
Differential Regulation of Interferon Signaling Pathways in CD4+ T Cells of the Low Type-2 Obesity-Associated Asthma Phenotype

 ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Results
2.1. Patient Selection and Clinical Parameters Comparison
2.2. Differentially Expressed Genes (DEGs) of CD4+ T Cells in Low Type-2 Obese and Non-Obese Asthmatics
2.3. Predominant Interferon Signaling Pathway in CD4+ T Cells of Low Type-2 Obese Asthmatics
2.4. Further Gene Regulation Pathways Involved in Low Type-2 Asthma Phenotypes
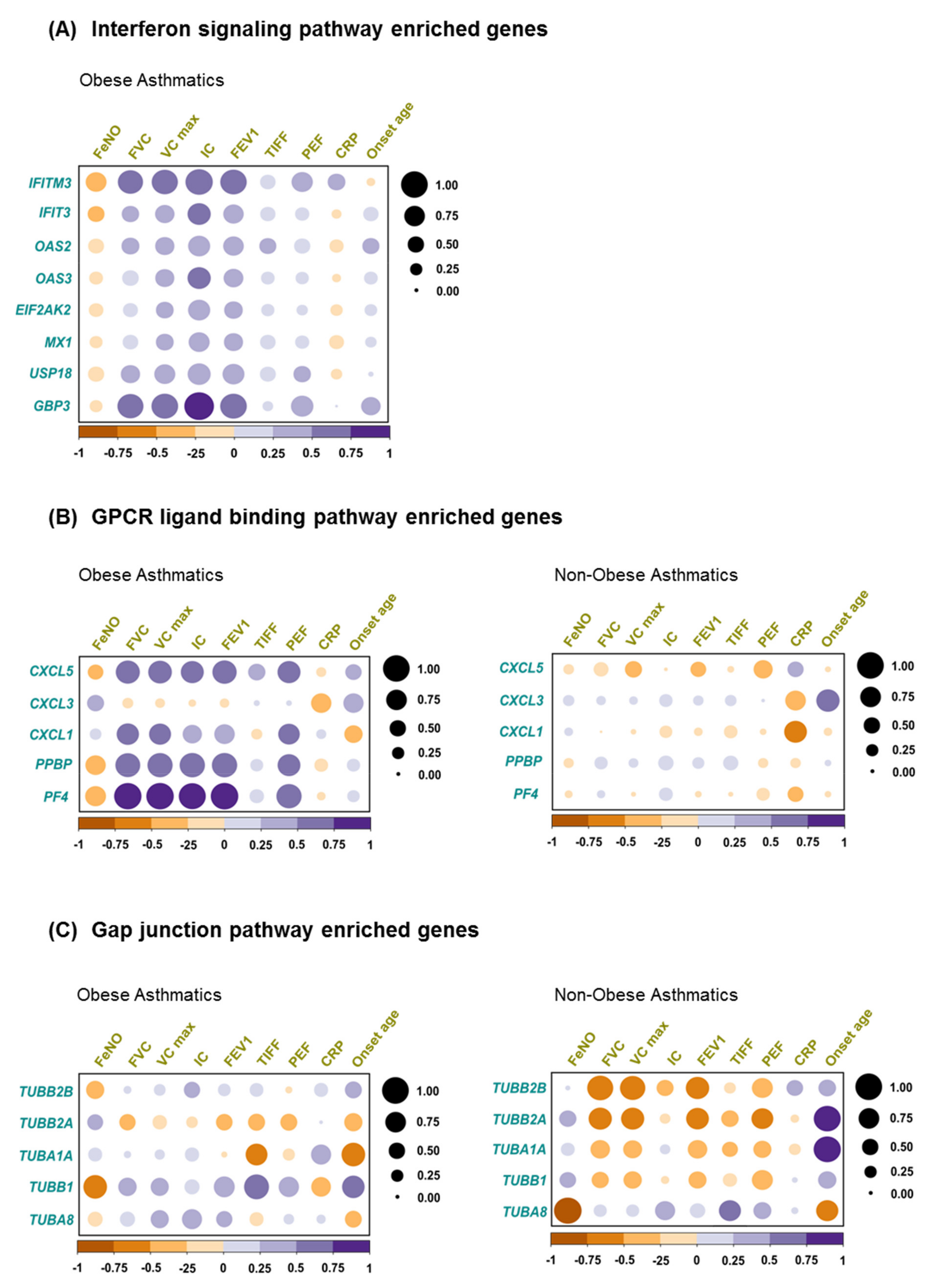
2.5. Correlation of Biological Pathway Enriched Genes with Clinical Variables
3. Discussion
4. Material and Methods
4.1. Study Population and Clinical Assessments
4.2. Blood CD4+ T Cells Isolation
4.3. RNA Extraction and Sequencing
4.4. Bioinformatics Analysis
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pearce, N.; Asher, I.; Billo, N.; Bissell, K.; Ellwood, P.; El Sony, A.; Garcia-Marcos, L.; Chiang, C.-Y.; Mallol, J.; Marks, G.; et al. Asthma in the global NCD agenda: A neglected epidemic. Lancet Respir. Med. 2013, 1, 96–98. [Google Scholar] [CrossRef]
- The Top 10 Causes of Death. 14 July 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 27 July 2021).
- Habib, S.H.; Saha, S. Burden of non-communicable disease: Global overview. Diabetes Metab. Syndr. Clin. Res. Rev. 2010, 4, 41–47. [Google Scholar] [CrossRef]
- Nugent, R.A.; Husain, M.J.; Kostova, D.; Chaloupka, F. Introducing the PLOS special collection of economic cases for NCD prevention and control: A global perspective. PLoS ONE 2020, 15, e0228564. [Google Scholar] [CrossRef] [PubMed]
- Kankeu, H.T.; Saksena, P.; Xu, K.; Evans, D.B. The financial burden from non-communicable diseases in low- and middle-income countries: A literature review. Health Res. Policy Syst. 2013, 11, 31. [Google Scholar] [CrossRef] [PubMed]
- Gowshall, M.; Taylor-Robinson, S.D. The increasing prevalence of non-communicable dis-eases in low-middle income countries: The view from Malawi. Int. J. Gen. Med. 2018, 11, 255–264. [Google Scholar] [CrossRef]
- Sivanantham, P.; Sahoo, J.; Lakshminarayanan, S.; Bobby, Z.; Kar, S.S. Profile of risk factors for Non-Communicable Diseases (NCDs) in a highly urbanized district of India: Findings from Puducherry district-wide STEPS Survey, 2019–2020. PLoS ONE 2021, 16, e0245254. [Google Scholar] [CrossRef]
- Noncommunicable Diseases. 16 July 2021. Available online: https://www.who.int/data/gho/data/themes/noncommunicable-diseases (accessed on 27 July 2021).
- Peters, R.; Ee, N.; Peters, J.; Beckett, N.; Booth, A.; Rockwood, K.; Anstey, K.J. Common risk factors for major noncommunicable disease, a systematic overview of reviews and commentary: The implied potential for targeted risk reduction. Ther. Adv. Chronic Dis. 2019, 10, 2040622319880392. [Google Scholar] [CrossRef] [PubMed]
- Nyberg, S.T.; Batty, G.; Pentti, J.; Virtanen, M.; Alfredsson, L.; Fransson, E.I.; Goldberg, M.; Heikkila, K.; Jokela, M.; Knutsson, A.; et al. Obesity and loss of disease-free years owing to major non-communicable diseases: A multicohort study. Lancet Public Health 2018, 3, e490–e497. [Google Scholar] [CrossRef]
- Kuruvilla, M.E.; Lee, F.E.-H.; Lee, G.B. Understanding Asthma Phenotypes, Endotypes, and Mechanisms of Disease. Clin. Rev. Allergy Immunol. 2019, 56, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Poole, J.A. Asthma is a major noncommunicable disease affecting over 230 million people worldwide and represents the most common chronic disease among children. Int. Immunopharmacol. 2014, 23, 315. [Google Scholar] [CrossRef]
- Wenzel, S.E. Asthma phenotypes: The evolution from clinical to molecular approaches. Nat. Med. 2012, 18, 716–725. [Google Scholar] [CrossRef]
- Potaczek, D.P.; Miethe, S.; Schindler, V.; Alhamdan, F.; Garn, H. Role of airway epithelial cells in the development of different asthma phenotypes. Cell. Signal. 2020, 69, 109523. [Google Scholar] [CrossRef] [PubMed]
- Ali, Z.; Ulrik, C.S. Obesity and asthma: A coincidence or a causal relationship? A systematic review. Respir. Med. 2013, 107, 1287–1300. [Google Scholar] [CrossRef] [PubMed]
- Miethe, S.; Guarino, M.; Alhamdan, F.; Simon, H.-U.; Renz, H.; Dufour, J.-F.; Potaczek, D.P.; Garn, H. Effects of obesity on asthma: Immunometabolic links. Polish Arch. Intern. Med. 2018, 128, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Peters, U.; Dixon, A.E.; Forno, E. Obesity and asthma. J. Allergy Clin. Immunol. 2018, 141, 1169–1179. [Google Scholar] [CrossRef]
- McAlees, J.W.; Lajoie, S.; Dienger, K.; Sproles, A.A.; Richgels, P.K.; Yang, Y.; Khodoun, M.; Azuma, M.; Yagita, H.; Fulkerson, P.C.; et al. Differential control of CD4(+) T-cell subsets by the PD-1/PD-L1 axis in a mouse model of allergic asthma. Eur. J. Immunol. 2015, 45, 1019–1029. [Google Scholar] [CrossRef]
- Muehling, L.M.; Lawrence, M.G.; Woodfolk, J.A. Pathogenic CD4+ T cells in patients with asthma. J. Allergy Clin. Immunol. 2017, 140, 1523–1540. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.S.; Wong, N.; Rao, G.; Nguyen, A.; Avery, D.T.; Payne, K.; Torpy, J.; O’Young, P.; Deenick, E.; Bustamante, J.; et al. Unique and shared signaling pathways cooperate to regulate the differentiation of human CD4+ T cells into distinct effector subsets. J. Exp. Med. 2016, 213, 1589–1608. [Google Scholar] [CrossRef]
- Global Initiative for Asthma—GINA. Global Initiative for Asthma—Global Initiative for Asthma—GINA. 29 July 2021. Available online: https://ginasthma.org/ (accessed on 30 July 2021).
- Radens, C.M.; Blake, D.; Jewell, P.; Barash, Y.; Lynch, K.W. Meta-analysis of transcriptomic varia-tion in T-cell populations reveals both variable and consistent signatures of gene expres-sion and splicing. RNA 2020, 26, 1320–1333. [Google Scholar] [CrossRef]
- Gelfand, E.W.; Schedel, M. Molecular Endotypes Contribute to the Heterogeneity of Asthma. Immunol. Allergy Clin. 2018, 38, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Oriss, T.B.; Wenzel, S.E. Emerging molecular phenotypes of asthma. American journal of physiology. Lung Cell. Mol. Physiol. 2015, 308, L130–L140. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kaur, R.; Chupp, G. Phenotypes and endotypes of adult asthma: Moving toward precision medicine. J. Allergy Clin. Immunol. 2019, 144, 1–12. [Google Scholar] [CrossRef]
- Schoettler, N.; Strek, M.E. Recent Advances in Severe Asthma: From Phenotypes to Personalized Medicine. Chest 2020, 157, 516–528. [Google Scholar] [CrossRef]
- Miura, K.; Inoue, K.; Ogura, A.; Kaminuma, O. Role of CD4+ T Cells in Allergic Airway Diseases: Learning from Murine Models. Int. J. Mol. Sci. 2020, 21, 7480. [Google Scholar] [CrossRef] [PubMed]
- Pepper, A.N.; Renz, H.; Casale, T.B.; Garn, H. Biologic Therapy and Novel Molecular Targets of Severe Asthma. J. Allergy Clin. Immunol. Pract. 2017, 5, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Potaczek, D.P.; Harb, H.; Michel, S.; Alhamwe, B.A.; Renz, H.; Tost, J. Epigenetics and allergy: From basic mechanisms to clinical applications. Epigenomics 2017, 9, 539–571. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.W.; Morris, D.; DelProposto, J.L.; Geletka, L.; Zamarron, B.; Martinez-Santibanez, G.; Meyer, K.A.; Singer, K.; O’Rourke, R.; Lumeng, C.N. An MHC II-dependent activation loop between adipose tissue macrophages and CD4+ T cells controls obesity-induced inflammation. Cell Rep. 2014, 9, 605–617. [Google Scholar] [CrossRef]
- Endo, Y.; Asou, H.K.; Matsugae, N.; Hirahara, K.; Shinoda, K.; Tumes, D.; Tokuyama, H.; Yokote, K.; Nakayama, T. Obesity Drives Th17 Cell Differentiation by Inducing the Lipid Metabolic Kinase, ACC1. Cell Rep. 2015, 12, 1042–1055. [Google Scholar] [CrossRef]
- Duan, Y.; Li, F.; Wang, W.; Guo, Q.; Wen, C.; Li, Y.; Yin, Y. Interleukin-15 in obesity and metabolic dysfunction: Current understanding and future perspectives. Obes. Rev. 2017, 18, 1147–1158. [Google Scholar] [CrossRef]
- Barra, N.G.; Reid, S.; MacKenzie, R.; Werstuck, G.; Trigatti, B.L.; Richards, C.; Holloway, A.C.; Ashkar, A.A. Interleukin-15 contributes to the regulation of murine adipose tissue and human adipocytes. Obesity 2010, 18, 1601–1607. [Google Scholar] [CrossRef]
- Reyes, J.L.; Vannan, D.T.; Vo, T.; Gulamhusein, A.; Beck, P.L.; Reimer, R.A.; Eksteen, B. Neutralization of IL-15 abrogates experimental immune-mediated cholangitis in diet-induced obese mice. Sci. Rep. 2018, 8, 3127. [Google Scholar] [CrossRef]
- Zampieri, T.T.; Da Silva, T.E.O.; de Paula Romeu, D.; Da Silva Torrão, A.; Donato, J., Jr. SOCS3 expression within leptin receptor-expressing cells regulates food intake and leptin sensitivity but does not affect weight gain in pregnant mice consuming a high-fat diet. Physiol. Behav. 2016, 157, 109–115. [Google Scholar] [CrossRef]
- Pedroso, J.; Silveira, M.; Lima, L.B.; Furigo, I.C.; Zampieri, T.T.; Ramos-Lobo, A.M.; Buonfiglio, D.C.; Teixeira, P.; Frazao, R.; Donato, J. Changes in Leptin Signaling by SOCS3 Modulate Fasting-Induced Hyperphagia and Weight Regain in Mice. Endocrinology 2016, 157, 3901–3914. [Google Scholar] [CrossRef]
- Pedroso, J.A.B.; Ramos-Lobo, A.M.; Donato, J. SOCS3 as a future target to treat metabolic dis-orders. Hormones 2019, 18, 127–136. [Google Scholar] [CrossRef]
- Palanivel, R.; Fullerton, M.D.; Galic, S.; Honeyman, J.; Hewitt, K.A.; Jorgensen, S.B.; Steinberg, G.R. Reduced Socs3 expression in adipose tissue protects female mice against obesi-ty-induced insulin resistance. Diabetologia 2012, 55, 3083–3093. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Liu, F. Regulation, Communication, and Functional Roles of Adipose Tissue-Resident CD4+ T Cells in the Control of Metabolic Homeostasis. Front. Immunol. 2018, 9, 1961. [Google Scholar] [CrossRef] [PubMed]
- Kiernan, K.; MacIver, N.J. The Role of the Adipokine Leptin in Immune Cell Function in Health and Disease. Front. Immunol. 2020, 11, 622468. [Google Scholar] [CrossRef] [PubMed]
- Korn, S.; Wilk, M.; Voigt, S.; Weber, S.; Keller, T.; Buhl, R. Measurement of Fractional Exhaled Nitric Oxide: Comparison of Three Different Analysers. RES 2020, 99, 1–8. [Google Scholar] [CrossRef]
- Carty, M.; Bowie, A.G. Recent insights into the role of Toll-like receptors in viral infection. Clin. Exp. Immunol. 2010, 161, 397–406. [Google Scholar] [CrossRef]
- Zakeri, A.; Russo, M. Dual Role of Toll-like Receptors in Human and Experimental Asthma Models. Front. Immunol. 2018, 9, 1027. [Google Scholar] [CrossRef]
- Tang, M.; Henderson, R.J.; Holbrook, J.T.; Que, L.G.; Mathews, A.M.; Wise, R.A.; Dixon, A.E.; Peters, S.P.; Rogers, L.; Smith, L.J.; et al. Does Obesity Increase Respiratory Tract Infections in Patients with Asthma? J. Allergy Clin. Immunol. Pract. 2019, 7, 954–961.e6. [Google Scholar] [CrossRef]
- Jartti, T.; Bønnelykke, K.; Elenius, V.; Feleszko, W. Role of viruses in asthma. Semin. Immunopathol. 2020, 42, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Maccioni, L.; Weber, S.; Elgizouli, M.; Stoehlker, A.-S.; Geist, I.; Peter, H.-H.; Vach, W.; Nieters, A. Obesity and risk of respiratory tract infections: Results of an infection-diary based cohort study. BMC Public Health 2018, 18, 271. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Piernas, C.; Astbury, N.M.; Hippisley-Cox, J.; O’Rahilly, S.; Aveyard, P.; Jebb, S.A. Associa-tions between body-mass index and COVID-19 severity in 6·9 million people in England: A prospective, community-based, cohort study. Lancet Diabetes Endocrinol. 2021, 9, 350–359. [Google Scholar] [CrossRef]
- Yates, T.; Razieh, C.; Zaccardi, F.; Rowlands, A.V.; Seidu, S.; Davies, M.J.; Khunti, K. Obesity, walking pace and risk of severe COVID-19 and mortality: Analysis of UK Biobank. Int. J. Obes. 2021, 45, 1155–1159. [Google Scholar] [CrossRef] [PubMed]
- Wendell, S.G.; Fan, H.; Zhang, C. G Protein-Coupled Receptors in Asthma Therapy: Pharmacology and Drug Action. Pharmacol. Rev. 2020, 72, 1–49. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, D.A.; Penn, R.B. Targeting G protein-coupled receptor signaling in asthma. Cell. Signal. 2006, 18, 2105–2120. [Google Scholar] [CrossRef]
- Hauser, A.S.; Attwood, M.M.; Rask-Andersen, M.; Schiöth, H.B.; Gloriam, D.E. Trends in GPCR drug discovery: New agents, targets and indications. Nat. Rev. Drug Discov. 2017, 16, 829–842. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.; Ursu, O.; Gaulton, A.; Bento, A.P.; Donadi, R.S.; Bologa, C.G.; Karlsson, A.; Al-Lazikani, B.; Hersey, A.; Oprea, T.I.; et al. A comprehensive map of molecular drug targets. Nat. Rev. Drug Discov. 2017, 16, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Elgueta, R.; Tobar, J.A.; Shoji, K.F.; De Calisto, J.; Kalergis, A.; Bono, M.R.; Rosemblatt, M.; Sáez, J.C. Gap junctions at the dendritic cell-T cell interface are key elements for antigen-dependent T cell activation. J. Immunol. 2009, 183, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Giepmans, B.N.; Verlaan, I.; Hengeveld, T.; Janssen, H.; Calafat, J.; Falk, M.M.; Moolenaar, W.H. Gap junction protein connexin-43 interacts directly with microtubules. Curr. Biol. 2001, 11, 1364–1368. [Google Scholar] [CrossRef]
- Bermudez-Fajardo, A.; Ylihärsilä, M.; Evans, W.H.; Newby, A.C.; Oviedo-Orta, E. CD4+ T lympho-cyte subsets express connexin 43 and establish gap junction channel communication with macrophages in vitro. J. Leukoc. Biol. 2007, 82, 608–612. [Google Scholar] [CrossRef]
- Kim, Y.J.; Lee, J.-S.; Kim, H.; Jang, J.H.; Choung, Y.-H. Gap Junction-Mediated Intercellular Commu-nication of cAMP Prevents CDDP-Induced Ototoxicity via cAMP/PKA/CREB Pathway. Int. J. Mol. Sci. 2021, 22, 6327. [Google Scholar] [CrossRef]
- Ring, S.; Karakhanova, S.; Johnson, T.; Enk, A.H.; Mahnke, K. Gap junctions between regulatory T cells and dendritic cells prevent sensitization of CD8(+) T cells. J. Allergy Clin. Immunol. 2010, 125, 237–246. [Google Scholar] [CrossRef]
- Bopp, T.; Becker, C.; Klein, M.; Klein-Heßling, S.; Palmetshofer, A.; Serfling, E.; Heib, V.; Becker, M.; Kubach, J.; Schmitt, S.; et al. Cyclic adeno-sine monophosphate is a key component of regulatory T cell–mediated suppression. J. Exp. Med. 2007, 204, 1303–1310. [Google Scholar] [CrossRef]
- Ram, A.; Singh, S.K.; Singh, V.P.; Kumar, S.; Ghosh, B. Inhaled carbenoxolone prevents allergic airway inflammation and airway hyperreactivity in a mouse model of asthma. Int. Arch. Allergy Immunol. 2009, 149, 38–46. [Google Scholar] [CrossRef]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.X.; Son, E.W.; Yao, R. iDEP: An integrated web application for differential expression and pathway analysis of RNA-Seq data. BMC Bioinform. 2018, 19, 534. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’Ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene Set Knowledge Discovery with Enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Obese Asthmatics | Non-Obese Asthmatics | p Value | |
|---|---|---|---|
| N | 10 | 10 | 1.00 |
| Age (years) | 46.30 ± 10.75 | 39.30 ± 13.94 | 0.23 |
| Male (%) | 30 | 10 | 0.29 |
| BMI (kg/m2) | 36.67 ± 6.90 | 23.88 ± 2.73 | <0.0001 |
| FeNO (ppb) | 15.72 ± 7.93 | 16.78 ± 11.64 | 0.81 |
| FEV1 (L) | 2.89 ± 0.80 | 2.73 ± 1.10 | 0.58 |
| FVC (L) | 3.79 ± 1.00 | 3.50 ± 1.00 | 0.52 |
| VC max (L) | 3.96 ± 1.02 | 3.58 ± 1.00 | 0.42 |
| PEF (L/s) | 6.68 ± 1.54 | 5.96 ± 2.04 | 0.38 |
| FEV1/FVC (%) | 71.32 ± 4.90 | 74.09 ± 12.27 | 0.52 |
| IC (L) | 3.36 ± 0.91 | 2.50 ± 0.67 | 0.03 |
| CRP (mg/L) | 6.80 ± 10.26 | 1.78 ± 2.06 | 0.15 |
| Age of onset (years) | 37.00 ± 15.63 | 28.10 ± 14.11 | 0.20 |
| Obese Asthmatics vs. Ctrl | Non-Obese Asthmatics vs. Ctrl | |||
|---|---|---|---|---|
| Th2 genes | Log2(FC) | FDR | Log2(FC) | FDR |
| GATA3 | −0.489 | 0.001 | −0.446 | 0.004 |
| TNFSF11 | −0.026 | 0.981 | 0.316 | 0.753 |
| IL17RB | −0.062 | 0.909 | 0.083 | 0.909 |
| AKAP12 | 1.031 | 0.368 | 0.916 | 0.511 |
| HPGDS | 0.033 | 0.981 | 0.331 | 0.812 |
| LRRC32 | −0.591 | 0.222 | −0.616 | 0.258 |
| PTGDR2 | 0.160 | 0.726 | 0.598 | 0.114 |
| BACE2 | 0.048 | 0.962 | 0.306 | 0.768 |
| NRIP3 | −0.124 | 0.824 | 0.058 | 0.943 |
| CHDH | −0.396 | 0.442 | 0.106 | 0.902 |
| NCS1 | −0.593 | 0.750 | 0.293 | 0.909 |
| PKP2 | 0.250 | 0.619 | 0.723 | 0.094 |
| LIMA1 | 0.088 | 0.631 | 0.126 | 0.533 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alhamdan, F.; Marsh, L.M.; Pedersen, F.; Alhamwe, B.A.; Thölken, C.; Pfefferle, P.I.; Bahmer, T.; Greulich, T.; Potaczek, D.P.; Garn, H. Differential Regulation of Interferon Signaling Pathways in CD4+ T Cells of the Low Type-2 Obesity-Associated Asthma Phenotype. Int. J. Mol. Sci. 2021, 22, 10144. https://doi.org/10.3390/ijms221810144
Alhamdan F, Marsh LM, Pedersen F, Alhamwe BA, Thölken C, Pfefferle PI, Bahmer T, Greulich T, Potaczek DP, Garn H. Differential Regulation of Interferon Signaling Pathways in CD4+ T Cells of the Low Type-2 Obesity-Associated Asthma Phenotype. International Journal of Molecular Sciences. 2021; 22(18):10144. https://doi.org/10.3390/ijms221810144
Chicago/Turabian StyleAlhamdan, Fahd, Leigh M. Marsh, Frauke Pedersen, Bilal Alashkar Alhamwe, Clemens Thölken, Petra Ina Pfefferle, Thomas Bahmer, Timm Greulich, Daniel P. Potaczek, and Holger Garn. 2021. "Differential Regulation of Interferon Signaling Pathways in CD4+ T Cells of the Low Type-2 Obesity-Associated Asthma Phenotype" International Journal of Molecular Sciences 22, no. 18: 10144. https://doi.org/10.3390/ijms221810144
APA StyleAlhamdan, F., Marsh, L. M., Pedersen, F., Alhamwe, B. A., Thölken, C., Pfefferle, P. I., Bahmer, T., Greulich, T., Potaczek, D. P., & Garn, H. (2021). Differential Regulation of Interferon Signaling Pathways in CD4+ T Cells of the Low Type-2 Obesity-Associated Asthma Phenotype. International Journal of Molecular Sciences, 22(18), 10144. https://doi.org/10.3390/ijms221810144