
Targeting Autophagy with Natural Products as a Potential Therapeutic Approach for Cancer

 ,
,  ,
, 
Abstract
1. Introduction
2. Molecular Mechanisms and Morphological Features of Autophagy
2.1. Autophagy Initiation
2.2. Cargo Nucleation, Elongation and Enclosure of IM
2.3. Transport of Autophagosomes
2.4. Autolysosome Formation, Vesicle Degradation and ALR Cycle
3. Natural Products as Inhibitors of Autophagy in Cancer
3.1. Class III-PI3K Complex Inhibitors
3.2. IM Elongation and Enclosure Stage Inhibitors
3.3. Docking and Fusion Stage Inhibitors
3.4. Late-Stage Disruptors
3.4.1. Acidification Stage Inhibitors
3.4.2. Vacuolar-Type H+-ATPase (V-ATPase) Inhibitors
3.4.3. Lysosomal Hydrolytic Enzyme Inhibitors
4. Natural Products as Inducers of Autophagy in Cancer
4.1. Initiation Stage Activators: mTOR Inhibitors
4.2. Polyphenolic Compounds
4.2.1. Quercetin
4.2.2. Magnolol
4.2.3. Kaempferol
4.2.4. Apigenin
4.2.5. Coffee and Tea: (−)-epigallocatechin-3-gallate (EGCG), Catechin and Epicatechin
4.2.6. Genistein
4.2.7. Curcumin Derivatives
4.2.8. Resveratrol
4.2.9. Propolis Extract: Chrysin
4.2.10. Fisetin
4.2.11. Rottlerin
4.3. Terpenoids
4.3.1. γ-Tocotrienol
4.3.2. Ursolic Acid
4.3.3. β-Elemene
4.3.4. (−)-Guaiol
4.3.5. Sesquiterpene Lactones: F1012-2
4.4. Saponin Compounds
4.4.1. Tubeimoside-1
4.4.2. Paris Polyphylla
4.4.3. Ophiopogonin B
4.4.4. Betulinic Acid
4.5. Alkaloids
4.5.1. Camptothecin
4.5.2. Berberine
4.5.3. Tetrandrine
4.5.4. Protopine
4.5.5. Neferine
4.5.6. Graveoline
4.6. Quinonoids
4.6.1. Thymoquinone
4.6.2. Celastrol
4.6.3. Pristimerin
4.6.4. Plumbagin
4.7. Omega-3 Polyunsaturated Fatty Acids (ω-3-PUFA)
4.8. Miscellaneous
4.8.1. Trichostatin A
4.8.2. 6-Shogaol
4.8.3. 7-Trehalose
5. Concluding Remarks and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| UPS | ubiquitin-proteasome system |
| ER | endoplasmic reticulum |
| ATG | Autophagy-related genes |
| FDA | Food and Drug Administration |
| IM | isolation membrane |
| ULK1 | Unc-51-like autophagy activating kinase 1 |
| PI3K | Phosphatidylinositide 3-kinase |
| mTORC1 | mechanistic target of rapamycin complex 1 |
| AMPK | AMP-activated protein kinase |
| MOM | Mitochondrial outer membrane |
| MOMP | MOM potential |
| ERES | ER exit sites |
| MAMs | mitochondria-associated ER membranes |
| PI3P | Phosphatidylinositol 3-phosphate |
| DFCP1 | double FYVE domain–containing protein 1 |
| Ubl | ubiquitin-like |
| LC3 | MT-associated protein 1 light chain 3 |
| LC3-PE | LC3-phosphatidylethanolamine |
| LIR | LC3 interacting regions |
| Ambra1 | Activating Molecule In Beclin1-Regulated Autophagy Protein 1 |
| p62/ SQSTM1 | Sequestosome-1 |
| MTs | Microtubules |
| ALR | autophagic lysosome regeneration |
| IP3R | inositol 1,4,5-trisphosphate (IP3) receptor |
| JNK | c-Jun N-terminal kinase |
| BH3 | Bcl-2 homology (BH) domain 3 |
| 3-MA | 3-methyladenine |
| MLCK | myosin light chain kinase |
| PLK1 | Polo-like kinase 1 |
| DNA-PK | DNA-dependent protein kinase |
| ATM | ataxia–telangiectasia mutated |
| NP | Nanoparticle |
| NCT | National Clinical Trial |
| PSM | Petrosaspongiolide M |
| EPI | Epirubicin |
| Bcl-2 | B-cell lymphoma 2 |
| TP | tea polyphenol |
| MTAs | MT-targeting agents |
| CA-4 | Combretastatin A-4 |
| CA-4P | CA-4 phosphate |
| NAC | N-acetyl cysteine |
| ROS | reactive oxygen species |
| rhArg | recombinant human arginase |
| 5-FU | 5-fluorouracil |
| VCP | Valosin Containing Protein or p97 |
| XN | xanthohumol |
| Sal A | salvianolic acid A |
| Sal B | salvianolic acid B |
| SCC | squamous cell carcinoma |
| CCC | circulating cancer cells |
| PTEN | phosphatase and tensin homolog deleted on chromosome 10 |
| EMT | epithelial-mesenchymal transition |
| AKT | protein kinase B (PKB) |
| HIF-1α | hypoxia-inducible factor 1α |
| VEGF | Vascular endothelial growth factor |
| HCC | hepatocellular carcinoma |
| CRC | colorectal cancer |
| Dox | Doxorubicin |
| Cdk4 | cyclin-dependent kinase 4 |
| Hsp-90 | heat shock protein-90 |
| PARP-1 | poly (ADP-ribose) polymerase 1 |
| ALL | acute lymphoblastic leukaemia |
| CQ | Chloroquine |
| Baf A1 | Bafilomycin A1 |
| OC | Oblongifolin C |
| GUTK | guttiferone K |
| v-ATPase | vacuolar-type ATPase |
| LMP | lysosomal membrane permeabilization |
| SAR | structure-activity relationship |
| CR | Caloric restriction |
| SIRT1 | Sirtuin 1 |
| BNIP3/BNIP3L | BCL2/adenovirus E1B 19 kDa protein-interacting protein 3/ligand |
| STAT3 | Signal transducer and activator of transcription 3 |
| RAGE | receptor for advanced glycation end products |
| MDR1 | multidrug resistance protein 1 |
| P-gp | P-glycoprotein |
| NLRP3 | NLR family pyrin domain containing 3 |
| EGCG | (−)-epigallocatechin-3-gallate |
| CGA | chlorogenic acid |
| EGC | (−)-epigallocatechin |
| ECG | (−)-epicatechin-3-gallate |
| NSCLC | non-small cell lung cancer |
| MAPK | mitogen-activated protein kinase |
| N-CoR | nuclear receptor co-repressor |
| HSC70 | Heat shock cognate 71 kDa protein |
| TFEB | transcription factor EB |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| MMP | mitochondrial membrane potential |
| Nrf2 | Nuclear Factor, Erythroid 2 Like 2 |
| ARDs | age-related diseases |
| CRM | caloric restriction mimetic |
| p70S6K | phosphorylation of p70 ribosomal protein S6 kinase |
| SOCE | store operated calcium entry |
| TMZ | Temozolomide |
| MGMT | O6-methylguanine-DNA methyltransferase |
| TSC2 | Tuberous Sclerosis Complex 2 |
| PKC-δ | protein kinase C δ |
| TG2 | Transglutaminase |
| CSCs | cancer stem cells |
| 6-CEPN | 6-C-(E-phenylethenyl)naringenin |
| BUC | bladder urothelial carcinoma |
| PERK | phosphorylated extracellular signal-regulated kinase |
| eIF2α | Eukaryotic initiation factor 2α |
| CHOP | C/EBP homologous protein |
| CaMMK | Calmodulin- dependent kinase protein kinase |
| IRE1α | inositol-requiring enzyme 1α |
| TNBC | triple-negative breast cancer |
| MPT | mitochondrial permeability transition |
| SERCA | Sarcoplasmic-/ endoplasmic reticulum sarco/ER Ca2+ ATPase |
| TG | Thapsigargin |
| TBMS1 | Tubeimoside-1 |
| ERK1/2 | Extracellular signal-regulated kinase 1/2 |
| PTP1B | protein-tyrosine phosphatase 1B |
| DRP1 | dynamin-related protein |
| EEPP | ethanolic extract from P. polyphylla |
| DPPE | Diosgenin enriched P. polyphylla rhizome extract |
| BA | Betulinic acid |
| GRP78 | glucose-regulated protein 78 |
| CPT | Camptothecin |
| AP-1 | Activator protein 1 |
| miR-15a | microRNA-15a |
| BBR | Berberine |
| PDT | photodynamic therapy |
| TET | Tetrandrine |
| APL | acute promyelocytic leukemia |
| PA | pituitary adenoma |
| TRAIL | Tumor necrosis factor (TNF)-related apoptosis-inducing ligand |
| TRAF2 | TNF receptor-associated factor 2 |
| FAK | focal adhesion kinase |
| S6K1 | 70-kDa ribosomal S6 kinase 1 |
| TQ | Thymoquinone |
| GBM | glioblastoma multiforme |
| LXRα | liver-X receptors α |
| ABCA1 | ATP-binding cassette transporter A1 |
| ccRCC | clear cell renal cell carcinoma |
| AR | androgen receptor |
| ω3-PUFAs | omega-3 polyunsaturated fatty acids |
| EPA | eicosapentaenoic acid |
| EPADA | EPA-dopamine |
| DHA | docosahexaenoic acid |
| DHADA | DHA-dopamine |
| MM | multiple myeloma |
| PBMCs | peripheral blood mononuclear cells |
| DCs | dendritic cells |
| PPARγ | peroxisome proliferator-activated receptor γ |
| Delta-T3 | vitamin E Delta-tocotrienol |
| HDACIs | histone deacetylase inhibitors |
| TSA | trichostatin A |
| SAHA | suberoylanilide hydroxamic acid |
| FOXO1 | forkhead box protein1 |
| PRMT5 | protein arginine methyltransferase 5 |
| TRPV6 | transient receptor potential cation channel, subfamily V, member 6 |
| STC1 | stanniocalcin 1 |
| IGFBP2 | insulin-like growth factor (IGF) binding protein-2 |
| VSOAC | volume-sensitive organic anion channel |
| TauT | taurine transporter |
| MTP | mitochondrial transmembrane potential |
| CLL | Chronic Lymphocytic Leukaemia |
| ANT | Active, not recruiting |
| T | terminated |
| C | Completed |
| S | Suspended |
| R | Recruitment |
| NR | Not yet recruiting |
| U | Unknown |
| ATRA | all-trans retinoic acid |
| CyP | Cyclophosphamide |
| Cape | Capecitabine |
| PCOS | Polycystic ovarian syndrome |
| FME | familial Mediterranean fever |
| HEKs | human epidermal keratinocytes |
| CSCC | cutaneous squamous cell carcinoma cell |
| NTM | nontuberculous mycobacteria |
| LYNUS | lysosomal nutrient sensing machinery |
References
- Pohl, C.; Dikic, I. Cellular quality control by the ubiquitin-proteasome system and autophagy. Science 2019, 366, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Saftig, P.; Klumperman, J. Lysosome biogenesis and lysosomal membrane proteins: Trafficking meets function. Nat. Rev. Mol. Cell Biol. 2009, 10, 623–635. [Google Scholar] [CrossRef]
- Horibe, A.; Eid, N.; Ito, Y.; Hamaoka, H.; Tanaka, Y.; Kondo, Y. Upregulated autophagy in Sertoli cells of ethanol-treated rats is associated with induction of inducible nitric oxide synthase (iNOS), androgen receptor suppression and germ cell apoptosis. Int. J. Mol. Sci. 2017, 18, 1061. [Google Scholar] [CrossRef] [PubMed]
- Levy, J.M.M.; Towers, C.G.; Thorburn, A. Targeting autophagy in cancer. Nat. Rev. Cancer 2017, 17, 528–542. [Google Scholar] [CrossRef] [PubMed]
- Kaur, J.; Debnath, J. Autophagy at the crossroads of catabolism and anabolism. Nat. Rev. Mol. Cell Biol. 2015, 16, 461–472. [Google Scholar] [CrossRef]
- Dikic, I.; Elazar, Z. Mechanism and medical implications of mammalian autophagy. Nat. Rev. Mol. Cell Biol. 2018, 19, 349–364. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Abdel-Aziz, A.K.; Abdelfatah, S.; Abdellatif, M.; Abdoli, A.; Abel, S.; Abeliovich, H.; Abildgaard, M.H.; Abudu, Y.P.; Acevedo-Arozena, A.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy (4th edition). Autophagy 2021, 17, 1–382. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Pietrocola, F.; Bravo-San Pedro, J.M.; Amaravadi, R.K.; Baehrecke, E.H.; Cecconi, F.; Codogno, P.; Debnath, J.; Gewirtz, D.A.; Karantza, V.; et al. Autophagy in malignant transformation and cancer progression. EMBO J. 2015, 34, 856–880. [Google Scholar] [CrossRef]
- Amaravadi, R.K.; Kimmelman, A.C.; Debnath, J. Targeting Autophagy in Cancer: Recent Advances and Future Directions. Cancer Discov. 2019, 9, 1167–1181. [Google Scholar] [CrossRef]
- Jung, S.; Jeong, H.; Yu, S.W. Autophagy as a decisive process for cell death. Exp. Mol. Med. 2020, 52, 921–930. [Google Scholar] [CrossRef]
- Deng, L.J.; Qi, M.; Li, N.; Lei, Y.H.; Zhang, D.M.; Chen, J.X. Natural products and their derivatives: Promising modulators of tumor immunotherapy. J. Leukoc. Biol. 2020, 108, 493–508. [Google Scholar] [CrossRef]
- Yin, B.; Fang, D.M.; Zhou, X.L.; Gao, F. Natural products as important tyrosine kinase inhibitors. Eur. J. Med. Chem. 2019, 182, 111664. [Google Scholar] [CrossRef]
- Lichota, A.; Gwozdzinski, K. Anticancer Activity of Natural Compounds from Plant and Marine Environment. Int. J. Mol. Sci. 2018, 19, 3533. [Google Scholar] [CrossRef]
- Khalifa, S.A.M.; Elias, N.; Farag, M.A.; Chen, L.; Saeed, A.; Hegazy, M.F.; Moustafa, M.S.; Abd El-Wahed, A.; Al-Mousawi, S.M.; Musharraf, S.G.; et al. Marine Natural Products: A Source of Novel Anticancer Drugs. Mar. Drugs 2019, 17, 491. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Morel, E.; Mehrpour, M.; Botti, J.; Dupont, N.; Hamai, A.; Nascimbeni, A.C.; Codogno, P. Autophagy: A Druggable Process. Annu. Rev. Pharmacol. Toxicol. 2017, 57, 375–398. [Google Scholar] [CrossRef]
- Abada, A.; Elazar, Z. Getting ready for building: Signaling and autophagosome biogenesis. EMBO Rep. 2014, 15, 839–852. [Google Scholar] [CrossRef] [PubMed]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef]
- Sanchez-Wandelmer, J.; Ktistakis, N.T.; Reggiori, F. ERES: Sites for autophagosome biogenesis and maturation? J. Cell. Sci. 2015, 128, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Geng, J.; Klionsky, D.J. Direct quantification of autophagic flux by a single molecule-based probe. Autophagy 2017, 13, 639–641. [Google Scholar] [CrossRef] [PubMed]
- Morishita, H.; Kaizuka, T.; Hama, Y.; Mizushima, N. A new probe to measure autophagic flux in vitro and in vivo. Autophagy 2017, 13, 757–758. [Google Scholar] [CrossRef] [PubMed]
- Shibutani, S.T.; Yoshimori, T. A current perspective of autophagosome biogenesis. Cell Res. 2014, 24, 58–68. [Google Scholar] [CrossRef]
- Hamasaki, M.; Furuta, N.; Matsuda, A.; Nezu, A.; Yamamoto, A.; Fujita, N.; Oomori, H.; Noda, T.; Haraguchi, T.; Hiraoka, Y.; et al. Autophagosomes form at ER-mitochondria contact sites. Nature 2013, 495, 389–393. [Google Scholar] [CrossRef]
- Krols, M.; Bultynck, G.; Janssens, S. ER-Mitochondria contact sites: A new regulator of cellular calcium flux comes into play. J. Cell Biol. 2016, 214, 367–370. [Google Scholar] [CrossRef]
- Polson, H.E.; de Lartigue, J.; Rigden, D.J.; Reedijk, M.; Urbe, S.; Clague, M.J.; Tooze, S.A. Mammalian Atg18 (WIPI2) localizes to omegasome-anchored phagophores and positively regulates LC3 lipidation. Autophagy 2010, 6, 506–522. [Google Scholar] [CrossRef]
- Dooley, H.C.; Razi, M.; Polson, H.E.; Girardin, S.E.; Wilson, M.I.; Tooze, S.A. WIPI2 links LC3 conjugation with PI3P, autophagosome formation, and pathogen clearance by recruiting Atg12-5-16L1. Mol. Cell 2014, 55, 238–252. [Google Scholar] [CrossRef]
- Xie, Z.; Klionsky, D.J. Autophagosome formation: Core machinery and adaptations. Nat. Cell Biol. 2007, 9, 1102–1109. [Google Scholar] [CrossRef]
- Nedelsky, N.B.; Todd, P.K.; Taylor, J.P. Autophagy and the ubiquitin-proteasome system: Collaborators in neuroprotection. Biochim. Biophys. Acta 2008, 1782, 691–699. [Google Scholar] [CrossRef]
- Zientara-Rytter, K.; Subramani, S. AIM/LIR-based fluorescent sensors-new tools to monitor mAtg8 functions. Autophagy 2018, 14, 1074–1078. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Jun, Y.W.; Choi, H.E.; Huh, Y.H.; Kaang, B.K.; Jang, D.J.; Lee, J.A. Development of LC3/GABARAP sensors containing a LIR and a hydrophobic domain to monitor autophagy. EMBO J. 2017, 36, 1100–1116. [Google Scholar] [CrossRef] [PubMed]
- Fracchiolla, D.; Sawa-Makarska, J.; Martens, S. Beyond Atg8 binding: The role of AIM/LIR motifs in autophagy. Autophagy 2017, 13, 978–979. [Google Scholar] [CrossRef]
- Deng, Z.; Lim, J.; Wang, Q.; Purtell, K.; Wu, S.; Palomo, G.M.; Tan, H.; Manfredi, G.; Zhao, Y.; Peng, J.; et al. ALS-FTLD-linked mutations of SQSTM1/p62 disrupt selective autophagy and NFE2L2/NRF2 anti-oxidative stress pathway. Autophagy 2020, 16, 917–931. [Google Scholar] [CrossRef] [PubMed]
- Kast, D.J.; Dominguez, R. The Cytoskeleton-Autophagy Connection. Curr. Biol. 2017, 27, R318–R326. [Google Scholar] [CrossRef] [PubMed]
- Gross, S.P.; Vershinin, M.; Shubeita, G.T. Cargo transport: Two motors are sometimes better than one. Curr. Biol. 2007, 17, R478–R486. [Google Scholar] [CrossRef]
- Korolchuk, V.I.; Saiki, S.; Lichtenberg, M.; Siddiqi, F.H.; Roberts, E.A.; Imarisio, S.; Jahreiss, L.; Sarkar, S.; Futter, M.; Menzies, F.M.; et al. Lysosomal positioning coordinates cellular nutrient responses. Nat. Cell. Biol. 2011, 13, 453–460. [Google Scholar] [CrossRef]
- Pous, C.; Codogno, P. Lysosome positioning coordinates mTORC1 activity and autophagy. Nat. Cell. Biol. 2011, 13, 342–344. [Google Scholar] [CrossRef] [PubMed]
- Pu, J.; Guardia, C.M.; Keren-Kaplan, T.; Bonifacino, J.S. Mechanisms and functions of lysosome positioning. J. Cell Sci. 2016, 129, 4329–4339. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Lian, S.; Khan, A.; Llop, J.R.; Samuelson, A.V.; Chen, W.; Klionsky, D.J.; Kishi, S. Autolysosome biogenesis and developmental senescence are regulated by both Spns1 and v-ATPase. Autophagy 2017, 13, 386–403. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Yoshimori, T. New insights into autophagosome-lysosome fusion. J. Cell Sci. 2017, 130, 1209–1216. [Google Scholar] [CrossRef]
- Chen, Y.; Yu, L. Development of Research into Autophagic Lysosome Reformation. Mol. Cells 2018, 41, 45–49. [Google Scholar] [PubMed]
- Al-Bari, M.A.A.; Xu, P. Molecular regulation of autophagy machinery by mTOR-dependent and -independent pathways. Ann. N. Y. Acad. Sci. 2020, 1467, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Eid, N.; Ito, Y.; Horibe, A.; Otsuki, Y.; Kondo, Y. Ethanol-Induced Mitochondrial Damage in Sertoli Cells is Associated with Parkin Overexpression and Activation of Mitophagy. Cells 2019, 25, 283. [Google Scholar] [CrossRef] [PubMed]
- Tisi, R.; Gaponenko, V.; Vanoni, M.; Sacco, E. Natural Products Attenuating Biosynthesis, Processing, and Activity of Ras Oncoproteins: State of the Art and Future Perspectives. Biomolecules 2020, 10, 1535. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Hannan, M.A.; Dash, R.; Rahman, M.H.; Islam, R.; Uddin, M.J.; Sohag, A.A.M.; Rahman, M.H.; Rhim, H. Phytochemicals as a Complement to Cancer Chemotherapy: Pharmacological Modulation of the Autophagy-Apoptosis Pathway. Front. Pharmacol. 2021, 12, 639628. [Google Scholar] [CrossRef]
- Dischler, N.M.; Xu, L.; Li, Y.; Nichols, C.B.; Alspaugh, J.A.; Bills, G.F.; Gloer, J.B. Wortmannin and Wortmannine Analogues from an Undescribed Niesslia sp. J. Nat. Prod. 2019, 82, 532–538. [Google Scholar] [CrossRef]
- Petiot, A.; Ogier-Denis, E.; Blommaart, E.F.; Meijer, A.J.; Codogno, P. Distinct classes of phosphatidylinositol 3′-kinases are involved in signaling pathways that control macroautophagy in HT-29 cells. J. Biol. Chem. 2000, 275, 992–998. [Google Scholar] [CrossRef]
- Nakanishi, S.; Kakita, S.; Takahashi, I.; Kawahara, K.; Tsukuda, E.; Sano, T.; Yamada, K.; Yoshida, M.; Kase, H.; Matsuda, Y.; et al. Wortmannin, a microbial product inhibitor of myosin light chain kinase. J. Biol. Chem. 1992, 267, 2157–2163. [Google Scholar] [CrossRef]
- Brunn, G.J.; Williams, J.; Sabers, C.; Wiederrecht, G.; Lawrence, J.C., Jr.; Abraham, R.T. Direct inhibition of the signaling functions of the mammalian target of rapamycin by the phosphoinositide 3-kinase inhibitors, wortmannin and LY294002. EMBO J. 1996, 15, 5256–5267. [Google Scholar] [CrossRef]
- Harder, B.G.; Peng, S.; Sereduk, C.P.; Sodoma, A.M.; Kitange, G.J.; Loftus, J.C.; Sarkaria, J.N.; Tran, N.L. Inhibition of phosphatidylinositol 3-kinase by PX-866 suppresses temozolomide-induced autophagy and promotes apoptosis in glioblastoma cells. Mol. Med. 2019, 25, 49. [Google Scholar] [CrossRef]
- Hotte, S.J.; Chi, K.N.; Joshua, A.M.; Tu, D.; Macfarlane, R.J.; Gregg, R.W.; Ruether, J.D.; Basappa, N.S.; Finch, D.; Salim, M.; et al. A Phase II Study of PX-866 in Patients With Recurrent or Metastatic Castration-resistant Prostate Cancer: Canadian Cancer Trials Group Study IND205. Clin. Genitourin. Cancer 2019, 17, 201–208.e1. [Google Scholar] [CrossRef]
- Yam, C.; Xu, X.; Davies, M.A.; Gimotty, P.A.; Morrissette, J.J.D.; Tetzlaff, M.T.; Wani, K.M.; Liu, S.; Deng, W.; Buckley, M.; et al. A Multicenter Phase I Study Evaluating Dual PI3K and BRAF Inhibition with PX-866 and Vemurafenib in Patients with Advanced BRAF V600-Mutant Solid Tumors. Clin. Cancer Res. 2018, 24, 22–32. [Google Scholar] [CrossRef]
- Pitz, M.W.; Eisenhauer, E.A.; MacNeil, M.V.; Thiessen, B.; Easaw, J.C.; Macdonald, D.R.; Eisenstat, D.D.; Kakumanu, A.S.; Salim, M.; Chalchal, H.; et al. Phase II study of PX-866 in recurrent glioblastoma. Neuro-Oncol. 2015, 17, 1270–1274. [Google Scholar] [CrossRef] [PubMed]
- Watanabe-Asano, T.; Kuma, A.; Mizushima, N. Cycloheximide inhibits starvation-induced autophagy through mTORC1 activation. Biochem. Biophys. Res. Commun. 2014, 445, 334–339. [Google Scholar] [CrossRef]
- Blommaart, E.F.; Luiken, J.J.; Blommaart, P.J.; van Woerkom, G.M.; Meijer, A.J. Phosphorylation of ribosomal protein S6 is inhibitory for autophagy in isolated rat hepatocytes. J. Biol. Chem. 1995, 270, 2320–2326. [Google Scholar] [CrossRef] [PubMed]
- Della Sala, G.; Agriesti, F.; Mazzoccoli, C.; Tataranni, T.; Costantino, V.; Piccoli, C. Clogging the Ubiquitin-Proteasome Machinery with Marine Natural Products: Last Decade Update. Mar. Drugs 2018, 16, 467. [Google Scholar] [CrossRef] [PubMed]
- Monti, M.C.; Margarucci, L.; Riccio, R.; Bonfili, L.; Mozzicafreddo, M.; Eleuteri, A.M.; Casapullo, A. Mechanistic insights on petrosaspongiolide M inhibitory effects on immunoproteasome and autophagy. Biochim. Biophys. Acta 2014, 1844, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Geng, X.; Ren, Y.; Wang, F.; Tian, D.; Yao, X.; Zhang, Y.; Tang, J. Harmines inhibit cancer cell growth through coordinated activation of apoptosis and inhibition of autophagy. Biochem. Biophys. Res. Commun. 2018, 498, 99–104. [Google Scholar] [CrossRef]
- Kaul, R.; Risinger, A.L.; Mooberry, S.L. Microtubule-Targeting Drugs: More than Antimitotics. J. Nat. Prod. 2019, 82, 680–685. [Google Scholar] [CrossRef]
- Mukhtar, E.; Adhami, V.M.; Mukhtar, H. Targeting microtubules by natural agents for cancer therapy. Mol. Cancer Ther. 2014, 13, 275–284. [Google Scholar] [CrossRef]
- Karatoprak, G.S.; Kupeli Akkol, E.; Genc, Y.; Bardakci, H.; Yucel, C.; Sobarzo-Sanchez, E. Combretastatins: An Overview of Structure, Probable Mechanisms of Action and Potential Applications. Molecules 2020, 25, 2560. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, W.; Xu, J.; Zhang, T.; Zuo, D.; Zhou, Z.; Lin, B.; Wang, G.; Wang, Z.; Sun, W.; et al. NDRG1 inhibition sensitizes osteosarcoma cells to combretastatin A-4 through targeting autophagy. Cell Death Dis. 2017, 8, e3048. [Google Scholar] [CrossRef]
- Greene, L.M.; O’Boyle, N.M.; Nolan, D.P.; Meegan, M.J.; Zisterer, D.M. The vascular targeting agent Combretastatin-A4 directly induces autophagy in adenocarcinoma-derived colon cancer cells. Biochem. Pharmacol. 2012, 84, 612–624. [Google Scholar] [CrossRef]
- Li, Y.; Luo, P.; Wang, J.; Dai, J.; Yang, X.; Wu, H.; Yang, B.; He, Q. Autophagy blockade sensitizes the anticancer activity of CA-4 via JNK-Bcl-2 pathway. Toxicol. Appl. Pharmacol. 2014, 274, 319–327. [Google Scholar] [CrossRef]
- Zafarullah, M.; Li, W.Q.; Sylvester, J.; Ahmad, M. Molecular mechanisms of N-acetylcysteine actions. Cell Mol. Life Sci. 2003, 60, 6–20. [Google Scholar] [CrossRef]
- Wu, M.S.; Lien, G.S.; Shen, S.C.; Yang, L.Y.; Chen, Y.C. N-acetyl-L-cysteine enhances fisetin-induced cytotoxicity via induction of ROS-independent apoptosis in human colonic cancer cells. Mol. Carcinog. 2014, 53 (Suppl. S1), E119–E129. [Google Scholar] [CrossRef]
- Sasazawa, Y.; Kanagaki, S.; Tashiro, E.; Nogawa, T.; Muroi, M.; Kondoh, Y.; Osada, H.; Imoto, M. Xanthohumol impairs autophagosome maturation through direct inhibition of valosin-containing protein. ACS Chem. Biol. 2012, 7, 892–900. [Google Scholar] [CrossRef]
- Ma, L.; Tang, L.; Yi, Q. Salvianolic Acids: Potential Source of Natural Drugs for the Treatment of Fibrosis Disease and Cancer. Front. Pharmacol. 2019, 10, 97. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Chen, S.; Zhang, W.; Zheng, X.; Hu, S.; Pang, C.; Lu, J.; Xing, J.; Dong, Y. Salvianolic acid A reverses paclitaxel resistance in human breast cancer MCF-7 cells via targeting the expression of transgelin 2 and attenuating PI3 K/Akt pathway. Phytomedicine 2014, 21, 1725–1732. [Google Scholar] [CrossRef]
- Zheng, X.; Chen, S.; Yang, Q.; Cai, J.; Zhang, W.; You, H.; Xing, J.; Dong, Y. Salvianolic acid A reverses the paclitaxel resistance and inhibits the migration and invasion abilities of human breast cancer cells by inactivating transgelin 2. Cancer Biol. Ther. 2015, 16, 1407–1414. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Qiu, S.; Qian, L.; Tian, Y.; Chen, Y.; Bi, L.; Chen, W. OCF can repress tumor metastasis by inhibiting epithelial-mesenchymal transition involved in PTEN/PI3K/AKT pathway in lung cancer cells. PLoS ONE 2017, 12, e0174021. [Google Scholar] [CrossRef]
- Tao, L.; Wang, S.; Zhao, Y.; Sheng, X.; Wang, A.; Zheng, S.; Lu, Y. Phenolcarboxylic acids from medicinal herbs exert anticancer effects through disruption of COX-2 activity. Phytomedicine 2014, 21, 1473–1482. [Google Scholar] [CrossRef]
- Gong, L.; Di, C.; Xia, X.; Wang, J.; Chen, G.; Shi, J.; Chen, P.; Xu, H.; Zhang, W. AKT/mTOR signaling pathway is involved in salvianolic acid B-induced autophagy and apoptosis in hepatocellular carcinoma cells. Int. J. Oncol. 2016, 49, 2538–2548. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.D.; Zhao, Y.; Zhang, M.; He, R.Z.; Shi, X.H.; Guo, X.J.; Shi, C.J.; Peng, F.; Wang, M.; Shen, M.; et al. Inhibition of Autophagy by Deguelin Sensitizes Pancreatic Cancer Cells to Doxorubicin. Int. J. Mol. Sci. 2017, 18, 370. [Google Scholar] [CrossRef]
- Yang, Y.L.; Ji, C.; Bi, Z.G.; Lu, C.C.; Wang, R.; Gu, B.; Cheng, L. Deguelin induces both apoptosis and autophagy in cultured head and neck squamous cell carcinoma cells. PLoS ONE 2013, 8, e54736. [Google Scholar] [CrossRef]
- Khan, N.; Yilmaz, S.; Aksoy, S.; Uzel, A.; Tosun, C.; Kirmizibayrak, P.B.; Bedir, E. Polyethers isolated from the marine actinobacterium Streptomyces cacaoi inhibit autophagy and induce apoptosis in cancer cells. Chem. Biol. Interact. 2019, 307, 167–178. [Google Scholar] [CrossRef]
- Gao, G.; Liu, F.; Xu, Z.; Wan, D.; Han, Y.; Kuang, Y.; Wang, Q.; Zhi, Q. Evidence of nigericin as a potential therapeutic candidate for cancers: A review. Biomed. Pharmacother. 2021, 137, 111262. [Google Scholar] [CrossRef]
- Choi, H.S.; Jeong, E.H.; Lee, T.G.; Kim, S.Y.; Kim, H.R.; Kim, C.H. Autophagy Inhibition with Monensin Enhances Cell Cycle Arrest and Apoptosis Induced by mTOR or Epidermal Growth Factor Receptor Inhibitors in Lung Cancer Cells. Tuberc. Respir. Dis. 2013, 75, 9–17. [Google Scholar] [CrossRef][Green Version]
- Knott, S.R.V.; Wagenblast, E.; Khan, S.; Kim, S.Y.; Soto, M.; Wagner, M.; Turgeon, M.O.; Fish, L.; Erard, N.; Gable, A.L.; et al. Asparagine bioavailability governs metastasis in a model of breast cancer. Nature 2018, 554, 378–381. [Google Scholar] [CrossRef]
- Heo, Y.A.; Syed, Y.Y.; Keam, S.J. Pegaspargase: A Review in Acute Lymphoblastic Leukaemia. Drugs 2019, 79, 767–777. [Google Scholar] [CrossRef]
- Takahashi, H.; Inoue, J.; Sakaguchi, K.; Takagi, M.; Mizutani, S.; Inazawa, J. Autophagy is required for cell survival under L-asparaginase-induced metabolic stress in acute lymphoblastic leukemia cells. Oncogene 2017, 36, 4267–4276. [Google Scholar] [CrossRef]
- Li, H.; Meng, X.X.; Zhang, L.; Zhang, B.J.; Liu, X.Y.; Fu, W.W.; Tan, H.S.; Lao, Y.Z.; Xu, H.X. Oblongifolin C and guttiferone K extracted from Garcinia yunnanensis fruit synergistically induce apoptosis in human colorectal cancer cells in vitro. Acta Pharmacol. Sin. 2017, 38, 252–263. [Google Scholar] [CrossRef]
- Feng, C.; Zhou, L.Y.; Yu, T.; Xu, G.; Tian, H.L.; Xu, J.J.; Xu, H.X.; Luo, K.Q. A new anticancer compound, oblongifolin C, inhibits tumor growth and promotes apoptosis in HeLa cells through Bax activation. Int. J. Cancer 2012, 131, 1445–1454. [Google Scholar] [CrossRef]
- Lao, Y.; Wan, G.; Liu, Z.; Wang, X.; Ruan, P.; Xu, W.; Xu, D.; Xie, W.; Zhang, Y.; Xu, H.; et al. The natural compound oblongifolin C inhibits autophagic flux and enhances antitumor efficacy of nutrient deprivation. Autophagy 2014, 10, 736–749. [Google Scholar] [CrossRef]
- Wu, M.; Lao, Y.; Xu, N.; Wang, X.; Tan, H.; Fu, W.; Lin, Z.; Xu, H. Guttiferone K induces autophagy and sensitizes cancer cells to nutrient stress-induced cell death. Phytomedicine 2015, 22, 902–910. [Google Scholar] [CrossRef]
- Kan, W.L.; Yin, C.; Xu, H.X.; Xu, G.; To, K.K.; Cho, C.H.; Rudd, J.A.; Lin, G. Antitumor effects of novel compound, guttiferone K, on colon cancer by p21Waf1/Cip1-mediated G(0)/G(1) cell cycle arrest and apoptosis. Int. J. Cancer 2013, 132, 707–716. [Google Scholar] [CrossRef]
- Renna, M.; Schaffner, C.; Brown, K.; Shang, S.; Tamayo, M.H.; Hegyi, K.; Grimsey, N.J.; Cusens, D.; Coulter, S.; Cooper, J.; et al. Azithromycin blocks autophagy and may predispose cystic fibrosis patients to mycobacterial infection. J. Clin. Investig. 2011, 121, 3554–3563. [Google Scholar] [CrossRef]
- Nakamura, M.; Kikukawa, Y.; Takeya, M.; Mitsuya, H.; Hata, H. Clarithromycin attenuates autophagy in myeloma cells. Int. J. Oncol. 2010, 37, 815–820. [Google Scholar]
- Wang, Z.; Zhang, J.; Wang, Y.; Xing, R.; Yi, C.; Zhu, H.; Chen, X.; Guo, J.; Guo, W.; Li, W.; et al. Matrine, a novel autophagy inhibitor, blocks trafficking and the proteolytic activation of lysosomal proteases. Carcinogenesis 2013, 34, 128–138. [Google Scholar] [CrossRef]
- Xie, S.B.; He, X.X.; Yao, S.K. Matrine-induced autophagy regulated by p53 through AMP-activated protein kinase in human hepatoma cells. Int. J. Oncol. 2015, 47, 517–526. [Google Scholar] [CrossRef]
- Zou, Y.; Sarem, M.; Xiang, S.; Hu, H.; Xu, W.; Shastri, V.P. Autophagy inhibition enhances Matrine derivative MASM induced apoptosis in cancer cells via a mechanism involving reactive oxygen species-mediated PI3K/Akt/mTOR and Erk/p38 signaling. BMC Cancer 2019, 19, 949. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Fang, Y.; Yang, Y.; Qin, Y.; Wu, P.; Wang, T.; Lai, H.; Meng, L.; Wang, D.; Zheng, Z.; et al. Elaiophylin, a novel autophagy inhibitor, exerts antitumor activity as a single agent in ovarian cancer cells. Autophagy 2015, 11, 1849–1863. [Google Scholar] [CrossRef]
- Wang, G.; Zhou, P.; Chen, X.; Zhao, L.; Tan, J.; Yang, Y.; Fang, Y.; Zhou, J. The novel autophagy inhibitor elaiophylin exerts antitumor activity against multiple myeloma with mutant TP53 in part through endoplasmic reticulum stress-induced apoptosis. Cancer Biol. Ther. 2017, 18, 584–595. [Google Scholar] [CrossRef]
- Pinto, M.M.M.; Palmeira, A.; Fernandes, C.; Resende, D.; Sousa, E.; Cidade, H.; Tiritan, M.E.; Correia-da-Silva, M.; Cravo, S. From Natural Products to New Synthetic Small Molecules: A Journey through the World of Xanthones. Molecules 2021, 26, 431. [Google Scholar] [CrossRef]
- Mattsson, J.P.; Vaananen, K.; Wallmark, B.; Lorentzon, P. Omeprazole and bafilomycin, two proton pump inhibitors: Differentiation of their effects on gastric, kidney and bone H+-translocating ATPases. Biochim. Biophys. Acta 1991, 1065, 261–268. [Google Scholar] [CrossRef]
- Yuan, N.; Song, L.; Zhang, S.; Lin, W.; Cao, Y.; Xu, F.; Fang, Y.; Wang, Z.; Zhang, H.; Li, X.; et al. Bafilomycin A1 targets both autophagy and apoptosis pathways in pediatric B-cell acute lymphoblastic leukemia. Haematologica 2015, 100, 345–356. [Google Scholar] [CrossRef]
- Yamamoto, A.; Tagawa, Y.; Yoshimori, T.; Moriyama, Y.; Masaki, R.; Tashiro, Y. Bafilomycin A1 prevents maturation of autophagic vacuoles by inhibiting fusion between autophagosomes and lysosomes in rat hepatoma cell line, H-4-II-E cells. Cell. Struct. Funct. 1998, 23, 33–42. [Google Scholar] [CrossRef]
- Huss, M.; Ingenhorst, G.; Konig, S.; Gassel, M.; Drose, S.; Zeeck, A.; Altendorf, K.; Wieczorek, H. Concanamycin A, the specific inhibitor of V-ATPases, binds to the V(o) subunit c. J. Biol. Chem. 2002, 277, 40544–40548. [Google Scholar] [CrossRef]
- Kallifatidis, G.; Hoepfner, D.; Jaeg, T.; Guzman, E.A.; Wright, A.E. The marine natural product manzamine A targets vacuolar ATPases and inhibits autophagy in pancreatic cancer cells. Mar. Drugs 2013, 11, 3500–3516. [Google Scholar] [CrossRef]
- Guzman, E.A.; Johnson, J.D.; Linley, P.A.; Gunasekera, S.E.; Wright, A.E. A novel activity from an old compound: Manzamine A reduces the metastatic potential of AsPC-1 pancreatic cancer cells and sensitizes them to TRAIL-induced apoptosis. Investig. New Drugs 2011, 29, 777–785. [Google Scholar] [CrossRef]
- Chen, C.; Lu, L.; Yan, S.; Yi, H.; Yao, H.; Wu, D.; He, G.; Tao, X.; Deng, X. Autophagy and doxorubicin resistance in cancer. Anticancer Drugs 2018, 29, 1–9. [Google Scholar] [CrossRef]
- Shabalala, S.; Muller, C.J.F.; Louw, J.; Johnson, R. Polyphenols, autophagy and doxorubicin-induced cardiotoxicity. Life Sci. 2017, 180, 160–170. [Google Scholar] [CrossRef]
- Piao, S.; Amaravadi, R.K. Targeting the lysosome in cancer. Ann. N. Y. Acad. Sci. 2016, 1371, 45–54. [Google Scholar] [CrossRef]
- Haspel, J.; Shaik, R.S.; Ifedigbo, E.; Nakahira, K.; Dolinay, T.; Englert, J.A.; Choi, A.M. Characterization of macroautophagic flux in vivo using a leupeptin-based assay. Autophagy 2011, 7, 629–642. [Google Scholar] [CrossRef] [PubMed]
- Lima, R.T.; Sousa, D.; Paiva, A.M.; Palmeira, A.; Barbosa, J.; Pedro, M.; Pinto, M.M.; Sousa, E.; Vasconcelos, M.H. Modulation of Autophagy by a Thioxanthone Decreases the Viability of Melanoma Cells. Molecules 2016, 21, 1343. [Google Scholar] [CrossRef]
- Madeo, F.; Zimmermann, A.; Maiuri, M.C.; Kroemer, G. Essential role for autophagy in life span extension. J. Clin. Investig. 2015, 125, 85–93. [Google Scholar] [CrossRef]
- Li, J.; Kim, S.G.; Blenis, J. Rapamycin: One drug, many effects. Cell. Metab. 2014, 19, 373–379. [Google Scholar] [CrossRef]
- Lamming, D.W.; Ye, L.; Sabatini, D.M.; Baur, J.A. Rapalogs and mTOR inhibitors as anti-aging therapeutics. J. Clin. Investig. 2013, 123, 980–989. [Google Scholar] [CrossRef]
- Liu, Y.; Li, C.L.; Xu, Q.Q.; Cheng, D.; Liu, K.D.; Sun, Z.Q. Quercetin inhibits invasion and angiogenesis of esophageal cancer cells. Pathol. Res. Pract. 2021, 222, 153455. [Google Scholar] [CrossRef] [PubMed]
- Almatroodi, S.A.; Alsahli, M.A.; Almatroudi, A.; Verma, A.K.; Aloliqi, A.; Allemailem, K.S.; Khan, A.A.; Rahmani, A.H. Potential Therapeutic Targets of Quercetin, a Plant Flavonol, and Its Role in the Therapy of Various Types of Cancer through the Modulation of Various Cell Signaling Pathways. Molecules 2021, 26, 1315. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Tavakol, S.; Ahmadi, Z.; Roomiani, S.; Mohammadinejad, R.; Samarghandian, S. Therapeutic effects of kaempferol affecting autophagy and endoplasmic reticulum stress. Phytother. Res. 2020, 34, 911–923. [Google Scholar] [CrossRef]
- Sameiyan, E.; Hayes, A.W.; Karimi, G. The effect of medicinal plants on multiple drug resistance through autophagy: A review of in vitro studies. Eur. J. Pharmacol. 2019, 852, 244–253. [Google Scholar] [CrossRef]
- Yang, J.; Pi, C.; Wang, G. Inhibition of PI3K/Akt/mTOR pathway by apigenin induces apoptosis and autophagy in hepatocellular carcinoma cells. Biomed. Pharmacother. 2018, 103, 699–707. [Google Scholar] [CrossRef]
- Fujiki, H.; Watanabe, T.; Sueoka, E.; Rawangkan, A.; Suganuma, M. Cancer Prevention with Green Tea and Its Principal Constituent, EGCG: From Early Investigations to Current Focus on Human Cancer Stem Cells. Mol. Cells 2018, 41, 73–82. [Google Scholar]
- Holczer, M.; Besze, B.; Zambo, V.; Csala, M.; Banhegyi, G.; Kapuy, O. Epigallocatechin-3-Gallate (EGCG) Promotes Autophagy-Dependent Survival via Influencing the Balance of mTOR-AMPK Pathways upon Endoplasmic Reticulum Stress. Oxid. Med. Cell Longev. 2018, 2018, 6721530. [Google Scholar] [CrossRef] [PubMed]
- Prietsch, R.F.; Monte, L.G.; da Silva, F.A.; Beira, F.T.; Del Pino, F.A.; Campos, V.F.; Collares, T.; Pinto, L.S.; Spanevello, R.M.; Gamaro, G.D.; et al. Genistein induces apoptosis and autophagy in human breast MCF-7 cells by modulating the expression of proapoptotic factors and oxidative stress enzymes. Mol. Cell. Biochem. 2014, 390, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, R.; Kang, Y.; Li, X.; Roife, D.; Zhang, R.; Fleming, J.B. Genistein potentiates the antitumor effect of 5-Fluorouracil by inducing apoptosis and autophagy in human pancreatic cancer cells. Anticancer Res. 2014, 34, 4685–4692. [Google Scholar]
- Wedel, S.; Manola, M.; Cavinato, M.; Trougakos, I.P.; Jansen-Durr, P. Targeting Protein Quality Control Mechanisms by Natural Products to Promote Healthy Ageing. Molecules 2018, 23, 1219. [Google Scholar] [CrossRef]
- Hsiao, Y.T.; Kuo, C.L.; Chueh, F.S.; Liu, K.C.; Bau, D.T.; Chung, J.G. Curcuminoids Induce Reactive Oxygen Species and Autophagy to Enhance Apoptosis in Human Oral Cancer Cells. Am. J. Chin. Med. 2018, 46, 1145–1168. [Google Scholar] [CrossRef] [PubMed]
- Hashemzaei, M.; Entezari Heravi, R.; Rezaee, R.; Roohbakhsh, A.; Karimi, G. Regulation of autophagy by some natural products as a potential therapeutic strategy for cardiovascular disorders. Eur. J. Pharmacol. 2017, 802, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Shanmugam, M.K.; Kumar, A.P.; Yap, C.T.; Sethi, G.; Bishayee, A. Targeting autophagy using natural compounds for cancer prevention and therapy. Cancer 2019, 125, 1228–1246. [Google Scholar] [CrossRef]
- Xiaokaiti, Y.; Li, X. Natural Product Regulates Autophagy in Cancer. Adv. Exp. Med. Biol. 2020, 1207, 709–724. [Google Scholar]
- Zhang, S.F.; Wang, X.L.; Yang, X.Q.; Chen, N. Autophagy-associated targeting pathways of natural products during cancer treatment. Asian Pac. J. Cancer Prev. 2014, 15, 10557–10563. [Google Scholar] [CrossRef]
- Huang, X.M.; Yang, Z.J.; Xie, Q.; Zhang, Z.K.; Zhang, H.; Ma, J.Y. Natural products for treating colorectal cancer: A mechanistic review. Biomed. Pharmacother. 2019, 117, 109142. [Google Scholar] [CrossRef]
- Wang, S.F.; Wu, M.Y.; Cai, C.Z.; Li, M.; Lu, J.H. Autophagy modulators from traditional Chinese medicine: Mechanisms and therapeutic potentials for cancer and neurodegenerative diseases. J. Ethnopharmacol. 2016, 194, 861–876. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, L.X.; Ouyang, L.; Cheng, Y.; Liu, B. Plant natural compounds: Targeting pathways of autophagy as anti-cancer therapeutic agents. Cell Prolif. 2012, 45, 466–476. [Google Scholar] [CrossRef]
- Endo, S.; Hoshi, M.; Matsunaga, T.; Inoue, T.; Ichihara, K.; Ikari, A. Autophagy inhibition enhances anticancer efficacy of artepillin C, a cinnamic acid derivative in Brazilian green propolis. Biochem. Biophys. Res. Commun. 2018, 497, 437–443. [Google Scholar] [CrossRef]
- Park, B.S.; Choi, N.E.; Lee, J.H.; Kang, H.M.; Yu, S.B.; Kim, H.J.; Kang, H.K.; Kim, I.R. Crosstalk between Fisetin-induced Apoptosis and Autophagy in Human Oral Squamous Cell Carcinoma. J. Cancer 2019, 10, 138–146. [Google Scholar] [CrossRef]
- Lee, J.W.; Choi, H.J.; Kim, E.J.; Hwang, W.Y.; Jung, M.H.; Kim, K.S. Fisetin induces apoptosis in uterine leiomyomas through multiple pathways. Sci. Rep. 2020, 10, 7993. [Google Scholar] [CrossRef]
- Jia, S.; Xu, X.; Zhou, S.; Chen, Y.; Ding, G.; Cao, L. Fisetin induces autophagy in pancreatic cancer cells via endoplasmic reticulum stress- and mitochondrial stress-dependent pathways. Cell Death Dis. 2019, 10, 142. [Google Scholar] [CrossRef]
- Liu, X.; Duan, C.; Ji, J.; Zhang, T.; Yuan, X.; Zhang, Y.; Ma, W.; Yang, J.; Yang, L.; Jiang, Z.; et al. Cucurbitacin B induces autophagy and apoptosis by suppressing CIP2A/PP2A/mTORC1 signaling axis in human cisplatin resistant gastric cancer cells. Oncol. Rep. 2017, 38, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Sha, T.; Guo, J.; Li, W.; Lu, J.; Chen, X. Cucurbitacin B induces DNA damage and autophagy mediated by reactive oxygen species (ROS) in MCF-7 breast cancer cells. J. Nat. Med. 2015, 69, 522–530. [Google Scholar] [CrossRef]
- Tan, H.; Li, X.; Yang, W.H.; Kang, Y. A flavone, Wogonin from Scutellaria baicalensis inhibits the proliferation of human colorectal cancer cells by inducing of autophagy, apoptosis and G2/M cell cycle arrest via modulating the PI3K/AKT and STAT3 signalling pathways. J. BUON 2019, 24, 1143–1149. [Google Scholar]
- Feng, Q.; Wang, H.; Pang, J.; Ji, L.; Han, J.; Wang, Y.; Qi, X.; Liu, Z.; Lu, L. Prevention of Wogonin on Colorectal Cancer Tumorigenesis by Regulating p53 Nuclear Translocation. Front. Pharmacol. 2018, 9, 1356. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Park, S.H. Induction of cytoprotective autophagy by morusin via AMP-activated protein kinase activation in human non-small cell lung cancer cells. Nutr. Res. Pract. 2020, 14, 478–489. [Google Scholar] [CrossRef]
- Wang, J.; Liu, X.; Zheng, H.; Liu, Q.; Zhang, H.; Wang, X.; Shen, T.; Wang, S.; Ren, D. Morusin induces apoptosis and autophagy via JNK, ERK and PI3K/Akt signaling in human lung carcinoma cells. Chem. Biol. Interact. 2020, 331, 109279. [Google Scholar] [CrossRef]
- Singh, B.N.; Kumar, D.; Shankar, S.; Srivastava, R.K. Rottlerin induces autophagy which leads to apoptotic cell death through inhibition of PI3K/Akt/mTOR pathway in human pancreatic cancer stem cells. Biochem. Pharmacol. 2012, 84, 1154–1163. [Google Scholar] [CrossRef]
- Kumar, D.; Shankar, S.; Srivastava, R.K. Rottlerin induces autophagy and apoptosis in prostate cancer stem cells via PI3K/Akt/mTOR signaling pathway. Cancer Lett. 2014, 343, 179–189. [Google Scholar] [CrossRef]
- Maioli, E.; Daveri, E.; Maellaro, E.; Ietta, F.; Cresti, L.; Valacchi, G. Non-conventional rottlerin anticancer properties. Arch. Biochem. Biophys. 2018, 645, 50–53. [Google Scholar] [CrossRef]
- Mosca, L.; Ilari, A.; Fazi, F.; Assaraf, Y.G.; Colotti, G. Taxanes in cancer treatment: Activity, chemoresistance and its overcoming. Drug Resist. Update 2021, 54, 100742. [Google Scholar] [CrossRef]
- Zou, S.H.; Du, X.; Lin, H.; Wang, P.C.; Li, M. Paclitaxel inhibits the progression of cervical cancer by inhibiting autophagy via lncRNARP11-381N20.2. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3010–3017. [Google Scholar] [PubMed]
- Tiwari, R.V.; Parajuli, P.; Sylvester, P.W. Synergistic anticancer effects of combined gamma-tocotrienol and oridonin treatment is associated with the induction of autophagy. Mol. Cell Biochem. 2015, 408, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yu, H.; Wu, R.; Chen, Z.Y.; Hu, Q.; Zhang, Y.F.; Gao, S.H.; Zhou, G.B. Autophagy inhibition enhances the inhibitory effects of ursolic acid on lung cancer cells. Int. J. Mol. Med. 2020, 46, 1816–1826. [Google Scholar] [CrossRef]
- Lin, C.W.; Chin, H.K.; Lee, S.L.; Chiu, C.F.; Chung, J.G.; Lin, Z.Y.; Wu, C.Y.; Liu, Y.C.; Hsiao, Y.T.; Feng, C.H.; et al. Ursolic acid induces apoptosis and autophagy in oral cancer cells. Environ. Toxicol. 2019, 34, 983–991. [Google Scholar] [CrossRef]
- Zhang, R.; Pan, T.; Xiang, Y.; Zhang, M.; Feng, J.; Liu, S.; Duan, T.; Chen, P.; Zhai, B.; Chen, X.; et al. beta-Elemene Reverses the Resistance of p53-Deficient Colorectal Cancer Cells to 5-Fluorouracil by Inducing Pro-death Autophagy and Cyclin D3-Dependent Cycle Arrest. Front. Bioeng. Biotechnol. 2020, 8, 378. [Google Scholar] [CrossRef]
- Liu, S.; Li, Q.; Li, G.; Zhang, Q.; Zhuo, L.; Han, X.; Zhang, M.; Chen, X.; Pan, T.; Yan, L.; et al. The mechanism of m(6)A methyltransferase METTL3-mediated autophagy in reversing gefitinib resistance in NSCLC cells by beta-elemene. Cell Death Dis. 2020, 11, 969. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhu, J.; Wu, J.; Huang, N.; Cui, Z.; Luo, Y.; Sun, F.; Pan, Q.; Li, Y.; Yang, Q. (−)-Guaiol regulates autophagic cell death depending on mTOR signaling in NSCLC. Cancer Biol. Ther. 2018, 19, 706–714. [Google Scholar] [CrossRef]
- Tian, S.; Chen, Y.; Yang, B.; Lou, C.; Zhu, R.; Zhao, Y.; Zhao, H. F1012-2 inhibits the growth of triple negative breast cancer through induction of cell cycle arrest, apoptosis, and autophagy. Phytother. Res. 2018, 32, 908–922. [Google Scholar] [CrossRef]
- Jiang, S.L.; Guan, Y.D.; Chen, X.S.; Ge, P.; Wang, X.L.; Lao, Y.Z.; Xiao, S.S.; Zhang, Y.; Yang, J.M.; Xu, X.J.; et al. Tubeimoside-1, a triterpenoid saponin, induces cytoprotective autophagy in human breast cancer cells in vitro via Akt-mediated pathway. Acta Pharmacol. Sin. 2019, 40, 919–928. [Google Scholar] [CrossRef]
- Islam, M.S.; Wang, C.; Zheng, J.; Paudyal, N.; Zhu, Y.; Sun, H. The potential role of tubeimosides in cancer prevention and treatment. Eur. J. Med. Chem. 2019, 162, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Ruan, C.; You, L.; Qiu, Y.; Cui, X.; Wu, D. Tubeimoside I induces autophagy in HepG2 cells by activating the AMP-activated protein kinase signaling pathway. Oncol. Lett. 2020, 20, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.T.; Uen, W.C.; Choong, C.Y.; Shi, Y.C.; Lee, B.H.; Tai, C.J.; Tai, C.J. Paris Polyphylla Inhibits Colorectal Cancer Cells via Inducing Autophagy and Enhancing the Efficacy of Chemotherapeutic Drug Doxorubicin. Molecules 2019, 24, 2102. [Google Scholar] [CrossRef]
- Chen, M.; Guo, Y.; Zhao, R.; Wang, X.; Jiang, M.; Fu, H.; Zhang, X. Ophiopogonin B induces apoptosis, mitotic catastrophe and autophagy in A549 cells. Int. J. Oncol. 2016, 49, 316–324. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, J.; Chen, Y. Betulinic acid and the pharmacological effects of tumor suppression (Review). Mol. Med. Rep. 2016, 14, 4489–4495. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Li, S.; Qu, Z.; Luo, Y.; Chen, R.; Wei, S.; Yang, X.; Wang, Q. Betulinic acid induces autophagy-mediated apoptosis through suppression of the PI3K/AKT/mTOR signaling pathway and inhibits hepatocellular carcinoma. Am. J. Transl. Res. 2019, 11, 6952–6964. [Google Scholar] [PubMed]
- Wang, S.; Wang, K.; Zhang, C.; Zhang, W.; Xu, Q.; Wang, Y.; Zhang, Y.; Li, Y.; Zhang, Y.; Zhu, H.; et al. Overaccumulation of p53-mediated autophagy protects against betulinic acid-induced apoptotic cell death in colorectal cancer cells. Cell Death Dis. 2017, 8, e3087. [Google Scholar] [CrossRef]
- Hua, F.; Shang, S.; Hu, Z.W. Seeking new anti-cancer agents from autophagy-regulating natural products. J. Asian Nat. Prod. Res. 2017, 19, 305–313. [Google Scholar] [CrossRef]
- Jayasooriya, R.; Dilshara, M.G.; Karunarathne, W.; Molagoda, I.M.N.; Choi, Y.H.; Kim, G.Y. Camptothecin enhances c-Myc-mediated endoplasmic reticulum stress and leads to autophagy by activating Ca2+-mediated AMPK. Food Chem. Toxicol. 2018, 121, 648–656. [Google Scholar] [CrossRef]
- Heng, Y.; Liang, Y.; Zhang, J.; Li, L.; Zhang, W.; Jiang, Y.; Wang, S.; Jia, L. Camptothecin Inhibits Neddylation to Activate the Protective Autophagy Through NF-kappaB/AMPK/mTOR/ULK1 Axis in Human Esophageal Cancer Cells. Front. Oncol. 2021, 11, 671180. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, X.; Cao, S.; Sun, Y.; He, X.; Jiang, B.; Yu, Y.; Duan, J.; Qiu, F.; Kang, N. Berberine represses human gastric cancer cell growth in vitro and in vivo by inducing cytostatic autophagy via inhibition of MAPK/mTOR/p70S6K and Akt signaling pathways. Biomed. Pharmacother. 2020, 128, 110245. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Y.; Du, X.; Ma, H.; Yao, J. The Anti-Cancer Mechanisms of Berberine: A Review. Cancer Manag. Res. 2020, 12, 695–702. [Google Scholar] [CrossRef]
- Lopes, T.Z.; de Moraes, F.R.; Tedesco, A.C.; Arni, R.K.; Rahal, P.; Calmon, M.F. Berberine associated photodynamic therapy promotes autophagy and apoptosis via ROS generation in renal carcinoma cells. Biomed. Pharmacother. 2020, 123, 109794. [Google Scholar] [CrossRef]
- Luan, F.; He, X.; Zeng, N. Tetrandrine: A review of its anticancer potentials, clinical settings, pharmacokinetics and drug delivery systems. J. Pharm. Pharmacol. 2020, 72, 1491–1512. [Google Scholar] [CrossRef]
- Wang, J.; Yao, Z.; Lai, X.; Bao, H.; Li, Y.; Li, S.; Chang, L.; Zhang, G. Tetrandrine sensitizes nasopharyngeal carcinoma cells to irradiation by inducing autophagy and inhibiting MEK/ERK pathway. Cancer Med. 2020, 9, 7268–7278. [Google Scholar] [CrossRef]
- Guo, Y.; Pei, X. Tetrandrine-Induced Autophagy in MDA-MB-231 Triple-Negative Breast Cancer Cell through the Inhibition of PI3K/AKT/mTOR Signaling. Evid. Based Complement. Alternat. Med. 2019, 2019, 7517431. [Google Scholar] [CrossRef]
- Son, Y.; An, Y.; Jung, J.; Shin, S.; Park, I.; Gwak, J.; Ju, B.G.; Chung, Y.H.; Na, M.; Oh, S. Protopine isolated from Nandina domestica induces apoptosis and autophagy in colon cancer cells by stabilizing p53. Phytother Res. 2019, 33, 1689–1696. [Google Scholar] [CrossRef]
- Marthandam Asokan, S.; Mariappan, R.; Muthusamy, S.; Velmurugan, B.K. Pharmacological benefits of neferine—A comprehensive review. Life Sci. 2018, 199, 60–70. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, X.; Li, Y.; Lu, S.; Lu, S.; Li, J.; Wang, Y.; Tian, X.; Wei, J.J.; Shao, C.; et al. Neferine induces autophagy of human ovarian cancer cells via p38 MAPK/ JNK activation. Tumour Biol. 2016, 37, 8721–8729. [Google Scholar] [CrossRef] [PubMed]
- Law, B.Y.K.; Michelangeli, F.; Qu, Y.Q.; Xu, S.W.; Han, Y.; Mok, S.W.F.; Dias, I.; Javed, M.U.; Chan, W.K.; Xue, W.W.; et al. Neferine induces autophagy-dependent cell death in apoptosis-resistant cancers via ryanodine receptor and Ca2+-dependent mechanism. Sci. Rep. 2019, 9, 20034. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Bishayee, K.; Khuda-Bukhsh, A.R. Graveoline isolated from ethanolic extract of Ruta graveolens triggers apoptosis and autophagy in skin melanoma cells: A novel apoptosis-independent autophagic signaling pathway. Phytother Res. 2014, 28, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Mostofa, A.G.M.; Hossain, M.K.; Basak, D.; Bin Sayeed, M.S. Thymoquinone as a Potential Adjuvant Therapy for Cancer Treatment: Evidence from Preclinical Studies. Front. Pharmacol. 2017, 8, 295. [Google Scholar] [CrossRef]
- Almajali, B.; Al-Jamal, H.A.N.; Taib, W.R.W.; Ismail, I.; Johan, M.F.; Doolaanea, A.A.; Ibrahim, W.N. Thymoquinone, as a Novel Therapeutic Candidate of Cancers. Pharmaceuticals 2021, 14, 369. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fan, Y.; Huang, S.; Wang, G.; Han, R.; Lei, F.; Luo, A.; Jing, X.; Zhao, L.; Gu, S.; et al. Thymoquinone inhibits the metastasis of renal cell cancer cells by inducing autophagy via AMPK/mTOR signaling pathway. Cancer Sci. 2018, 109, 3865–3873. [Google Scholar] [CrossRef]
- Bashmail, H.A.; Alamoudi, A.A.; Noorwali, A.; Hegazy, G.; Ajabnoor, G.; Choudhry, H.; Al-Abd, A.M. Thymoquinone synergizes gemcitabine anti-breast cancer activity via modulating its apoptotic and autophagic activities. Sci. Rep. 2018, 8, 11674. [Google Scholar] [CrossRef]
- Ren, B.; Liu, H.; Gao, H.; Liu, S.; Zhang, Z.; Fribley, A.M.; Callaghan, M.U.; Xu, Z.; Zeng, Q.; Li, Y. Celastrol induces apoptosis in hepatocellular carcinoma cells via targeting ER-stress/UPR. Oncotarget 2017, 8, 93039–93050. [Google Scholar] [CrossRef]
- Chen, Y.; Ou, Y.; Tao, Y.; Liu, H.; Yin, H.; Zhong, S.; Yu, H.; Zhao, Z.; He, B. Effect and mechanisms of celastrol on the apoptosis of HOS osteosarcoma cells. Oncol. Rep. 2018, 40, 2260–2268. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Y.; Zhang, J.; Sun, L.L.; Li, B.H.; Gao, H.L.; Xie, T.; Zhang, N.; Ye, Z.M. Celastrol induces apoptosis and autophagy via the ROS/JNK signaling pathway in human osteosarcoma cells: An in vitro and in vivo study. Cell Death Dis. 2015, 6, e1604. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Liu, Y.; Zhong, J.; Bi, Y.; Liu, Y.; Ren, Z.; Li, X.; Jia, J.; Yu, M.; Yu, X. Pristimerin induces apoptosis and autophagy via activation of ROS/ASK1/JNK pathway in human breast cancer in vitro and in vivo. Cell Death Discov. 2019, 5, 125. [Google Scholar] [CrossRef]
- Li, J.J.; Yan, Y.Y.; Sun, H.M.; Liu, Y.; Su, C.Y.; Chen, H.B.; Zhang, J.Y. Anti-Cancer Effects of Pristimerin and the Mechanisms: A Critical Review. Front. Pharmacol. 2019, 10, 746. [Google Scholar] [CrossRef]
- Yin, Z.; Zhang, J.; Chen, L.; Guo, Q.; Yang, B.; Zhang, W.; Kang, W. Anticancer Effects and Mechanisms of Action of Plumbagin: Review of Research Advances. Biomed. Res. Int. 2020, 2020, 6940953. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Kan, H.; Liu, Y.; Ding, W. Plumbagin induces Ishikawa cell cycle arrest, autophagy, and apoptosis via the PI3K/Akt signaling pathway in endometrial cancer. Food Chem. Toxicol. 2021, 148, 111957. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Chen, Y.; Wang, S.; Ma, J.; Peng, Y.; Yuan, X.; Lv, B.; Chen, W.; Wei, Y. Plumbagin induces autophagy and apoptosis of SMMC-7721 cells in vitro and in vivo. J. Cell Biochem. 2019, 120, 9820–9830. [Google Scholar] [CrossRef] [PubMed]
- Xue, D.; Pan, S.T.; Zhou, X.; Ye, F.; Zhou, Q.; Shi, F.; He, F.; Yu, H.; Qiu, J. Plumbagin Enhances the Anticancer Efficacy of Cisplatin by Increasing Intracellular ROS in Human Tongue Squamous Cell Carcinoma. Oxid. Med. Cell. Longev. 2020, 2020, 5649174. [Google Scholar] [CrossRef]
- Zhang, J.; Ng, S.; Wang, J.; Zhou, J.; Tan, S.H.; Yang, N.; Lin, Q.; Xia, D.; Shen, H.M. Histone deacetylase inhibitors induce autophagy through FOXO1-dependent pathways. Autophagy 2015, 11, 629–642. [Google Scholar] [CrossRef]
- Gilardini Montani, M.S.; Granato, M.; Santoni, C.; Del Porto, P.; Merendino, N.; D’Orazi, G.; Faggioni, A.; Cirone, M. Histone deacetylase inhibitors VPA and TSA induce apoptosis and autophagy in pancreatic cancer cells. Cell Oncol. 2017, 40, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.H.; Cao, Y.M.; Rong, Z.P.; Ding, J.; Pan, X. Trichostatin A Induces Autophagy in Cervical Cancer Cells by Regulating the PRMT5-STC1-TRPV6-JNK Pathway. Pharmacology 2021, 106, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Jing, K.; Song, K.S.; Shin, S.; Kim, N.; Jeong, S.; Oh, H.R.; Park, J.H.; Seo, K.S.; Heo, J.Y.; Han, J.; et al. Docosahexaenoic acid induces autophagy through p53/AMPK/mTOR signaling and promotes apoptosis in human cancer cells harboring wild-type p53. Autophagy 2011, 7, 1348–1358. [Google Scholar] [CrossRef]
- Shin, S.; Jing, K.; Jeong, S.; Kim, N.; Song, K.S.; Heo, J.Y.; Park, J.H.; Seo, K.S.; Han, J.; Park, J.I.; et al. The omega-3 polyunsaturated fatty acid DHA induces simultaneous apoptosis and autophagy via mitochondrial ROS-mediated Akt-mTOR signaling in prostate cancer cells expressing mutant p53. Biomed. Res. Int. 2013, 2013, 568671. [Google Scholar] [CrossRef]
- Kim, N.; Jeong, S.; Jing, K.; Shin, S.; Kim, S.; Heo, J.Y.; Kweon, G.R.; Park, S.K.; Wu, T.; Park, J.I.; et al. Docosahexaenoic Acid Induces Cell Death in Human Non-Small Cell Lung Cancer Cells by Repressing mTOR via AMPK Activation and PI3K/Akt Inhibition. Biomed. Res. Int. 2015, 2015, 239764. [Google Scholar] [CrossRef]
- Farkas, T.; Daugaard, M.; Jaattela, M. Identification of small molecule inhibitors of phosphatidylinositol 3-kinase and autophagy. J. Biol. Chem. 2011, 286, 38904–38912. [Google Scholar] [CrossRef]
- Liu, Y.; Jiang, N.; Wu, J.; Dai, W.; Rosenblum, J.S. Polo-like kinases inhibited by wortmannin. Labeling site and downstream effects. J. Biol. Chem. 2007, 282, 2505–2511. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Rao, S.; Tokuno, O.; Yamamoto, K.; Takata, M.; Takeda, S.; Utsumi, H. DNA-PK: The major target for wortmannin-mediated radiosensitization by the inhibition of DSB repair via NHEJ pathway. J. Radiat. Res. 2003, 44, 151–159. [Google Scholar] [CrossRef][Green Version]
- Sarkaria, J.N.; Tibbetts, R.S.; Busby, E.C.; Kennedy, A.P.; Hill, D.E.; Abraham, R.T. Inhibition of phosphoinositide 3-kinase related kinases by the radiosensitizing agent wortmannin. Cancer Res. 1998, 58, 4375–4382. [Google Scholar] [PubMed]
- Bain, J.; Plater, L.; Elliott, M.; Shpiro, N.; Hastie, C.J.; McLauchlan, H.; Klevernic, I.; Arthur, J.S.; Alessi, D.R.; Cohen, P. The selectivity of protein kinase inhibitors: A further update. Biochem. J. 2007, 408, 297–315. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.T.; Tan, H.L.; Shui, G.; Bauvy, C.; Huang, Q.; Wenk, M.R.; Ong, C.N.; Codogno, P.; Shen, H.M. Dual role of 3-methyladenine in modulation of autophagy via different temporal patterns of inhibition on class I and III phosphoinositide 3-kinase. J. Biol. Chem. 2010, 285, 10850–10861. [Google Scholar] [CrossRef] [PubMed]
- Klionsky, D.J.; Cuervo, A.M.; Seglen, P.O. Methods for monitoring autophagy from yeast to human. Autophagy 2007, 3, 181–206. [Google Scholar] [CrossRef]
- Karve, S.; Werner, M.E.; Sukumar, R.; Cummings, N.D.; Copp, J.A.; Wang, E.C.; Li, C.; Sethi, M.; Chen, R.C.; Pacold, M.E.; et al. Revival of the abandoned therapeutic wortmannin by nanoparticle drug delivery. Proc. Natl. Acad. Sci. USA 2012, 109, 8230–8235. [Google Scholar] [CrossRef]
- Gu, W.; Lin, Y.; Gou, X.; He, W. Tea Polyphenol inhibits autophagy to sensitize Epirubicin-induced apoptosis in human bladder cancer cells. Neoplasma 2017, 64, 674–680. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Li, H.; Qaed, E.; Zhang, J.; Song, Y.; Wu, R.; Bu, X.; Wang, Q.; Tang, Z. Salinomycin, as an autophagy modulator—A new avenue to anticancer: A review. J. Exp. Clin. Cancer Res. 2018, 37, 26. [Google Scholar] [CrossRef]
- Zhou, J.; Li, G.; Zheng, Y.; Shen, H.M.; Hu, X.; Ming, Q.L.; Huang, C.; Li, P.; Gao, N. A novel autophagy/mitophagy inhibitor liensinine sensitizes breast cancer cells to chemotherapy through DNM1L-mediated mitochondrial fission. Autophagy 2015, 11, 1259–1279. [Google Scholar] [CrossRef]
- Liu, C.M.; Wu, Z.; Pan, B.; An, L.; Zhu, C.; Zhou, J.; Jiang, Y. The antiandrogenic effect of neferine, liensinine, and isoliensinine by inhibiting 5-alpha-reductase and androgen receptor expression via PI3K/AKT signaling pathway in prostate cancer. Pharmazie 2021, 76, 225–231. [Google Scholar] [PubMed]
- Al-Bari, M.A. Chloroquine analogues in drug discovery: New directions of uses, mechanisms of actions and toxic manifestations from malaria to multifarious diseases. J. Antimicrob. Chemother. 2015, 70, 1608–1621. [Google Scholar] [CrossRef]
- Collins, M.P.; Forgac, M. Regulation and function of V-ATPases in physiology and disease. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183341. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, B.K.; Pennypacker, J.K. Drugs that modulate aging: The promising yet difficult path ahead. Transl. Res. 2014, 163, 456–465. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, W.; Gao, Q.; Yang, J.; Yan, X.; Zhao, H.; Su, L.; Yang, M.; Gao, C.; Yao, Y.; et al. Rapamycin directly activates lysosomal mucolipin TRP channels independent of mTOR. PLoS Biol. 2019, 17, e3000252. [Google Scholar] [CrossRef]
- Kim, Y.C.; Guan, K.L. mTOR: A pharmacologic target for autophagy regulation. J. Clin. Investig. 2015, 125, 25–32. [Google Scholar] [CrossRef]
- Yessenkyzy, A.; Saliev, T.; Zhanaliyeva, M.; Masoud, A.R.; Umbayev, B.; Sergazy, S.; Krivykh, E.; Gulyayev, A.; Nurgozhin, T. Polyphenols as Caloric-Restriction Mimetics and Autophagy Inducers in Aging Research. Nutrients 2020, 12, 1344. [Google Scholar] [CrossRef]
- Messeha, S.S.; Zarmouh, N.O.; Soliman, K.F.A. Polyphenols Modulating Effects of PD-L1/PD-1 Checkpoint and EMT-Mediated PD-L1 Overexpression in Breast Cancer. Nutrients 2021, 13, 1718. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, S.; Xu, S.; Nabavi, S.M.; Amirkhani, M.A.; Sureda, A.; Tejada, S.; Lorigooini, Z. Epigenetic targeting of cancer stem cells by polyphenols (cancer stem cells targeting). Phytother. Res. 2021, 35, 3649–3664. [Google Scholar] [CrossRef]
- Musial, C.; Siedlecka-Kroplewska, K.; Kmiec, Z.; Gorska-Ponikowska, M. Modulation of Autophagy in Cancer Cells by Dietary Polyphenols. Antioxidants 2021, 10, 123. [Google Scholar] [CrossRef]
- Wang, K.; Liu, R.; Li, J.; Mao, J.; Lei, Y.; Wu, J.; Zeng, J.; Zhang, T.; Wu, H.; Chen, L.; et al. Quercetin induces protective autophagy in gastric cancer cells: Involvement of Akt-mTOR- and hypoxia-induced factor 1alpha-mediated signaling. Autophagy 2011, 7, 966–978. [Google Scholar] [CrossRef] [PubMed]
- Enogieru, A.B.; Haylett, W.; Hiss, D.C.; Bardien, S.; Ekpo, O.E. Rutin as a Potent Antioxidant: Implications for Neurodegenerative Disorders. Oxid. Med. Cell. Longev. 2018, 2018, 6241017. [Google Scholar] [CrossRef]
- Shao, J.; Wang, C.; Li, L.; Liang, H.; Dai, J.; Ling, X.; Tang, H. Luteoloside Inhibits Proliferation and Promotes Intrinsic and Extrinsic Pathway-Mediated Apoptosis Involving MAPK and mTOR Signaling Pathways in Human Cervical Cancer Cells. Int. J. Mol. Sci. 2018, 19, 1664. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Wang, L.; Yi, T.; Xu, J.; Wang, J.; Qin, J.-J.; Chen, Q.; Yip, K.-M.; Pan, Y.H.; Hong, P.; et al. Synergistic effects of autophagy/mitophagy inhibitors and magnolol promote apoptosis and antitumor efficacy. Acta Pharm. Sin. B 2021, in press. [Google Scholar] [CrossRef]
- Gao, A.M.; Zhang, X.Y.; Hu, J.N.; Ke, Z.P. Apigenin sensitizes hepatocellular carcinoma cells to doxorubic through regulating miR-520b/ATG7 axis. Chem. Biol. Interact. 2018, 280, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Mohan, N.; Banik, N.L.; Ray, S.K. Combination of N-(4-hydroxyphenyl) retinamide and apigenin suppressed starvation-induced autophagy and promoted apoptosis in malignant neuroblastoma cells. Neurosci. Lett. 2011, 502, 24–29. [Google Scholar] [CrossRef][Green Version]
- Zhang, L.; Cheng, X.; Gao, Y.; Zheng, J.; Xu, Q.; Sun, Y.; Guan, H.; Yu, H.; Sun, Z. Apigenin induces autophagic cell death in human papillary thyroid carcinoma BCPAP cells. Food. Funct. 2015, 6, 3464–3472. [Google Scholar] [CrossRef]
- He, M.; Min, J.W.; Kong, W.L.; He, X.H.; Li, J.X.; Peng, B.W. A review on the pharmacological effects of vitexin and isovitexin. Fitoterapia 2016, 115, 74–85. [Google Scholar] [CrossRef]
- Bhardwaj, M.; Cho, H.J.; Paul, S.; Jakhar, R.; Khan, I.; Lee, S.J.; Kim, B.Y.; Krishnan, M.; Khaket, T.P.; Lee, H.G.; et al. Vitexin induces apoptosis by suppressing autophagy in multi-drug resistant colorectal cancer cells. Oncotarget 2018, 9, 3278–3291. [Google Scholar] [CrossRef]
- Landais, E.; Moskal, A.; Mullee, A.; Nicolas, G.; Gunter, M.J.; Huybrechts, I.; Overvad, K.; Roswall, N.; Affret, A.; Fagherazzi, G.; et al. Coffee and Tea Consumption and the Contribution of Their Added Ingredients to Total Energy and Nutrient Intakes in 10 European Countries: Benchmark Data from the Late 1990s. Nutrients 2018, 10, 725. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Song, J.; Zhou, Y.; Cao, L.; Gong, Y.; Wei, Y.; Yang, H.; Tang, L. Methylxanthine derivatives promote autophagy in gastric cancer cells targeting PTEN. Anticancer Drugs 2019, 30, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Saeed, M.; Naveed, M.; BiBi, J.; Ali Kamboh, A.; Phil, L.; Chao, S. Potential nutraceutical and food additive properties and risks of coffee: A comprehensive overview. Crit. Rev. Food Sci. Nutr. 2019, 59, 3293–3319. [Google Scholar] [CrossRef]
- Buldak, R.J.; Hejmo, T.; Osowski, M.; Buldak, L.; Kukla, M.; Polaniak, R.; Birkner, E. The Impact of Coffee and Its Selected Bioactive Compounds on the Development and Progression of Colorectal Cancer In Vivo and In Vitro. Molecules 2018, 23, 3309. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Freedman, N.D.; Haiman, C.A.; Le Marchand, L.; Wilkens, L.R.; Setiawan, V.W. Prospective Study of Coffee Consumption and Cancer Incidence in Non-White Populations. Cancer Epidemiol. Biomark. Prev. 2018, 27, 928–935. [Google Scholar] [CrossRef]
- Pietrocola, F.; Malik, S.A.; Marino, G.; Vacchelli, E.; Senovilla, L.; Chaba, K.; Niso-Santano, M.; Maiuri, M.C.; Madeo, F.; Kroemer, G. Coffee induces autophagy in vivo. Cell Cycle 2014, 13, 1987–1994. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.K.; Xu, S.; Li, S.; Zhang, Y. Proanthocyanidins Should Be a Candidate in the Treatment of Cancer, Cardiovascular Diseases and Lipid Metabolic Disorder. Molecules 2020, 25, 5971. [Google Scholar] [CrossRef]
- Wang, L.; Huang, W.; Zhan, J. Grape Seed Proanthocyanidins Induce Autophagy and Modulate Survivin in HepG2 Cells and Inhibit Xenograft Tumor Growth in Vivo. Nutrients 2019, 11, 2983. [Google Scholar] [CrossRef]
- Ali, A.B.; Nin, D.S.; Tam, J.; Khan, M. Role of chaperone mediated autophagy (CMA) in the degradation of misfolded N-CoR protein in non-small cell lung cancer (NSCLC) cells. PLoS ONE 2011, 6, e25268. [Google Scholar] [CrossRef]
- Moskot, M.; Montefusco, S.; Jakobkiewicz-Banecka, J.; Mozolewski, P.; Wegrzyn, A.; Di Bernardo, D.; Wegrzyn, G.; Medina, D.L.; Ballabio, A.; Gabig-Ciminska, M. The phytoestrogen genistein modulates lysosomal metabolism and transcription factor EB (TFEB) activation. J. Biol. Chem. 2014, 289, 17054–17069. [Google Scholar] [CrossRef]
- Argüello, G.; Balboa, E.; Tapia, P.J.; Castro, J.; Yañez, M.J.; Mattar, P.; Pulgar, R.; Zanlungo, S. Genistein Activates Transcription Factor EB and Corrects Niemann-Pick C Phenotype. Int. J. Mol. Sci. 2021, 22, 4220. [Google Scholar] [CrossRef]
- Astanina, E.; Bussolino, F.; Doronzo, G. Multifaceted activities of transcription factor EB in cancer onset and progression. Mol. Oncol. 2021, 15, 327–346. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, J.; Xu, J.; Lu, Y.; Jiang, J.; Wang, L.; Shen, H.M.; Xia, D. Curcumin targets the TFEB-lysosome pathway for induction of autophagy. Oncotarget 2016, 7, 75659–75671. [Google Scholar] [CrossRef]
- Qu, W.; Xiao, J.; Zhang, H.; Chen, Q.; Wang, Z.; Shi, H.; Gong, L.; Chen, J.; Liu, Y.; Cao, R.; et al. B19, a novel monocarbonyl analogue of curcumin, induces human ovarian cancer cell apoptosis via activation of endoplasmic reticulum stress and the autophagy signaling pathway. Int. J. Biol. Sci. 2013, 9, 766–777. [Google Scholar] [CrossRef]
- Wu, J.C.; Lai, C.S.; Badmaev, V.; Nagabhushanam, K.; Ho, C.T.; Pan, M.H. Tetrahydrocurcumin, a major metabolite of curcumin, induced autophagic cell death through coordinative modulation of PI3K/Akt-mTOR and MAPK signaling pathways in human leukemia HL-60 cells. Mol. Nutr. Food Res. 2011, 55, 1646–1654. [Google Scholar] [CrossRef]
- Zhou, T.; Ye, L.; Bai, Y.; Sun, A.; Cox, B.; Liu, D.; Li, Y.; Liotta, D.; Snyder, J.P.; Fu, H.; et al. Autophagy and apoptosis in hepatocellular carcinoma induced by EF25-(GSH)2: A novel curcumin analog. PLoS ONE 2014, 9, e107876. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Takeda, T.; Tsuiji, K.; Wong, T.F.; Tadakawa, M.; Kondo, A.; Nagase, S.; Yaegashi, N. Curcumin induces cross-regulation between autophagy and apoptosis in uterine leiomyosarcoma cells. Int. J. Gynecol. Cancer 2013, 23, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Jiang, T.; Long, M.; Chen, J.; Ren, D.M.; Wong, P.K.; Chapman, E.; Zhou, B.; Zhang, D.D. A Curcumin Derivative That Inhibits Vinyl Carbamate-Induced Lung Carcinogenesis via Activation of the Nrf2 Protective Response. Antioxid. Redox Signal. 2015, 23, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Yue, Y.; Chen, L.; Xu, C.; Wang, Y.; Du, L.; Xue, X.; Liu, Q.; Wang, Y.; Fan, F. Resveratrol Sensitizes Carfilzomib-Induced Apoptosis via Promoting Oxidative Stress in Multiple Myeloma Cells. Front. Pharmacol. 2018, 9, 334. [Google Scholar] [CrossRef]
- Cuanalo-Contreras, K.; Moreno-Gonzalez, I. Natural Products as Modulators of the Proteostasis Machinery: Implications in Neurodegenerative Diseases. Int. J. Mol. Sci. 2019, 20, 4666. [Google Scholar] [CrossRef]
- Mani, R.; Natesan, V. Chrysin: Sources, beneficial pharmacological activities, and molecular mechanism of action. Phytochemistry 2018, 145, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Sundarraj, K.; Raghunath, A.; Panneerselvam, L.; Perumal, E. Fisetin Inhibits Autophagy in HepG2 Cells via PI3K/Akt/mTOR and AMPK Pathway. Nutr. Cancer 2020, 1–13. [Google Scholar] [CrossRef]
- Kumar, D.; Shankar, S.; Srivastava, R.K. Rottlerin-induced autophagy leads to the apoptosis in breast cancer stem cells: Molecular mechanisms. Mol. Cancer 2013, 12, 171. [Google Scholar] [CrossRef]
- Raha, S.; Kim, S.M.; Lee, H.J.; Yumnam, S.; Saralamma, V.V.; Ha, S.E.; Lee, W.S.; Kim, G.S. Naringin Induces Lysosomal Permeabilization and Autophagy Cell Death in AGS Gastric Cancer Cells. Am. J. Chin. Med. 2020, 48, 679–702. [Google Scholar] [CrossRef]
- Zhao, Y.; Fan, D.; Ru, B.; Cheng, K.W.; Hu, S.; Zhang, J.; Li, E.T.; Wang, M. 6-C-(E-phenylethenyl)naringenin induces cell growth inhibition and cytoprotective autophagy in colon cancer cells. Eur. J. Cancer 2016, 68, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.; Rao, X.; Jiang, Q. Gamma-tocotrienol profoundly alters sphingolipids in cancer cells by inhibition of dihydroceramide desaturase and possibly activation of sphingolipid hydrolysis during prolonged treatment. J. Nutr. Biochem. 2017, 46, 49–56. [Google Scholar] [CrossRef]
- Tran, A.T.; Ramalinga, M.; Kedir, H.; Clarke, R.; Kumar, D. Autophagy inhibitor 3-methyladenine potentiates apoptosis induced by dietary tocotrienols in breast cancer cells. Eur. J. Nutr. 2015, 54, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Yami, A.; Hamzeloo-Moghadam, M.; Darbandi, A.; Karami, A.; Mashati, P.; Takhviji, V.; Gharehbaghian, A. Ergolide, a potent sesquiterpene lactone induces cell cycle arrest along with ROS-dependent apoptosis and potentiates vincristine cytotoxicity in ALL cell lines. J. Ethnopharmacol. 2020, 253, 112504. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Han, X. Anthecotulide Sesquiterpene Lactone Exhibits Selective Anticancer Effects in Human Malignant Melanoma Cells by Activating Apoptotic and Autophagic Pathways, S-Phase Cell Cycle Arrest, Caspase Activation, and Inhibition of NF-kappaB Signalling Pathway. Med. Sci. Monit. 2019, 25, 2852–2858. [Google Scholar] [CrossRef]
- Tian, L.; Cheng, F.; Wang, L.; Qin, W.; Zou, K.; Chen, J. CLE-10 from Carpesium abrotanoides L. Suppresses the Growth of Human Breast Cancer Cells (MDA-MB-231) In Vitro by Inducing Apoptosis and Pro-Death Autophagy Via the PI3K/Akt/mTOR Signaling Pathway. Molecules 2019, 24, 1091. [Google Scholar] [CrossRef]
- Bai, M.; Chen, J.J.; Xu, W.; Dong, S.H.; Liu, Q.B.; Lin, B.; Huang, X.X.; Yao, G.D.; Song, S.J. Elephantopinolide A-P, germacrane-type sesquiterpene lactones from Elephantopus scaber induce apoptosis, autophagy and G2/M phase arrest in hepatocellular carcinoma cells. Eur. J. Med. Chem. 2020, 198, 112362. [Google Scholar] [CrossRef]
- Wang, B.; Zhou, T.Y.; Nie, C.H.; Wan, D.L.; Zheng, S.S. Bigelovin, a sesquiterpene lactone, suppresses tumor growth through inducing apoptosis and autophagy via the inhibition of mTOR pathway regulated by ROS generation in liver cancer. Biochem. Biophys. Res. Commun. 2018, 499, 156–163. [Google Scholar] [CrossRef]
- Lindner, P.; Christensen, S.B.; Nissen, P.; Moller, J.V.; Engedal, N. Cell death induced by the ER stressor thapsigargin involves death receptor 5, a non-autophagic function of MAP1LC3B, and distinct contributions from unfolded protein response components. Cell. Commun. Signal. 2020, 18, 12. [Google Scholar] [CrossRef]
- Du, J.; Dong, Z.; Tan, L.; Tan, M.; Zhang, F.; Zhang, K.; Pan, G.; Li, C.; Shi, S.; Zhang, Y.; et al. Tubeimoside I Inhibits Cell Proliferation and Induces a Partly Disrupted and Cytoprotective Autophagy Through Rapidly Hyperactivation of MEK1/2-ERK1/2 Cascade via Promoting PTP1B in Melanoma. Front. Cell Dev. Biol. 2020, 8, 607757. [Google Scholar] [CrossRef]
- Wang, K.; Zhan, Y.; Chen, B.; Lu, Y.; Yin, T.; Zhou, S.; Zhang, W.; Liu, X.; Du, B.; Wei, X.; et al. Tubeimoside I-induced lung cancer cell death and the underlying crosstalk between lysosomes and mitochondria. Cell Death Dis. 2020, 11, 708. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.D.; Mishra, S.; Verma, S.S.; Shekher, A.; Rai, V.; Awasthee, N.; Das, T.J.; Paul, D.; Das, S.K.; Tag, H.; et al. Evaluation of antioxidant, anti-inflammatory and anticancer activities of diosgenin enriched Paris polyphylla rhizome extract of Indian Himalayan landraces. J. Ethnopharmacol. 2021, 270, 113842. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Liu, P.; Wang, N.; Wang, S.; Yang, B.; Li, M.; Chen, J.; Situ, H.; Xie, M.; Lin, Y.; et al. Betulinic Acid Suppresses Breast Cancer Metastasis by Targeting GRP78-Mediated Glycolysis and ER Stress Apoptotic Pathway. Oxid. Med. Cell. Longev. 2019, 2019, 8781690. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.H.; Mun, J.G.; Jeon, H.D.; Kee, J.Y.; Hong, S.H. Betulin Inhibits Lung Metastasis by Inducing Cell Cycle Arrest, Autophagy, and Apoptosis of Metastatic Colorectal Cancer Cells. Nutrients 2019, 12, 66. [Google Scholar] [CrossRef]
- Wang, K.; Tu, Y.; Wan, J.B.; Chen, M.; He, C. Synergistic anti-breast cancer effect of pulsatilla saponin D and camptothecin through interrupting autophagic-lysosomal function and promoting p62-mediated ubiquitinated protein aggregation. Carcinogenesis 2020, 41, 804–816. [Google Scholar] [CrossRef]
- Jang, M.; Kim, H.; Park, R.; Jo, D.; Lee, E.J.; Oh, W.K.; Park, J. 2,2′-Methylenebis (6-tert-butyl 4-methylphenol) enhances the antitumor efficacy of belotecan, a derivative of camptothecin, by inducing autophagy. Oncotarget 2017, 8, 115068–115078. [Google Scholar] [CrossRef]
- Ramesh, G.; Das, S.; Bola Sadashiva, S.R. Berberine, a natural alkaloid sensitizes human hepatocarcinoma to ionizing radiation by blocking autophagy and cell cycle arrest resulting in senescence. J. Pharm. Pharmacol. 2020, 72, 1893–1908. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Men, Q.; Wu, G.; Yu, C.; Huang, Z.; Liu, X.; Li, W. Tetrandrine induces autophagy and differentiation by activating ROS and Notch1 signaling in leukemia cells. Oncotarget 2015, 6, 7992–8006. [Google Scholar] [CrossRef]
- Lyu, L.; Hu, Y.; Yin, S.; Wang, L.; Ye, F.; Wang, M.; Zhou, Y.; Ma, W.; Chen, C.; Jiang, Y.; et al. Autophagy inhibition enhances anti-pituitary adenoma effect of tetrandrine. Phytother Res. 2021, 35, 4007–4021. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Su, M.; Xie, F.; Ai, J.; Ren, Y.; Zhang, J.; Guan, R.; He, W.; Gong, Y.; Guo, Y. Tetrandrine blocks autophagic flux and induces apoptosis via energetic impairment in cancer cells. Cell Death Dis. 2014, 5, e1123. [Google Scholar] [CrossRef] [PubMed]
- Sato, E.; Ohta, S.; Kawakami, K.; Ikeda, M.; Takahashi, T.; Kobayashi, S.; Nomura, M. Tetrandrine Increases the Sensitivity of Human Lung Adenocarcinoma PC14 Cells to Gefitinib by Lysosomal Inhibition. Anticancer Res. 2019, 39, 6585–6593. [Google Scholar] [CrossRef]
- Ye, L.Y.; Hu, S.; Xu, H.E.; Xu, R.R.; Kong, H.; Zeng, X.N.; Xie, W.P.; Wang, H. The effect of tetrandrine combined with cisplatin on proliferation and apoptosis of A549/DDP cells and A549 cells. Cancer Cell Int. 2017, 17, 40. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yue, W.; Lang, H.; Ding, X.; Chen, X.; Chen, H. Resuming Sensitivity of Tamoxifen-Resistant Breast Cancer Cells to Tamoxifen by Tetrandrine. Integr. Cancer Ther. 2021, 20, 1534735421996822. [Google Scholar] [CrossRef] [PubMed]
- Nazim, U.M.; Yin, H.; Park, S.Y. Neferine treatment enhances the TRAILinduced apoptosis of human prostate cancer cells via autophagic flux and the JNK pathway. Int. J. Oncol. 2020, 56, 1152–1161. [Google Scholar]
- Pham, D.C.; Chang, Y.C.; Lin, S.R.; Fuh, Y.M.; Tsai, M.J.; Weng, C.F. FAK and S6K1 Inhibitor, Neferine, Dually Induces Autophagy and Apoptosis in Human Neuroblastoma Cells. Molecules 2018, 23, 3110. [Google Scholar] [CrossRef]
- Dasari, S.; Bakthavachalam, V.; Chinnapaka, S.; Venkatesan, R.; Samy, A.; Munirathinam, G. Neferine, an alkaloid from lotus seed embryo targets HeLa and SiHa cervical cancer cells via pro-oxidant anticancer mechanism. Phytother Res. 2020, 34, 2366–2384. [Google Scholar] [CrossRef] [PubMed]
- Kalai Selvi, S.; Vinoth, A.; Varadharajan, T.; Weng, C.F.; Vijaya Padma, V. Neferine augments therapeutic efficacy of cisplatin through ROS- mediated non-canonical autophagy in human lung adenocarcinoma (A549 cells). Food Chem. Toxicol. 2017, 103, 28–40. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, Y.; Miao, C.; Wang, Y.; Xu, Y.; Dong, R.; Zhang, Z.; Griffin, B.B.; Yuan, C.; Yan, S.; et al. Anti-angiogenesis effect of Neferine via regulating autophagy and polarization of tumor-associated macrophages in high-grade serous ovarian carcinoma. Cancer Lett. 2018, 432, 144–155. [Google Scholar] [CrossRef]
- Liou, Y.F.; Chen, P.N.; Chu, S.C.; Kao, S.H.; Chang, Y.Z.; Hsieh, Y.S.; Chang, H.R. Thymoquinone suppresses the proliferation of renal cell carcinoma cells via reactive oxygen species-induced apoptosis and reduces cell stemness. Environ. Toxicol. 2019, 34, 1208–1220. [Google Scholar] [CrossRef]
- Alaufi, O.M.; Noorwali, A.; Zahran, F.; Al-Abd, A.M.; Al-Attas, S. Cytotoxicity of thymoquinone alone or in combination with cisplatin (CDDP) against oral squamous cell carcinoma in vitro. Sci. Rep. 2017, 7, 13131. [Google Scholar] [CrossRef]
- Pazhouhi, M.; Sariri, R.; Rabzia, A.; Khazaei, M. Thymoquinone synergistically potentiates temozolomide cytotoxicity through the inhibition of autophagy in U87MG cell line. Iran. J. Basic. Med. Sci. 2016, 19, 890–898. [Google Scholar]
- Chen, M.C.; Lee, N.H.; Hsu, H.H.; Ho, T.J.; Tu, C.C.; Hsieh, D.J.; Lin, Y.M.; Chen, L.M.; Kuo, W.W.; Huang, C.Y. Thymoquinone induces caspase-independent, autophagic cell death in CPT-11-resistant lovo colon cancer via mitochondrial dysfunction and activation of JNK and p38. J. Agric. Food Chem. 2015, 63, 1540–1546. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, P.; Wang, X.; Wang, L.; Zhu, Y.; Song, Y.; Gao, W. Celastrol mediates autophagy and apoptosis via the ROS/JNK and Akt/mTOR signaling pathways in glioma cells. J. Exp. Clin. Cancer Res. 2019, 38, 184. [Google Scholar] [CrossRef]
- Lee, H.W.; Jang, K.S.; Choi, H.J.; Jo, A.; Cheong, J.H.; Chun, K.H. Celastrol inhibits gastric cancer growth by induction of apoptosis and autophagy. BMB Rep. 2014, 47, 697–702. [Google Scholar] [CrossRef]
- Liu, M.; Fan, Y.; Li, D.; Han, B.; Meng, Y.; Chen, F.; Liu, T.; Song, Z.; Han, Y.; Huang, L.; et al. Ferroptosis inducer erastin sensitizes NSCLC cells to celastrol through activation of the ROS-mitochondrial fission-mitophagy axis. Mol. Oncol. 2021, 15, 2084–2105. [Google Scholar] [CrossRef]
- Xu, S.W.; Law, B.Y.K.; Qu, S.L.Q.; Hamdoun, S.; Chen, J.; Zhang, W.; Guo, J.R.; Wu, A.G.; Mok, S.W.F.; Zhang, D.W.; et al. SERCA and P-glycoprotein inhibition and ATP depletion are necessary for celastrol-induced autophagic cell death and collateral sensitivity in multidrug-resistant tumor cells. Pharmacol. Res. 2020, 153, 104660. [Google Scholar] [CrossRef]
- Dai, C.H.; Zhu, L.R.; Wang, Y.; Tang, X.P.; Du, Y.J.; Chen, Y.C.; Li, J. Celastrol acts synergistically with afatinib to suppress non-small cell lung cancer cell proliferation by inducing paraptosis. J. Cell Physiol. 2021, 236, 4538–4554. [Google Scholar] [CrossRef]
- Zhang, C.J.; Zhu, N.; Long, J.; Wu, H.T.; Wang, Y.X.; Liu, B.Y.; Liao, D.F.; Qin, L. Celastrol induces lipophagy via the LXRalpha/ABCA1 pathway in clear cell renal cell carcinoma. Acta Pharmacol. Sin. 2020, 42, 1472–1485. [Google Scholar] [CrossRef]
- Hu, M.; Luo, Q.; Alitongbieke, G.; Chong, S.; Xu, C.; Xie, L.; Chen, X.; Zhang, D.; Zhou, Y.; Wang, Z.; et al. Celastrol-Induced Nur77 Interaction with TRAF2 Alleviates Inflammation by Promoting Mitochondrial Ubiquitination and Autophagy. Mol. Cell 2017, 66, 141–153.e6. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Huang, X.; Wang, H.; Yang, H. Celastrol Induces Autophagy by Targeting AR/miR-101 in Prostate Cancer Cells. PLoS ONE 2015, 10, e0140745. [Google Scholar] [CrossRef]
- Nazim, U.M.; Yin, H.; Park, S.Y. Autophagy flux inhibition mediated by celastrol sensitized lung cancer cells to TRAILinduced apoptosis via regulation of mitochondrial transmembrane potential and reactive oxygen species. Mol. Med. Rep. 2019, 19, 984–993. [Google Scholar] [PubMed]
- Tan, X.; Zou, L.; Qin, J.; Xia, D.; Zhou, Y.; Jin, G.; Jiang, Z.; Li, H. SQSTM1/p62 is involved in docosahexaenoic acid-induced cellular autophagy in glioblastoma cell lines. In Vitro Cell Dev. Biol. Anim. 2019, 55, 703–712. [Google Scholar] [CrossRef]
- Kim, S.; Jing, K.; Shin, S.; Jeong, S.; Han, S.H.; Oh, H.; Yoo, Y.S.; Han, J.; Jeon, Y.J.; Heo, J.Y.; et al. Omega3-polyunsaturated fatty acids induce cell death through apoptosis and autophagy in glioblastoma cells: In vitro and in vivo. Oncol. Rep. 2018, 39, 239–246. [Google Scholar]
- Tan, R.H.; Wang, F.; Fan, C.L.; Zhang, X.H.; Zhao, J.S.; Zhang, J.J.; Yang, Y.; Xi, Y.; Zou, Z.Q.; Bu, S.Z. Algal oil rich in n-3 polyunsaturated fatty acids suppresses B16F10 melanoma lung metastasis by autophagy induction. Food Funct. 2018, 9, 6179–6186. [Google Scholar] [CrossRef]
- D’Eliseo, D.; Di Renzo, L.; Santoni, A.; Velotti, F. Docosahexaenoic acid (DHA) promotes immunogenic apoptosis in human multiple myeloma cells, induces autophagy and inhibits STAT3 in both tumor and dendritic cells. Genes Cancer 2017, 8, 426–437. [Google Scholar] [CrossRef]
- Giordano, C.; Plastina, P.; Barone, I.; Catalano, S.; Bonofiglio, D. n-3 Polyunsaturated Fatty Acid Amides: New Avenues in the Prevention and Treatment of Breast Cancer. Int. J. Mol. Sci. 2020, 21, 2279. [Google Scholar] [CrossRef]
- Rovito, D.; Giordano, C.; Plastina, P.; Barone, I.; De Amicis, F.; Mauro, L.; Rizza, P.; Lanzino, M.; Catalano, S.; Bonofiglio, D.; et al. Omega-3 DHA- and EPA-dopamine conjugates induce PPARgamma-dependent breast cancer cell death through autophagy and apoptosis. Biochim. Biophys. Acta 2015, 1850, 2185–2195. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Lin, G.; Song, C.; Wu, Y.; Feng, N.; Chen, W.; He, Z.; Chen, Y.Q. RA and omega-3 PUFA co-treatment activates autophagy in cancer cells. Oncotarget 2017, 8, 109135–109150. [Google Scholar] [CrossRef]
- Jeong, S.; Kim, D.Y.; Kang, S.H.; Yun, H.K.; Kim, J.L.; Kim, B.R.; Park, S.H.; Na, Y.J.; Jo, M.J.; Jeong, Y.A.; et al. Docosahexaenoic Acid Enhances Oxaliplatin-Induced Autophagic Cell Death via the ER Stress/Sesn2 Pathway in Colorectal Cancer. Cancers 2019, 11, 982. [Google Scholar] [CrossRef] [PubMed]
- Pizato, N.; Kiffer, L.; Luzete, B.C.; Assumpcao, J.A.F.; Correa, L.H.; Melo, H.A.B.; Sant’Ana, L.P.; Ito, M.K.; Magalhaes, K.G. Omega 3-DHA and Delta-Tocotrienol Modulate Lipid Droplet Biogenesis and Lipophagy in Breast Cancer Cells: The Impact in Cancer Aggressiveness. Nutrients 2019, 11, 1199. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Yao, R.; Zhao, D.; Zhou, L.; Wu, Y.; Yang, Y.; Sun, Y.; Lu, L.; Gao, W. Trichostatin A reverses the chemoresistance of lung cancer with high IGFBP2 expression through enhancing autophagy. Sci. Rep. 2018, 8, 3917. [Google Scholar] [CrossRef] [PubMed]
- Lambert, I.H.; Nielsen, D.; Sturup, S. Impact of the histone deacetylase inhibitor trichostatin A on active uptake, volume-sensitive release of taurine, and cell fate in human ovarian cancer cells. Am. J. Physiol. Cell Physiol. 2020, 318, C581–C597. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Sun, X.; Zhang, Q.; Chen, X.; Zhao, T.; Lu, L.; Zhang, J.; Hong, Y. Histone deacetylase inhibitor trichostatin A and autophagy inhibitor chloroquine synergistically exert anti-tumor activity in H-ras transformed breast epithelial cells. Mol. Med. Rep. 2018, 17, 4345–4350. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Zhu, Y.; Xu, X.; Zhu, Y.; Song, Y.; Pang, L.; Chen, Z. The anti-tumor effects of dual PI3K/mTOR inhibitor BEZ235 and histone deacetylase inhibitor Trichostatin A on inducing autophagy in esophageal squamous cell carcinoma. J. Cancer 2018, 9, 987–997. [Google Scholar] [CrossRef]
- Chen, L.; Jin, T.; Zhu, K.; Piao, Y.; Quan, T.; Quan, C.; Lin, Z. PI3K/mTOR dual inhibitor BEZ235 and histone deacetylase inhibitor Trichostatin A synergistically exert anti-tumor activity in breast cancer. Oncotarget 2017, 8, 11937–11949. [Google Scholar] [CrossRef]
- Nazim, U.M.; Park, S.Y. Attenuation of autophagy flux by 6-shogaol sensitizes human liver cancer cells to TRAIL-induced apoptosis via p53 and ROS. Int. J. Mol. Med. 2019, 43, 701–708. [Google Scholar] [CrossRef]
- Wu, J.J.; Omar, H.A.; Lee, Y.R.; Teng, Y.N.; Chen, P.S.; Chen, Y.C.; Huang, H.S.; Lee, K.H.; Hung, J.H. 6-Shogaol induces cell cycle arrest and apoptosis in human hepatoma cells through pleiotropic mechanisms. Eur. J. Pharmacol. 2015, 762, 449–458. [Google Scholar] [CrossRef]
- Hung, J.Y.; Hsu, Y.L.; Li, C.T.; Ko, Y.C.; Ni, W.C.; Huang, M.S.; Kuo, P.L. 6-Shogaol, an active constituent of dietary ginger, induces autophagy by inhibiting the AKT/mTOR pathway in human non-small cell lung cancer A549 cells. J. Agric. Food Chem. 2009, 57, 9809–9816. [Google Scholar] [CrossRef]
- Li, T.Y.; Chiang, B.H. 6-shogaol induces autophagic cell death then triggered apoptosis in colorectal adenocarcinoma HT-29 cells. Biomed. Pharmacother. 2017, 93, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Pei, X.D.; He, Z.L.; Yao, H.L.; Xiao, J.S.; Li, L.; Gu, J.Z.; Shi, P.Z.; Wang, J.H.; Jiang, L.H. 6-Shogaol from ginger shows anti-tumor effect in cervical carcinoma via PI3K/Akt/mTOR pathway. Eur. J. Nutr. 2021. [Google Scholar] [CrossRef]
- Bawadood, A.S.; Al-Abbasi, F.A.; Anwar, F.; El-Halawany, A.M.; Al-Abd, A.M. 6-Shogaol suppresses the growth of breast cancer cells by inducing apoptosis and suppressing autophagy via targeting notch signaling pathway. Biomed. Pharmacother. 2020, 128, 110302. [Google Scholar] [CrossRef]
- Ray, A.; Vasudevan, S.; Sengupta, S. 6-Shogaol Inhibits Breast Cancer Cells and Stem Cell-Like Spheroids by Modulation of Notch Signaling Pathway and Induction of Autophagic Cell Death. PLoS ONE 2015, 10, e0137614. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, M.; Makuch, S.; Winograd, K.; Wisniewski, J.; Ziolkowski, P.; Agrawal, S. 6-Shogaol enhances the anticancer effect of 5-fluorouracil, oxaliplatin, and irinotecan via increase of apoptosis and autophagy in colon cancer cells in hypoxic/aglycemic conditions. BMC Complement. Med. Ther. 2020, 20, 141. [Google Scholar] [CrossRef]
- Cristofani, R.; Montagnani Marelli, M.; Cicardi, M.E.; Fontana, F.; Marzagalli, M.; Limonta, P.; Poletti, A.; Moretti, R.M. Dual role of autophagy on docetaxel-sensitivity in prostate cancer cells. Cell Death Dis. 2018, 9, 889. [Google Scholar] [CrossRef]
- Eid, N.; Kondo, Y. Parkin in cancer: Mitophagy-related/unrelated tasks. World J. Hepatol. 2017, 9, 349–351. [Google Scholar] [CrossRef] [PubMed]
- Eid, N.; Ito, Y.; Otsuki, Y. Triggering of Parkin Mitochondrial Translocation in Mitophagy: Implications for Liver Diseases. Front. Pharmacol. 2016, 7, 100. [Google Scholar] [CrossRef]
- Eid, N.; Ito, Y.; Otsuki, Y. The autophagic response to alcohol toxicity: The missing layer. J. Hepatol. 2013, 59, 398. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Geng, F.; Cheng, X.; Guo, Q.; Zhong, Y.; Cloughesy, T.F.; Yong, W.H.; Chakravarti, A.; Guo, D. Lipid Droplets Maintain Energy Homeostasis and Glioblastoma Growth via Autophagic Release of Stored Fatty Acids. iScience 2020, 23, 101569. [Google Scholar] [CrossRef]
- Ghirga, F.; Quaglio, D.; Mori, M.; Cammarone, S.; Lazzetti, A.; Goggiamani, A.; Ingallina, C.; Botta, B.; Calcaterra, A. A unique high-diversity natural product collection as a reservoir of new therapeutic leads. Org. Chem. Front. 2021, 8, 996–1025. [Google Scholar] [CrossRef]
- Ito, Y.; Taniguchi, K.; Kuranaga, Y.; Eid, N.; Inomata, Y.; Lee, S.W.; Uchiyama, K. Uptake of MicroRNAs from Exosome-Like Nanovesicles of Edible Plant Juice by Rat Enterocytes. Int. J. Mol. Sci. 2021, 22, 3749. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
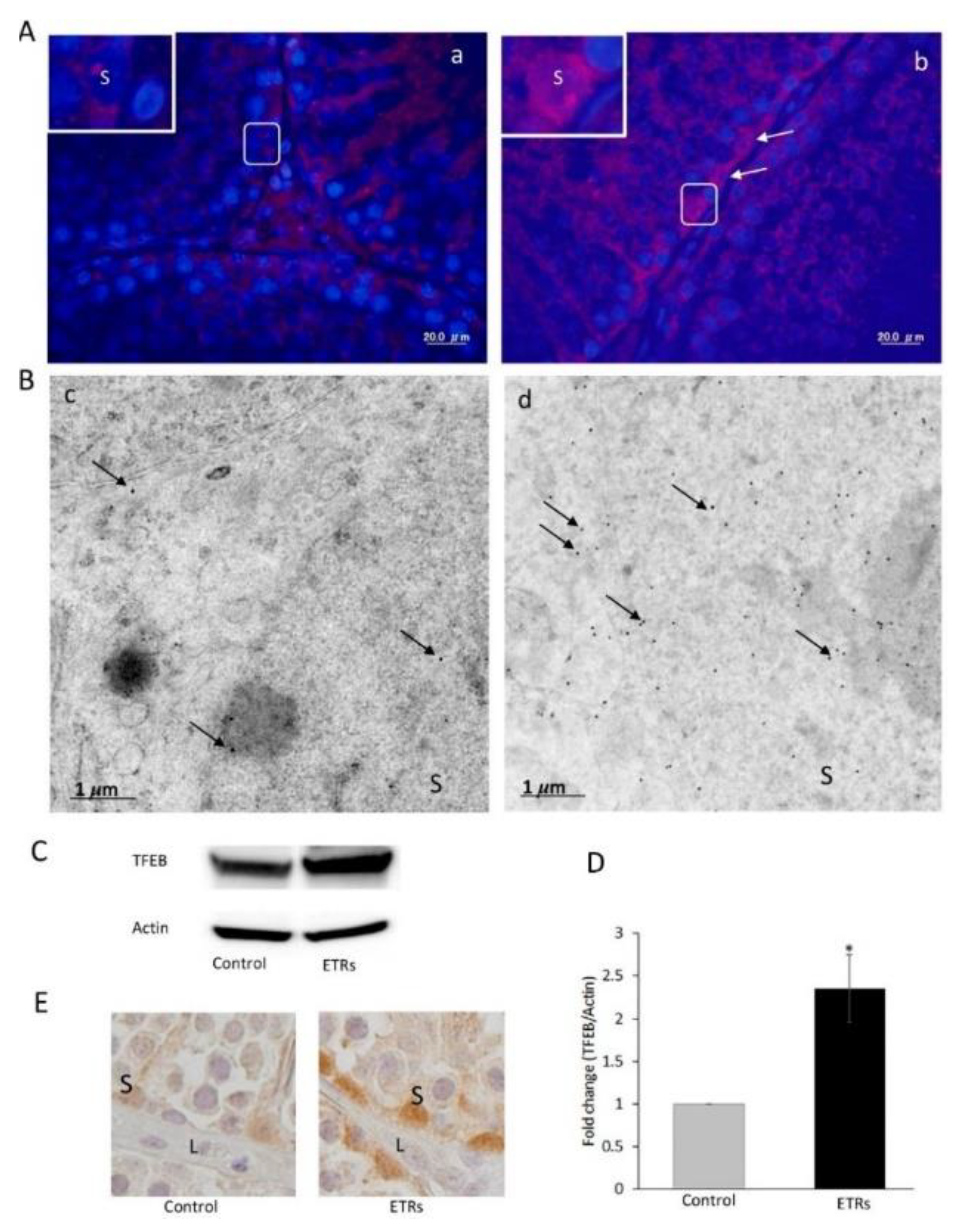{kind=link}
| Target Stage of Autophagy | Natural Product | Chemical Structure | Derivatives | Biological Sources | Autophagy-Related M/A | Traditional Indications | Affected Cancer Cell Type Tested | Remarks | |
|---|---|---|---|---|---|---|---|---|---|
| Autophagy inhibitors: Initiation and nucleation stage inhibitors | |||||||||
| Nucleation | Wortmannin |  | Fungal steroidal furan | Penicillium wortmannin, Talaromyces wortmannin KY12420 | Inhibition of class III PI3K activity | N/A, toxic for prolonged uses | Experimental agent | Nonspecific inhibition both class I and III PI3K [46,47,48,49] | |
| Nucleation | Sonolisib (PX-866) |  | Furan-ring-opened | Wortmannin derivative | Inhibition of class III PI3K activity | - | NSCLC, SCCHN, Glioblastoma | Irreversible, and pan- PI3K inhibitor [50,51,52,53] | |
| Initiation | Cycloheximide |  | Bacterial actidione antibiotic | Streptomyces griseus | ULK1 suppression and activation of mTORC1. | Highly toxic | In vitro research only | Interference translocation step in protein synthesis [54,55] | |
| Nucleation | Petrosaspongiolide M |  | Marine γ-hydroxybutenolide terpenoid | Sponge, Petrosaspongia nigra | Down-regulation of Beclin1 levels | Preclinical studies | U937 cells | Inhibition both of proteasome and autophagy [56,57] | |
| Nucleation | Harmine |  | β-carboline alkaloid | Peganum harmala Lemon balm (Melissa officinalis) | Reduction in LC3-II expression | Anti-viral infection | A wide range of human cancers | Precise inhibitory mechanism of cellular enzymes remains elusive [58] | |
| Elongation and fusion stage inhibitors | |||||||||
| Elongation and fusion | Vinblastine |  | Vinca alkaloids | Catharanthus roseus (L.) or Vinca rosea | Depolymerization of whole MT network | Approved cancers | A wide range of human cancers | No protection the entry of autophagic flux [59,60] | |
| Elongation and fusion | Vincristine |  | Vinca alkaloids | Catharanthus roseus (L.) or Vinca rosea | Depolymerization of whole MT network | Approved cancers | Leukaemia, lymphomas | No protection the entry of autophagic flux [59,60] | |
| Elongation and fusion | Colchicine |  | Colchicum alkaloid | Autumn crocus Colchicum autumnale | Depolarization and interruption of microtubule dynamics | Gout, Behçet’s disease, FMF, swelling | Lung cancer cells | Inducer of ROS mediated autophagy at clinically admissible concentration [59] | |
| Elongation and fusion | Combretastatin |  | Cis-stilbenes (phenols) | South African tree Combretum caffrum (Combretacae) | Inhibition of MT polymerization | - | CT-26, Caco-2 and HT-29 cells | Vascular disrupting agents (VDA) [61,62,63,64] | |
| Elongation | N-Acetyl-L-Cysteine |  | Prodrug of L-cysteine | Onion, Allium cepa | Limiting ROS and blocking ATG4 | Chest pain, ALS, Alzheimer’s disease | A wide range of human cancers | In specific cases, autophagy activator [65,66] | |
| Elongation | Xanthohumol |  | Natural Prenylated chalcone | Hop plant Humulus lupulus L. | Inhibition of VCP function; Up-regulation of p62 and LC3-II | Preclinical cancers | A wide range of human cancers | Activation of ER stress by suppressing of NF-κB, [67] | |
| Elongation | Salvianolic acid B |  | Stilbenoid | Salvia miltiorrhiza (Danshen) | Inhibition of LC3 lipidation | Cardiovascular-related disease, Fibrosis Disease | A wide range of human cancers | Induction of autophagy by suppressing the mTOR pathway [68,69,70,71,72,73] | |
| Elongation | Deguelin |  | Natural rotenoid | African plant, Mundulea sericea (Willd) | Inhibition of LC3 lipidation | Preclinical cancer | A wide range of human cancers | Inhibition of autophagy flux by accumulating p62 [74,75] | |
| Docking and fusion stage inhibitors | |||||||||
| Fusion | Monensin |  | Polyether monocarboxylic acid antibiotic | Streptomyces cinnamonensis | Na+/H+ ionophore | Coccidiosis | A wide range of human cancers | Act as proton exchanger for K+/Na+ [76,77,78] | |
| Fusion and degradation | Ionomycin |  | Natural ionophore | Bacterium Streptomyces conglobatus | Ca2+ ionophore, releases Ca2+ from intracellular stores | Antibiotic | Experimental agent | mTOR-independent autophagy inhibition [76,77,78] | |
| Fusion | L-asparagine |  | Nonessential amino acid | Dietary foods | Inhibition of efflux mechanism of lysosome | Development of brain | A wide range of human cancers | Low-asparagine diet can slow breast cancer metastasis | |
| Fusion and degradation | Liensinine |  | Isoquinoline alkaloid | Seed embryo of Nelumbo nucifera Gaertn | Impairment of RAB7A recruitment to lysosomes | Anti-arrhythmias, anti-hypertension, anti-pulmonary fibrosis | A wide range of human cancers | Unaffected lysosomal pH [79,80,81] | |
| Fusion and degradation | Oblongifolin C |  | Polycyclic polyprenylated acylphloroglucinols | Garcinia yunnanensis Hu | Inhibition of lysosomal proteolytic activity | Antiseptics, antidepressants, and antibiotics | A wide range of human cancers | Guttiferone K is another active component of the plant causes autophagy induction [82,83,84,85,86] | |
| Autolysosomal acidification stageinhibitors | |||||||||
| Fusion and degradation | Azithromycin |  | Macrolide antibiotic erythromycin analogue | Actinomycete, Streptomyces erythreus (Saccharopolyspora erythraea) | Prevention of autophagosomal acidification | Many pharmacological actions including COVID-19 | A wide range of human cancers | Failure to kill intracellular mycobacteria in NTM, Mycobacterium abscessus in CF patients [87] | |
| Fusion and degradation | Clarithromycin |  | Macrolide erythromycin analogue | Actinomycete, Streptomyces erythreus (Saccharopolyspora erythraea) | Prevention of autophagosomal acidification | Antibiotic | A wide range of human cancers | Failure to kill intracellular mycobacteria [88] | |
| Diffusion and degradation | Matrine |  | Quinolizidine alkaloid | Plants Sophora flavescens Ait | Inhibition of endosomal/lysosomal acidification | Anticancer, anti-inflammatory, antiviral actions | A wide range of human cancers | Induction of autophagy by activating the AMPK pathway [89,90,91] | |
| Degradation | Elaiophylin |  | Macrodiolide antibiotic | Streptomyces melanosporus | Disruption of lysosomal degradation | Antibacterial and antihelminthic activities | A wide range of human cancers | Induction of LMP [92,93] | |
| Degradation | Lucanthone |  | Thioxanthenones | Plant Gentiana lutea and mycelium of Aspergillus stellatus derivative | Blocking autophagic flux | Approved for schistosomiasis | A wide range of human cancers | A blocker of DNA base excision repair [94] | |
| Vacuolar H+-ATPase inhibitors | |||||||||
| Fusion and degradation | Bafilomycin A1 |  | 16-membered plecomacrolide | Streptomyces griseus | Lysosomal V-ATPase inhibition | Antibiotic, High toxicity profile | Experimental agent | Universal V-ATPase inhibitor (e.g., osteoclast) [95,96,97] | |
| Fusion and degradation | Concanamycin A |  | 18-membered plecomacrolide | Mycelium Streptomyces diastatochromogenes S-45 | Lysosomal V-ATPase inhibition | Antibiotic, Neoplasm | Experimental agent | Universal V-ATPase inhibitor (e.g., osteoclast) [98] | |
| Fusion and degradation | Manzamine A |  | Manzamine alkaloid | marine sponges of the genera Haliclona sp., Xestospongia sp. and Pellina sp. | Lysosomal V-ATPase inhibition | Several pharmacological actions | A wide range of human cancers | v-ATPase inhibition is similar to bafilomycin A [99,100] | |
| Fusion and degradation | Doxorubicin |  | Anthracycline antibiotic | Streptomyces peucetius, Streptomyces coeruleorubidus | Lysosomal V-ATPase suppression | Antibiotic Antitumor | Approved for range of human cancers | Universal V-ATPase inhibitor [101,102] | |
| Fusion and degradation | Cleistanthin-A |  | Diphyllin glycoside | Tropical plant Cleistanthus collinus | Lysosomal V-ATPase inhibition | Homicide | Experimental agent (In vitro) | Highly toxic and universal V-ATPase inhibitor [103] | |
| Fusion and degradation | Archazolid A |  | Macrocyclic polyketide | Myxobacterium Archangium gephyra, Cystobacter violaceus | Lysosomal V-ATPase inhibition | Antibiotic | Experimental agent (In vitro) | Potent and specific V-ATPase inhibitors [103] | |
| Lysosomal hydrolytic enzyme inhibitors | |||||||||
| Degradation (partial) | Leupeptin |  | Tripeptide antibiotic | Multiple sources | Lysosomal thiolprotease and Ca2+-dependent calpain inhibitor | Serine protease inhibitor | Experimental agent (In vitro) | Agent for analyzing autophagy dynamics in vivo [104] | |
| Degradation (partial) | Pepstatin A |  | Hexapeptide metabolite | Actinomycetes | Lysosomal Aspartyl protease and cathepsin D inhibitor | Aspartyl protease inhibitor | Experimental agent (In vitro) | A reversible nonspecific inhibitor [105] | |
| Autophagy Inducers | |||||||||
| Initiation | Rapamycin (sirolimus) |  | Macrocyclic lactone | Bacterium Streptomyces hygroscopicus | Specific inhibitor of mTOR | Many pharmacological actions | A wide range of human cancers | Suppression of mTORC2 function [106,107,108] | |
| Initiation and nucleation | Quercetin |  | 3, 3′, 4′, 5, 7-pentahydroxyflavone | Many foods, including fruits, vegetables and beverages | Induction of FOXO1 and ATG5 levels; AMPK activator | Dietary supplement; many pharmacological actions | A wide range of human cancers | Well-known antioxidant [109,110] | |
| Initiation and nucleation | Kaempferol |  | Tetrahydroxyflavone | Different in fruits and vegetables such as Cuscuta chinensis and Hypericum perforatum. | Induction of AMPK signaling | Dietary supplement; several pharmacological actions | A wide range of human cancers | Multipotential neuroprotective action in CNS diseases [111] | |
| Initiation | Apigenin |  | 4′,5,7-trihydroxyflavone | Widely distributed in fruits and vegetables | Inhibition of the PI3K/AKT/mTOR pathway in HCC | Dietary supplement; several pharmacological actions | A wide range of human cancers | Inhibition of autophagy flux in in the HEKs and CSCC [112,113] | |
| Initiation | EGCG, catechin and epicatechin |  EGCG | Polyphenols | Coffee plant seeds, Tea leaves of Camellia sinensis | Activation of AMPK | Dietary supplement; several pharmacological actions | A wide range of human cancers | Amelioration of a variety of human diseases including cancers [114,115] | |
| Initiation | Genistein |  | 4′,5,7-trihydroxyisoflavone | Soybean and other legumes, such as Vigna angularis | Inhibition of the PI3K-AKT signaling pathway and N-CoR misfolding; induction of TFEB expression | Dietary supplement; several pharmacological actions | A wide range of human cancers | Beneficial agent in treatment of lysosomal storage diseases [116,117,118] | |
| Initiation; elongation | Curcumin |  | Turmeric polyphenol (beta-diketone) | Turmeric rhizome (Curcuma longa L.), Curcuma zedoaria (Christm). Rosc., and Curcuma petiolata | Increases transcriptional activity of TFEB, which in turn suppresses mTOR and increases LC3 levels | Dietary supplement; many pharmacological actions | Clinical trial for cancers | In specific cases, autophagy inhibition [119,120,121,122,123] | |
| Initiation | Resveratrol |  | 3,5,4′-trihydroxy-trans-stilbene (Stilbenoids) | Fruits such as berries, red grapes and peanuts, Pterocarpus marsupium, Vitis amurensis roots | Caloric restriction mimetic, AMPK activation | Dietary supplement; numerous pharmacological actions | A wide range of human cancers | Attenuation in cigarette smoke-induced autophagy (at higher concentrations) [118,124,125,126] | |
| Nucleation | Chrysin |  | Dihydroxyflavone | Propolis, honey and plants such as Passiflora caerulea, Passiflora incarnate | Decrease in LC3-II, Beclin1 and ATG7 levels | Dietary supplement; numerous pharmacological actions | A wide range of human cancers | A potent inhibitor of aromatase [127] | |
| Initiation | Fisetin |  | 3,7,3’,4’-tetrahydroxyflavone | Rhus verniciflua Stokes, fruits and vegetables (apples, persimmons, grapes, kiwis, strawberries, onions | Activation of AMPK, suppression mTOR activity | Several pharmacological actions | A wide range of human cancers | Inhibition of autophagy in MCF7 breast cancer cells [128,129,130] | |
| Initiation | Cucurbitacin B |  | Tetracyclic triterpene | Cucurbitaceous plants | Induction of ROS formation | Numerous pharmacological actions | A wide range of human cancers | Protection against pressure-overload cardiac hypertrophy by inhibiting the AKT-mTOR pathway [131,132] | |
| Initiation, nucleation | Wogonin |  | 5,7-Dihydroxy-8-methoxyflavone | Scutellaria baicalensis Georgi, Scutellaria radix | ER stress, upregulation of LC3-II and Beclin1 | Numerous pharmacological actions | A wide range of human cancers | Modulation of ER stress and autophagy and/or apoptosis in a cell-type- and context-dependent manner [133,134] | |
| Initiation | Morusin |  | Mulberrochromene, prenylated flavonoid | Root bark of mulberry tree (Morus alba L., Morus nigra L, M. australis) | mTOR1 inhibition, AMPK activation | Antitumor, antioxidant and anti-bacteria property | A wide range of human cancers | Inhibition of NF-κB and STAT3 activity [135,136] | |
| Initiation | Rottlerin (Mallotoxin or Kamala) |  | Polyphenolic ketone | Mallotus philippinensis | Inhibition of mTOR signaling and AMPK induction | Tapeworm, scabies and herpetic ringworm | A wide range of human cancers | Protein kinase C-δ inhibitor [137,138,139] | |
| Nucleation | Paclitaxel |  | Polyoxygenated taxane class diterpenoid | Stem bark of the Pacific yew tree (Taxus brevifolia Nutt), bark of Pacific Taxus chinensis | Increase LC3-II, ATG5 and Beclin1 levels | FDA-approved for metastatic ovarian cancer | Lung-, ovarian- and breast cancers | Inhibition of autophagy in MCF-7 and SK-BR-3 breast cancer cells [60,140,141] | |
| Initiation, nucleation | γ-Tocotrienol |  | Unsaturated tail of tocotrienol | Abundant in rice bran oil, palm oil, and annatto seeds | AMPK activation, increasing LC3-II, ATG5 and Beclin1 levels | Many pharmacological actions including malabsorptive conditions | A wide range of human cancers | Vitamin E deficiency treatment [121,142] | |
| Initiation, nucleation | Ursolic Acid |  | Pentacyclic triterpenoid | Plant, Mirabilis jalapa, apple peels | Increasing LC3-II, ATG5 and Beclin1 levels, inhibition of the mTOR pathway | Numerous pharmacological actions | A wide range of human cancers | mTOR-dependent and independent pathway followed [121,143,144] | |
| Initiation | β-Elemene |  | Sesquiterpene | Rhizoma Curcumae, Curcuma wenyujin | Inhibition of the mTOR pathway | - | KRAS mutant CRC cells | Rho kinase inhibitor [121,145,146] | |
| Initiation | (–)-Guaiol |  | Sesquiterpene alcohol | Guaiacum officinale Guaiacum sanctum | Inhibition of the mTOR pathway | Antibacterial activity Gout | NSCLC cells | Blocking of mTORC2-AKT signaling [147] | |
| Initiation | Thapsigargin |  | Sesquiterpene lactone | Root of umbelliferous plant Thapsia garganica | mTOR-independent Ca2+-dependent pathway | Many pharmacological actions | A wide range of human cancers | Inhibition of autophagosome–lysosome fusion [148] | |
| Initiation nucleation | Tubeimoside-1 |  | Triterpenoid saponin | Bolbostemma paniculatum (Maxim) | AMPK activation, inhibition of the mTOR pathway, increasing LC3-II level | Treatment of snake venoms and inflammation | A wide range of human cancers | [149,150,151] | |
| Nucleation | Polyphyllin D |  | Steroidal saponin | Paris polyphylla | Increasing LC3-II and Beclin1 levels | Fevers, headaches, burns, and wounds, and treatment of snake venom | Human breast cancer cells | [152] | |
| Nucleation | Ophiopogonin B |  | Saponin compound | Radix Ophiopogon japonicus | Increasing LC3-II and ATG5-ATG12 levels | Cardioprotective, diuretic and antibacterial activities | Gastric cancer, human cervical cancer | [153] | |
| Initiation, nucleation | Betulinic acid |  | Lupane-type pentacyclic triterpenoid saponin | White birch bark, Betula alba, Betula pubescens | Inhibition of mTOR path 1way | Numerous pharmacological actions | A wide range of human cancers | Nutraceutical in anxiety and stress [154,155,156] | |
| Initiation | Oleanolic acid |  | Pentacyclic triterpenoid | Olive oil, Phytolacca americana, Syzygium spp, garlic, etc. | ROS generation; AMPK activation and mTOR suppression | Numerous pharmacological actions | A wide range of human cancer cells | [143,144] | |
| Initiation | Camptothecin |  | A quinoline type of alkaloid | Bark, stem and leaves of the Chinese happy tree, Camptotheca acuminate, Chonemorpha fragrans | Phosphorylation of AMPK | Anti-HIV activity | Lung, ovarian, breast, pancreas and stomach cancers | Potent topoisomerase I inhibitor [157,158,159] | |
| Initiation, nucleation | Berberine |  | Isoquinoline alkaloid | Coptis chinensis Franch and Rhizoma coptidis herbs | Activation of Beclin1, inhibition of the mTOR pathway | Several pharmacological actions | A wide range of human cancers | An effective immunomodulator [160,161,162] | |
| Initiation, nucleation | Tetrandrine |  | Bisbenzylisoquinoline alkaloid | Root of Stephania tetrandra S Moore | ROS production, elevation of Beclin1 and LC3-II levels | Many pharmacological actions | A wide range of human cancers | A potent lysosomal inhibitor [163,164,165] | |
| Initiation nucleation | Protopine |  | Benzylisoquinoline alkaloid | Nandina domestica,Fumaria vaillantii | p53 phosphorylation, Increase LC3-II level | Several pharmacological actions | Colon cancer cells | An activator of the p53 pathway [166] | |
| Initiation | Neferine |  | Bisbenzylisoquinoline alkaloid | Green seed embryos of Nelumbo nucifera Gaertn (lotus) | ROS generation, inactivation of th mTOR pathway | Several pharmacological actions | A wide range of human cancers | [167,168,169] | |
| Initiation | Graveoline |  | Alkaloid | Ruta graveolens | ROS generation, elevation of Beclin1 levels | Numerous pharmacological actions | Skin melanoma cells | [170] | |
| Initiation | Peiminine |  | Alkaloid | Fritillaria thunbergii | Dephosphorylation of mTOR and AMPK activation | Numerous pharmacological actions | Colorectal cancer cells | Modulation of metabolic pathways | |
| Initiation, Nucleation | Thymoquinone |  | Edible monoterpene | Black cumin (seed) Nigella sativa L. | Induction of AMPK/mTOR signaling pathway, induction of LC3II expression | Dietary supplement; many pharmacological actions | A wide range of human cancers | Thymoquinone inhibits autophagy in glioblastoma cells [171,172,173,174] | |
| Initiation, Nucleation | Celastrol (triperine) |  | Quinone methide triterpenoid | Tripterygium wilfordii Hook (Thunder of God Vine), Tripterygium regelii (Regel’s threewingnut) | Induction of the ROS/JNK pathway, LC3B-II levels | Numerous pharmacological actions | A wide range of human cancers | Inhibition autophagy in prostate cancer cells [121,175,176,177] | |
| Initiation | Pristimerin (Celastrol methyl ester) |  | Quinone methide triterpenoid | Celastraceae and Hippocrateaceae families, | Induction of the ROS/JNK pathway | Numerous pharmacological actions | A wide range of human cancers | A potent and reversible monoacylglycerol lipase inhibitor [178,179] | |
| Initiation | Plumbagin |  | Naphthoquinone | Plants Plumbago zeylandica L (Chitrak), P. europaea, P. rosea, Juglans regia, J. cinereal, J. nigra | ROS generation, inhibition of the mTOR pathway | Numerous pharmacological actions | A wide range of human cancers | [180,181,182,183] | |
| Nucleation | Emodin |  | 3-methyl-1,6,8-trihydroxyanthraquinone | Chinese herbs such as Rheum palmatum (rhubarb), R. rhabarbarum L | Induction of LC3II expression | Several pharmacological actions including antiviral activity against SARS-CoV-2 | A wide range of human cancers | Attenuation of autophagy in in acute pancreatitis model | |
| Nucleation | Gossypol |  | Phenolic natural product | Cotton seed (Gossypium spp.) Malvaceae family | BH3 mimetic | Male oral contraceptive, antimalarial property | A wide range of human cancers | - | |
| Initiation | Anacardic acid |  | 6-Pentadecyl-salicylic acid | Nutshell of the cashew tree, Anacardium occidentale | Stimulation of ER-stress, repression of AKT signaling | Gastric ulcer, gastritis and gastric cancer | Multiple tumor cells | - | |
| Initiation, Nucleation | Withaferin-A |  | Steroidal lactone | Plant Withania somnifera, Withania somnifera(Indian Winter cherry) | ROS generation, induction of LC3-II | Numerous pharmacological actions including anti-inflammatory and antibacterial properties | A wide range of human cancers | Inhibition of lysosomal activity in breast cancer cells | |
| Initiation | Magnolol |  | 5,5’-diallyi-2,2’-dihydroxybiphenyl | Bark of Magnolia officinalis or M. grandiflora | Down-regulation of the Akt/mTOR pathway | Numerous pharmacological actions including anti-inflammatory effects | A wide range of human cancer cells | Mitochondria-targeted mito-magnolol is more effective than it. | |
| Initiation, nucleation | α-Mangostin |  | Xanthone derivative | Garcinia mangostana Linn (mangosteen fruit) | AMPK activation, induction of LC3-II | Numerous pharmacological actions including anti-aging | A wide range of human cancer cells | - | |
| Initiation | Trichostatin A |  | Natural Hydroxamate | Streptomyces hygroscopius | Inhibition of the mTOR pathway, enhancing the FOXO1-dependent pathway | Numerous pharmacological actions including antibiotic potential | A wide range of human cancer cells | Prolonged exposure (24h) leads to block autophagy [184,185,186] | |
| Initiation | Antroquinonol |  | an enone | Antrodia camphorata | Inhibition of mTOR pathways | Numerous pharmacological actions | Various cancer cells | Inhibition of isoprenyl transferase activity | |
| Initiation | ω-3 fatty acids (EPA, DHA) |  DHA |  EPA | PUFAs | Dietary fats | ROS generation, inhibition of mTOR signaling | Many pharmacological actions | A wide range of human cancer cells | Undergoing clinical trials for ovarian cancer [187,188,189] |
| Transcription and elongation | All-trans retinoic acid |  | Vitamin A metabolite | Dietary fats | RARα activation; enhanced Beclin1 and LC3 II levels | Numerous pharmacological actions | Approved for cancers | In specific cases, autophagy inhibition [187,188,189] | |
| Treatment Strategy | Disease/Conditions | Phase/Status | Identifier | Sponsor | |
|---|---|---|---|---|---|
| Inhibitor | Other Intervention | ||||
| PX-866 | Docetaxel | NSCLC; SCC | 1/2/C | NCT01204099 | Cascadian Therapeutics Inc. |
| - | Advanced Solid Tumors | 1/C | NCT00726583 | Cascadian Therapeutics Inc. | |
| Cetuximab | Colorectal Carcinoma | 1/2/C | NCT01252628 | Cascadian Therapeutics Inc | |
| - | Glioblastoma | 2/C | NCT01259869 | NCIC Clinical Trials Group | |
| Vinblastine | Sirolimus | CNS Tumors | 1/C | NCT01135563 | The Hospital for Sick Children |
| CyP, Cape, Nivolumab | Childhood Lymphoma | 1/2/R | NCT03585465 | Centre Oscar Lambret | |
| Selumetinib Sulfate | Astrocytoma | 3/R | NCT04576117 | National Cancer Institute (NCI) | |
| Bevacizumab | Low Grade Glioma | 2/R | NCT02840409 | The Hospital for Sick Children | |
| Nivolumab | Hodgkin Lymphoma | 2/R | NCT03580408 | Lymphoma Academic Research Org. | |
| Dox, Bleomycin, Nivolumab | Hodgkin Lymphoma | 1/2/R | NCT03033914 | Memorial Sloan Kettering Cancer Center | |
| Dox, Bleomycin, Dacarbazine | Hodgkin Lymphoma | 3/R | NCT03159897 | Fondazione Italiana Linfomi ONLUS | |
| Cisplatin, TMZ | Skin Cancer | 2/C | NCT00885534 | Memorial Sloan Kettering Cancer Center | |
| Vincristine | Cytarabine R-EphB4-HSA | Myelodysplastic neoplasm | 1/R | NCT03519984 | University of Southern California |
| CyP Vinorelbine Actino D | Rhabdomyosarcoma | 2/R | NCT04388839 | H. Lee Moffitt Cancer Center | |
| Venetoclax | B ALL | 1/2/R | NCT03504644 | ECOG-ACRIN Cancer Research Group | |
| Inotuzumab Ozogamicin | Recurrent B ALL | 1/2/R | NCT03851081 | Roswell Park Cancer Institute | |
| TMZ | Ewing Sarcoma | 2/R | NCT03359005 | Peking University People’s Hospital | |
| Camrelizumab, Dacarbazine | Hodgkin Lymphoma | 2/R | NCT04067037 | Henan Cancer Hospital | |
| Carboplatin Irinotecan | Pilocytic Astrocytoma | 1/2/R | NCT01837862 | Julie Krystal | |
| Etoposide, Dox, CyP | Recurrent B ALL | 1/R | NCT03991884 | University of Washington | |
| Pegaspargase, CyP, etoposide | NK/T Cell Lymphoma | 2/R | NCT04484506 | Peking University | |
| Melphalan, etoposide | Retinoblastoma | 2/3/R | NCT04681417 | Institut Curie | |
| Brentuximab, Dox Rituximab | Hodgkin Lymphoma | 2/R | NCT02398240 | Mitchell Cairo | |
| Colchicine | - | Hepatocellular Carcinoma | 2/R | NCT04264260 | Kaohsiung Medical University |
| - | Hepatocellular Carcinoma | 2/C | NCT01935700 | Kaohsiung Medical University | |
| CA-4P | Paclitaxel, Carboplatin | Cancer, Tumor | 2/C | NCT00113438 | Mateon Therapeutics |
| Bevacizumab (Avastin) | Tumors | 1/C | NCT00395434 | Mateon Therapeutics | |
| CA-2P | - | Neoplasm Metastasis | 1/C | NCT00960557 | Mateon Therapeutics |
| Fosbretabulin | - | Head and Neck Cancer | 2/C | NCT00060242 | Case Comprehensive Cancer Center |
| Paclitaxel, Bevacizumab | Tumors | 2/C | NCT00653939 | Mateon Therapeutics | |
| - | Adult Solid Tumor | 1/C | NCT00003698 | University of Glasgow | |
| - | Adult Solid Tumor | 1/C | NCT00003768 | Case Comprehensive Cancer Center | |
| - | Neuroendocrine Tumors | 2/C | NCT02132468 | Mateon Therapeutics | |
| - | Neuroendocrine Tumors | 2/C | NCT02279602 | Mateon Therapeutics | |
| NAC | - | Breast Cancer | 1/C | NCT01878695 | Thomas Jefferson University |
| - | Ovarian Cancer | 1/2/NR | NCT04520139 | University of California, Irvine | |
| Simethicone | Stomach Neoplasms | 4/C | NCT01653171 | Pontificia Universidad Catolica de Chile | |
| - | Bronchial Carcinoma | 4/C | NCT00196885 | German Cancer Research Center | |
| Neurofibromatosis 1 | 2/R | NCT04481048 | Children’s Hospital Med Center, Cincinnati | ||
| Sodium thiosulfate | Esophageal Carcinoma | R | NCT04764643 | Xijing Hospital of Digestive Diseases | |
| Azithromycin | Hydroxychloroquine | Cancer and COVID 19 | 2/R | NCT04341207 | Gustave Roussy, Cancer Campus, Grand Paris |
| - | Adenomatous Polyposis | 4/NR | NCT04454151 | Tel-Aviv Sourasky Medical Center | |
| Cefixime | Cervical Papilloma | U | NCT02830230 | Tata Memorial Hospital | |
| Clarithromycin | Abemaciclib | Neoplasm Metastasis | 1/C | NCT02117648 | Eli Lilly and Company |
| Prasterone | Multiple Myeloma | 2/C | NCT00006219 | Mayo Clinic | |
| Lenalidomide Dexamethasone | Multiple Myeloma | 2/R | NCT04063189 | The First Hospital of Jilin University | |
| Pomalidomide Dexamethasone | Multiple Myeloma | 2/NR | NCT04843579 | Weill Medical College of Cornell University | |
| Pomalidomide Dexamethasone | Multiple Myeloma | 2/NR | NCT04302324 | Weill Medical College of Cornell University | |
| Dexamethasone Lenalidomide | Plasma Cell Myeloma | 2/C | NCT00445692 | Fred Hutchinson Cancer Research Center | |
| Thalidomide dexamethasone | Multiple Myeloma | 2/C | NCT00182663 | Fred Hutchinson Cancer Research Center | |
| Dexamethasone Pomalidomide | Multiple Myeloma | 2/C | NCT01159574 | Weill Medical College of Cornell University | |
| Lenalidomide, dexamethasone | Multiple Myeloma | 3/R | NCT04287660 | First Affiliated Hospital of Soochow Uni | |
| Omeprazole, Amoxicillin, | Gastric MALT Lymphoma | NA/C | NCT00327132 | National Health Research Institutes, Taiwan | |
| Amoxicillin, Metronidazole | B-cell Lymphoma | 2/R | NCT02388581 | National Health Research Institutes, Taiwan | |
| Ciprofloxacin, lansoprazole | CLL | 2/C | NCT01279252 | King’s College Hospital NHS Trust | |
| Amoxicillin, Metronidazole | Lymphoma | 2/C | NCT00002682 | M.D. Anderson Cancer Center | |
| Bismuth subcitrate Amoxicillin | Lymphoma | 2/C | NCT00003151 | EORTC | |
| Lucanthone | TMZ, Radiation | Glioblastoma Multiforme | 2/T | NCT01587144 | Spectrum Pharmaceuticals, Inc |
| Dox | L-DOS47 | Pancreas Cancer | 1/2/R | NCT04203641 | Helix BioPharma Corporation |
| Irinotecan, 5-fluorouracil | Gastric Cancer | 2/R | NCT04358341 | Sixth Affiliated Hospital, Sun Yat-sen Uni | |
| Fludarabine | Ovarian Cancer | 2/R | NCT03335241 | Sun Yat-sen University | |
| CyP | Breast Cancer Patients | - | NCT04654195 | Damanhour University | |
| PD-1 | Bladder Cancer | 2/R | NCT04101812 | Tianjin Medical University Second Hospital | |
| - | Breast Cancer | 2/R | NCT03933319 | Chinese Academy of Medical Sciences | |
| Trastuzumab, paclitaxel | Breast Cancer | 1/2/R | NCT03994107 | Peking Union Medical College | |
| Bortezomib | Serous Carcinoma | 2/R | NCT03509246 | Seoul National University Hospital | |
| Bevacizumab, Pembrolizumab | Ovarian Cancer | 1/R | NCT03596281 | Gustave Roussy, Cancer Campus, Grand Paris | |
| CyP, Atezolizumab | Breast Cancer | 2/R | NCT03164993 | Oslo University Hospital | |
| Rapamycin | - | Breast Cancer | 2/R | NCT02642094 | LuZhe Sun |
| - | Bladder Cancer | 2/R | NCT04375813 | Emtora Biosciences | |
| - | Pancreatic Cancer | 1/2/R | NCT03662412 | Zhejiang University | |
| Nivolumab | Ewing Sarcoma | 1/2/R | NCT03190174 | Sarcoma Oncology Research Center, LLC | |
| - | Refractory Solid Tumors | 4/R | NCT02688881 | Samsung Medical Center | |
| Quercetin | Fisetin, Dasatinib | Frailty, Childhood Cancer | 2/NR | NCT04733534 | St. Jude Children’s Research Hospital |
| - | Squamous Cell Carcinoma | 2/R | NCT03476330 | Children’s Hospital Med Center, Cincinnati | |
| Green tea extract | Prostate Cancer | 1/ANR | NCT01912820 | Jonsson Comprehensive Cancer Center | |
| EGCG | - | Colon Cancer | 1/R | NCT02891538 | University of Texas Health Science Center |
| - | Breast Neoplasms | 2/U | NCT02580279 | Shandong Cancer Hospital and Institute | |
| - | Prostate Cancer | 2/C | NCT00676780 | Louisiana State University Health Sciences | |
| Catechin | - | Adult solid tumor | 1/C | NCT00091325 | University of Arizona |
| EGCG | Clomiphene Citrate, Letrozole | Uterine Fibroids | 1/R | NCT04177693 | Yale University |
| Genistein | Sugar pill | Bladder Cancer | 2/R | NCT01489813 | Emory University |
| Gemcitabine | Breast Cancer | 2/C | NCT00244933 | Barbara Ann Karmanos Cancer Institute | |
| Decitabine | NSCLC | 1/2/C | NCT01628471 | Uman Pharma | |
| - | Breast Cancer | 2/C | NCT00290758 | National Cancer Institute (NCI) | |
| Erlotinib HCl, gemcitabine HCl | Pancreatic Cancer | 2/C | NCT00376948 | Barbara Ann Karmanos Cancer Institute | |
| Curcumin | - | Prostate Cancer | 3/R | NCT03769766 | University of Texas Southwestern Med Center |
| - | Breast Cancer | 1/R | NCT03980509 | Medical University of South Carolina | |
| - | Breast Cancer | R | NCT03865992 | City of Hope Medical Center | |
| - | Head and Neck Cancer | 2/R | NCT04208334 | Phramongkutklao College Hospital | |
| - | Prostate Cancer | NCT02064673 | University of Texas Southwestern Med Center | ||
| Pembrolizumab, Vit D, CyP | Endometrial Cancer | NCT03192059 | University Hospital, Ghent | ||
| Resveratrol | Metformin | PCOS | 2/NR | NCT04867252 | Khyber Medical University Peshawar |
| Simvastatin | PCOS | 4/U | NCT02766803 | Poznan University of Medical Sciences | |
| - | PCOS | U | NCT01720459 | Poznan University of Medical Sciences | |
| Sirolimus | Lymphangioleiomyomatosis | 2/U | NCT03253913 | University of Cincinnati | |
| Fisetin | Dasatinib, Quercetin | Childhood Cancer | 2/NR | NCT04733534 | St. Jude Children’s Research Hospital |
| β-elemene | EGFR-TKIs | NSCLC | 2/U | NCT03123484 | China Medical University |
| - | Astrocytoma, Glioblastoma | 3/R | NCT02629757 | Sun Yat-sen University | |
| Ursolic Acid | Curcumin | Prostate Cancer | 1/NR | NCT04403568 | University of Texas Health Science Center |
| Tocotrienol | Bevacizumab | Ovarian Cancer Recurrent | 3/R | NCT04175470 | Vejle Hospital |
| Paclitaxel | - | Breast Cancer | 2/NR | NCT03959397 | Jilin University |
| Apatinib, Camrelizumab | Advanced Gastric Cancer | 1/2/NR | NCT04286711 | China Medical University | |
| Carboplatin | Prostate Cancer | 1/2/NR | NCT04148885 | Shanghai Jiao Tong University | |
| Pembrolizumab | Breast Cancer | 2/NR | NCT03841747 | Queen Mary University of London | |
| - | Breast Cancer | 2/NR | NCT04194684 | Chinese Academy of Medical Sciences | |
| Pyrotinib | Breast Cancer | 2/NR | NCT04659499 | Peking Union Medical College Hospital | |
| Oxaliplatin, Cape | Gastric Cancer | 1/NR | NCT03977220 | Shanghai Zhongshan Hospital | |
| Mipsagargin | G-202, Tapsigargin pro-drug | Advanced Solid Tumors | 1/C | NCT01056029 | GenSpera, Inc. |
| Betulinic acid | - | Dysplastic Nevus Syndrome | 1/2/S | NCT00346502 | University of Illinois at Chicago |
| * CRLX101 | Enzalutamide | Prostate Neoplasms | 2/R | NCT03531827 | National Cancer Institute (NCI) |
| * EP0057 | Olaparib | Urothelial Carcinoma | 1/2/R | NCT02769962 | National Cancer Institute (NCI) |
| Olaparib tablets | Ovarian Cancer | 2/R | NCT04669002 | Ellipses Pharma | |
| Camptothecin | - | Adult Solid Tumor | 1/C | NCT00059917 | Memorial Sloan Kettering Cancer Center |
| - | Solid Tumor | 1/2/C | NCT00333502 | NewLink Genetics Corporation | |
| Berberine HCl | - | Colorectal Adenomas | 2/3/R | NCT03333265 | xiaohua li |
| - | Colorectal Adenoma | 2/3/C | NCT02226185 | Shanghai Jiao Tong University | |
| Gefitinib | Lung Adenocarcinoma | 2/U | NCT03486496 | Fujian Cancer Hospital | |
| - | Colorectal Adenomas | 2/3/U | NCT03281096 | xiaohua li | |
| Thymoquinone | - | Premalignant Lesion | 2/C | NCT03208790 | Cairo University |
| Metformin | PCOS | 2/3/C | NCT04852510 | Saudi German Hospital—Madinah | |
| R-(–)-gossypol | - | Adult Glioblastoma | 2/C | NCT00540722 | National Cancer Institute (NCI) |
| - | Adrenocortical Carcinoma | 2/C | NCT00848016 | National Cancer Institute (NCI) | |
| TMZ | Brain and CNS Tumors | 1/C | NCT00390403 | Sidney Kimmel Com Cancer Center | |
| Paclitaxel, carboplatin | Follicular Lymphoma | 1/C | NCT00891072 | National Cancer Institute (NCI) | |
| Non-small Cell Lung Cancer | 3/U | NCT01977209 | Third Military Medical University | ||
| Cisplatin, etoposide | Small Cell Lung Cancer | 1/C | NCT00544596 | National Cancer Institute (NCI) | |
| Trichostatin A | - | Hematologic Malignancies | 1/U | NCT03838926 | Vanda Pharmaceuticals |
| Antroquinonol | - | Pancreatic Neoplasm | 1/2/R | NCT03310632 | Golden Biotechnology Corporation |
| - | Non-small Cell Lung Cancer | 1/C | NCT01134016 | Golden Biotechnology Corporation | |
| - | Non-small Cell Lung Cancer | 2/C | NCT02047344 | Golden Biotechnology Corporation | |
| - | Acute Myeloid Leukemia | 2/C | NCT03823352 | Golden Biotechnology Corporation | |
| EPA | AMR101 | Colorectal Adenoma | 1/2/R | NCT04216251 | Massachusetts General Hospital |
| AMR101 (VASCEPA) | Colon Cancer | 2/R | NCT03661047 | Mingyang Song | |
| EPA-FFA | - | Adenomatous Polyposis | 3/R | NCT03806426 | S.L.A. Pharma AG |
| EPA | Tyrosine kinase inhibitor | Chronic Myeloid Leukemia | 1/2/R | NCT04006847 | Milton S. Hershey Medical Center |
| Icosapent Ethyl | - | Liver Metastasis | 3/R | NCT03428477 | Mark A Hull, PhD FRCP |
| DHA | - | Breast Cancer | 2/R | NCT03831178 | AHS Cancer Control Alberta |
| EPA | NSCLC | NR | NCT04175769 | AHS Cancer Control Alberta | |
| EPA | Lung Cancer | 2/3/U | NCT01048970 | National Institute of Cancerología | |
| Paclitaxel | Pancreatic Cancer | 2/U | NCT00024375 | Theradex | |
| Paclitaxel | Prostate Cancer | 2/U | NCT00024414 | Theradex | |
| Paclitaxel | Colorectal Cancer | 2/U | NCT00024401 | Theradex | |
| ATRA | Lupron, 5-Azacitidine | Prostatic Neoplasms | 2/R | NCT03572387 | Icahn School of Medicine at Mount Sinai |
| Anastrozole | Breast Neoplasm | 2/R | NCT04113863 | Mario Negri Inst for Pharm Research | |
| - | Leukemia | R | NCT01064557 | Gruppo Italiano Malattie | |
| VEGFR inhibitor | Adenoid Cystic Carcinoma | 2/R | NCT04433169 | Shanghai Jiao Tong University | |
| Tranylcypromine, cytarabine | Acute Myeloid Leukemia | 1/2/R | NCT02717884 | Michael Luebbert | |
| - | APL | 2/R | NCT01064570 | Gruppo Italiano Malattie | |
| Arsenic Trioxide, Mylotarg | APL | 2/R | NCT04793919 | Associazione Italiana Ematologia | |
| Cytarabine, Arsenic Trioxide | AML | 1/2/R | NCT03031249 | Institute of Hematology and Blood Diseases | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Bari, M.A.A.; Ito, Y.; Ahmed, S.; Radwan, N.; Ahmed, H.S.; Eid, N. Targeting Autophagy with Natural Products as a Potential Therapeutic Approach for Cancer. Int. J. Mol. Sci. 2021, 22, 9807. https://doi.org/10.3390/ijms22189807
Al-Bari MAA, Ito Y, Ahmed S, Radwan N, Ahmed HS, Eid N. Targeting Autophagy with Natural Products as a Potential Therapeutic Approach for Cancer. International Journal of Molecular Sciences. 2021; 22(18):9807. https://doi.org/10.3390/ijms22189807
Chicago/Turabian StyleAl-Bari, Md. Abdul Alim, Yuko Ito, Samrein Ahmed, Nada Radwan, Hend S. Ahmed, and Nabil Eid. 2021. "Targeting Autophagy with Natural Products as a Potential Therapeutic Approach for Cancer" International Journal of Molecular Sciences 22, no. 18: 9807. https://doi.org/10.3390/ijms22189807
APA StyleAl-Bari, M. A. A., Ito, Y., Ahmed, S., Radwan, N., Ahmed, H. S., & Eid, N. (2021). Targeting Autophagy with Natural Products as a Potential Therapeutic Approach for Cancer. International Journal of Molecular Sciences, 22(18), 9807. https://doi.org/10.3390/ijms22189807