
SMAD4 Feedback Activates the Canonical TGF-β Family Signaling Pathways


Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
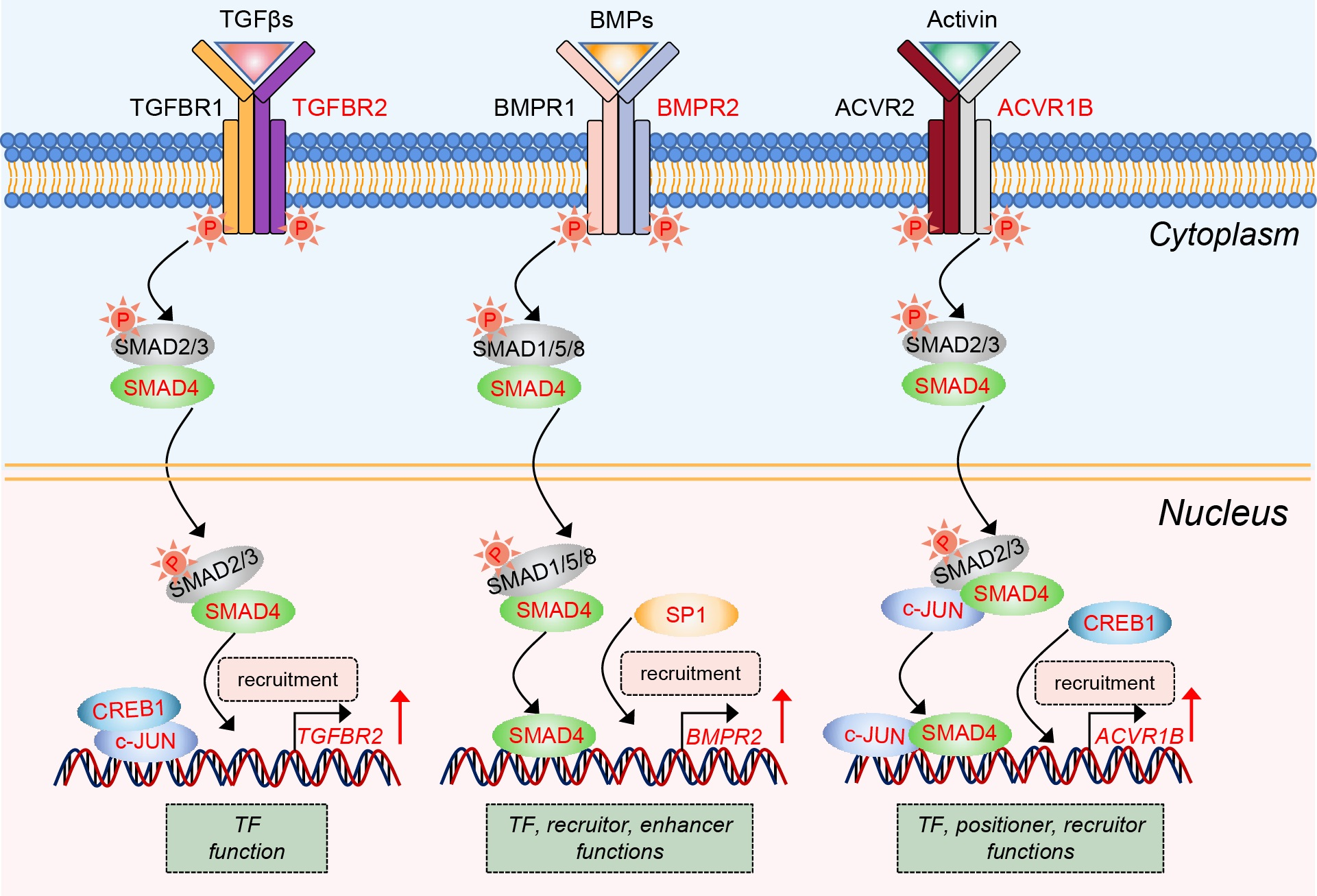
2.1. Reanalysis of the Transcriptomic Alteration in Porcine GCs after Knockdown of SMAD4
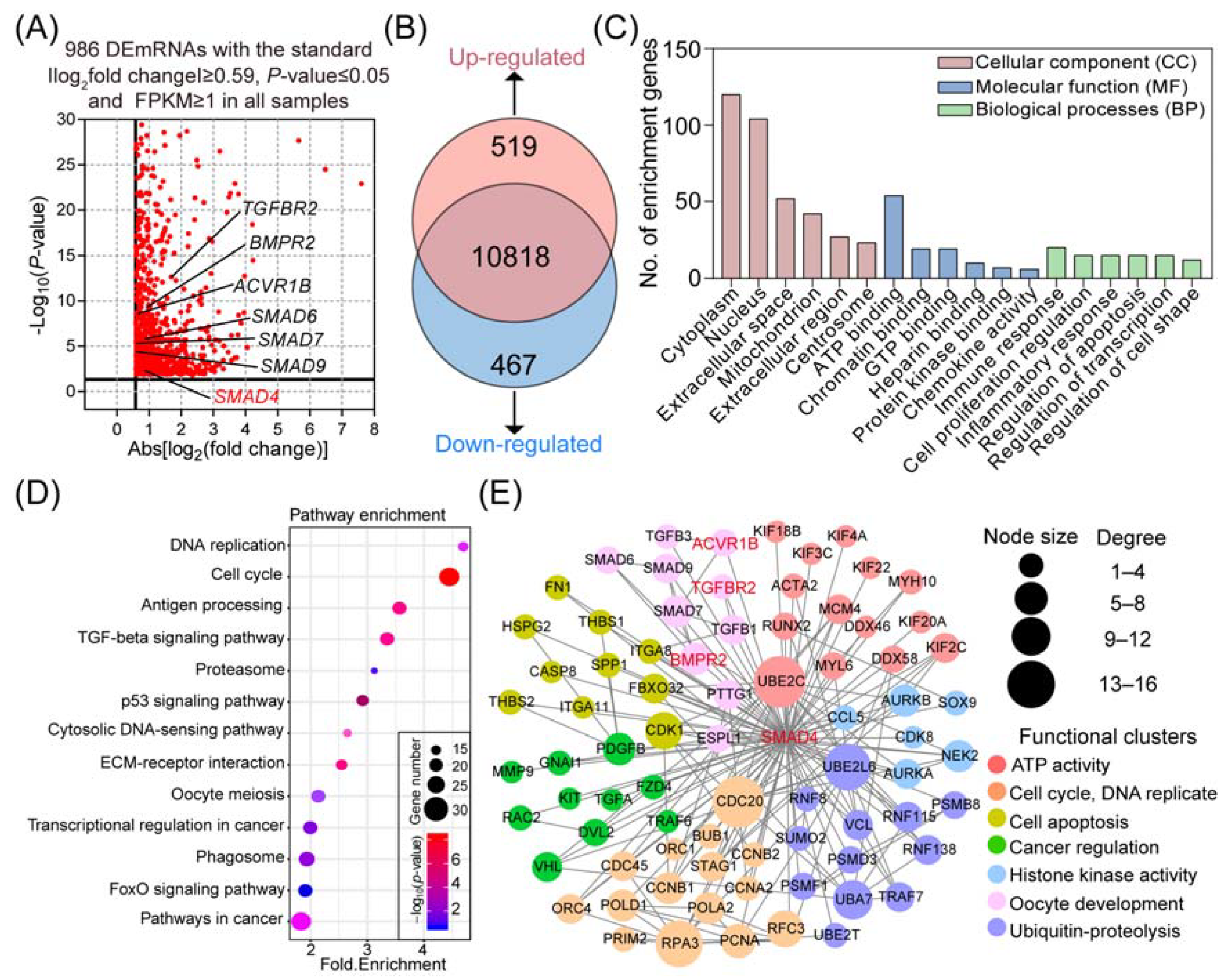
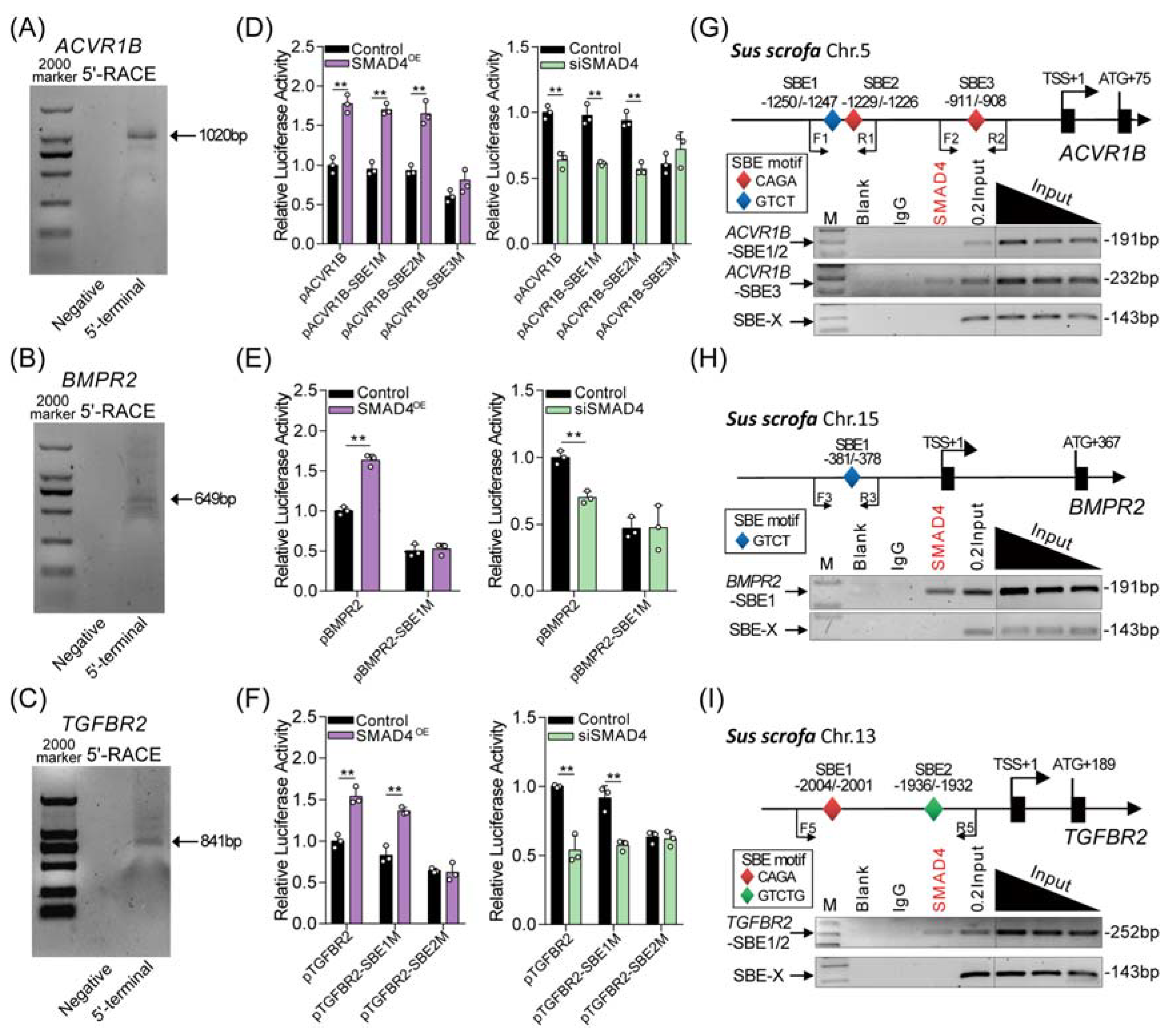
2.2. SMAD4 Is a Strong Inducer for Its Upstream Receptors
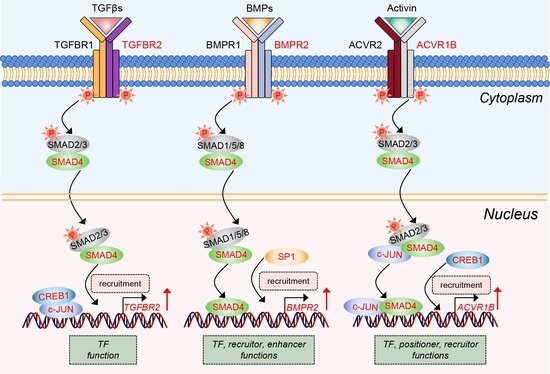
2.3. SMAD4 Feedback Regulates Its Upstream Receptors by Acting as a Transcription Factor
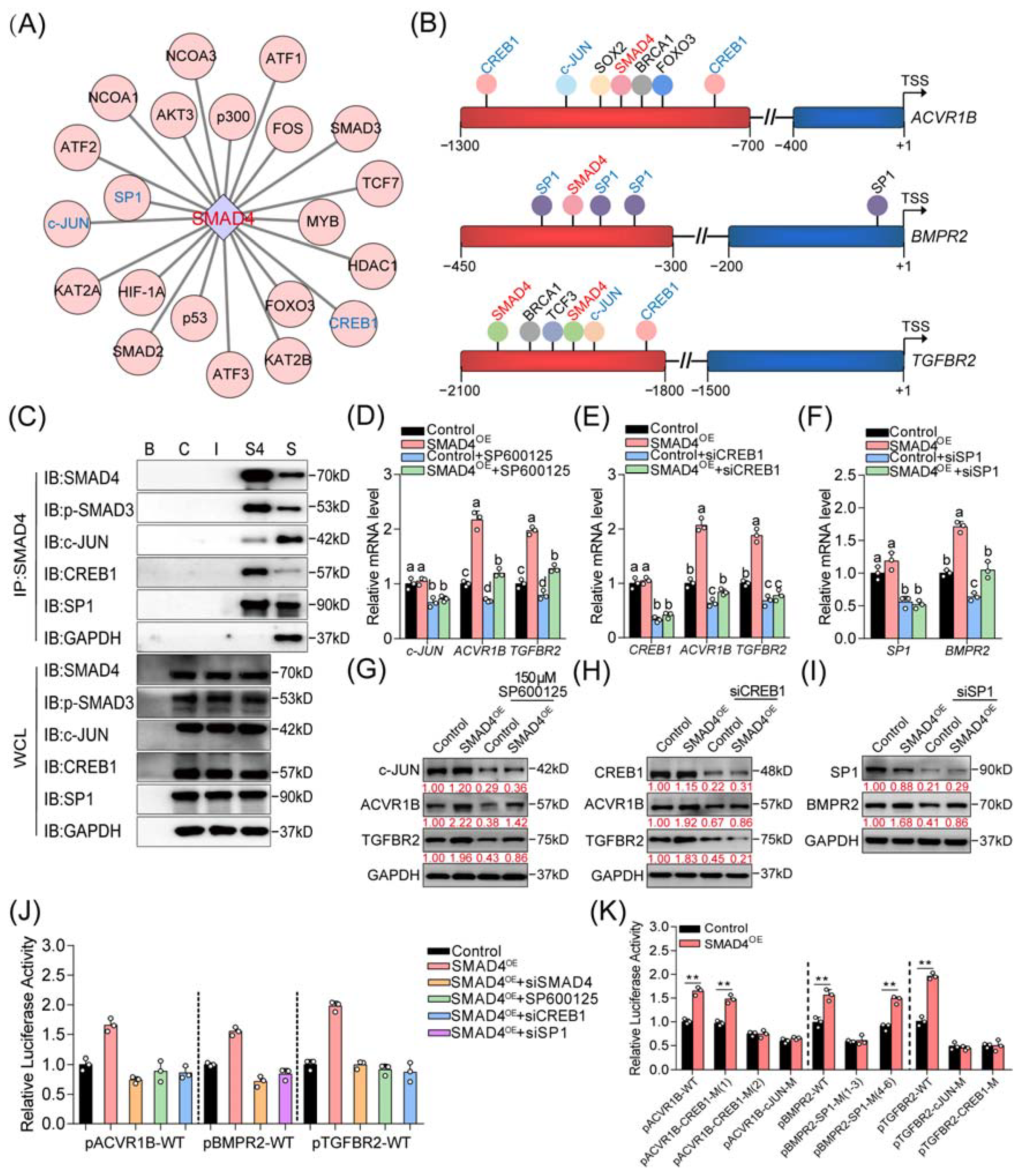
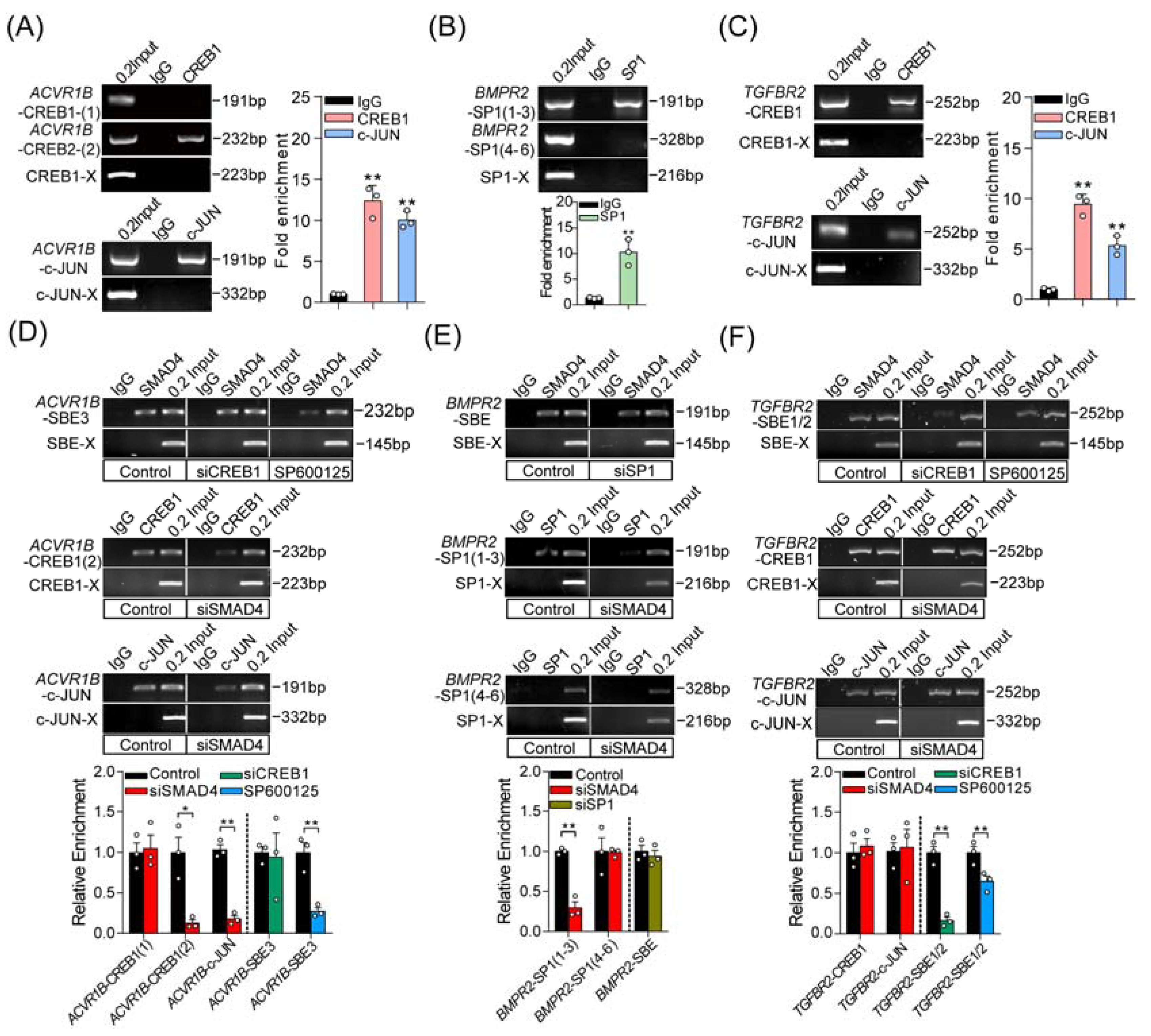
2.4. Three Coactivators (CREB1, c-JUN, and SP1) Are Essential for SMAD4-Mediated Feedback Regulation of TGF-β Family Signaling Pathways
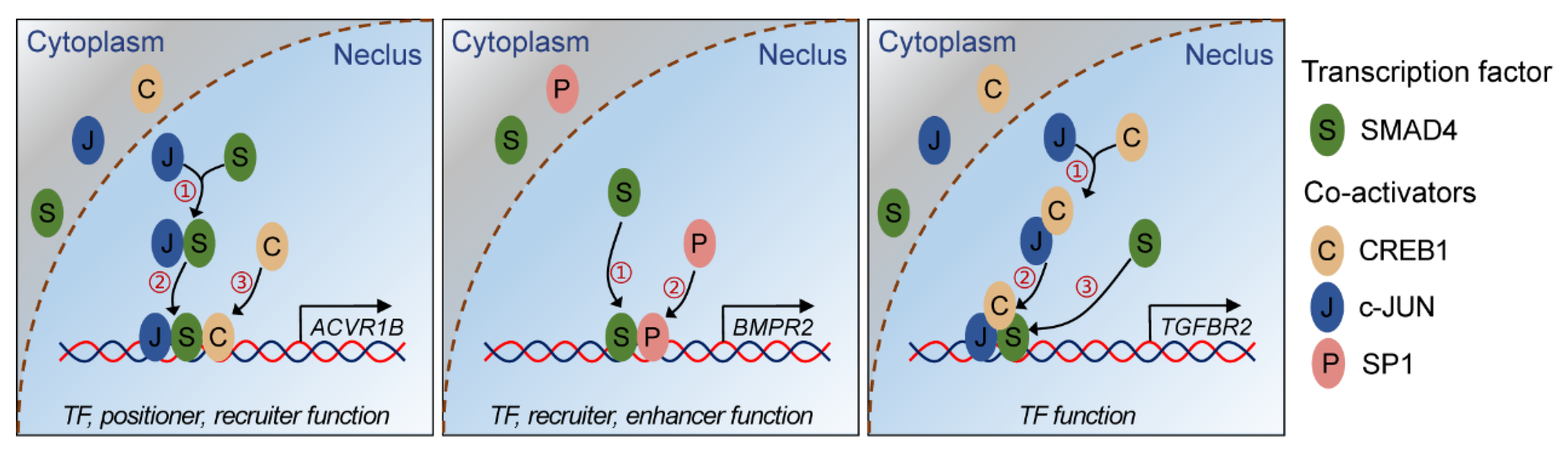
2.5. SMAD4 Interacts with Coactivators in Different Modes during Feedback Regulation Process
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals
4.3. Cell Culture and Treatment
4.4. RNA Isolation and Quantitative Real-Time PCR (qRT-PCR)
4.5. Rapid Amplification of cDNA Ends (RACE)
4.6. Remapping on Pig Reference Genome and Data Reanalysis
4.7. Bioinformatics Analysis
4.8. SiRNAs and Inhibitors
4.9. Plasmids Construction and Luciferase Activity Assay
4.10. Western Blotting
4.11. Immunoprecipitation (IP) and Chromatin Immunoprecipitation (ChIP)
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ChIP | chromatin immunoprecipitation |
| DEGs | differentially expressed genes |
| FBS | fetal bovine serum |
| FC | fold change |
| GC | granulosa cell |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| IP | immunoprecipitation |
| PPI | protein–protein interaction |
| RACE | rapid amplification of cDNA ends |
| RPKM | reads per kilobase transcriptome per million mapped reads |
| RNA-seq | high-throughput sequencing of RNA |
| SBE | SMAD4 binding element |
| siRNA | small interfering RNA |
| TF | transcription factor |
| TSS | transcription start site |
References
- Antebi, Y.E.; Linton, J.M.; Klumpe, H.; Bintu, B.; Gong, M.; Su, C.; McCardell, R.; Elowitz, M.B. Combinatorial signal perception in the BMP pathway. Cell 2017, 170, 1184–1196. [Google Scholar] [CrossRef] [PubMed]
- Grafe, I.; Alexander, S.; Peterson, J.R.; Snider, T.N.; Levi, B.; Lee, B.; Mishina, Y. TGF-beta family signaling in mesenchymal differentiation. Cold Spring Harb. Perspect. Biol. 2018, 10, a022202. [Google Scholar] [CrossRef] [PubMed]
- Seoane, J.; Gomis, R.R. TGF-beta family signaling in tumor suppression and cancer progression. Cold Spring Harb. Perspect. Biol. 2017, 9, a022277. [Google Scholar] [CrossRef] [PubMed]
- Tracy Cai, X.; Li, H.; Safyan, A.; Gawlik, J.; Pyrowolakis, G.; Jasper, H. AWD regulates timed activation of BMP signaling in intestinal stem cells to maintain tissue homeostasis. Nat. Commun. 2019, 10, 2988. [Google Scholar] [CrossRef]
- Yan, X.; Xiong, X.; Chen, Y.G. Feedback regulation of TGF-beta signaling. Acta Biochim. Biophys. Sin. (Shanghai) 2018, 50, 37–50. [Google Scholar] [CrossRef]
- Wang, L.; Jiang, P.; He, Y.; Hu, H.; Guo, Y.; Liu, X.; Qiu, H.; Ma, Q.; Ouyang, F. A novel mechanism of Smads/miR-675/TGFbetaR1 axis modulating the proliferation and remodeling of mouse cardiac fibroblasts. J. Cell. Physiol. 2019, 234, 20275–20285. [Google Scholar] [CrossRef]
- Du, X.; Pan, Z.; Li, Q.; Liu, H.; Li, Q. SMAD4 feedback regulates the canonical TGF-beta signaling pathway to control granulosa cell apoptosis. Cell Death Dis. 2018, 9, 151. [Google Scholar] [CrossRef]
- Sun, Q.; Mao, S.; Li, H.; Zen, K.; Zhang, C.; Li, L. Role of miR-17 family in the negative feedback loop of bone morphogenetic protein signaling in neuron. PLoS ONE 2013, 8, e83067. [Google Scholar]
- Sun, Y.; Wang, H.; Li, Y.; Liu, S.; Chen, J.; Ying, H. miR-24 and miR-122 negatively regulate the transforming growth factor-beta/Smad signaling pathway in skeletal muscle fibrosis. Mol. Ther. Nucleic Acids 2018, 11, 528–537. [Google Scholar] [CrossRef]
- Hu, J.; Zhang, L.; Mei, Z.; Jiang, Y.; Yi, Y.; Liu, L.; Meng, Y.; Zhou, L.; Zeng, J.; Wu, H.; et al. Interaction of E3 ubiquitin ligase MARCH7 with long noncoding rNA MALAT1 and autophagy-related protein ATG7 promotes autophagy and invasion in ovarian cancer. Cell. Physiol. Biochem. 2018, 47, 654–666. [Google Scholar] [CrossRef]
- Calvo-Sanchez, M.I.; Fernandez-Martos, S.; Carrasco, E.; Moreno-Bueno, G.; Bernabeu, C.; Quintanilla, M.; Espada, J. A role for the Tgf-beta/Bmp co-receptor Endoglin in the molecular oscillator that regulates the hair follicle cycle. J. Mol. Cell Biol. 2019, 11, 39–52. [Google Scholar] [CrossRef]
- Caja, L.; Tzavlaki, K.; Dadras, M.S.; Tan, E.J.; Hatem, G.; Maturi, N.P.; Moren, A.; Wik, L.; Watanabe, Y.; Savary, K.; et al. Snail regulates BMP and TGFbeta pathways to control the differentiation status of glioma-initiating cells. Oncogene 2018, 37, 2515–2531. [Google Scholar] [CrossRef]
- Rojas, A.; Zhang, P.; Wang, Y.; Foo, W.C.; Munoz, N.M.; Xiao, L.; Wang, J.; Gores, G.J.; Hung, M.C.; Blechacz, B. A positive TGF-beta/c-KIT feedback loop drives tumor progression in advanced primary liver cancer. Neoplasia 2016, 18, 371–386. [Google Scholar] [CrossRef]
- Tumelty, K.E.; Higginson-Scott, N.; Fan, X.; Bajaj, P.; Knowlton, K.M.; Shamashkin, M.; Coyle, A.J.; Lu, W.; Berasi, S.P. Identification of direct negative cross-talk between the SLIT2 and bone morphogenetic protein-Gremlin signaling pathways. J. Biol. Chem. 2018, 293, 3039–3055. [Google Scholar] [CrossRef]
- Wu, D.; Huang, D.; Li, L.; Ni, P.; Li, X.; Wang, B.; Han, Y.; Shao, X.; Zhao, D.; Chu, W.; et al. TGF-beta1-PML SUMOylation-peptidyl-prolyl cis-trans isomerase NIMA-interacting 1 (Pin1) form a positive feedback loop to regulate cardiac fibrosis. J. Cell. Physiol. 2019, 234, 6263–6273. [Google Scholar] [CrossRef]
- Xu, J.; Yang, Y.; Xie, R.; Liu, J.; Nie, X.; An, J.; Wen, G.; Liu, X.; Jin, H.; Tuo, B. The NCX1/TRPC6 complex mediates TGFbeta-driven migration and invasion of human hepatocellular carcinoma cells. Cancer Res. 2018, 78, 2564–2576. [Google Scholar] [CrossRef]
- Zheng, D.; Zhang, Y.; Hu, Y.; Guan, J.; Xu, L.; Xiao, W.; Zhong, Q.; Ren, C.; Lu, J.; Liang, J.; et al. Long noncoding RNA Crnde attenuates cardiac fibrosis via Smad3-Crnde negative feedback in diabetic cardiomyopathy. FEBS J. 2019, 286, 1645–1655. [Google Scholar] [CrossRef]
- Wan, C.; Yuan, G.; Luo, D.; Zhang, L.; Lin, H.; Liu, H.; Chen, L.; Yang, G.; Chen, S.; Chen, Z. The dentin sialoprotein (dsp) domain regulates dental mesenchymal cell differentiation through a novel surface receptor. Sci. Rep. 2016, 6, 29666. [Google Scholar] [CrossRef]
- Chen, L.; Toke, N.H.; Luo, S.; Vasoya, R.P.; Fullem, R.L.; Parthasarathy, A.; Perekatt, A.O.; Verzi, M.P. A reinforcing HNF4-SMAD4 feed-forward module stabilizes enterocyte identity. Nat. Genet. 2019, 51, 777–785. [Google Scholar] [CrossRef]
- Miyazawa, K.; Miyazono, K. Regulation of TGF-beta family signaling by inhibitory smads. Cold Spring Harb. Perspect. Biol. 2017, 9, a022095. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Pan, Z.; Du, X.; Zhang, J.; Li, Q. miR-181b-induced SMAD7 downregulation controls granulosa cell apoptosis through TGF-beta signaling by interacting with the TGFBR1 promoter. J. Cell. Physiol. 2018, 233, 6807–6821. [Google Scholar] [CrossRef]
- Zhang, L.; Du, X.; Wei, S.; Li, D.; Li, Q. A comprehensive transcriptomic view on the role of SMAD4 gene by RNAi-mediated knockdown in porcine follicular granulosa cells. Reproduction 2016, 152, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Li, Q.; Yang, L.; Liu, L.; Cao, Q.; Li, Q. SMAD4 activates Wnt signaling pathway to inhibit granulosa cell apoptosis. Cell Death Dis. 2020, 11, 373. [Google Scholar] [CrossRef]
- Wu, Q.; Fukuda, K.; Kato, Y.; Zhou, Z.; Deng, C.; Saga, Y. Sexual fate change of XX germ cells caused by the deletion of SMAD4 and STRA8 independent of somatic sex reprogramming. PLoS Biol. 2016, 14, e1002553. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ju, D.; Zhang, D.; Li, H.; Kong, L.; Guo, Y.; Li, C.; Wang, X.; Chen, Z.; Bian, H. Activation of TGF-beta1-CD147 positive feedback loop in hepatic stellate cells promotes liver fibrosis. Sci. Rep. 2015, 5, 16552. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Jin, E.; Cheng, X.; Feng, S.; Shang, X.; Deng, P.; Jiang, S.; Chang, Q.; Rahmy, S.; Zhao, R.; et al. Opposing roles of TGFbeta and BMP signaling in prostate cancer development. Genes Dev. 2017, 31, 2337–2342. [Google Scholar] [CrossRef]
- Cunha, S.I.; Magnusson, P.U.; Dejana, E.; Lampugnani, M.G. Deregulated TGF-beta/BMP signaling in vascular malformations. Circ. Res. 2017, 121, 981–999. [Google Scholar] [CrossRef] [PubMed]
- Berg, M.; Monnin, D.; Cho, J.; Nelson, L.; Crits-Christoph, A.; Shapira, M. TGFbeta/BMP immune signaling affects abundance and function of C. elegans gut commensals. Nat. Commun. 2019, 10, 604. [Google Scholar] [CrossRef]
- Wang, W.; Chun, H.; Baek, J.; Sadik, J.E.; Shirazyan, A.; Razavi, P.; Lopez, N.; Lyons, K.M. The TGFbeta type I receptor TGFbetaRI functions as an inhibitor of BMP signaling in cartilage. Proc. Natl. Acad. Sci. USA 2019, 116, 15570–15579. [Google Scholar] [CrossRef]
- Hiepen, C.; Jatzlau, J.; Hildebrandt, S.; Kampfrath, B.; Goktas, M.; Murgai, A.; Cuellar Camacho, J.L.; Haag, R.; Ruppert, C.; Sengle, G.; et al. BMPR2 acts as a gatekeeper to protect endothelial cells from increased TGFbeta responses and altered cell mechanics. PLoS Biol. 2019, 17, e3000557. [Google Scholar] [CrossRef]
- Yang, L.; Du, X.; Liu, L.; Cao, Q.; Pan, Z.; Li, Q. miR-1306 mediates the feedback regulation of the TGF-beta/SMAD signaling pathway in granulosa cells. Cells 2019, 8, 298. [Google Scholar] [CrossRef]
- Knight, P.G.; Glister, C. TGF-beta superfamily members and ovarian follicle development. Reproduction 2006, 132, 191–206. [Google Scholar] [CrossRef]
- Abdurahman, A.; Du, X.; Yao, Y.; Sulaiman, Y.; Aniwashi, J.; Li, Q. Smad4 feedback enhances BMPR1B transcription in ovine granulosa cells. Int. J. Mol. Sci. 2019, 20, 2732. [Google Scholar] [CrossRef]
- Heldin, C.H.; Miyazono, K.; ten Dijke, P. TGF-beta signalling from cell membrane to nucleus through SMAD proteins. Nature 1997, 390, 465–471. [Google Scholar] [CrossRef]
- Ishida, W.; Hamamoto, T.; Kusanagi, K.; Yagi, K.; Kawabata, M.; Takehara, K.; Sampath, T.K.; Kato, M.; Miyazono, K. Smad6 is a Smad1/5-induced smad inhibitor. Characterization of bone morphogenetic protein-responsive element in the mouse Smad6 promoter. J. Biol. Chem. 2000, 275, 6075–6079. [Google Scholar] [CrossRef] [PubMed]
- Nakao, A.; Afrakhte, M.; Moren, A.; Nakayama, T.; Christian, J.L.; Heuchel, R.; Itoh, S.; Kawabata, M.; Heldin, N.E.; Heldin, C.H.; et al. Identification of Smad7, a TGFbeta-inducible antagonist of TGF-beta signalling. Nature 1997, 389, 631–635. [Google Scholar] [CrossRef]
- Liu, S.; Long, J.; Yuan, B.; Zheng, M.; Xiao, M.; Xu, J.; Lin, X.; Feng, X.H. SUMO modification reverses inhibitory effects of Smad nuclear interacting protein-1 in TGF-beta responses. J. Biol. Chem. 2016, 291, 24418–24430. [Google Scholar] [CrossRef]
- Tecalco-Cruz, A.C.; Rios-Lopez, D.G.; Vazquez-Victorio, G.; Rosales-Alvarez, R.E.; Macias-Silva, M. Transcriptional cofactors Ski and SnoN are major regulators of the TGF-beta/Smad signaling pathway in health and disease. Signal. Transduct. Target. Ther. 2018, 3, 15. [Google Scholar] [CrossRef]
- Zhang, S.; Takaku, M.; Zou, L.; Gu, A.; Chou, W.; Zhang, G.; Wu, B.; Kong, Q.; Thomas, S.Y.; Serody, J.S.; et al. Reversing SKI-SMAD4-mediated suppression is essential for TH17 cell differentiation. Nature 2017, 551, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chu, J.; Yi, P.; Dong, W.; Saultz, J.; Wang, H.; Scoville, S.; Zhang, J.; Wu, L.; Deng, Y.; et al. SMAD4 promotes TGF-beta-independent NK cell homeostasis and maturation and antitumor immunity. J. Clin. Investig. 2018, 128, 5123–5136. [Google Scholar] [CrossRef]
- Light, A.; Hammes, S.R. LH-induced steroidogenesis in the mouse ovary, but not testis, requires matrix metalloproteinase 2- and 9-mediated cleavage of upregulated EGF receptor ligands. Biol. Reprod. 2015, 93, 65. [Google Scholar] [CrossRef]
- Hamil, K.G.; Conti, M.; Shimasaki, S.; Hall, S.H. Follicle-stimulating hormone regulation of AP-1: Inhibition of c-jun and stimulation of jun-B gene transcription in the rat Sertoli cell. Mol. Cell Endocrinl. 1994, 99, 269–277. [Google Scholar] [CrossRef]
- Pogrmic-Majkic, K.; Samardzija, D.; Fa, S.; Hrubik, J.; Glisic, B.; Kaisarevic, S.; Andric, N. Atrazine enhances progesterone production through activation of multiple signaling pathways in FSH-stimulated rat granulosa cells: Evidence for premature luteinization. Biol. Reprod. 2014, 91, 124. [Google Scholar] [CrossRef]
- Chu, Y.L.; Xu, Y.R.; Yang, W.X.; Sun, Y. The role of FSH and TGF-beta superfamily in follicle atresia. Aging 2018, 10, 305–321. [Google Scholar] [CrossRef]
- Ongaro, L.; Schang, G.; Ho, C.C.; Zhou, X.; Bernard, D.J. TGF-beta superfamily regulation of follicle-stimulating hormone synthesis by gonadotrope cells: Is there a role for bone morphogenetic proteins? Endocrinology 2019, 160, 675–683. [Google Scholar] [CrossRef]
- Fortin, J.; Boehm, U.; Deng, C.X.; Treier, M.; Bernard, D.J. Follicle-stimulating hormone synthesis and fertility depend on SMAD4 and FOXL2. FASEB J. 2014, 28, 3396–3410. [Google Scholar] [CrossRef]
- Wang, W.; Chen, X.; Li, X.; Wang, L.; Zhang, H.; He, Y.; Wang, J.; Zhao, Y.; Zhang, B.; Xu, Y. Interference RNA-based silencing of endogenous SMAD4 in porcine granulosa cells resulted in decreased FSH-mediated granulosa cells proliferation and steroidogenesis. Reproduction 2011, 141, 643–651. [Google Scholar] [CrossRef]
- Du, X.; Zhang, L.; Li, X.; Pan, Z.; Liu, H.; Li, Q. TGF-beta signaling controls FSHR signaling-reduced ovarian granulosa cell apoptosis through the SMAD4/miR-143 axis. Cell Death Dis. 2016, 7, e2476. [Google Scholar] [CrossRef]
- Du, X.; Li, Q.; Pan, Z. Androgen receptor and miRNA-126* axis controls follicle-stimulating hormone receptor expression in porcine ovarian granulosa cells. Reproduction 2016, 152, 161–169. [Google Scholar] [CrossRef][Green Version]
- Li, Q.; Du, X.; Liu, L.; Pan, Z.; Cao, S. MiR-126* is a novel functional target of transcription factor SMAD4 in ovarian granulosa cells. Gene 2019, 711, 143953. [Google Scholar] [CrossRef] [PubMed]
- Martin-Malpartida, P.; Batet, M.; Kaczmarska, Z.; Freier, R.; Gomes, T.; Aragon, E.; Zou, Y.; Wang, Q.; Xi, Q.; Ruiz, L.; et al. Structural basis for genome wide recognition of 5-bp GC motifs by SMAD transcription factors. Nat. Commun. 2017, 8, 2070. [Google Scholar] [CrossRef]
- Itoh, Y.; Koinuma, D.; Omata, C.; Ogami, T.; Motizuki, M.; Yaguchi, S.I.; Itoh, T.; Miyake, K.; Tsutsumi, S.; Aburatani, H.; et al. A comparative analysis of Smad-responsive motifs identifies multiple regulatory inputs for TGF-beta transcriptional activation. J. Biol. Chem. 2019, 294, 15466–15479. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Liu, L.; Wu, W.; Li, P.; Pan, Z.; Zhang, L.; Liu, J.; Li, Q. SMARCA2 is regulated by NORFA/miR-29c, a novel pathway related to female fertility, controls granulosa cell apoptosis. J. Cell Sci. 2020, 133, jcs249961. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, X.; Yao, Y.; Li, Q.; Pan, Z. miR-1275 controls granulosa cell apoptosis and estradiol synthesis by impairing LRH-1/CYP19A1 axis. Biochim. Biophys. Acta 2018, 1861, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Liu, L.; Li, Q.; Zhang, L.; Pan, Z.; Li, Q. NORFA, long intergenic noncoding RNA, maintains sow fertility by inhibiting granulosa cell death. Commun. Biol. 2020, 3, 131. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Li, Q.; Yang, L.; Li, Q.; Du, X. SMAD4 Feedback Activates the Canonical TGF-β Family Signaling Pathways. Int. J. Mol. Sci. 2021, 22, 10024. https://doi.org/10.3390/ijms221810024
Liu L, Li Q, Yang L, Li Q, Du X. SMAD4 Feedback Activates the Canonical TGF-β Family Signaling Pathways. International Journal of Molecular Sciences. 2021; 22(18):10024. https://doi.org/10.3390/ijms221810024
Chicago/Turabian StyleLiu, Lu, Qiqi Li, Liu Yang, Qifa Li, and Xing Du. 2021. "SMAD4 Feedback Activates the Canonical TGF-β Family Signaling Pathways" International Journal of Molecular Sciences 22, no. 18: 10024. https://doi.org/10.3390/ijms221810024
APA StyleLiu, L., Li, Q., Yang, L., Li, Q., & Du, X. (2021). SMAD4 Feedback Activates the Canonical TGF-β Family Signaling Pathways. International Journal of Molecular Sciences, 22(18), 10024. https://doi.org/10.3390/ijms221810024