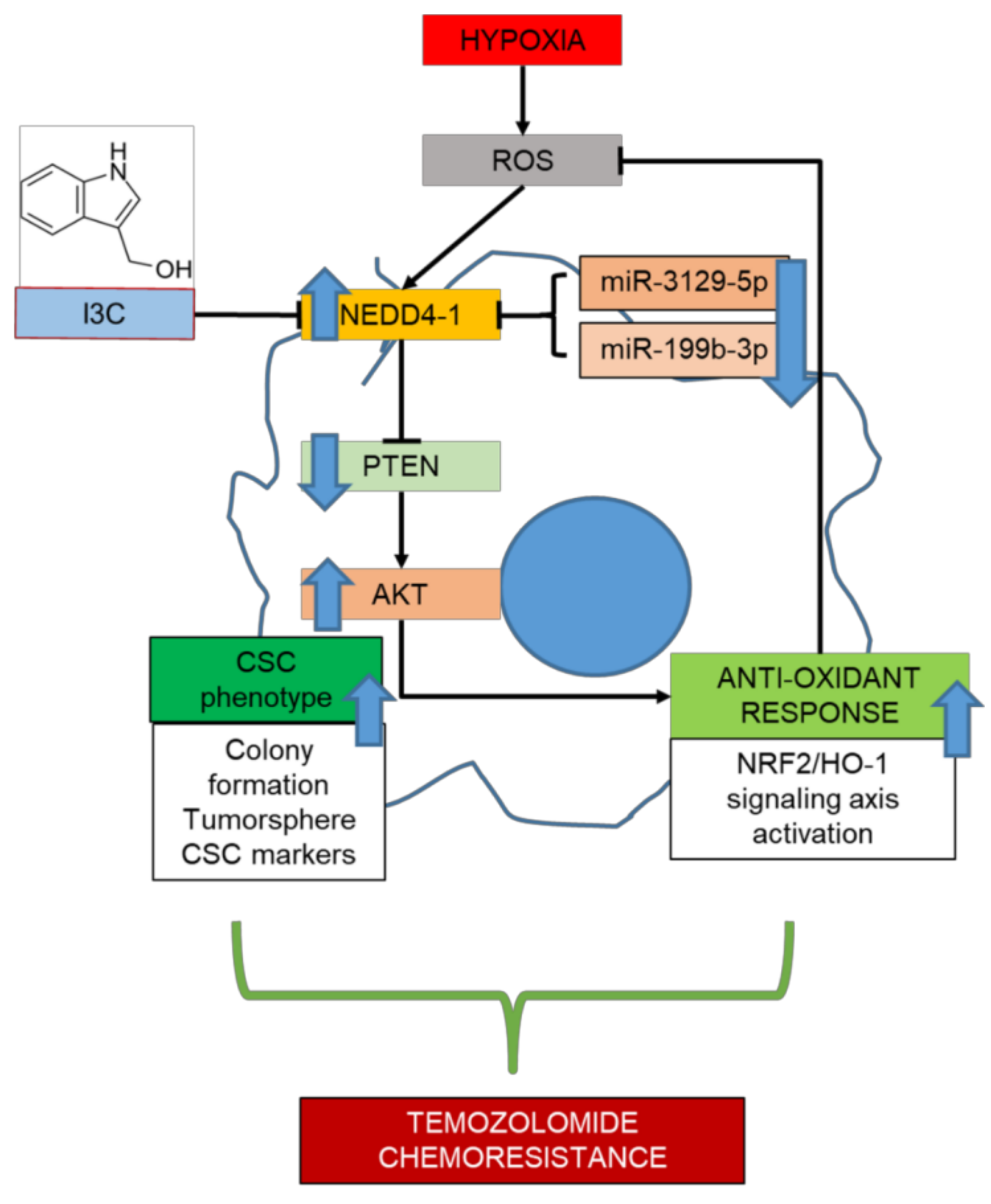
2. Results
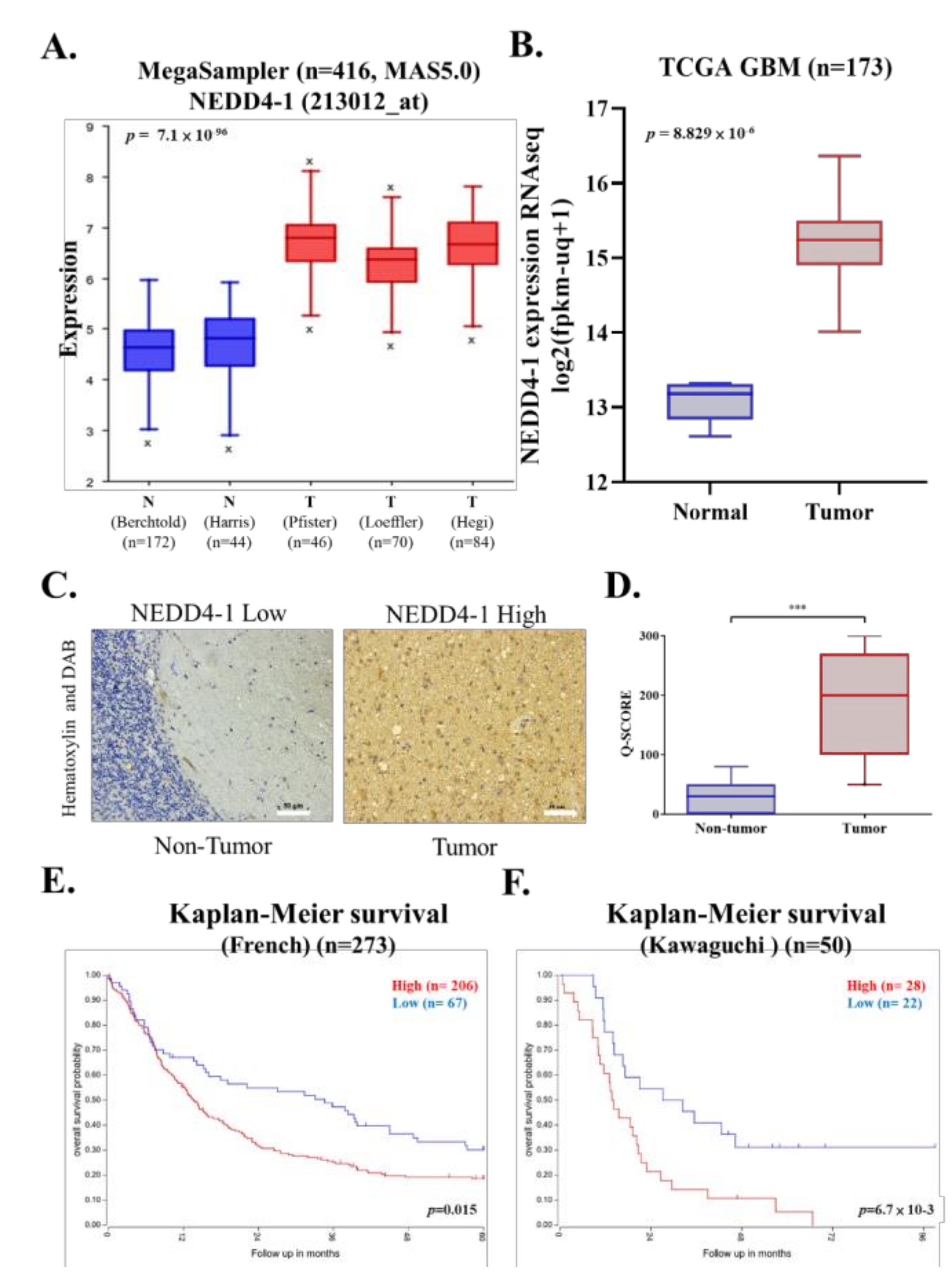
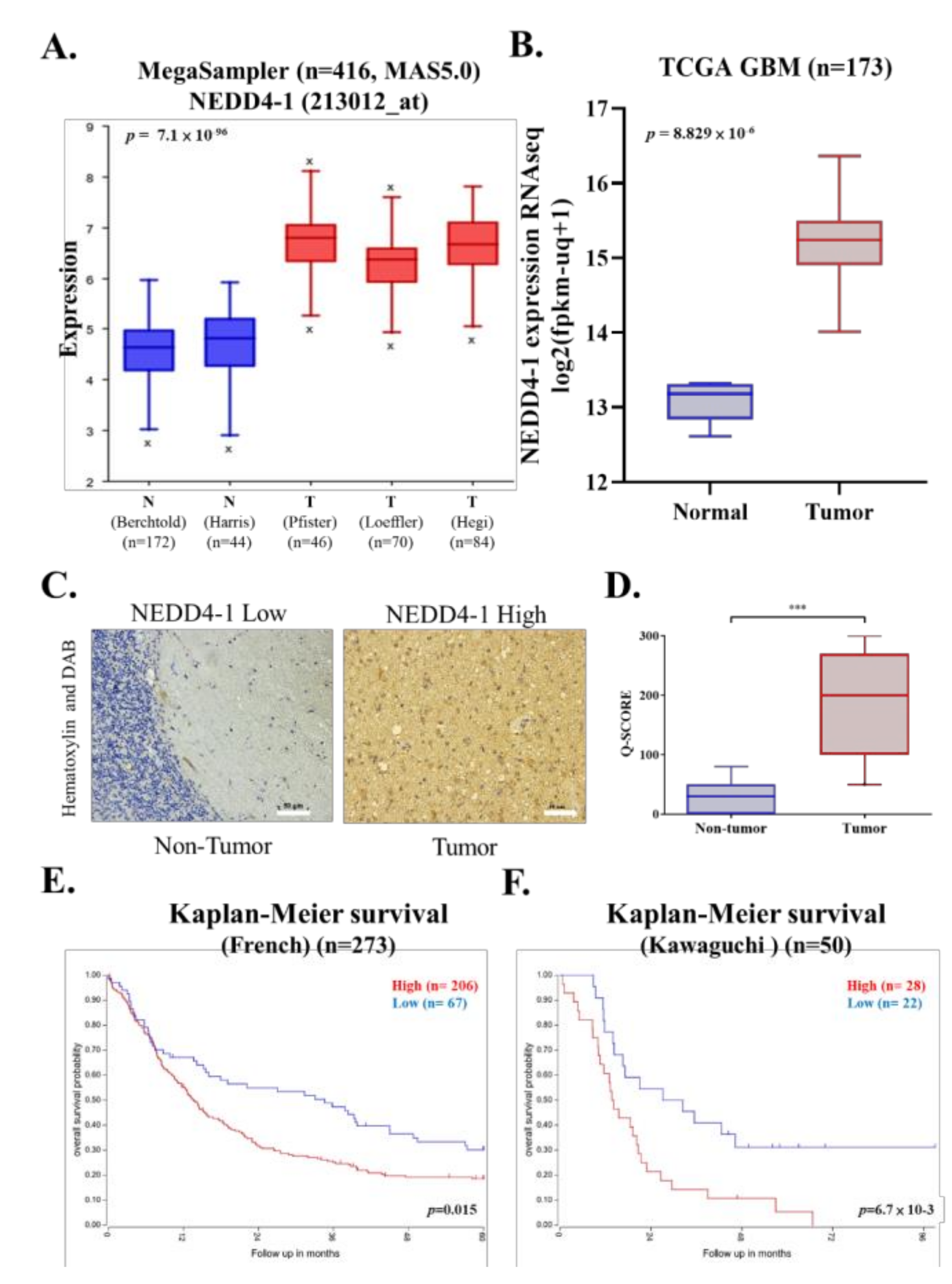
2.1. NEDD4-1 Was Overexpressed in GBM Tissue Samples
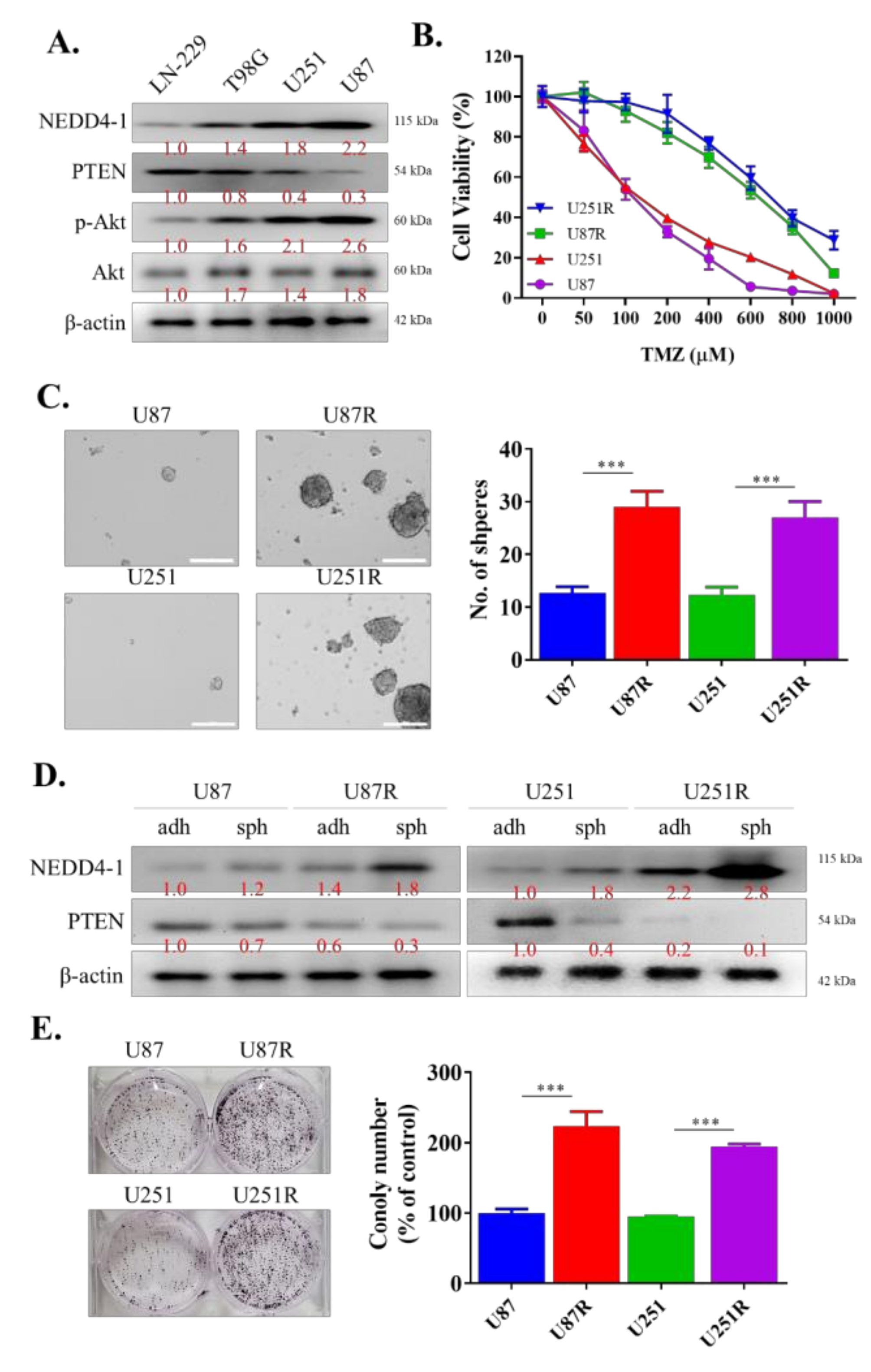
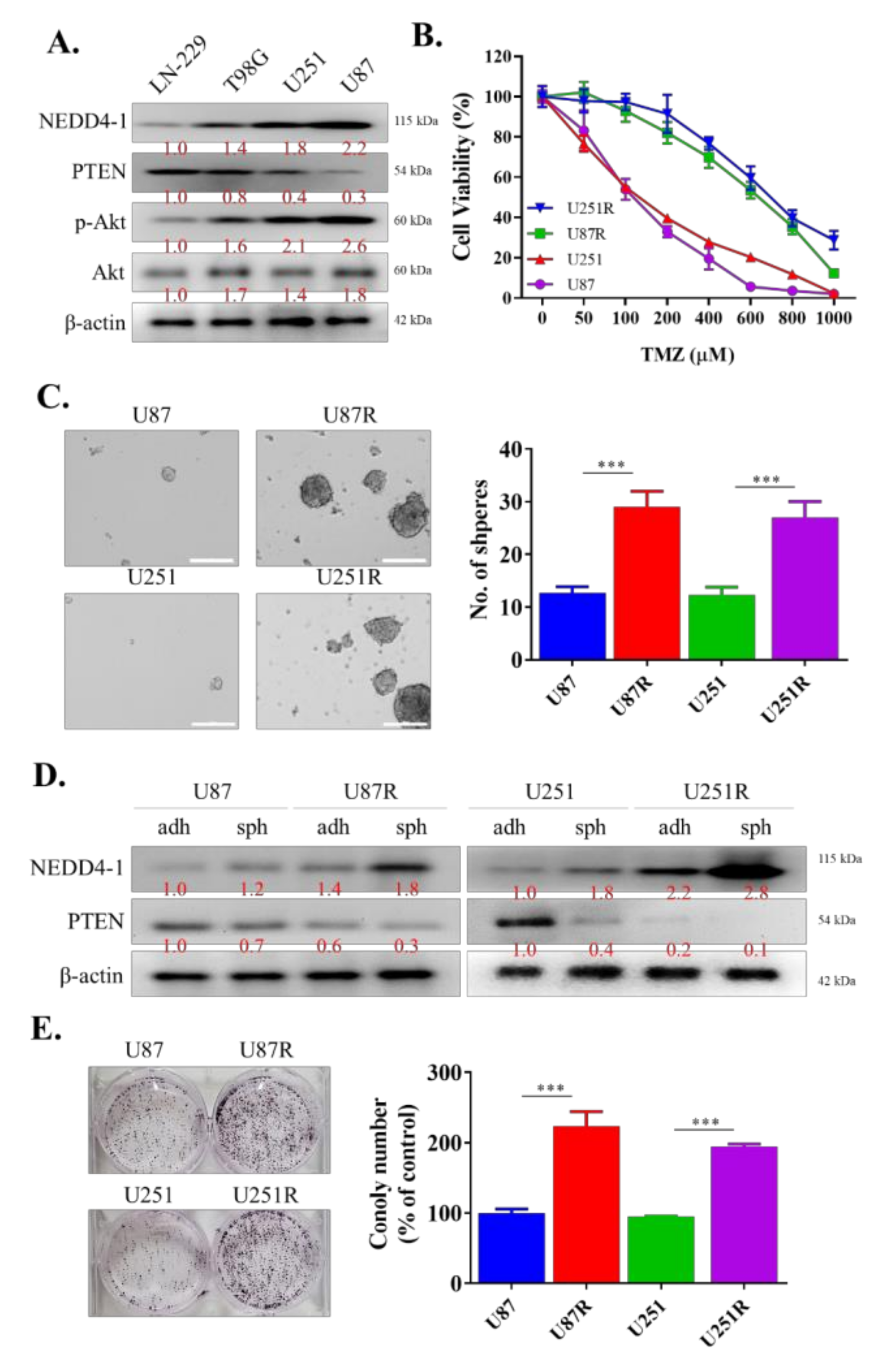
2.2. TMZ-Resistant GBM Cell Lines Had an Increased Endogenous Expression of NEDD4-1
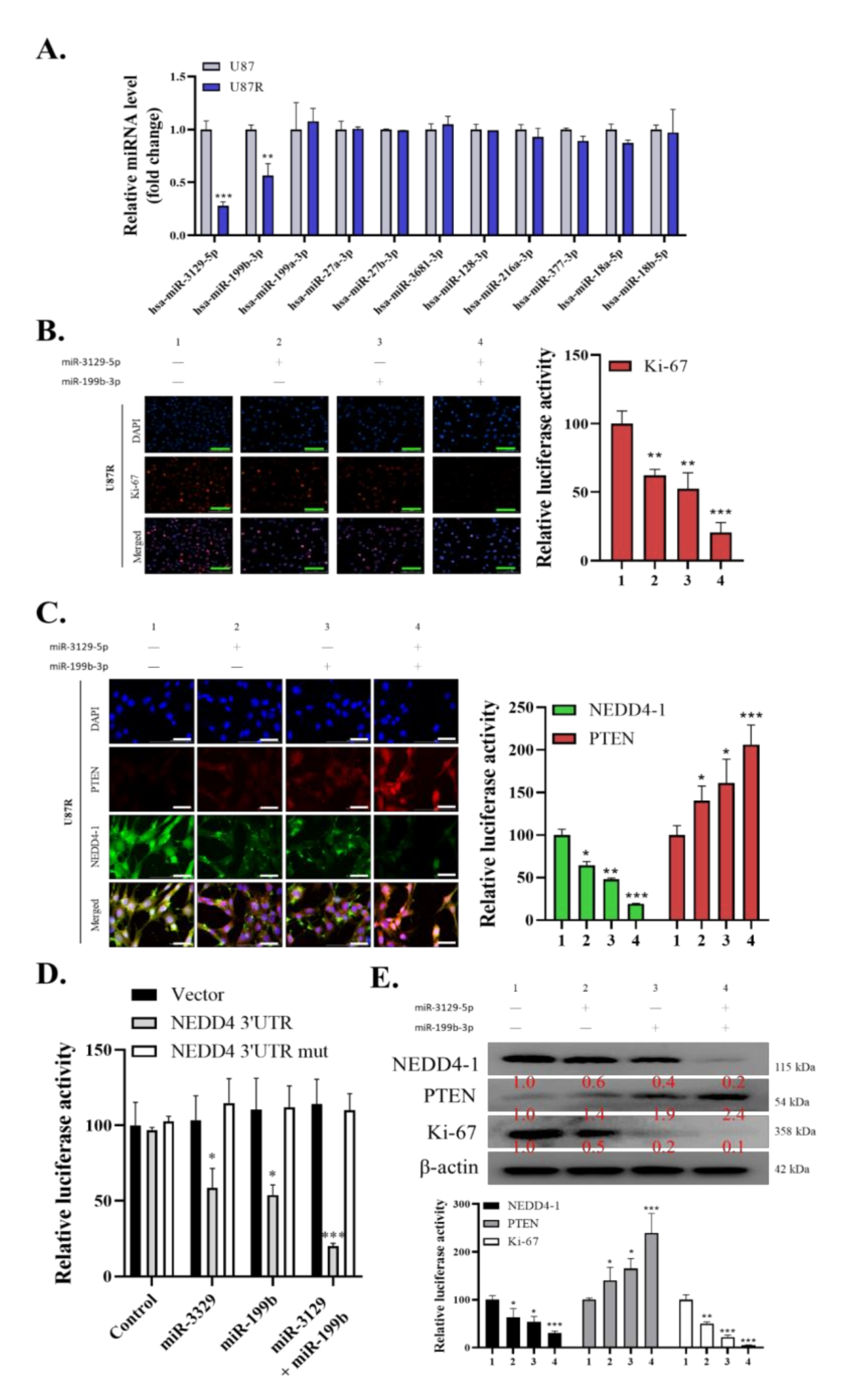
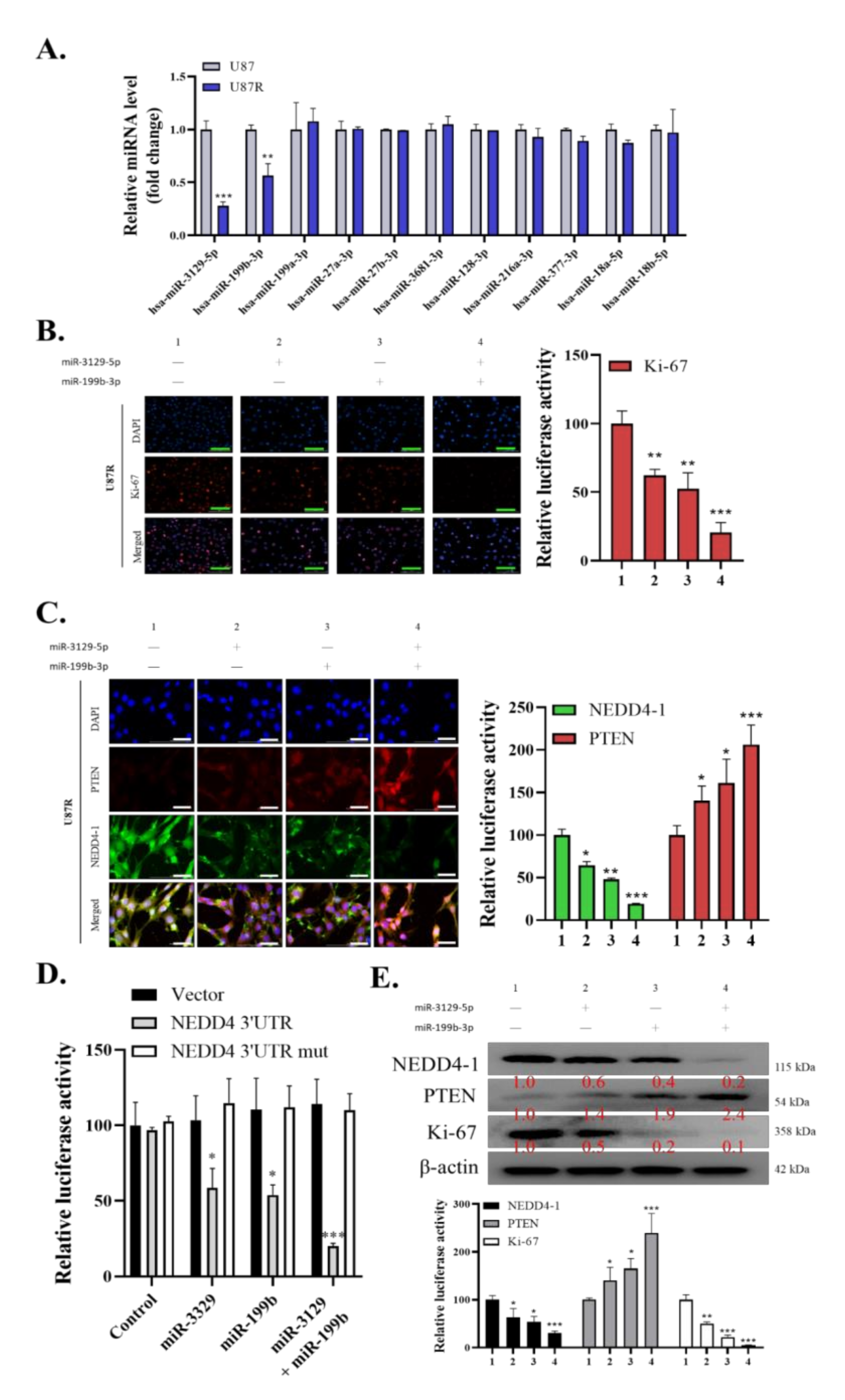
2.3. Dysregulation of miR-3129-5p and miR-199b-3p Consequently Leads to an Induced Expression of NEDD4-1 in TMZ-Resistant GBM Cells
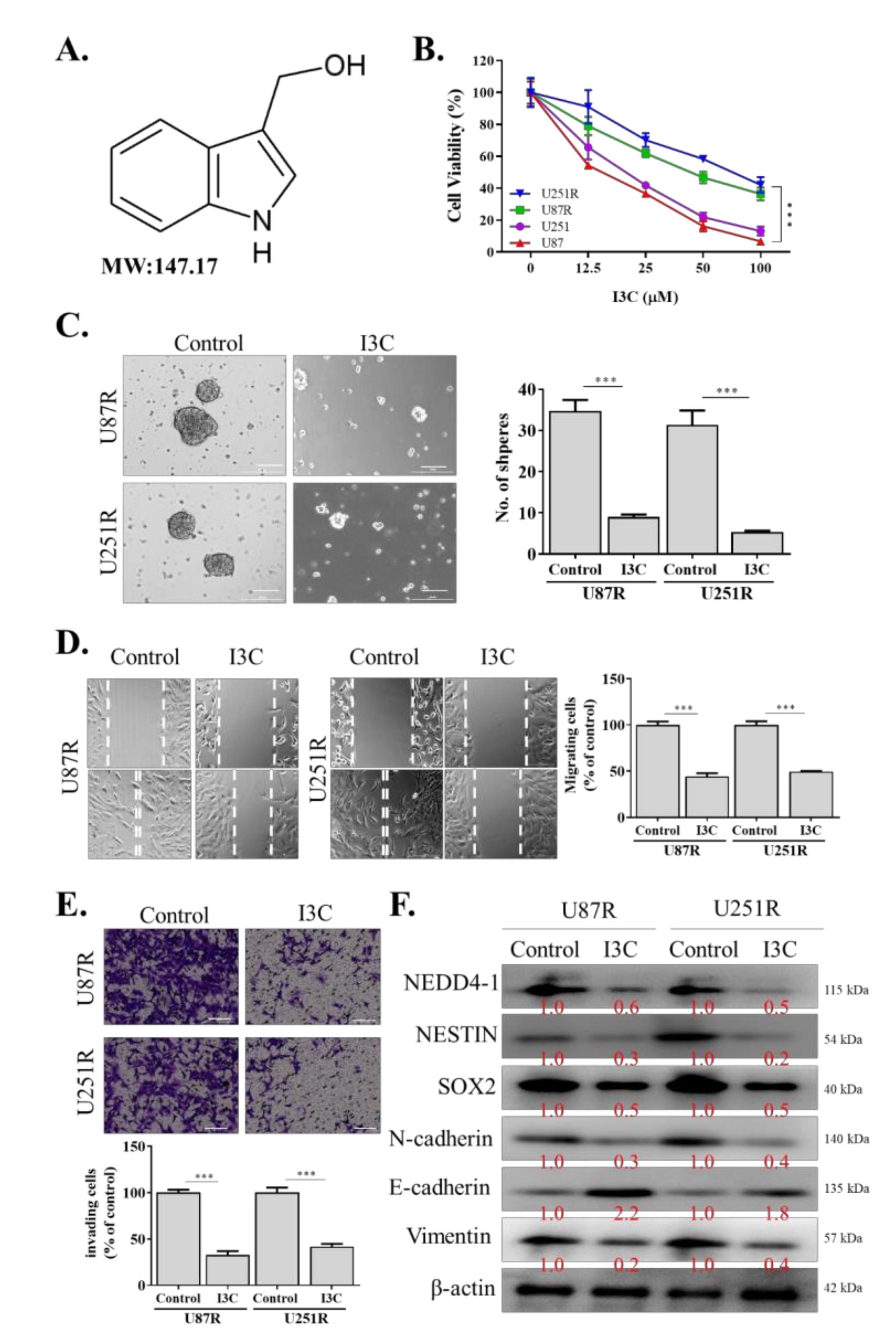
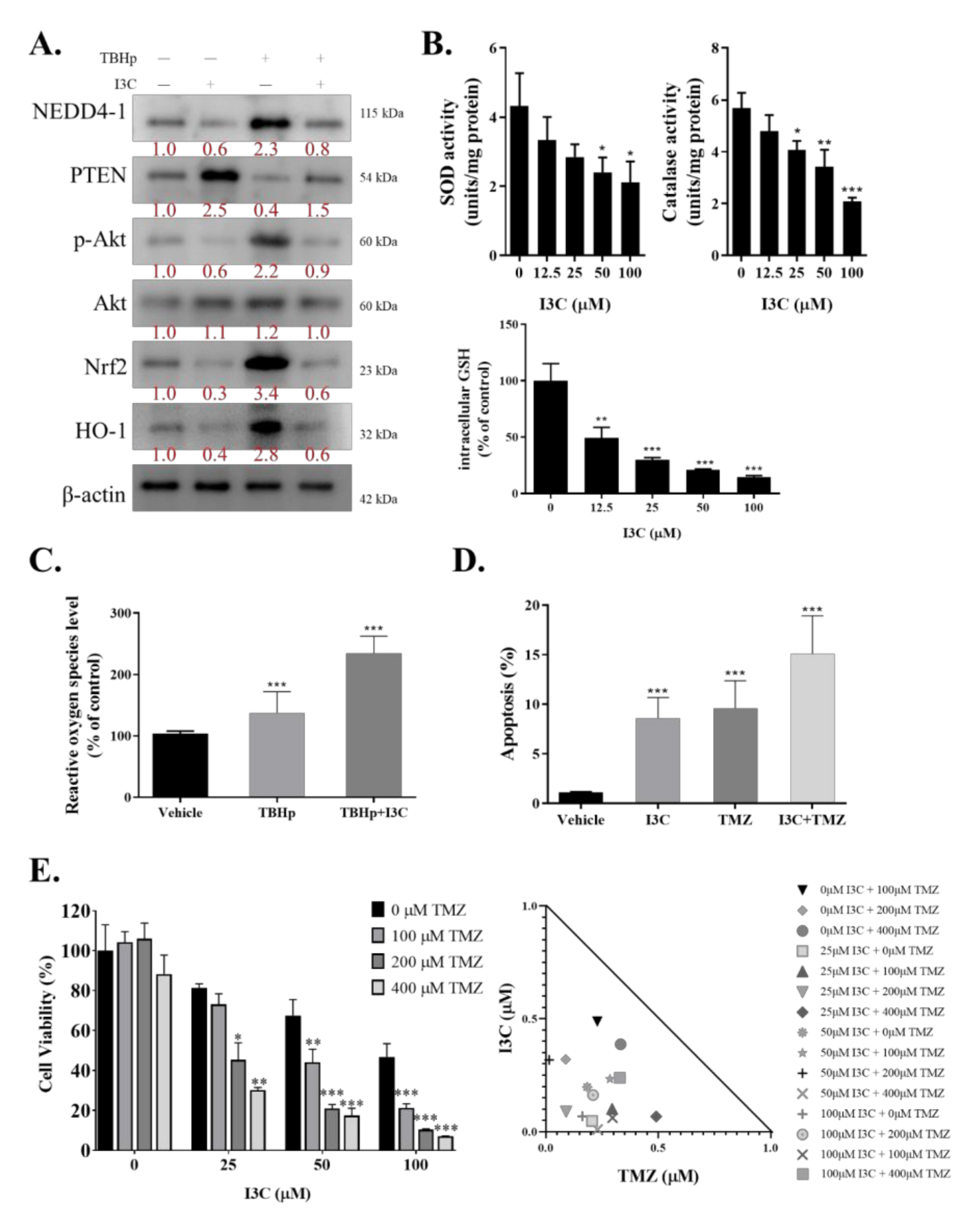
2.4. I3C Targets NEDD4-1 and Modulates the Oncogenic and Metastatic Phenotypes of TMZ-Resistant GBM Cells
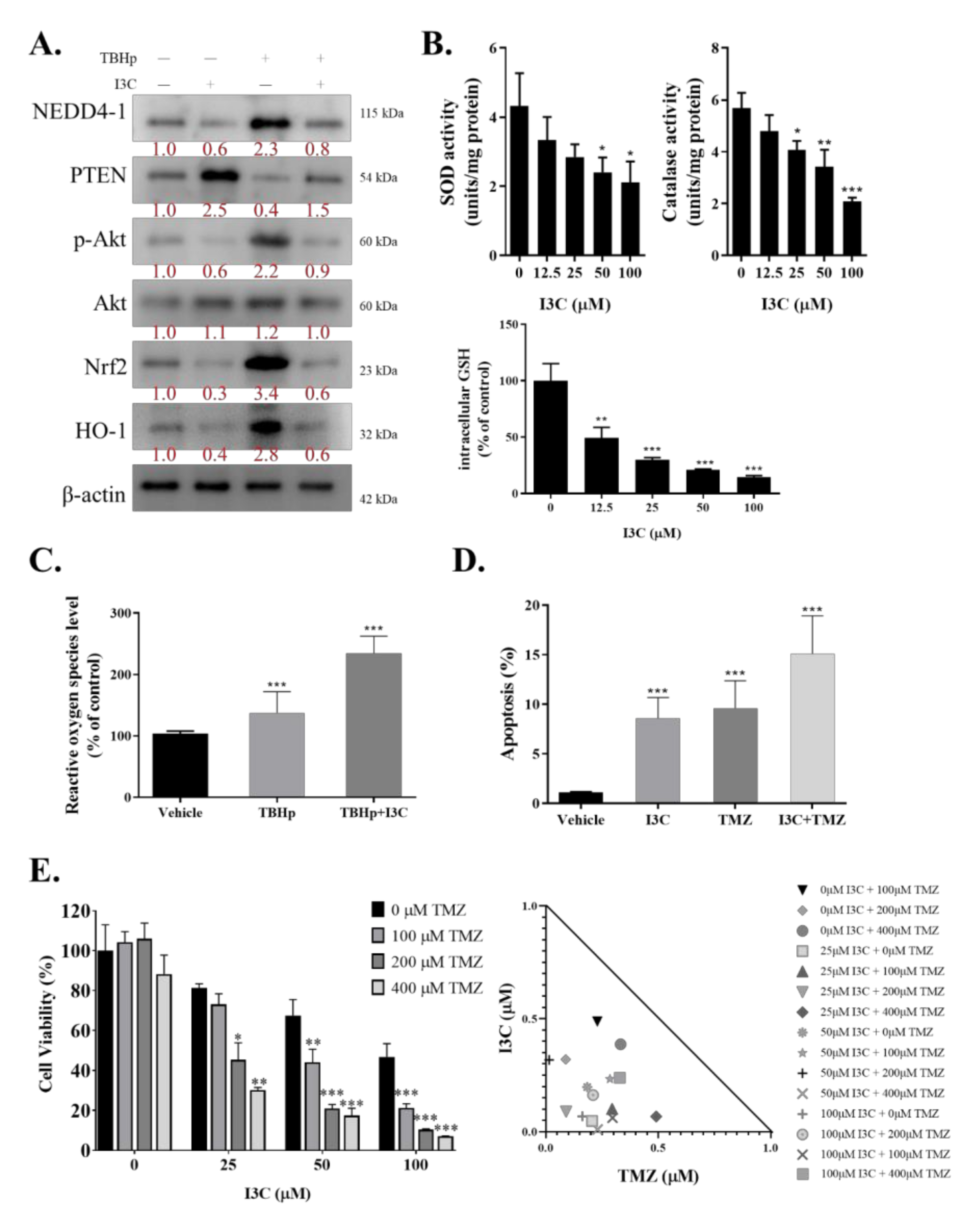
2.5. I3C Increased Susceptibility of TMZ-Resistant Cells to TMZ by Inhibiting NEDD4-1-Induced PTEN Ubiquitination and Subsequently Inhibiting NRF2/HO-1 Antioxidant Signaling Response
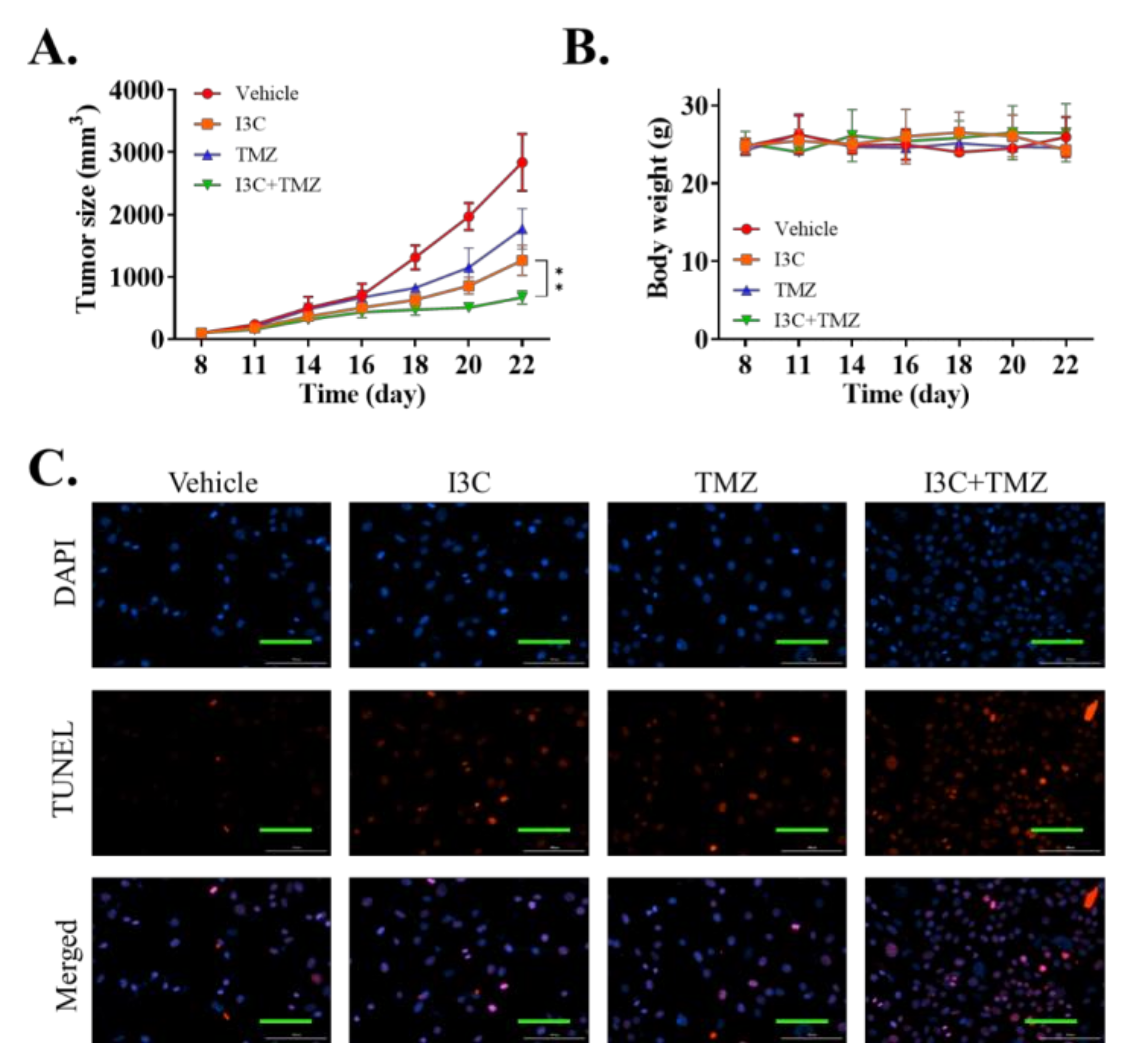
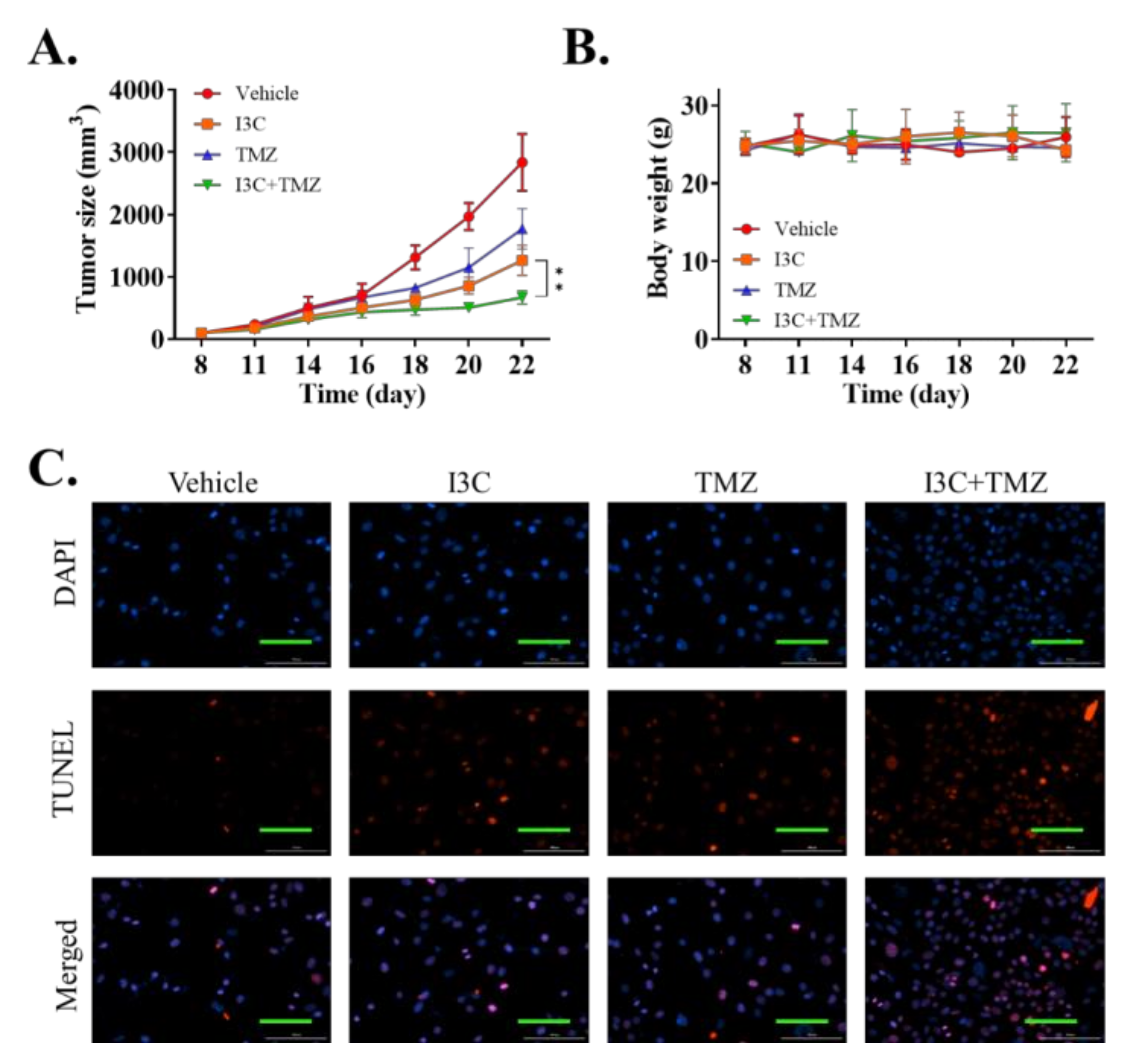
2.6. I3C Enhanced the Chemosensitivity of Xenografted TMZ-Resistant U87 Cell Line to TMZ
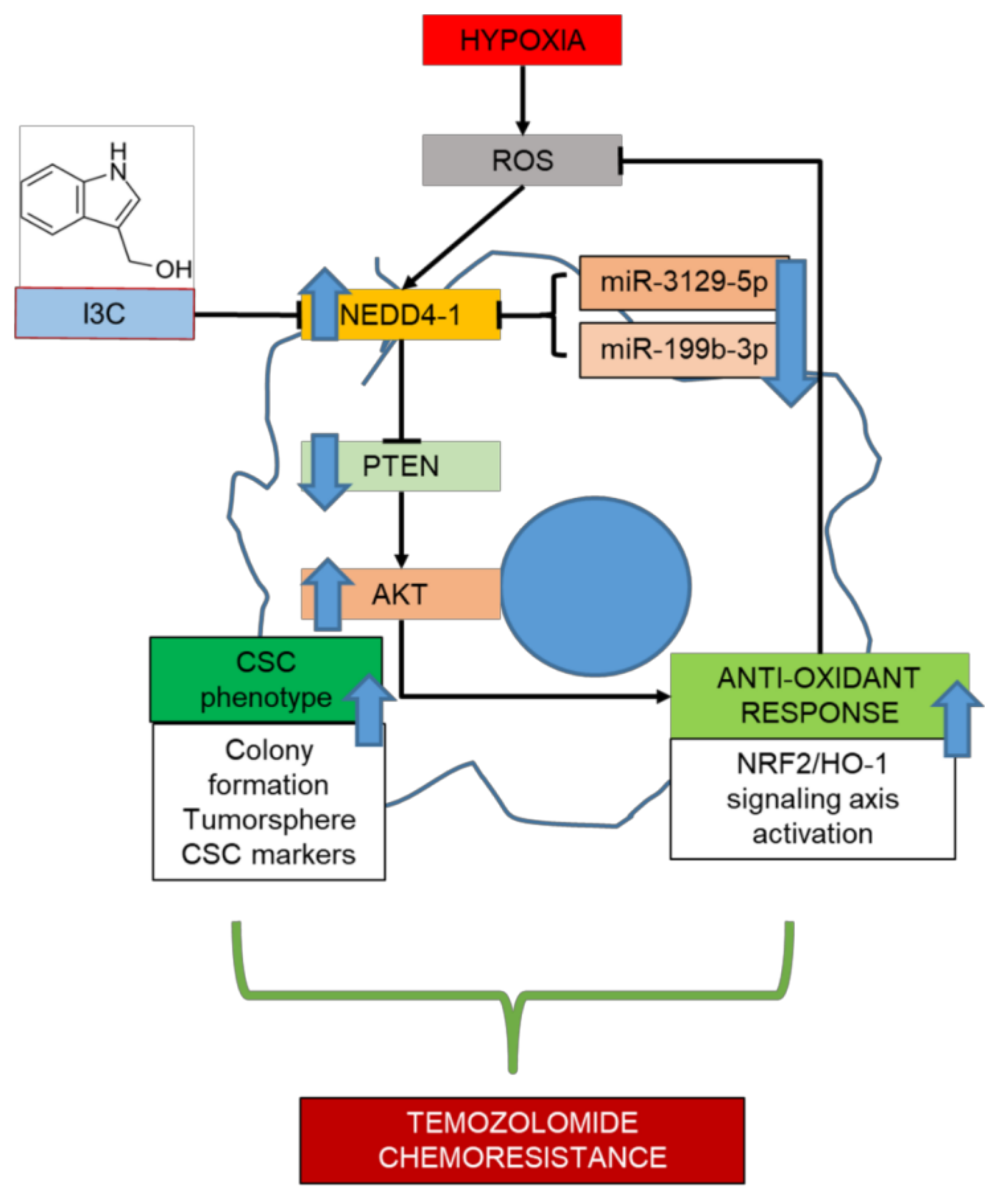
3. Discussion
4. Materials and Methods
4.1. Clinical Samples
4.2. Cell Culture and Generation of Temozolomide-Resistant Cell Line
4.3. Western Blotting
4.4. Transient Oligonucleotide Transfection
4.5. RNA Isolation and Reverse Transcription Quantitative Polymerase Chain Reaction
4.6. Cell Sensitivity Assay/Clonogenic Assay
4.7. Sulforhodamine B (SRB) Assay
4.8. Immunofluorescence Assay
4.9. Measurement of ROS Production
4.10. Tumor Xenograft Study
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Vargas Lopez, A.J. Glioblastoma in adults: A Society for Neuro-Oncology (SNO) and European Society of Neuro-Oncology (EANO) consensus review on current management and future directions. Neuro Oncol. 2021, 23, 502–503. [Google Scholar] [CrossRef] [PubMed]
- Zottel, A.; Jovcevska, I.; Samec, N.; Komel, R. Cytoskeletal proteins as glioblastoma biomarkers and targets for therapy: A systematic review. Crit. Rev. Oncol. Hematol. 2021, 160, 103283. [Google Scholar] [CrossRef]
- Tome, M.E.; Frye, J.B.; Coyle, D.L.; Jacobson, E.L.; Samulitis, B.K.; Dvorak, K.; Dorr, R.T.; Briehl, M.M. Lymphoma cells with increased anti-oxidant defenses acquire chemoresistance. Exp. Ther. Med. 2012, 3, 845–852. [Google Scholar] [CrossRef] [Green Version]
- Olivier, C.; Oliver, L.; Lalier, L.; Vallette, F.M. Drug Resistance in Glioblastoma: The Two Faces of Oxidative Stress. Front. Mol. Biosci. 2020, 7, 620677. [Google Scholar] [CrossRef]
- Jelic, M.D.; Mandic, A.D.; Maricic, S.M.; Srdjenovic, B.U. Oxidative stress and its role in cancer. J. Cancer Res. Ther. 2021, 17, 22–28. [Google Scholar] [CrossRef]
- Salazar-Ramiro, A.; Ramirez-Ortega, D.; Perez de la Cruz, V.; Hernandez-Pedro, N.Y.; Gonzalez-Esquivel, D.F.; Sotelo, J.; Pineda, B. Role of Redox Status in Development of Glioblastoma. Front. Immunol. 2016, 7, 156. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Shin, H.; Lee, J.K.; Liu, Z.; Rabadan, R.; Lee, J.; Shin, J.; Lee, C.; Yang, H.; Kim, D.; et al. Secretome analysis of patient-derived GBM tumor spheres identifies midkine as a potent therapeutic target. Exp. Mol. Med. 2019, 51, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sato, A.; Okada, M.; Shibuya, K.; Watanabe, E.; Seino, S.; Narita, Y.; Shibui, S.; Kayama, T.; Kitanaka, C. Pivotal role for ROS activation of p38 MAPK in the control of differentiation and tumor-initiating capacity of glioma-initiating cells. Stem. Cell Res. 2014, 12, 119–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gursel, D.B.; Shin, B.J.; Burkhardt, J.K.; Kesavabhotla, K.; Schlaff, C.D.; Boockvar, J.A. Glioblastoma stem-like cells-biology and therapeutic implications. Cancers 2011, 3, 2655–2666. [Google Scholar] [CrossRef] [Green Version]
- Dey, M.; Ulasov, I.V.; Tyler, M.A.; Sonabend, A.M.; Lesniak, M.S. Cancer stem cells: The final frontier for glioma virotherapy. Stem Cell Rev. Rep. 2011, 7, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawamura, Y.; Takouda, J.; Yoshimoto, K.; Nakashima, K. New aspects of glioblastoma multiforme revealed by similarities between neural and glioblastoma stem cells. Cell Biol. Toxicol. 2018, 34, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Afify, S.M.; Oo, A.K.K.; Hassan, G.; Seno, A.; Seno, M. How can we turn the PI3K/AKT/mTOR pathway down? Insights into inhibition and treatment of cancer. Expert Rev. Anticancer Ther. 2021, 21, 605–619. [Google Scholar] [CrossRef]
- Li, J.; Wang, J.; Xie, D.; Pei, Q.; Wan, X.; Xing, H.R.; Ye, T. Characteristics of the PI3K/AKT and MAPK/ERK pathways involved in the maintenance of self-renewal in lung cancer stem-like cells. Int. J. Biol. Sci. 2021, 17, 1191–1202. [Google Scholar] [CrossRef]
- Sanches, J.G.P.; Song, B.; Zhang, Q.; Cui, X.; Yabasin, I.B.; Ntim, M.; Li, X.; He, J.; Zhang, Y.; Mao, J.; et al. The Role of KDM2B and EZH2 in Regulating the Stemness in Colorectal Cancer Through the PI3K/AKT Pathway. Front. Oncol. 2021, 11, 637298. [Google Scholar] [CrossRef]
- He, H.; Huang, C.; Chen, Z.; Huang, H.; Wang, X.; Chen, J. An outlined review for the role of Nedd4-1 and Nedd4-2 in lung disorders. Biomed. Pharmacother. 2020, 125, 109983. [Google Scholar] [CrossRef] [PubMed]
- Sang, P.F.; Wang, H.; Wang, M.; Hu, C.; Zhang, J.S.; Li, X.J.; Zhu, F. NEDD4-1 and PTEN expression in keloid scarring. Genet. Mol. Res. 2015, 14, 13467–13475. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, X.; Zhang, Y. Correlation of NEDD4-1 and PTEN expression with the invasive capacity of pituitary adenomas. Mol. Clin. Oncol. 2017, 6, 96–100. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Matesic, L.E. The Nedd4-like family of E3 ubiquitin ligases and cancer. Cancer Metastasis Rev. 2007, 26, 587–604. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Seth, A.K.; Aplin, A.E. Genetic and expression aberrations of E3 ubiquitin ligases in human breast cancer. Mol. Cancer Res. 2006, 4, 695–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, Y.S.; Qian, Y.; Chen, X. Pirh2 RING-finger E3 ubiquitin ligase: Its role in tumorigenesis and cancer therapy. FEBS Lett. 2012, 586, 1397–1402. [Google Scholar] [CrossRef] [Green Version]
- Penas, C.; Ramachandran, V.; Ayad, N.G. The APC/C Ubiquitin Ligase: From Cell Biology to Tumorigenesis. Front. Oncol. 2011, 1, 60. [Google Scholar] [CrossRef] [Green Version]
- Senft, D.; Qi, J.; Ronai, Z.A. Ubiquitin ligases in oncogenic transformation and cancer therapy. Nat. Rev. Cancer 2018, 18, 69–88. [Google Scholar] [CrossRef] [PubMed]
- Samarzija, I. Post-Translational Modifications That Drive Prostate Cancer Progression. Biomolecules 2021, 11, 247. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Chen, J.; Cao, W.; Yang, L.; Chen, Q.; He, J.; Yi, Q.; Huang, H.; Zhang, E.; Cai, Z. The many substrates and functions of NEDD4-1. Cell Death Dis. 2019, 10, 904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.W.; Hu, X.; Ye, M.; Lin, M.; Chu, M.; Shen, X. NEDD4 E3 ligase: Functions and mechanism in human cancer. Semin. Cancer Biol. 2020, 67, 92–101. [Google Scholar] [CrossRef]
- Quirit, J.G.; Lavrenov, S.N.; Poindexter, K.; Xu, J.; Kyauk, C.; Durkin, K.A.; Aronchik, I.; Tomasiak, T.; Solomatin, Y.A.; Preobrazhenskaya, M.N.; et al. Indole-3-carbinol (I3C) analogues are potent small molecule inhibitors of NEDD4-1 ubiquitin ligase activity that disrupt proliferation of human melanoma cells. Biochem. Pharmacol. 2017, 127, 13–27. [Google Scholar] [CrossRef]
- Mesrian Tanha, H.; Mojtabavi Naeini, M.; Rahgozar, S.; Moafi, A.; Honardoost, M.A. Integrative computational in-depth analysis of dysregulated miRNA-mRNA interactions in drug-resistant pediatric acute lymphoblastic leukemia cells: An attempt to obtain new potential gene-miRNA pathways involved in response to treatment. Tumour Biol. 2016, 37, 7861–7872. [Google Scholar] [CrossRef]
- Pogribny, I.P.; Filkowski, J.N.; Tryndyak, V.P.; Golubov, A.; Shpyleva, S.I.; Kovalchuk, O. Alterations of microRNAs and their targets are associated with acquired resistance of MCF-7 breast cancer cells to cisplatin. Int. J. Cancer 2010, 127, 1785–1794. [Google Scholar] [CrossRef] [PubMed]
- Pandey, M.; Mukhopadhyay, A.; Sharawat, S.K.; Kumar, S. Role of microRNAs in regulating cell proliferation, metastasis and chemoresistance and their applications as cancer biomarkers in small cell lung cancer. Biochim. Biophys. Acta Rev. Cancer 2021, 1876, 188552. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Li, J.; Song, J.; Guo, F. miR-186 reverses cisplatin resistance and inhibits the formation of the glioblastoma-initiating cell phenotype by degrading Yin Yang 1 in glioblastoma. Int. J. Mol. Med. 2019, 43, 517–524. [Google Scholar] [CrossRef] [Green Version]
- Sana, J.; Busek, P.; Fadrus, P.; Besse, A.; Radova, L.; Vecera, M.; Reguli, S.; Stollinova Sromova, L.; Hilser, M.; Lipina, R.; et al. Identification of microRNAs differentially expressed in glioblastoma stem-like cells and their association with patient survival. Sci. Rep. 2018, 8, 2836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aronchik, I.; Kundu, A.; Quirit, J.G.; Firestone, G.L. The antiproliferative response of indole-3-carbinol in human melanoma cells is triggered by an interaction with NEDD4-1 and disruption of wild-type PTEN degradation. Mol. Cancer Res. 2014, 12, 1621–1634. [Google Scholar] [CrossRef] [Green Version]
- Oliva, C.R.; Moellering, D.R.; Gillespie, G.Y.; Griguer, C.E. Acquisition of chemoresistance in gliomas is associated with increased mitochondrial coupling and decreased ROS production. PLoS ONE 2011, 6, e24665. [Google Scholar] [CrossRef] [Green Version]
- Falone, S.; Lisanti, M.P.; Domenicotti, C. Oxidative Stress and Reprogramming of Mitochondrial Function and Dynamics as Targets to Modulate Cancer Cell Behavior and Chemoresistance. Oxid. Med. Cell. Longev. 2019, 2019, 4647807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koster, J. R2: Genomics Analysis and Visualization Platform. Available online: http://r2.amc.nl (accessed on 2 July 2020).
- Akiyama, Y.; Ashizawa, T.; Komiyama, M.; Miyata, H.; Oshita, C.; Omiya, M.; Iizuka, A.; Kume, A.; Sugino, T.; Hayashi, N.; et al. YKL-40 downregulation is a key factor to overcome temozolomide resistance in a glioblastoma cell line. Oncol. Rep. 2014, 32, 159–166. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Position in the UTR | Seed 1234match | Context++ Score | Context++ Score Percentile | Weighted Context++ Score | Conserved Branch Length | Pct |
|---|---|---|---|---|---|---|---|
| hsa-miR-3129-5p | 366–373 | 8mer | −0.31 | 97 | −0.31 | 7.3 | 0.87 |
| hsa-miR-199b-3p | 366–373 | 8mer | −0.34 | 96 | −0.34 | 7.3 | 0.87 |
| hsa-miR-199a-3p | 366–373 | 8mer | −0.34 | 96 | −0.34 | 7.3 | 0.87 |
| hsa-miR-27a-3p | 381–388 | 8mer | −0.43 | 99 | −0.43 | 6.942 | 0.96 |
| hsa-miR-27b-3p | 381–388 | 8mer | −0.43 | 99 | −0.43 | 6.942 | 0.96 |
| hsa-miR-3681-3p | 381–387 | 7mer-1A | −0.22 | 93 | −0.22 | 6.942 | 0.77 |
| hsa-miR-128-3p | 381–387 | 7mer-1A | −0.17 | 90 | −0.17 | 6.942 | 0.77 |
| hsa-miR-216a-3p | 381–387 | 7mer-1A | −0.15 | 88 | −0.15 | 6.942 | 0.77 |
| hsa-miR-377-3p | 408–415 | 8mer | −0.11 | 72 | −0.11 | 2.883 | N/A |
| hsa-miR-18a-5p | 538–545 | 8mer | −0.35 | 97 | −0.33 | 5.828 | 0.83 |
| hsa-miR-18b-5p | 538–545 | 8mer | −0.35 | 97 | −0.33 | 5.828 | 0.83 |
| Total no. Patients of Glioblastoma (n = 58) | No. (%) |
|---|---|
| Age | |
| <65 | 40 (68.4%) |
| ≥65 | 18 (31.6%) |
| Gender | |
| male | 34 (58.6%) |
| female | 24 (41.4%) |
| IDH1 status | |
| wildtype | 56 (96.6%) |
| mutation | 2 (3.4%) |
| Treatment modalities | |
| Chemotherapy | 24 (41.4%) |
| Radiotherapy | 34 (58.6%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chuang, H.-Y.; Hsu, L.-Y.; Pan, C.-M.; Pikatan, N.W.; Yadav, V.K.; Fong, I.-H.; Chen, C.-H.; Yeh, C.-T.; Chiu, S.-C. The E3 Ubiquitin Ligase NEDD4-1 Mediates Temozolomide-Resistant Glioblastoma through PTEN Attenuation and Redox Imbalance in Nrf2–HO-1 Axis. Int. J. Mol. Sci. 2021, 22, 10247. https://doi.org/10.3390/ijms221910247
Chuang H-Y, Hsu L-Y, Pan C-M, Pikatan NW, Yadav VK, Fong I-H, Chen C-H, Yeh C-T, Chiu S-C. The E3 Ubiquitin Ligase NEDD4-1 Mediates Temozolomide-Resistant Glioblastoma through PTEN Attenuation and Redox Imbalance in Nrf2–HO-1 Axis. International Journal of Molecular Sciences. 2021; 22(19):10247. https://doi.org/10.3390/ijms221910247
Chicago/Turabian StyleChuang, Hao-Yu, Li-Yun Hsu, Chih-Ming Pan, Narpati Wesa Pikatan, Vijesh Kumar Yadav, Iat-Hang Fong, Chao-Hsuan Chen, Chi-Tai Yeh, and Shao-Chih Chiu. 2021. "The E3 Ubiquitin Ligase NEDD4-1 Mediates Temozolomide-Resistant Glioblastoma through PTEN Attenuation and Redox Imbalance in Nrf2–HO-1 Axis" International Journal of Molecular Sciences 22, no. 19: 10247. https://doi.org/10.3390/ijms221910247
APA StyleChuang, H.-Y., Hsu, L.-Y., Pan, C.-M., Pikatan, N. W., Yadav, V. K., Fong, I.-H., Chen, C.-H., Yeh, C.-T., & Chiu, S.-C. (2021). The E3 Ubiquitin Ligase NEDD4-1 Mediates Temozolomide-Resistant Glioblastoma through PTEN Attenuation and Redox Imbalance in Nrf2–HO-1 Axis. International Journal of Molecular Sciences, 22(19), 10247. https://doi.org/10.3390/ijms221910247