
Oxidative Stress, Mutations and Chromosomal Aberrations Induced by In Vitro and In Vivo Exposure to Furan

 ,
, 
Abstract
:1. Introduction
2. Results
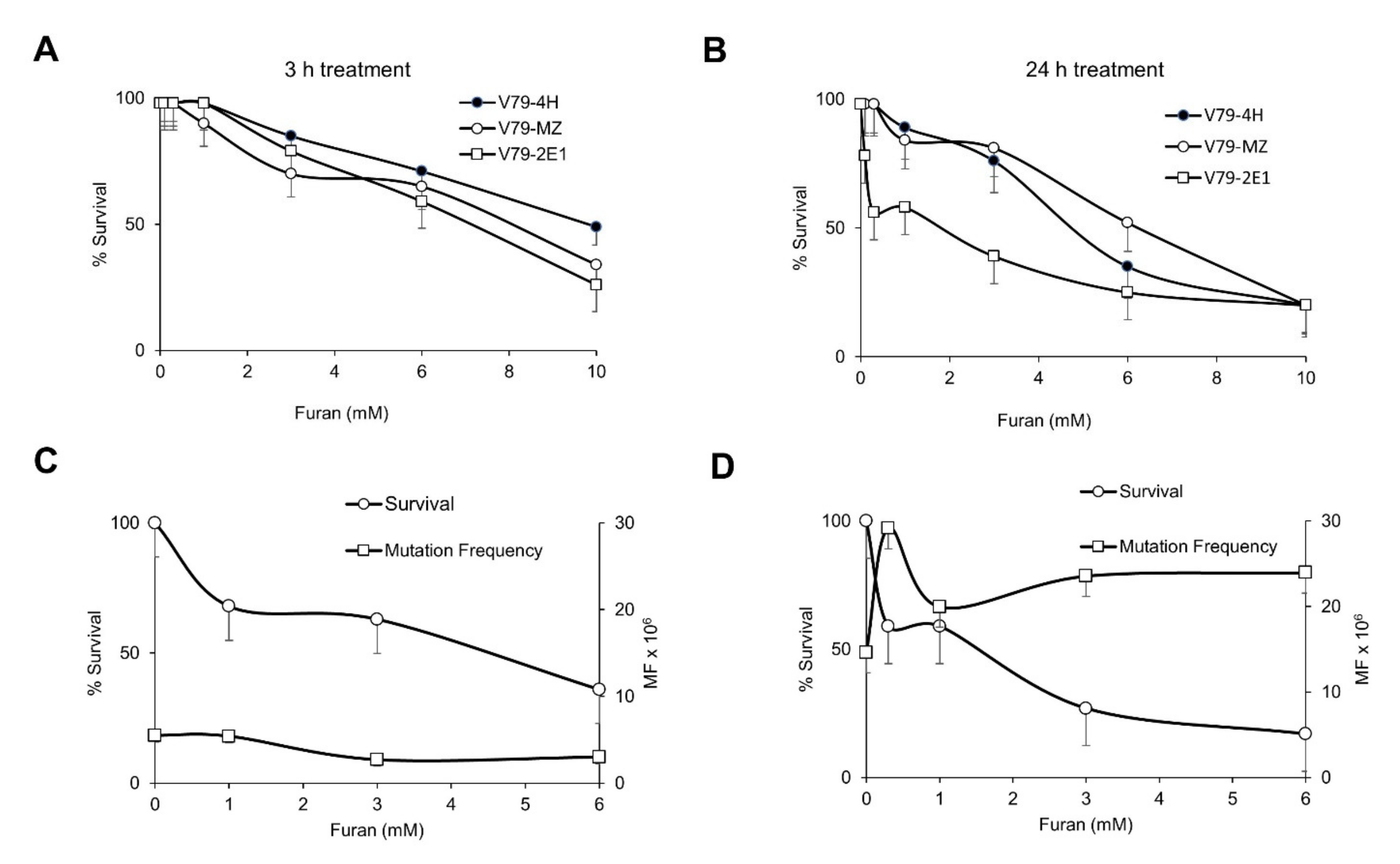
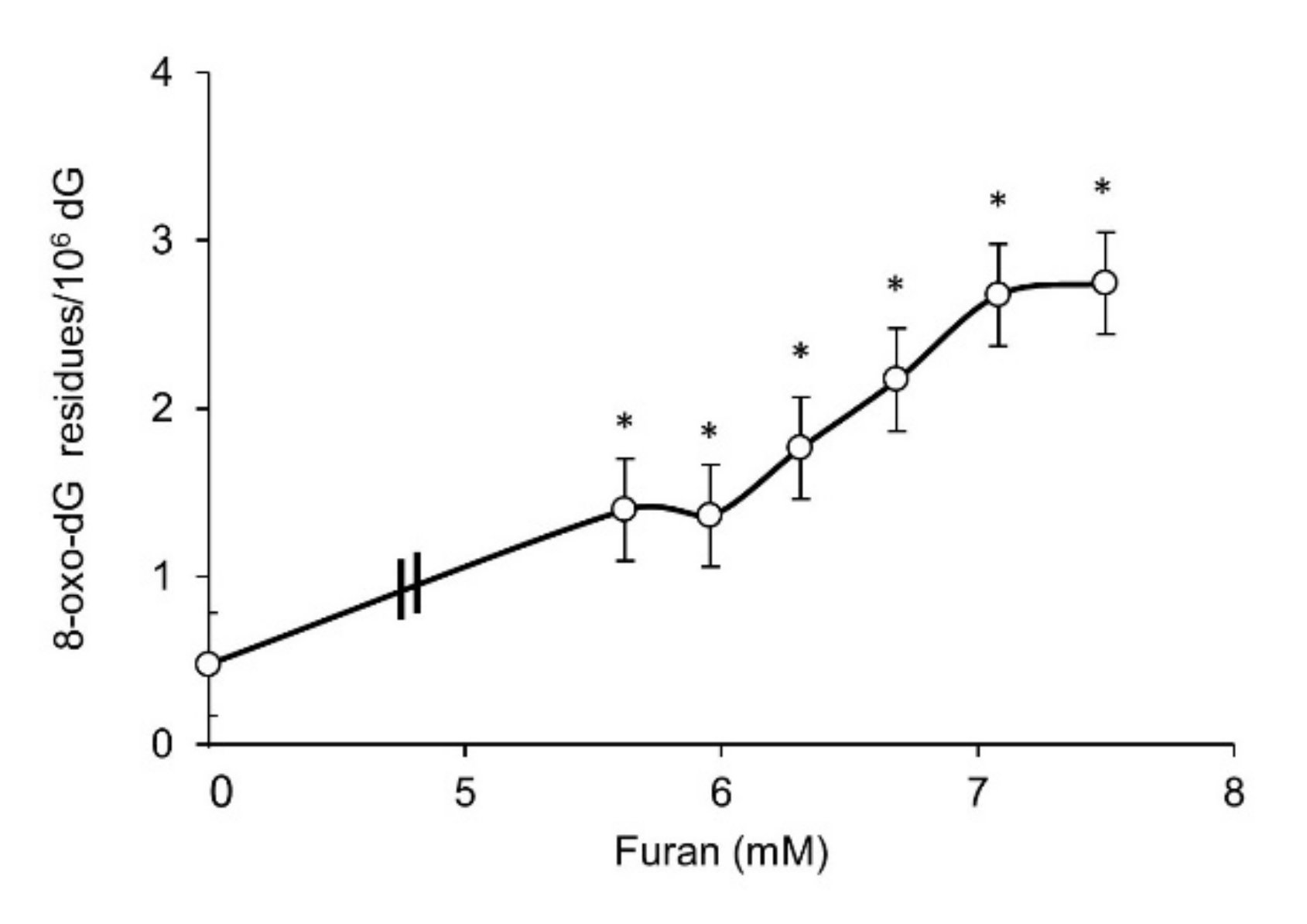
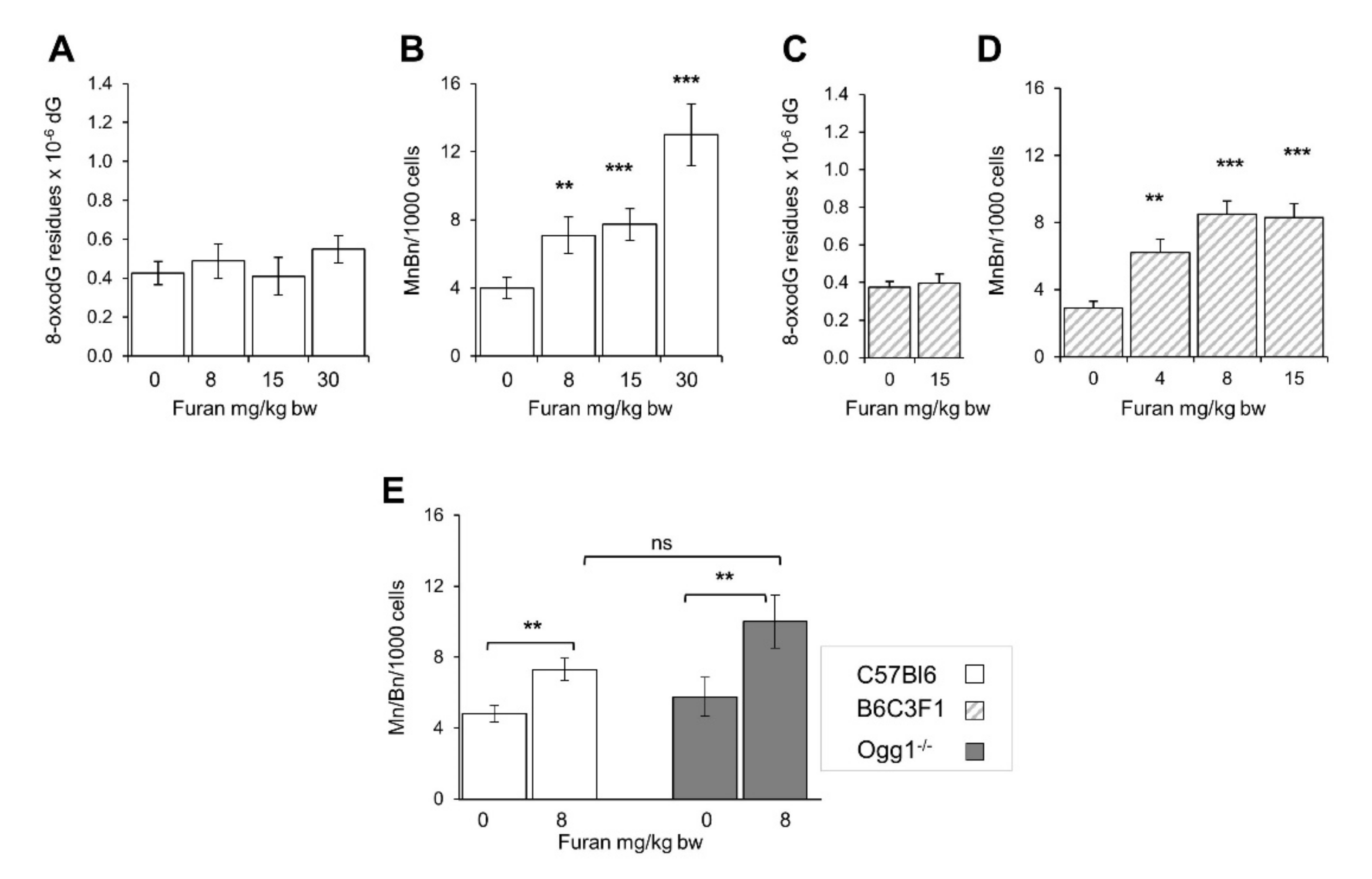
2.1. Furan Is Not Mutagenic at the Hprt and Tk Loci and Is a Strong Inducer of DNA Oxidation in Mammalian Cell Lines
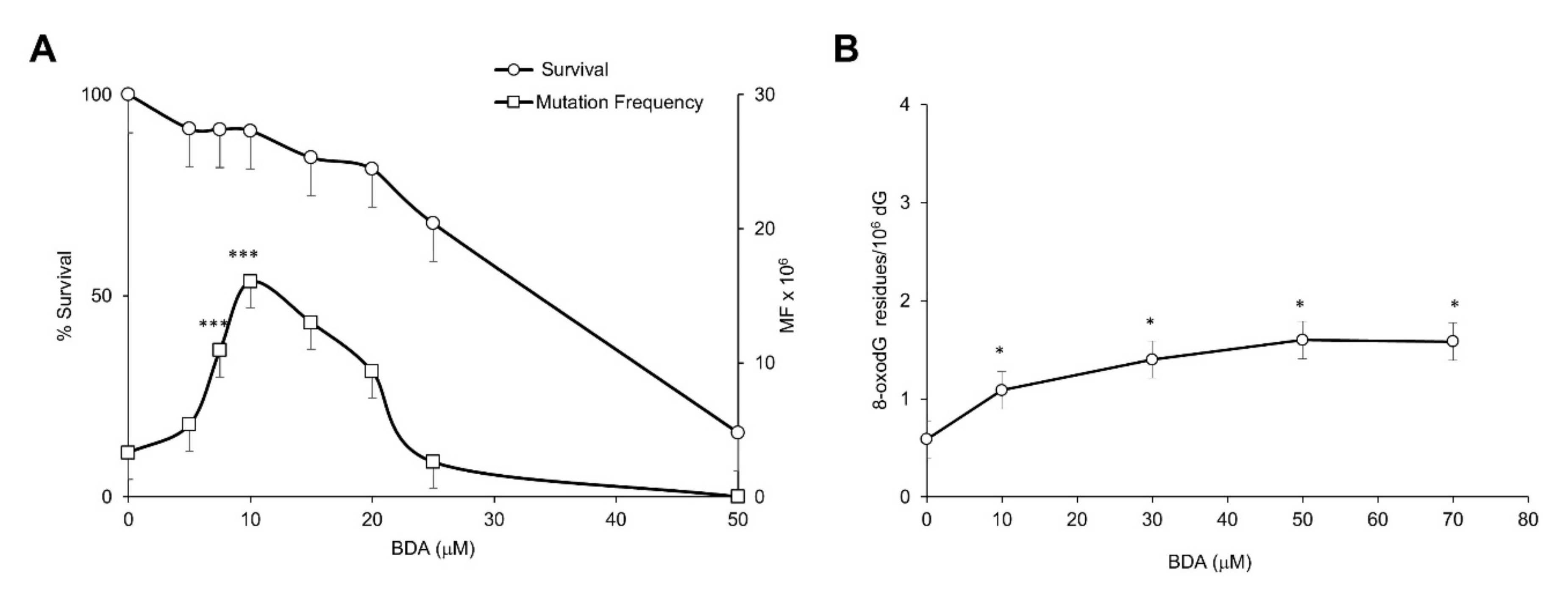
2.2. The Furan Metabolite BDA Is a Weak Mutagen at the Hprt and Tk Loci and a Poor Inducer of DNA Oxidation in Mammalian Cell Lines
2.3. BDA Induces Micronuclei in L5178Y Lymphoblastoid Cells
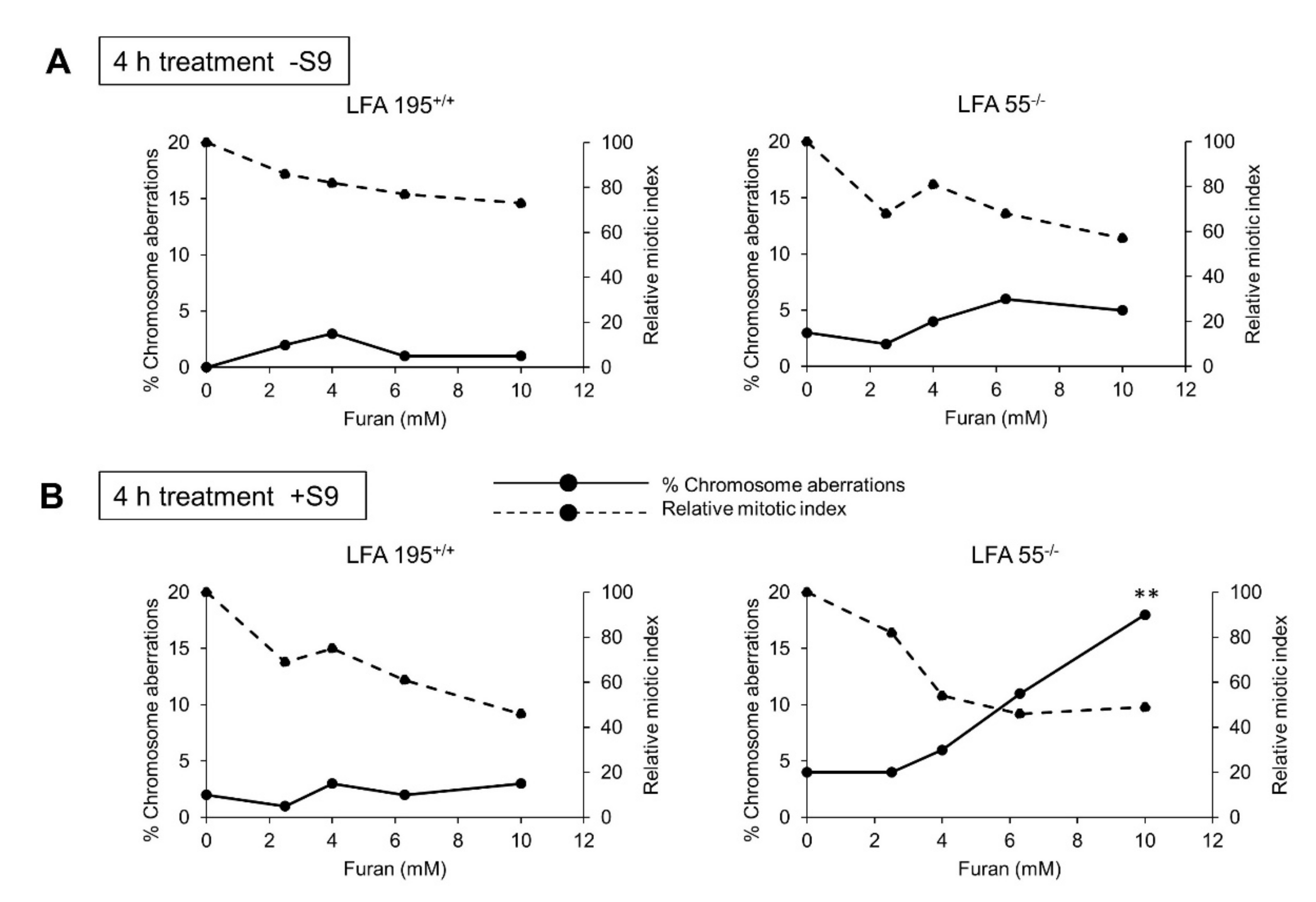
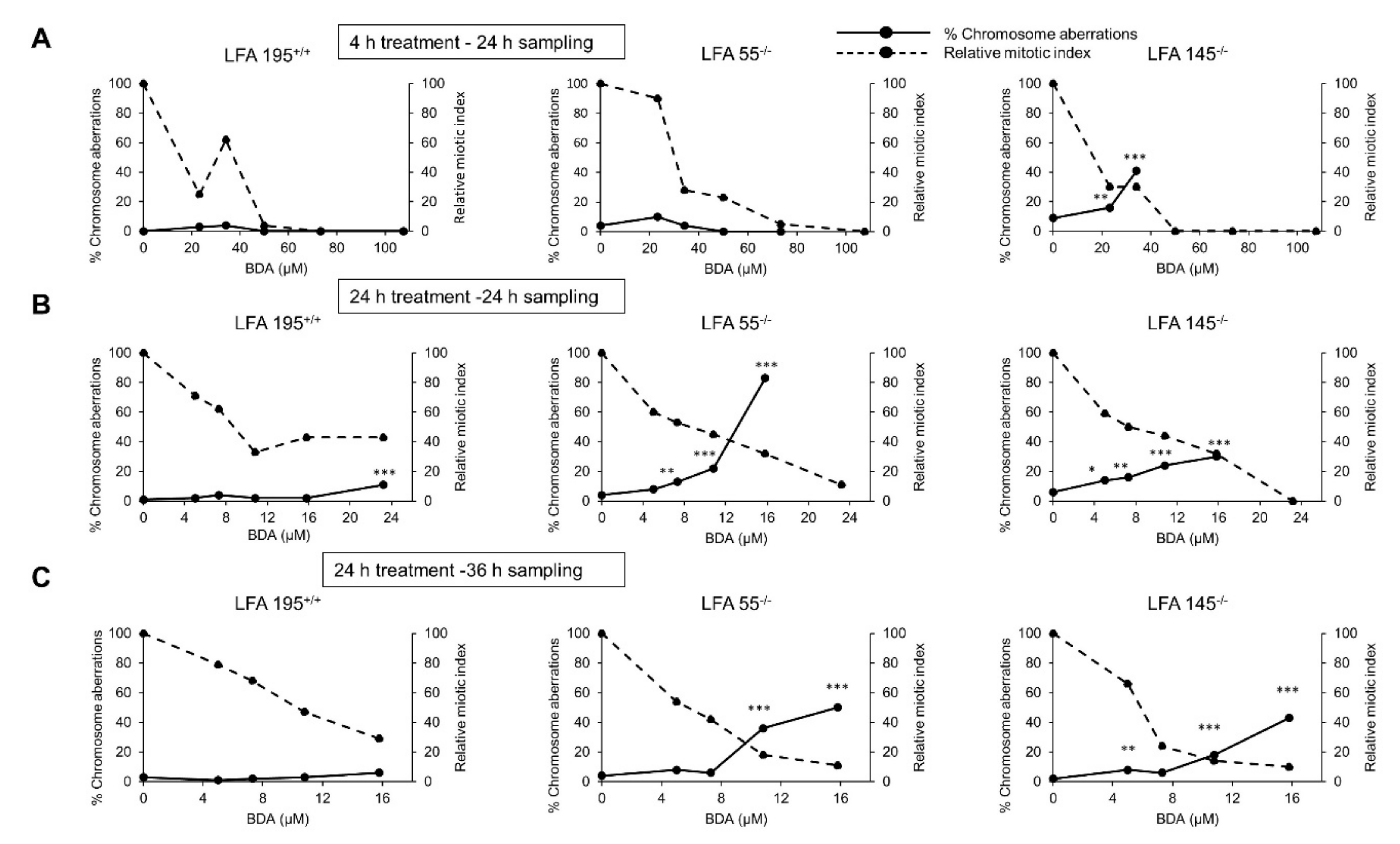
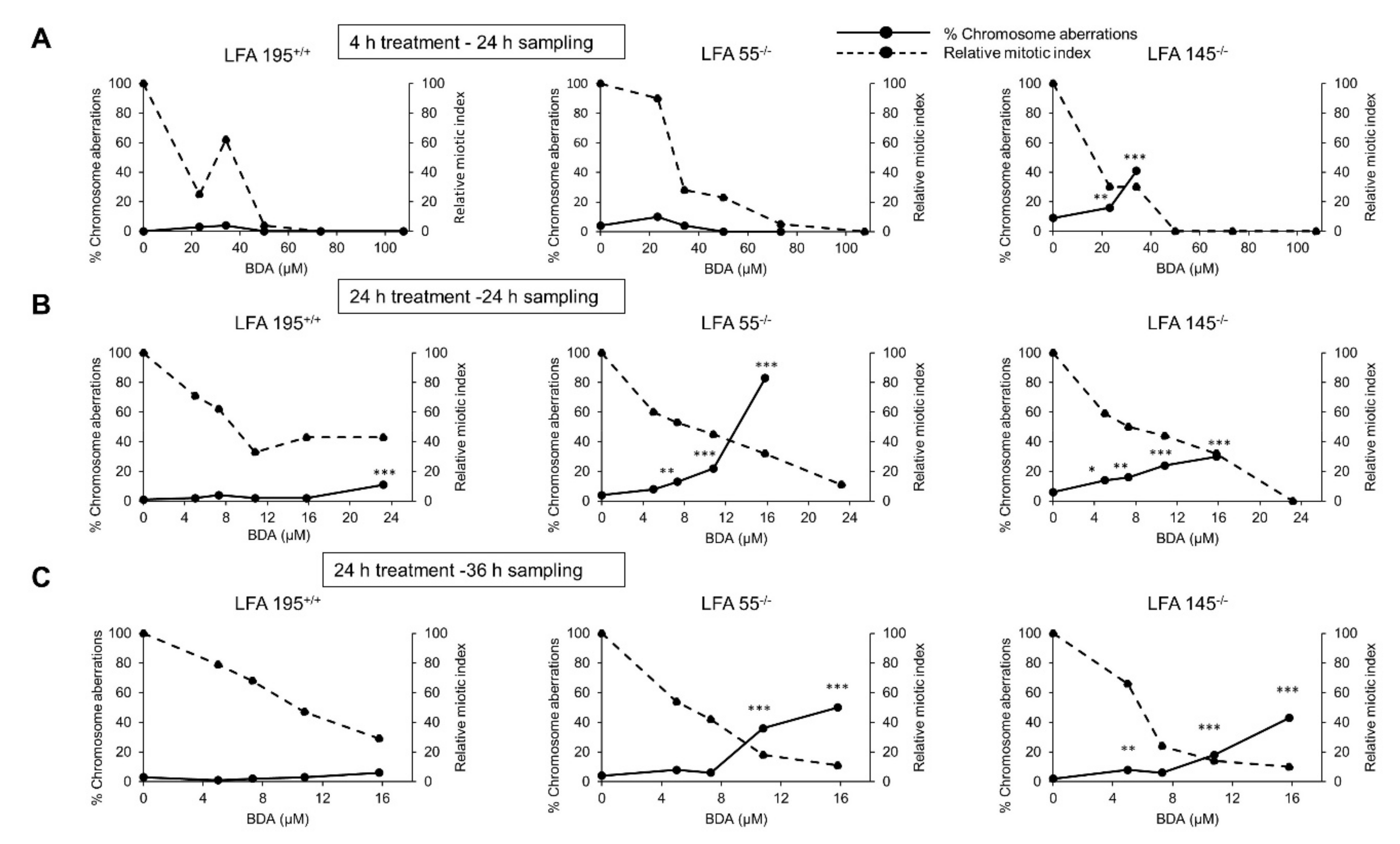
2.4. Furan Induces Chromosomal Aberrations in the Presence of Both S9 Metabolism and a Defect in DNA ICL Repair
2.5. A Defect in the Repair of DNA ICLs Significantly Increases the Frequency of Chromosomal Aberrations Induced by BDA
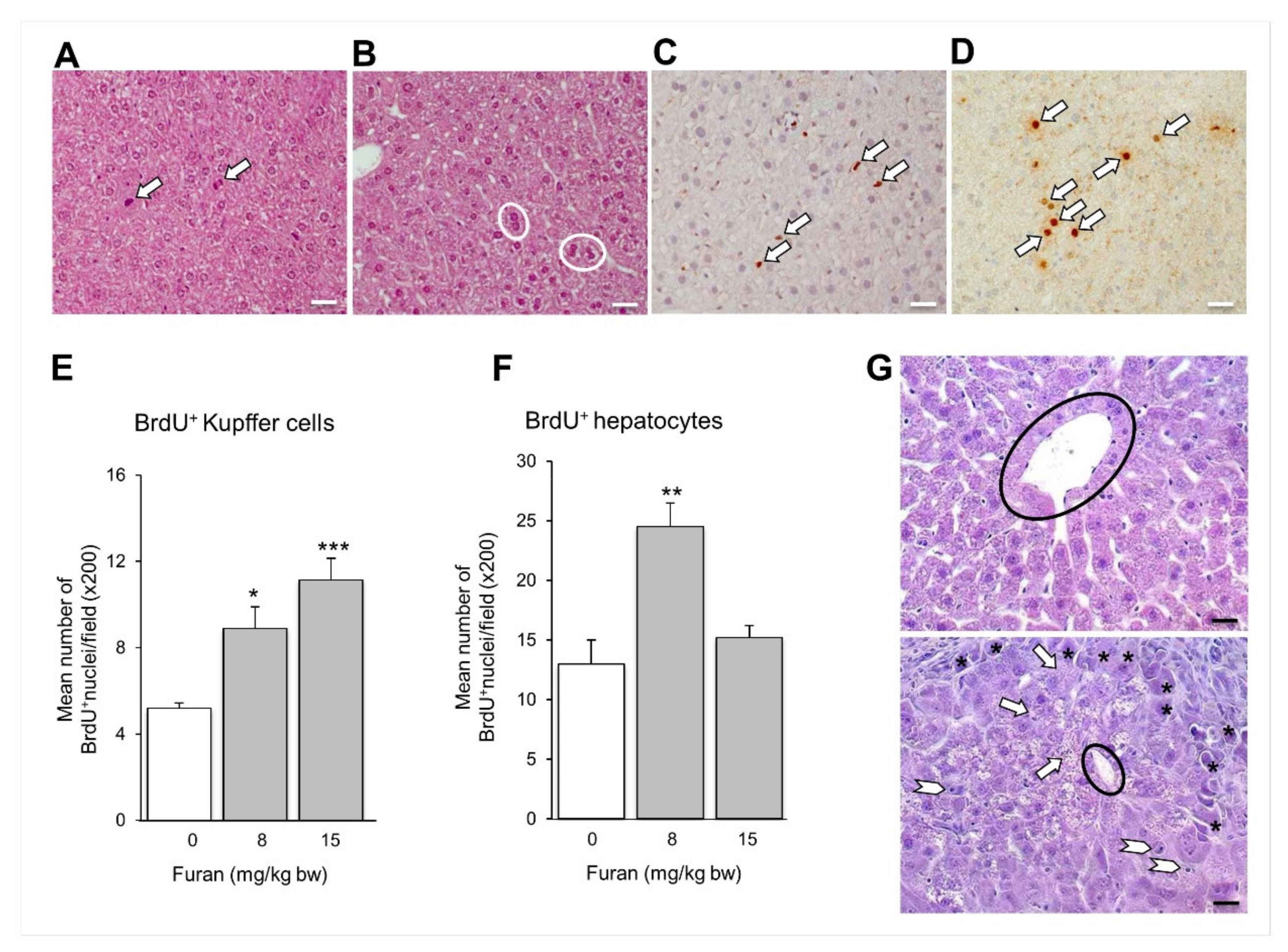
2.6. Subacute Oral Administration of Furan to C57BL6 Mouse Strain Causes Liver Toxicity
2.7. Subchronic Exposure to Furan Results in a Moderate Increase in Micronucleus Frequency in Splenocytes with No Signs of Spleen Toxicity
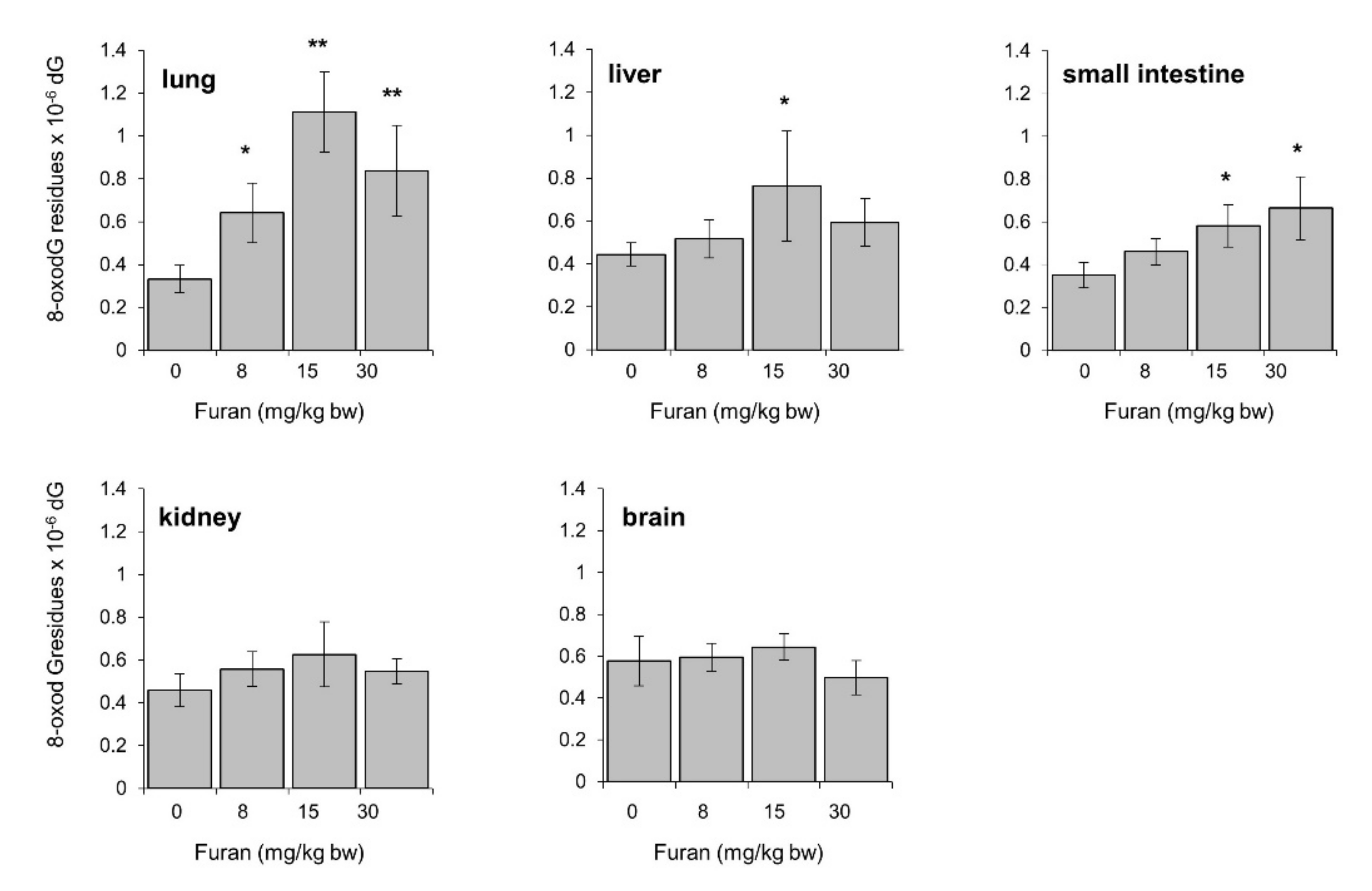
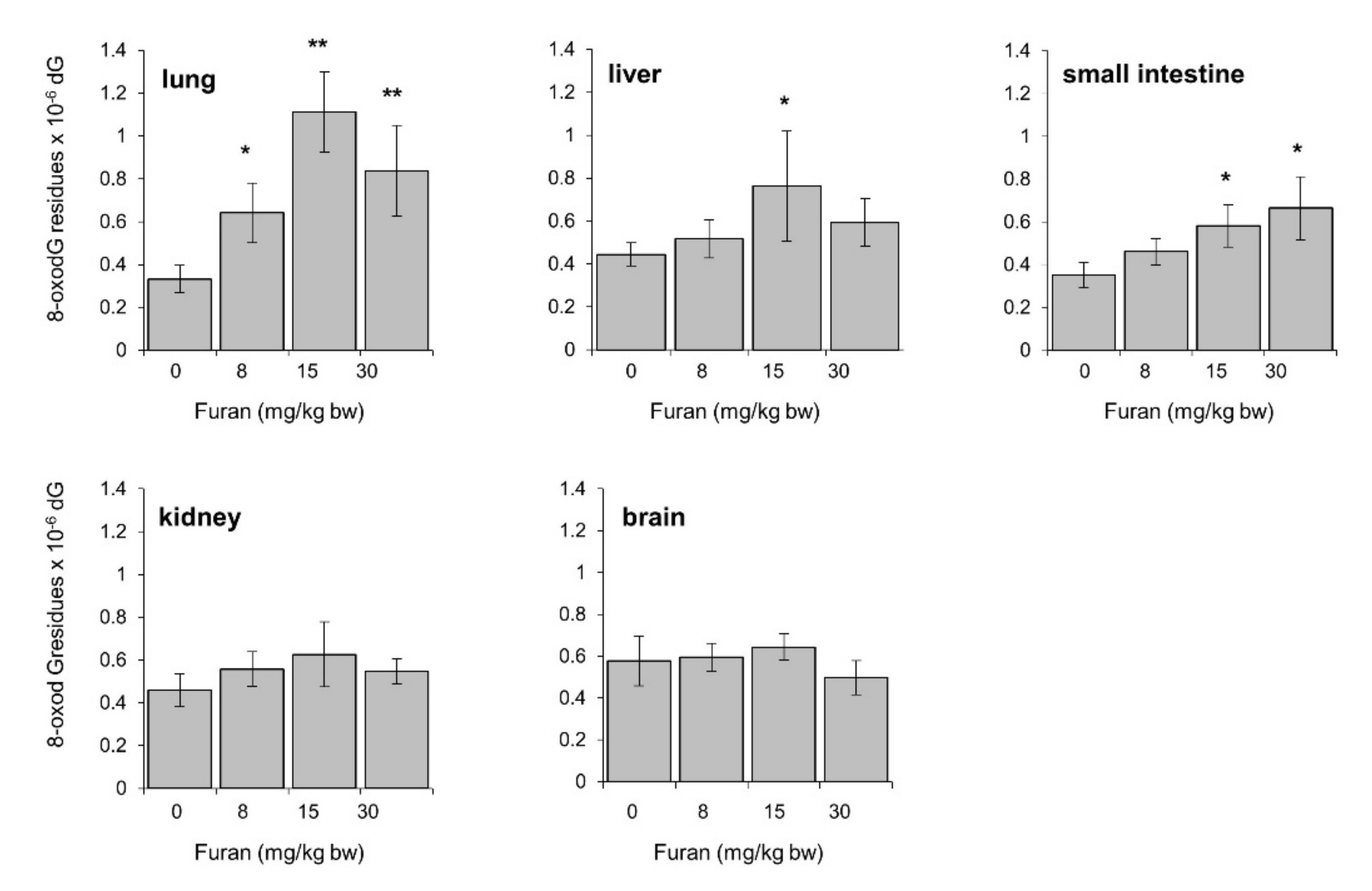
2.8. Furan-Induced Chromosomal Damage in the Spleen Is Independent from Oxidatively Induced DNA Damage
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Preparation of In Vitro Metabolic Activation System (S9)
4.3. Cell Lines
4.4. Treatment of Cells in Culture
4.5. V79 Hprt Mutation Assay
4.6. Mouse Lymphoma Tk+/− Mutation Assay
4.7. Cytokinesis-Block Micronucleus Assay (CBMN) in the L5178Y Tk+/− Mouse Lymphoma Cell Line with BDA
4.8. Chromosomal Aberration Assay in Fanconi Anaemia Lymphoblastoid Cell Lines
4.9. Animals
4.10. Treatment of Animals
4.11. Histopathology and BrdU Incorporation Assay
4.12. Isolation of Splenocytes
4.13. Cytokinesis-Block Micronucleus Assay (CBMN) In Vivo
4.14. Preparation of Liver, Spleen, Kidney, Brain, Small Intestine, and Lung DNA
4.15. Measurements of DNA 8-oxodG
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- National Toxicology Program (NTP). Proceedings of the 14th Report on Carcinogens, Research Triangle Park; Department of Health and Human Services, Public Health Service: Durham, NC, USA, 2016.
- National Toxicology Program (NTP). Toxicology and Carcinogenesis Studies of Furan (CAS No. 110-00-9) in F344/N Rats and B6C3Fl Mice (Gavage Studies) in NTP Technical Report No. 402; US Department of Health and Human Services, Public Health Service, National Institutes of Health: Durham, NC, USA, 1993.
- IARC (International Agency for Research on Cancer). Furan 1995, 63, 393–407.
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Risks for public health related to the presence of furan and methylfurans in food. EFSA J. 2017, 15, 1–142. [Google Scholar]
- NCTR (National Center for Toxicological Research). Two-Year Carcinogenicity Bioassay of Furan in F344 Rats; Technical Report for NCTR Experiment No E2168.01 (Test No. E2168.02); NCTR: Redfield, AR, USA, 2015; 102p. [Google Scholar]
- Von Tungeln, L.S.; Walker, N.J.; Olson, G.R.; Mendoza, M.C.; Felton, R.P.; Thorn, B.T.; Marques, M.M.; Pogribny, I.P.; Doerge, D.R.; Beland, F.A. Low dose assessment of the carcinogenicity of furan in male F344/N Nctr rats in a 2-year gavage study. Food Chem. Toxicol. 2017, 99, 170–181. [Google Scholar] [CrossRef] [Green Version]
- Mortelmans, K.; Haworth, S.; Lawlor, T.; Speck, W.; Tainer, B.; Zeiger, E. Salmonella mutagenicity tests: II. Results from the testing of 270 chemicals. Environ. Mutagen. 1986, 8 (Suppl. S7), 1–119. [Google Scholar] [CrossRef]
- Lee, H.; Bian, S.S.; Chen, Y.L. Genotoxicity of 1,3-dithiane and 1,4-dithiane in the CHO/SCE assay and the Salmonella/microsomal test. Mutat. Res. 1994, 321, 213–218. [Google Scholar] [CrossRef]
- McGregor, D.B.; Brown, A.; Cattanach, P.; Edwards, I.; McBride, D.; Riach, C.; Caspary, W.J. Responses of the L5178y tk+/tk− mouse lymphoma cell forward mutation assay: III. 72 Coded chemicals. Environ. Mutagen. 1988, 12, 85–154. [Google Scholar] [CrossRef]
- Kellert, M.; Brink, A.; Richter, I.; Schlatter, J.; Lutz, W.K. Tests for genotoxicity and mutagenicity of furan and its metabolite cis-2-butene-1,4-dial in L5178Y tk+/− mouse lymphoma cells. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2008, 657, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Stich, H.F.; Rosin, M.P.; Wu, C.H.; Powrie, W.D. Clastogenicity of furans found in food. Cancer Lett. 1991, 13, 89–95. [Google Scholar] [CrossRef]
- Glatt, H.; Schneider, H.; Liu, Y. V79-hCYP2E1-hSULT1A1, a cell line for the sensitive detection of genotoxic effects induced by carbohydrate pyrolysis products and other food-borne chemicals. Mutat. Res. 2005, 580, 41–52. [Google Scholar] [CrossRef]
- Durling, L.; Svensson, K.; Abramssonzetterberg, L. Furan is not genotoxic in the micronucleus assay in vivo or in vitro. Toxicol. Lett. 2007, 169, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Cordelli, E.; Leopardi, P.; Villani, P.; Marcon, F.; Macri, C.; Caiola, S.; Siniscalchi, E.; Conti, L.; Eleuteri, P.; Malchiodi-Albedi, F.; et al. Toxic and genotoxic effects of oral administration of furan in mouse liver. Mutagenesis 2010, 25, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Leopardi, P.; Cordelli, E.; Villani, P.; Cremona, T.P.; Conti, L.; De Luca, G.; Crebelli, R. Assessment of in vivo genotoxicity of the rodent carcinogen furan: Evaluation of DNA damage and induction of micronuclei in mouse splenocytes. Mutagenesis 2010, 25, 57–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDaniel, L.P.; Wei, D.; Dobrovolsky, V.N.; Shaddock, J.G., Jr.; Mittelstaedt, R.A.; Doerge, D.R.; Heflich, R.H. Genotoxicity of furan in Big Blue rats. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2012, 742, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Terrell, A.N.; Huynh, M.; Grill, A.E.; Kovi, R.C.; O’Sullivan, M.G.; Guttenplan, J.B.; Ho, Y.Y.; Peterson, L.A. Mutagenicity of furan in female Big Blue B6C3F1 mice. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2014, 770, 46–54. [Google Scholar] [CrossRef] [Green Version]
- Hoie, A.H.; Svendsen, C.; Brunborg, G.; Glatt, H.; Alexander, J.; Meinl, W.; Husoy, T. Genotoxicity of three food processing contaminants in transgenic mice expressing human sulfotransferases 1A1 and 1A2 as assessed by the in vivo alkaline single cell gel electrophoresis assay. Environ. Mol. Mutagen. 2015, 56, 709–714. [Google Scholar] [CrossRef] [Green Version]
- Hibi, D.; Yokoo, Y.; Suzuki, Y.; Ishii, Y.; Jin, M.; Kijima, A.; Nohmi, T.; Nishikawa, A.; Umemura, T. Lack of genotoxic mechanisms in early-stage furan-induced hepatocellular tumorigenesis in gpt delta rats. J. Appl. Toxicol. 2016, 37, 142–149. [Google Scholar] [CrossRef]
- Garle, M.J.; Fry, J.R. Reactive metabolite formation catalysed by cytochrome P-450. Toxicol. In Vitro 1990, 4, 493–496. [Google Scholar] [CrossRef]
- Chen, L.J.; Hecht, S.S.; Peterson, L.A. Identification of cis-2-butene-1,4-dial as a microsomal metabolite of furan. Chem. Res. Toxicol. 1995, 8, 903–906. [Google Scholar] [CrossRef]
- Parmar, D.; Burka, L.T. Studies on the interaction of furan with hepatic cytochrome P-450. J. Biochem. Toxicol. 1993, 8, 1–9. [Google Scholar] [CrossRef]
- Chen, L.J.; Hecht, S.S.; Peterson, L.A. Characterization of amino acid and glutathione adducts of cis-2-butene-1,4-dial, a reactive metabolite of furan. Chem. Res. Toxicol. 1997, 10, 866–874. [Google Scholar] [CrossRef]
- Gingipalli, L.; Dedon, P.C. Reaction of cis- and trans-2-Butene-1,4-dial with 2′-Deoxycytidine to Form Stable Oxadiazabicyclooctaimine Adducts. J. Am. Chem. Soc. 2001, 123, 2664–2665. [Google Scholar] [CrossRef] [PubMed]
- Byrns, M.C.; Predecki, D.P.; Peterson, L.A. Characterization of nucleoside adducts of cis-2-butene-1,4-dial, a reactive metabolite of furan. Chem. Res. Toxicol. 2002, 15, 373–379. [Google Scholar] [CrossRef]
- Byrns, M.C.; Vu, C.C.; Neidigh, J.W.; Abad, J.L.; Jones, R.A.; Peterson, L.A. Detection of DNA adducts derived from the reactive metabolite of furan, cis-2-butene-1,4-dial. Chem. Res. Toxicol. 2006, 19, 414–420. [Google Scholar] [CrossRef] [Green Version]
- Marinari, U.M.; Ferro, M.; Bassi, A.M.; Sciaba, L.; Finollo, R.; Brambilla, G. DNA-damaging activity of biotic and xenobiotic aldehydes in Chinese hamster ovary cells. Cell Biochem. Funct. 1984, 2, 243–248. [Google Scholar] [CrossRef]
- Byrns, M.C.; Vu, C.C.; Peterson, L.A. The formation of substituted 1,N-6-etheno-2′-deoxyadenosine and 1,N-2-etheno-2′-deoxyguanosine adducts by cis-2-butene-1,4-dial, a reactive metabolite of furan. Chem. Res. Toxicol. 2004, 17, 1607–1613. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L.A.; Naruko, K.C.; Predecki, D.P. A reactive metabolite of furan, cis-2-butene-1,4-dial, is mutagenic in the Ames assay. Chem. Res. Toxicol. 2000, 13, 531–534. [Google Scholar] [CrossRef] [PubMed]
- Hickling, K.C.; Hitchcock, J.M.; Oreffo, V.; Mally, A.; Hammond, T.G.; Evans, J.G.; Chipman, J.K. Evidence of oxidative stress and associated DNA damage, increased proliferative drive, and altered gene expression in rat liver produced by the cholangiocarcinogenic agent furan. Toxicol. Pathol. 2010, 38, 230–243. [Google Scholar] [CrossRef]
- Ding, W.; Petibone, D.M.; Latendresse, J.R.; Pearce, M.G.; Muskhelishvili, L.; White, G.A.; Chang, C.W.; Mittelstaedt, R.A.; Shaddock, J.G.; McDaniel, L.P.; et al. In vivo genotoxicity of furan in F344 rats at cancer bioassay doses. Toxicol. Appl. Pharmacol. 2012, 261, 164–171. [Google Scholar] [CrossRef]
- Ceccaldi, R.; Sarangi, P.; D’Andrea, A.D. The Fanconi anaemia pathway: New players and new functions. Nat. Rev. Mol. Cell Biol. 2016, 17, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.T.; De Luca, G.; Degan, P.; Parlanti, E.; Dogliotti, E.; Barnes, D.E.; Lindahl, T.; Yang, H.; Miller, J.H.; Bignami, M. Accumulation of the oxidative base lesion 8-hydroxyguanine in DNA of tumor-prone mice defective in both the Myh and Ogg1 DNA glycosylases. Cancer Res. 2004, 64, 4411–4414. [Google Scholar] [CrossRef] [Green Version]
- Neuwirth, C.; Mosesso, P.; Pepe, G.; Fiore, M.; Malfatti, M.; Turteltaub, K.; Dekant, W.; Mally, A. Furan carcinogenicity: DNA binding and genotoxicity of furan in rats in vivo. Mol. Nutr. Food Res. 2012, 56, 1363–1374. [Google Scholar] [CrossRef]
- Alizadeh, M.; Barati, M.; Saleh-Ghadimi, S.; Neda Roshanravan, N.; Zeinalian, R.; Jabbari, M. Industrial furan and its biological effects on the body systems. J. Food Biochem. 2018, 42, e12597. [Google Scholar] [CrossRef]
- Halila, S.; Velasco, T.; Clercq, P.D.; Madder, A. Fine-tuning furan toxicity: Fast and quantitative DNA interchain cross-link formation upon selective oxidation of a furan containing oligonucleotide. Chem. Commun. 2005, 7, 936. [Google Scholar] [CrossRef]
- Castella, M.; Pujol, R.; Callén, E.; Trujillo, J.P.; Casado, J.A.; Gille, H.; Lach, F.P.; Auerbach, A.D.; Schindler, D.; Benítez, J.; et al. Origin, functional role, and clinical impact of Fanconi anemia FANCA mutations. Blood 2011, 117, 3759–3769. [Google Scholar] [CrossRef] [PubMed]
- Palma, N.; Cinelli, S.; Sapora, O.; Wilson, S.H.; Dogliotti, E. Ochratoxin A-Induced Mutagenesis in Mammalian Cells Is Consistent with the Production of Oxidative Stress. Chem. Res. Toxicol. 2007, 20, 1031–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, W.D. Statistical Evaluation of Mutagenicity Test Data; Kirkland, D.J., Ed.; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Moore, M.M.; Honma, M.; Clements, J.; Bolcsfoldi, G.; Burlinson, B.; Cifone, M.; Clarke, J.; Delongchamp, R.; Durward, R.; Fellows, M.; et al. Mouse Lymphoma Thymidine Kinase Gene Mutation Assay: Follow-Up Meeting of the International Workshop on Genotoxicity Tests—Aberdeen, Scotland, 2003—Assay Acceptance Criteria, Positive Controls, and Data Evaluation. Environ. Mol. Mutagen. 2006, 47, 1–5. [Google Scholar] [CrossRef]
- Savage, J.R.K. Classification and relationships of induced chromosomal structural changes. J. Med. Genet. 1976, 13, 103–122. [Google Scholar] [CrossRef] [Green Version]
- Klungland, A.; Rosewell, I.; Hollenbach, S.; Larsen, E.; Daly, G.; Epe, B.; Seeberg, E.; Lindahl, T.; Barnes, D.E. Accumulation of premutagenic DNA lesions in mice defective in removal of oxidative base damage. Proc. Natl. Acad. Sci. USA 1999, 96, 13300–13305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landis, J.R.; Koch, G.G. The measurement of observer agreement for categorical data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dose (mM) | RTG (%) | MF (×10−6 Cells) | Induced MF (×10−6 Cells) | Small Colonies MF (×10−6 Cells) | Large Colonies MF (×10−6 Cells) | Small/Large Colonies (%) |
|---|---|---|---|---|---|---|
| 0 | 100 | 90.7 | - | 38.4 | 48.9 | 0.44 |
| 1.78 | 109 | 70.6 | 0 | 32.0 | 41.1 | 0.44 |
| 2.37 | 133 | 74.2 | 0 | 28.3 | 40.7 | 0.41 |
| 3.13 | 125 | 61.7 | 0 | 15.2 | 40.8 | 0.27 |
| 4.22 | 97 | 72.3 | 0 | 30.5 | 38.6 | 0.44 |
| 5.62 | 96 | 76.1 | 0 | 29.2 | 42.9 | 0.41 |
| 7.50 § | 3 | 294.5 | 203.8 | 182.3 | 94.7 | 0.66 |
| Linear trend | NS | NS | NS | |||
| MMS 45.4 × 10−3 | 125 | 344.1 ** | 253.4 | 154.5 ** | 131.3 ** | 0.54 |
| 0 | 100 | 66.8 | - | 14.4 | 50.6 | 0.22 |
| 3.06 | 82 | 55.6 | 0 | 15.4 | 38.7 | 0.28 |
| 3.35 | 70 | 74.5 | 7.71 | 16.1 | 56.0 | 0.22 |
| 3.67 | 51 | 75.0 | 8.24 | 18.4 | 54.7 | 0.25 |
| 4.03 | 46 | 80.4 | 13.60 | 28.4 | 49.5 | 0.36 |
| 4.42 | 50 | 103.4 | 36.62 | 27.5 | 72.3 | 0.28 |
| 4.84 | 24 | 113.2 | 46.43 | 32.2 | 77.5 | 0.29 |
| 5.31 § | 11 | 145.0 | 78.16 | 65.1 | 71.2 | 0.48 |
| 5.82 § | 8 | 127.5 | 60.68 | 51.4 | 70.0 | 0.42 |
| Linear trend | * p < 0.05 | * p < 0.05 | NS | |||
| MMS 45.4 × 10−3 | 125 | 424.4 ** | 357.6 | 119.2 ** | 242.4 ** | 0.54 |
| Dose (μM) | RTG (%) | MF (×10−6 Cells) | Induced MF (×10−6 Cells) | Small Colonies MF (×10−6 Cells) | Large Colonies MF (×10−6 Cells) | Small/Large Colonies (%) |
|---|---|---|---|---|---|---|
| 0 | 100 | 57.9 | - | 21.7 | 35.3 | 0.38 |
| 7.5 | 84 | 68.6 | 10.71 | 32.0 | 34.5 | 0.48 |
| 10 | 85 | 70.4 | 12.50 | 21.4 | 46.5 | 0.32 |
| 15 | 93 | 71.3 | 13.38 | 25.6 | 42.9 | 0.37 |
| 20 | 82 | 82.8 | 24.95 | 28.9 | 50.4 | 0.36 |
| 40 | 64 | 118.7 ** | 60.86 | 64.9 ** | 45.5 | 0.59 |
| 80 § | 6 | 268.4 | 210.5 | 133.8 | 109.3 | 0.55 |
| Linear trend | *** | *** | NS | |||
| MMS 90.8 | 88 | 343.1 ** | 285.2 | 136.6 ** | 155.7 ** | 0.47 |
| 0 | 100 | 62.0 | - | 26.3 | 34.3 | 0.43 |
| 20 | 110 | 63.9 | 1.86 | 29.5 | 33.3 | 0.47 |
| 30 | 105 | 66.5 | 4.53 | 27.9 | 37.6 | 0.43 |
| 40 | 65 | 87.0 | 25.01 | 49.6 | 36.4 | 0.58 |
| 50 § | 10 | 483.2 | 421.2 | 248.1 | 200.9 | 0.55 |
| 60 § | 5 | 516.5 | 454.5 | 302.4 | 182.3 | 0.62 |
| Linear trend | NS | NS | NS | |||
| MMS 90.8 | 127 | 244.3 ** | 182.3 | 117.3 ** | 79.1 * | 0.60 |
| Treatment | Dose (µM) | Cytotoxicity (%) | MnBn Cells (‰) | Total Micronuclei | Distribution of Micronuclei | |||
|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | |||||
| 4 h treatment | ||||||||
| Untreated | 0 | - | 8 | 16 | 1984 | 16 | - | - |
| BDA | 10 | 10 | 14 | 28 | 1972 | 28 | - | - |
| BDA | 15 | 23 | 22 *** | 44 | 1956 | 44 | - | - |
| BDA | 20 | 27 | 18 * | 36 | 1964 | 36 | - | - |
| 24 h treatment | ||||||||
| Untreated | 0 | - | 12 | 24 | 1976 | 24 | - | - |
| BDA | 10 | 49 | 20 | 40 | 1960 | 40 | - | - |
| BDA | 15 | 55 | 27 *** | 54 | 1947 | 52 | 1 | - |
| BDA | 20 | 62 | 36 *** | 75 | 1931 | 69 | 3 | - |
| MMC | 1.5 μg/mL | 2 | 24 *** | 51 | 1952 | 46 | 1 | 1 |
| Furan (mg/kg bw) | Survived Mice (No.) | Final Body Weight (g) | Liver | Spleen | ||
|---|---|---|---|---|---|---|
| Mean ± SE | Weight (g) Mean ± SE | Hepatic a Index Mean ± SE | Weight (mg) Mean ± SE | Splenic a Index Mean ± SE | ||
| 0 | 5 | 23.3 ± 1.4 | 1.03 ± 0.1 | 4.40 ± 0.6 | 73.9 ± 11.5 | 0.32 ± 0.5 |
| 8 | 6 § | 24.1 ± 0.4 | 1.29 ± 0.1 | 5.37 ± 0.3 * | 84.2 ± 8.6 | 0.35 ± 0.02 |
| 15 | 5 | 22.6 ± 2.7 | 1.25 ± 1.2 | 5.50 ± 0.4 ** | 85.1 ± 24.4 | 0.39 ± 0.15 |
| 30 | 6 | 22.0 ± 0.9 | 1.18 ± 0.3 | 5.31 ± 1.3 | 75.7 ± 16.4 | 0.34 ± 0.08 |
| MnBn | ||||
|---|---|---|---|---|
| Furan Dose | Mice | MnBn/1000 Cells | Frequency | |
| mg/kg bw | No. | Individual Animal Data | Group Mean ± SE | |
| Exp. 1 | 0 | 5 | 5, 3, 2, 5, 1 | 3.2 ± 0.8 |
| 8 | 5 | 6, 1, 9, 8, 8 | 6.4 ± 1.4 | |
| 15 | 5 | 8, 9, 6, 3, 7 | 6.6 * ± 1.1 | |
| 30 | 6 | 7, 16, 7,10, 6, 9 | 9.2 ** ± 1.5 | |
| Exp. 2 | 0 | 6 | 2, 4, 2, 1, 3, 2 | 4.6 ± 0.8 |
| 8 | 6 | 1, 3, 5, 3, 7, 4 | 7.6 ± 0.4 | |
| 15 | 6 | 6, 3, 3,4, 7, 3 | 8.6 * ± 1.1 | |
| 30 | 5 | 12, 5, 10, 8, 9 | 17.6 ** ± 1.5 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Russo, M.T.; De Luca, G.; Palma, N.; Leopardi, P.; Degan, P.; Cinelli, S.; Pepe, G.; Mosesso, P.; Di Carlo, E.; Sorrentino, C.; et al. Oxidative Stress, Mutations and Chromosomal Aberrations Induced by In Vitro and In Vivo Exposure to Furan. Int. J. Mol. Sci. 2021, 22, 9687. https://doi.org/10.3390/ijms22189687
Russo MT, De Luca G, Palma N, Leopardi P, Degan P, Cinelli S, Pepe G, Mosesso P, Di Carlo E, Sorrentino C, et al. Oxidative Stress, Mutations and Chromosomal Aberrations Induced by In Vitro and In Vivo Exposure to Furan. International Journal of Molecular Sciences. 2021; 22(18):9687. https://doi.org/10.3390/ijms22189687
Chicago/Turabian StyleRusso, Maria Teresa, Gabriele De Luca, Nieves Palma, Paola Leopardi, Paolo Degan, Serena Cinelli, Gaetano Pepe, Pasquale Mosesso, Emma Di Carlo, Carlo Sorrentino, and et al. 2021. "Oxidative Stress, Mutations and Chromosomal Aberrations Induced by In Vitro and In Vivo Exposure to Furan" International Journal of Molecular Sciences 22, no. 18: 9687. https://doi.org/10.3390/ijms22189687
APA StyleRusso, M. T., De Luca, G., Palma, N., Leopardi, P., Degan, P., Cinelli, S., Pepe, G., Mosesso, P., Di Carlo, E., Sorrentino, C., Musiani, P., Crebelli, R., Bignami, M., & Dogliotti, E. (2021). Oxidative Stress, Mutations and Chromosomal Aberrations Induced by In Vitro and In Vivo Exposure to Furan. International Journal of Molecular Sciences, 22(18), 9687. https://doi.org/10.3390/ijms22189687