
The CXC Chemokine Receptors in Four-Eyed Sleeper (Bostrychus sinensis) and Their Involvement in Responding to Skin Injury


 , and
, and
Abstract
:1. Introduction
2. Results
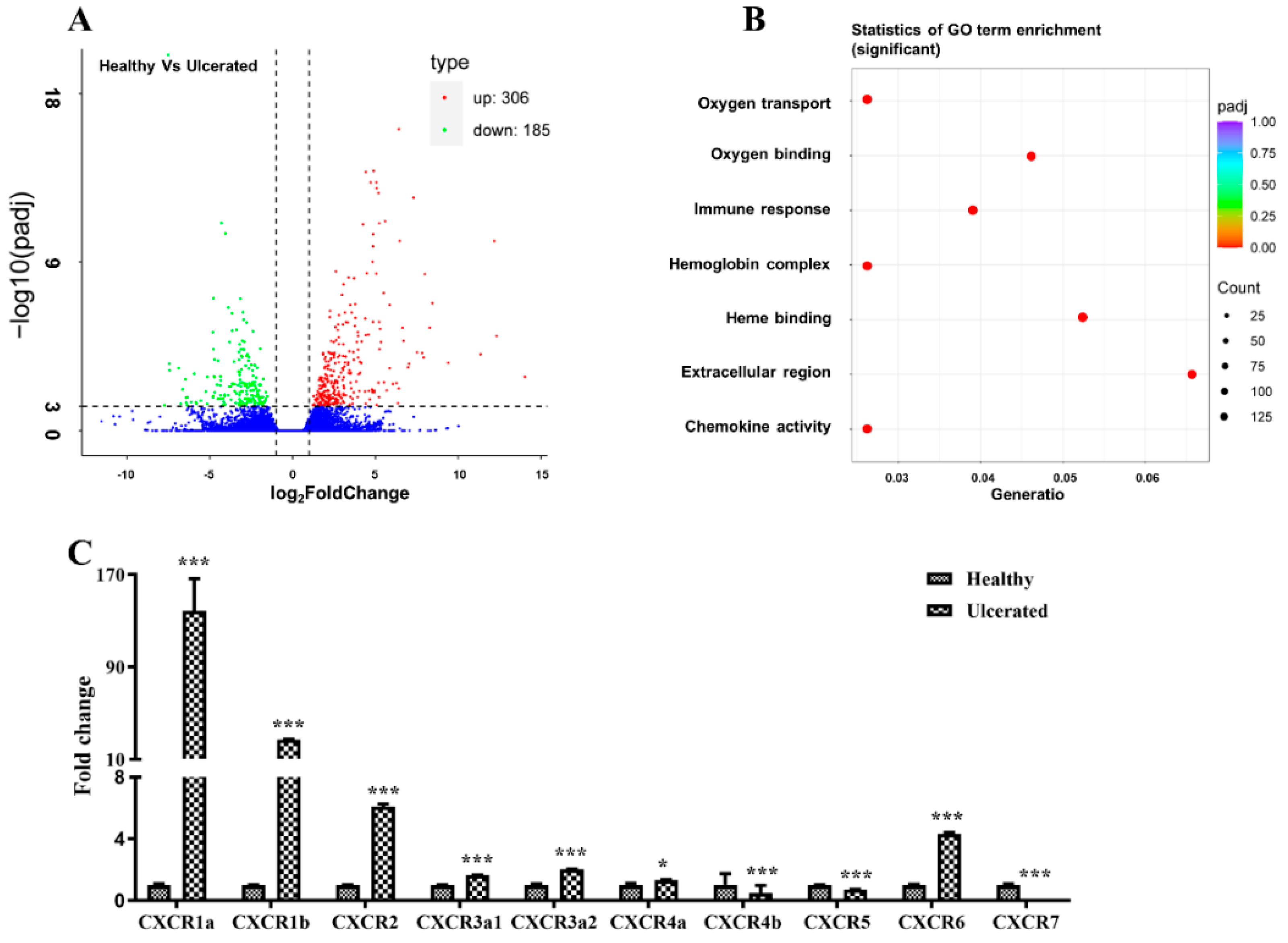
2.1. Transcriptome Analysis of the Healthy and Skin Ulcerated Fish
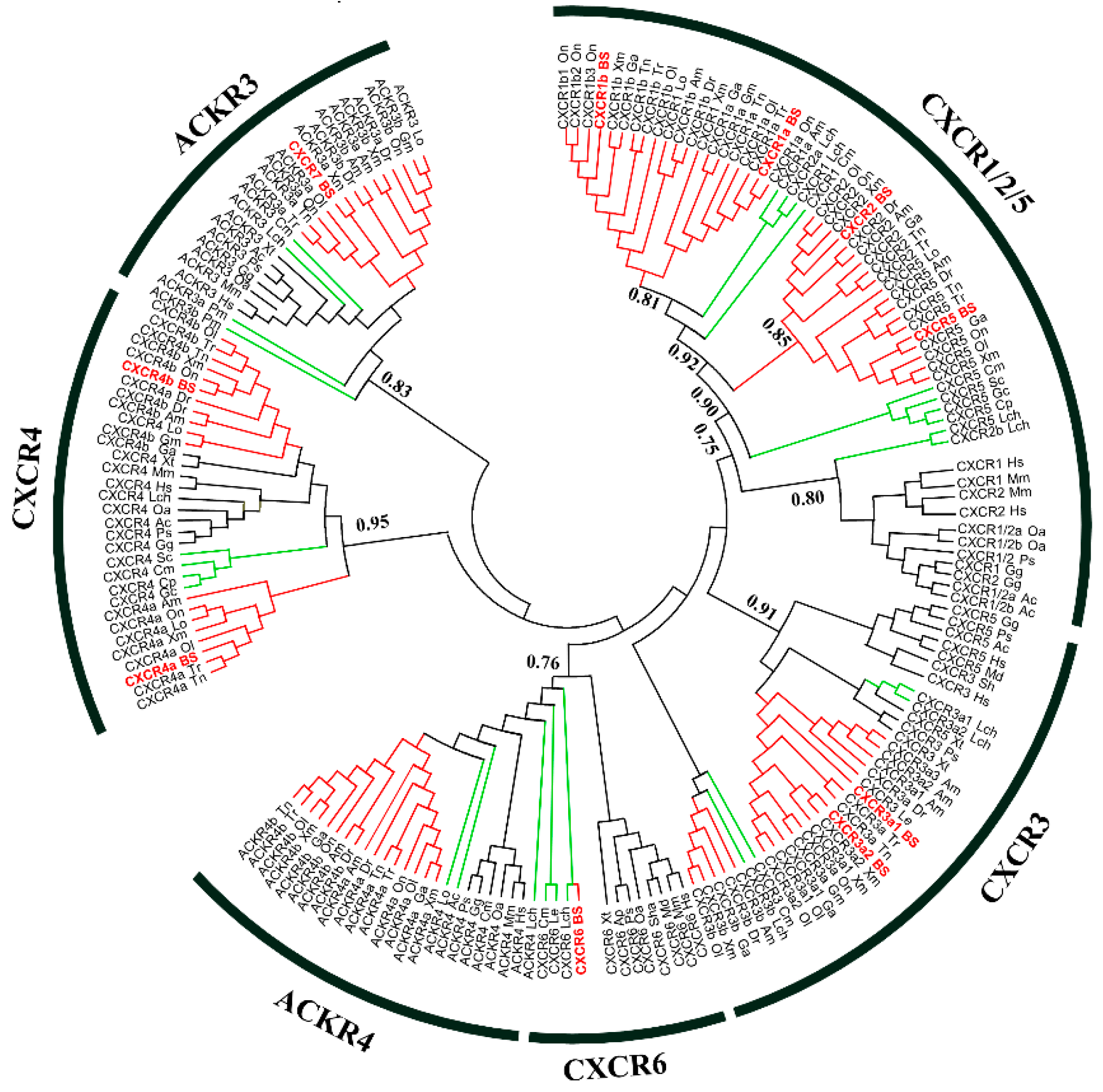
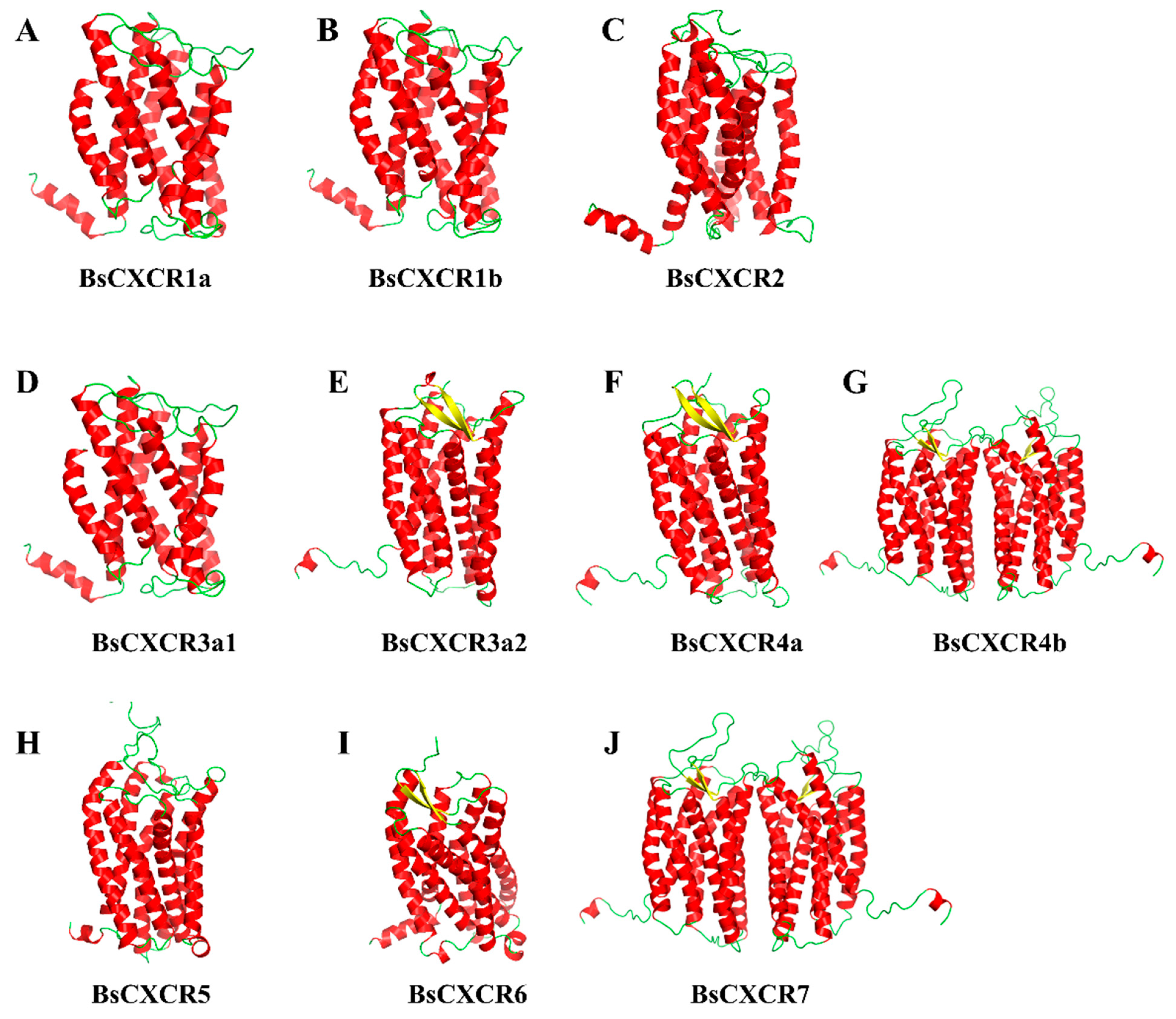
2.2. Sequence Analysis of CXCRs
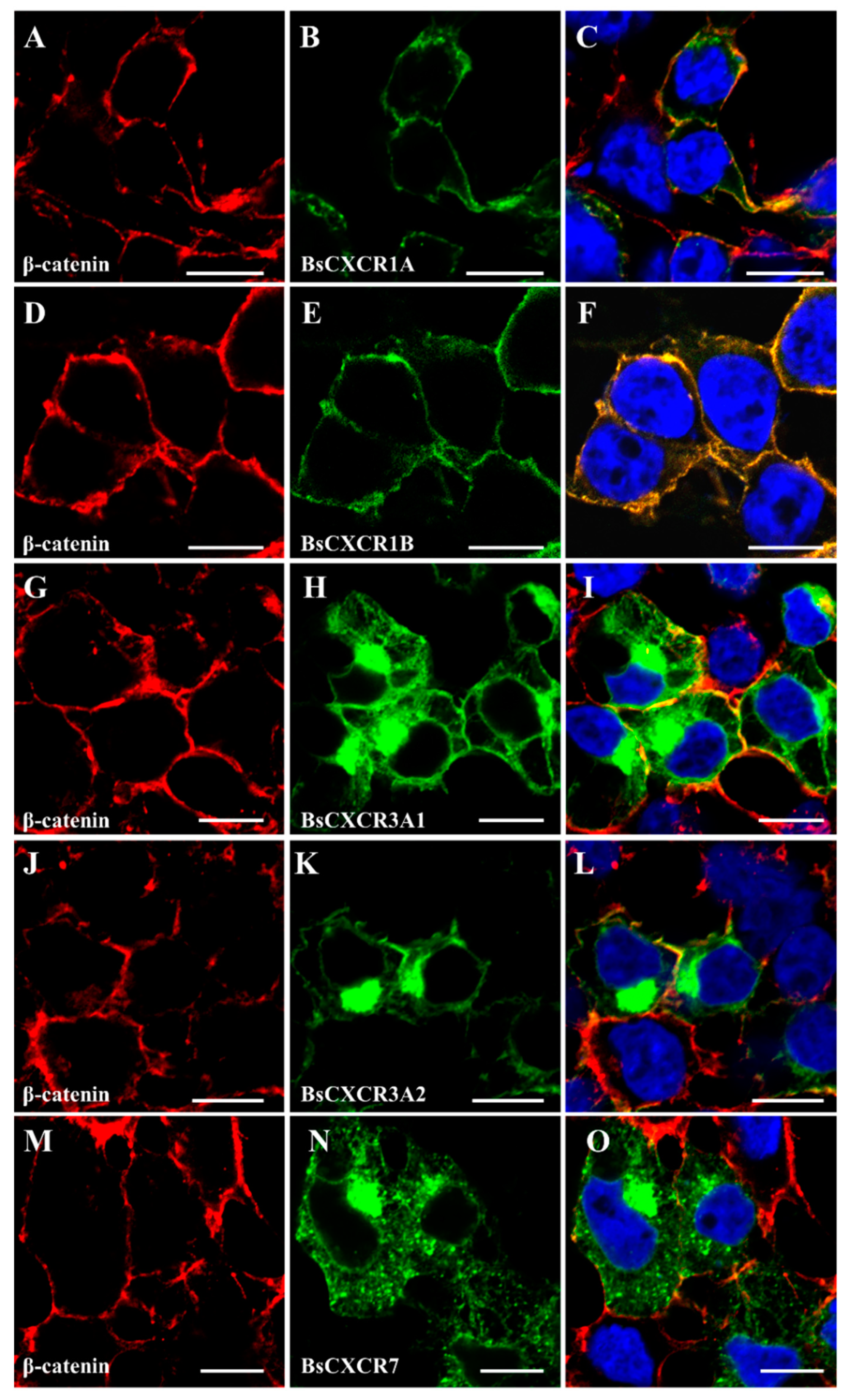
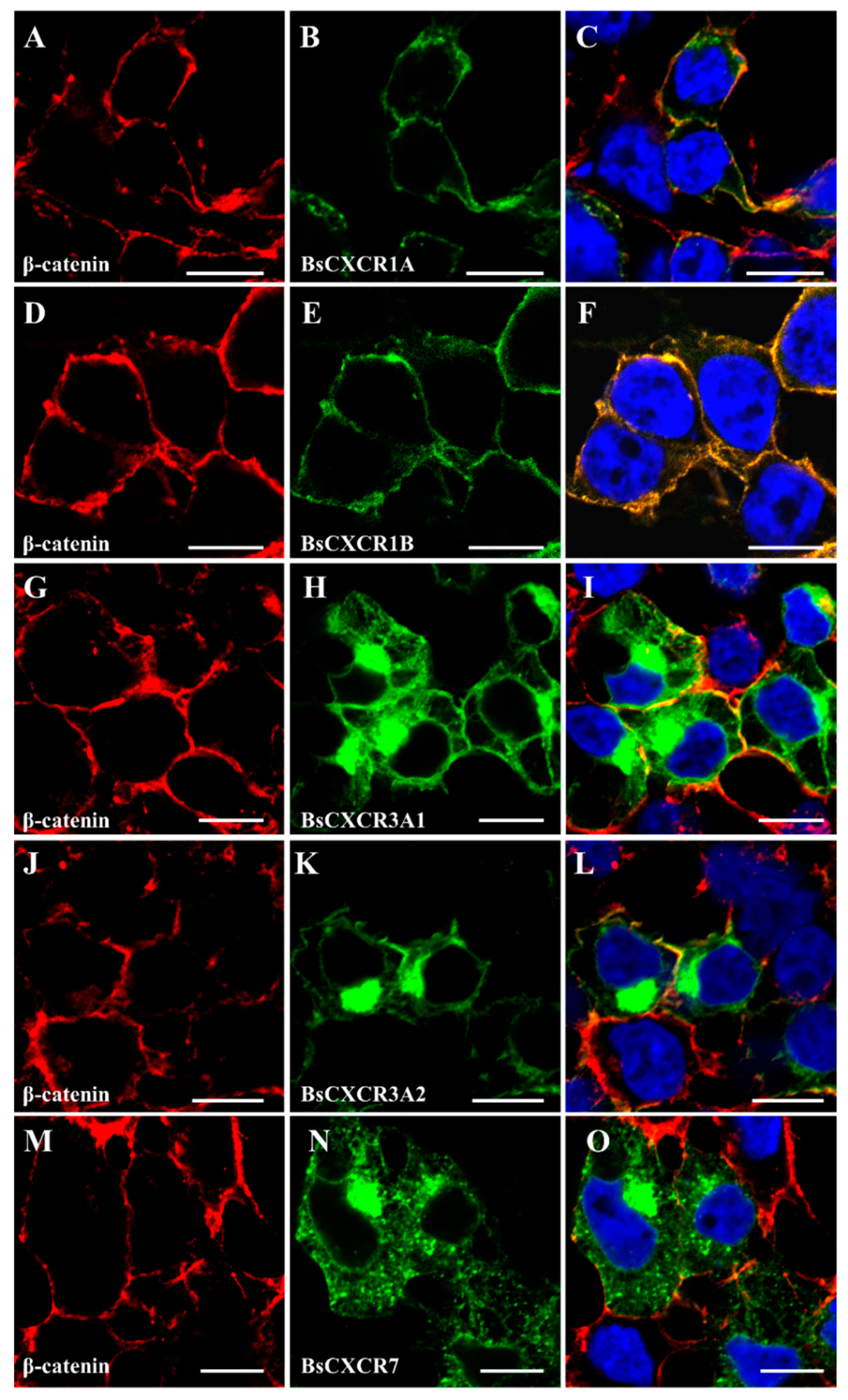
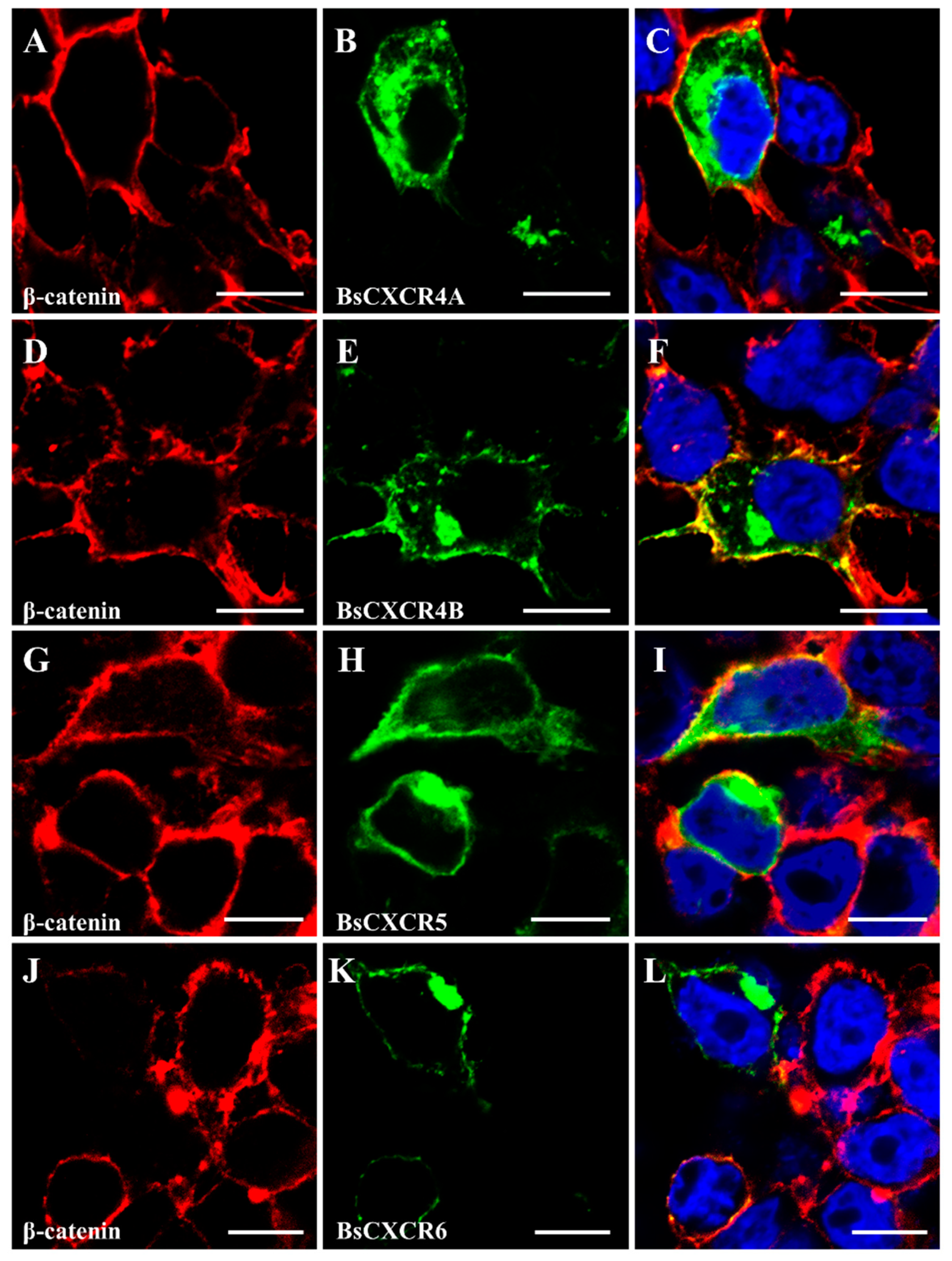
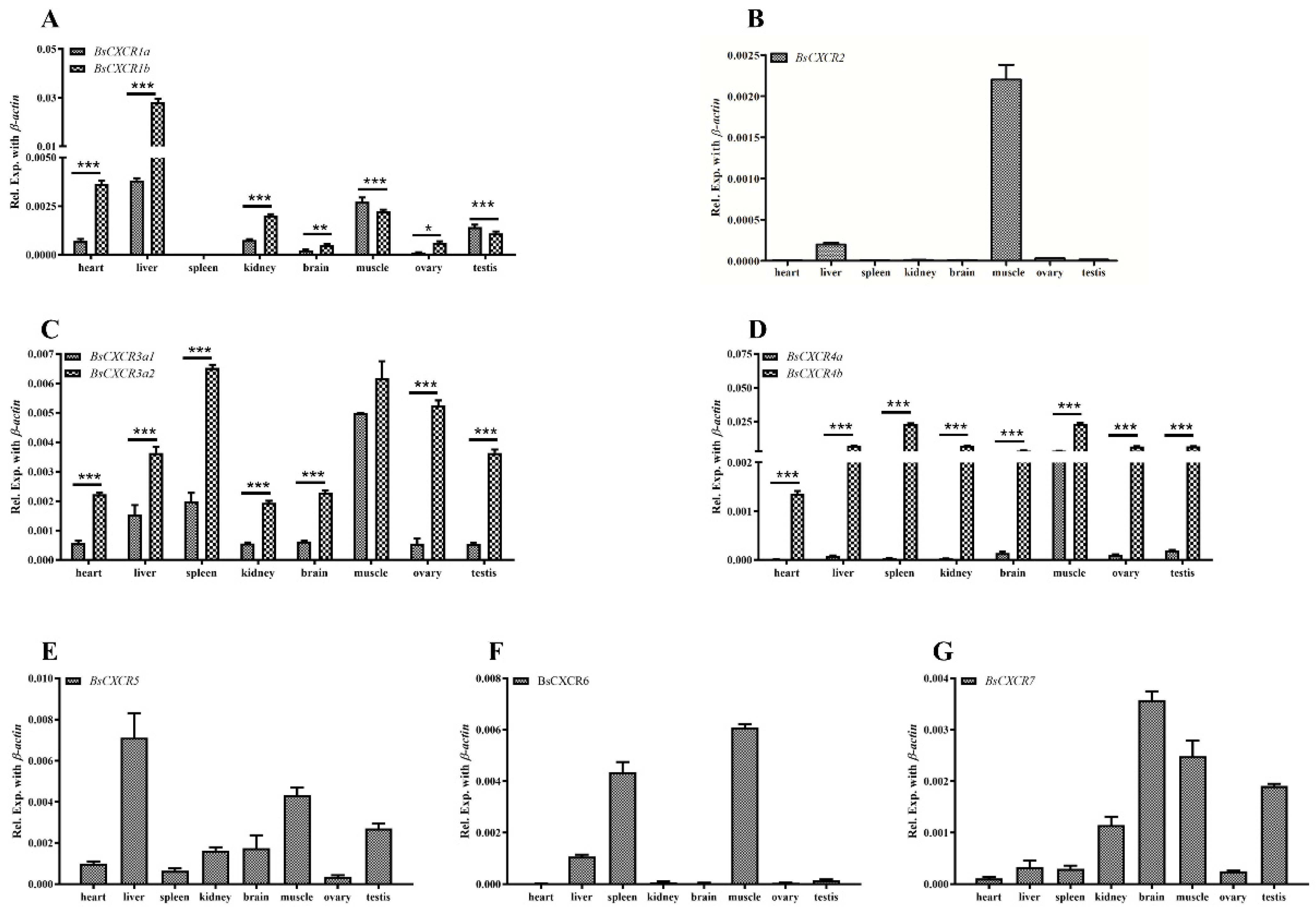
2.3. Cellular Localizations and Tissue Distributions of BsCXCRs
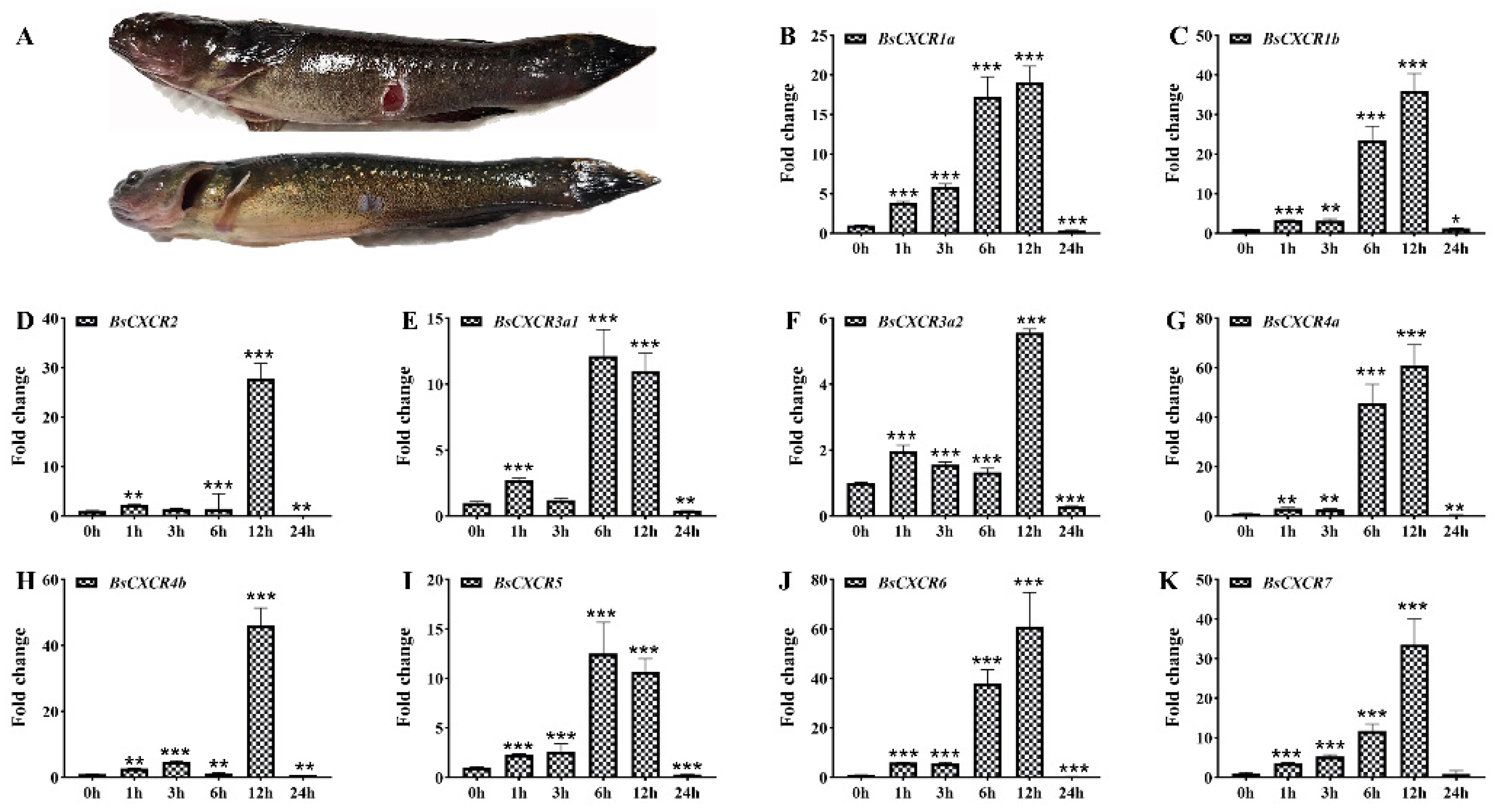
2.4. Dynamic Expression of BsCXCR Transcripts in Response to Skin Injury
3. Discussion
4. Materials and Methods
4.1. Ethical Procedures
4.2. Fish, Cells and Sample Collection
4.3. Total RNA Isolation and Illumine Sequencing
4.4. TranscriptomeSsequencing and Analysis
4.5. Bioinformatic Analysis of BsCXCR Family
4.6. Quantitative PCR (qPCR)
4.7. Plasmid Construction
4.8. Immunofluorescence
4.9. Mechanical Skin Injury
4.10. Statistics Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CXCR | CXC chemokine receptor |
| CCR | CC chemokine receptor |
| CX3CR | CX3C chemokine receptor |
| DAPI | 4′,6-diamidino-2-phenylindole |
| DEGs | Differentially expressed genes |
| DMEM | Dulbecco’s modified eagle medium |
| ECL | Extracellular loop |
| GO | Gene ontology |
| ICL | Intracellular loop |
| KEGG | Kyoto encyclopedia of genes and genomes |
| KOG | Eukaryotic ortholog groups |
| ORF | Open Reading Frame |
| PBS | Phosphate buffered saline |
| qPCR | Quantitative polymerase chain reaction |
| RT | Room temperature |
| TM | Transmembrane domain |
| XCR | XC chemokine receptor |
| CXCL | CXC chemokine ligand |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) | Comments |
|---|---|---|
| BsCXCR1a-F | ACCGAATTCATGTCAGATGAAATCACCTCGTCTTTTAT | Plamid construction |
| BsCXCR1a-R | CGCGGATCCCATAAATGTCGATGTGATCTCTGAAGAC | |
| BsCXCR1b-F | ACCGAATTCATGTCTTTGTCTTTCGACTTCAGT | |
| BsCXCR1b-R | ACCGGATCCGAGAACTGTCCCATTGCTTTCTGAGG | |
| BsCXCR3a1-F | ACCGAATTCTCCTCAGTAATTCACCTGTTACT | |
| BsCXCR3a1-R | ACCGGATCCGATGGCAATGGAGTTGGAGGTGT | |
| BsCXCR3a2-F | ACCGAATTCTCCTCAGTAATTCACCTGTTACT | |
| BsCXCR3a2-R | ACCGGATCCGATGGCAATGGAGTTGGAGGTGT | |
| BsCXCR4a-F | ACCGCTAGCATGTCTTACTATGAGCACAT | |
| BsCXCR4a-R | CGCGGATCCACTGGAGTGTAGACTGGATGACTCG | |
| BsCXCR4b-F | ACCGCTAGCACTAACGTAACGTAACATCACGA | |
| BsCXCR4b-R | AGCGAATTCGAACTGGACAGCACACTTGAGGATTCCGA | |
| BsCXCR5-F | CTAGCTAGCATGGAGGAATCTGAGAATTATACATAT | |
| BsCXCR5-R | ACCGAATTCGAGAAGGTGTTGGTGGTTGTTGCTCC | |
| BsCXCR6-F | CTAGCTAGCATGACGTCTGTGTACC | |
| BsCXCR6-R | ACCGAATTCGACAAAATATTGCTGCCTGGAAACTTAAA | |
| BsCXCR7-R | ACCGAATTCGGTCAGTGGTCCTCTGTCAGC | |
| BsCXCR7-R | ACCGGATCCGATCTGCGGCGGATTGTCCAC | |
| BsActin-F | GACAGAGCGTGGCTACTCATTCACC | qRT-PCR |
| BsActin-R | GGTTTCATGGATACCGCAGGATTCC | |
| BsCXCR1a RT-F | AAGTGCTGAGGGGAGGAGTGAGGA | |
| BsCXCR1a RT-R | CATGAAATGGTGAAATGGAGAAATGGAG | |
| BsCXCR1b RT-F | CCTGACCATATGCCTCCTCC | |
| BsCXCR1b RT-R | ACAGAGCCATGAGACCATCC | |
| BsCXCR3a1 RT-F | CCTCTGAGGCTGTGAGTGGA | |
| BsCXCR3a1 RT-R | AGAACCACACCAGCAAGCAG | |
| BsCXCR3a2 RT-F | GGATGGTGCTGTGGTGTGTT | |
| BsCXCR3a2 RT-R | GCCTGGGTGTCTTCTGGGTA | |
| BsCXCR4a RT-F | CATAGTGTTGGGATGCCAGC | |
| BsCXCR4a RT-R | CAAGTAGCAGCTCCACAACG | |
| BsCXCR4b RT-F | CACGAGCGTTCAACATGGAT | |
| BsCXCR4b RT-R | CGATCTCTCACATGGCTCCT | |
| BsCXCR5 RT-F | GCTGCCTGAACCCTTTTCTC | |
| BsCXCR5 RT-R | AAGGTGTTGGTGGTTGTTGC | |
| BsCXCR6 RT-F | CTCCAAGCCACCCATACTCA | |
| BsCXCR6 RT-R | CACACACCGCAGCTATCTTC | |
| BsCXCR7 RT-F | GTACATTGCCCACTTAGCCG | |
| BsCXCR7 RT-R | GGAAGAAGATGCTGCCGAAG |
References
- Esser, C.; Bargen, I.; Weighardt, H.; Haarmann-Stemmann, T.; Krutmann, J. Functions of the aryl hydrocarbon receptor in the skin. Semin. Immunopathol. 2013, 35, 677–691. [Google Scholar] [CrossRef] [PubMed]
- Singer, A.J.; Clark, R.A. Cutaneous wound healing. N. Engl. J. Med. 1999, 341, 738–746. [Google Scholar] [CrossRef]
- Zaja-Milatovic, S.; Richmond, A. CXC chemokines and their receptors: A case for a significant biological role in cutaneous wound healing. Histol. Histopathol. 2008, 23, 1399–1407. [Google Scholar] [PubMed]
- Gillitzer, R.; Goebeler, M. Chemokines in cutaneous wound healing. J. Leukoc. Biol. 2001, 69, 513–521. [Google Scholar]
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef]
- Aghaallaei, N.; Bajoghli, B.; Schwarz, H.; Schorpp, M.; Boehm, T. Characterization of mononuclear phagocytic cells in medaka fish transgenic for a cxcr3a:gfp reporter. Proc. Natl. Acad. Sci. USA 2010, 107, 18079–18084. [Google Scholar] [CrossRef] [Green Version]
- Vågesjö, E.; Öhnstedt, E.; Mortier, A.; Lofton, H.; Huss, F.; Proost, P.; Roos, S.; Phillipson, M. Accelerated wound healing in mice by on-site production and delivery of CXCL12 by transformed lactic acid bacteria. Proc. Natl. Acad. Sci. USA 2018, 115, 1895–1900. [Google Scholar] [CrossRef] [Green Version]
- Bajoghli, B. Evolution and function of chemokine receptors in the immune system of lower vertebrates. Eur. J. Immunol. 2013, 43, 1686–1692. [Google Scholar] [CrossRef]
- Lu, W.J.; Zhou, L.; Gao, F.X.; Sun, Z.H.; Li, Z.; Liu, X.C.; Li, S.S.; Wang, Y.; Gui, J.F. Divergent expression patterns and function of two cxcr4 paralogs in hermaphroditic Epinephelus coioides. Int. J. Mol. Sci. 2018, 19, 2943. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.; Li, S.; Xu, Q.; Zhu, D.; Zhang, Q.; Luo, K.; Zhang, W. Molecular characterization and expression analysis of Asian swamp eel (Monopterus albus) CXC chemokine receptor (CXCR) 1a, CXCR1b, CXCR2, CXCR3a, CXCR3b, and CXCR4 after bacteria and poly I:C challenge. Fish. Shellfish Immunol. 2019, 84, 572–586. [Google Scholar] [CrossRef] [PubMed]
- Doitsidou, M.; Reichman-Fried, M.; Stebler, J.; Koprunner, M.; Dorries, J.; Meyer, D.; Esguerra, C.V.; Leung, T.; Raz, E. Guidance of primordial germ cell migration by the chemokine SDF-1. Cell 2002, 111, 647–659. [Google Scholar] [CrossRef] [Green Version]
- Badillo, A.T.; Chung, S.; Zhang, L.; Zoltick, P.; Liechty, K.W. Lentiviral gene transfer of SDF-1alpha to wounds improves diabetic wound healing. J. Surg. Res. 2007, 143, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Feng, G.; Hao, D.; Chai, J. Processing of CXCL12 impedes the recruitment of endothelial progenitor cells in diabetic wound healing. FEBS J. 2014, 281, 5054–5062. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Zúñiga, S.; Morales, R.A.; Muñoz-Sánchez, S.; Muñoz-Montecinos, C.; Parada, M.; Tapia, K.; Rubilar, C.; Allende, M.L.; Peña, O.A. CXCL12a/CXCR4b acts to retain neutrophils in caudal hematopoietic tissue and to antagonize recruitment to an injury site in the zebrafish larva. Immunogenetics 2017, 69, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Qiu, F.; Li, H.; Lin, H.; Ding, S.; Miyamoto, M.M. Phylogeography of the inshore fish, Bostrychus sinensis, along the Pacific coastline of China. Mol. Phylogenet. Evol. 2016, 96, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.J.; Lin, H.D.; Zhang, L.Y.; Ding, S.X. Development and characterization of 20 microsatellite mrkers for Chinese back sleeper, Bostrychus sinensis. Int. J. Mol. Sci. 2011, 12, 9570–9575. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Wang, H.; Ding, S. Reproductive biology and annual reproductive cycles of two sympatric lineages of Bostrychus sinensis with a natural habitat on southeastern coast of China. Anim. Reprod. Sci. 2021, 232, 106821. [Google Scholar] [CrossRef]
- Zou, J.; Redmond, A.K.; Qi, Z.; Dooley, H.; Secombes, C.J. The CXC chemokine receptors of fish: Insights into CXCR evolution in the vertebrates. Gen. Comp. Endocrinol. 2015, 215, 117–131. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, S.; Zhang, Y.; Yu, Y.; Zhao, H.; Yang, H.; Zheng, L.; Yang, M.; Qin, Q. The CXC chemokines and CXC chemokine receptors in orange-spotted grouper (Epinephelus coioides) and their expression after Singapore grouper iridovirus infection. Dev. Comp. Immunol. 2019, 90, 10–20. [Google Scholar] [CrossRef]
- Sun, Z.; Qin, Y.; Liu, D.; Wang, B.; Jia, Z.; Wang, J.; Gao, Q.; Zou, J.; Pang, Y. The evolution and functional characterization of CXC chemokines and receptors in lamprey. Dev. Comp. Immunol. 2021, 116, 103905. [Google Scholar] [CrossRef]
- Liao, Z.; Wan, Q.; Xiao, X.; Ji, J.; Su, J. A systematic investigation on the composition, evolution and expression characteristics of chemokine superfamily in grass carp Ctenopharyngodon idella. Dev. Comp. Immunol. 2018, 82, 72–82. [Google Scholar] [CrossRef]
- Liu, X.; Kang, L.; Liu, W.; Lou, B.; Wu, C.; Jiang, L. Molecular characterization and expression analysis of the large yellow croaker (Larimichthys crocea) chemokine receptors CXCR2, CXCR3, and CXCR4 after bacterial and poly I:C challenge. Fish. Shellfish Immunol. 2017, 70, 228–239. [Google Scholar] [CrossRef]
- Lu, X.J.; Chen, Q.; Rong, Y.J.; Chen, F.; Chen, J. CXCR3.1 and CXCR3.2 Differentially Contribute to Macrophage Polarization in Teleost Fish. J. Immunol. 2017, 198, 4692–4706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.H.; Liu, Q.; Xu, B.; Chen, W.; Yang, Q.; Wang, Z.X.; Sun, Y.H. Identification of nuclear localization sequence of CXCR4 in renal cell carcinoma by constructing expression plasmids of different deletants. Plasmid 2010, 63, 68–72. [Google Scholar] [CrossRef]
- Bajoghli, B.; Guo, P.; Aghaallaei, N.; Hirano, M.; Strohmeier, C.; McCurley, N.; Bockman, D.E.; Schorpp, M.; Cooper, M.D.; Boehm, T. A thymus candidate in lampreys. Nature 2011, 470, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, B.; Lee, A.P.; Ravi, V.; Maurya, A.K.; Lian, M.M.; Swann, J.B.; Ohta, Y.; Flajnik, M.F.; Sutoh, Y.; Kasahara, M.; et al. Elephant shark genome provides unique insights into gnathostome evolution. Nature 2014, 505, 174–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Clercq, E. Recent advances on the use of the CXCR4 antagonist plerixafor (AMD3100, Mozobil™) and potential of other CXCR4 antagonists as stem cell mobilizers. Pharmacology 2010, 128, 509–518. [Google Scholar] [CrossRef]
- Umasuthan, N.; Wan, Q.; Revathy, K.S.; Whang, I.; Noh, J.K.; Kim, S.; Park, M.A.; Lee, J. Molecular aspects, genomic arrangement and immune responsive mRNA expression profiles of two CXC chemokine receptor homologs (CXCR1 and CXCR2) from rock bream, Oplegnathus fasciatus. Fish. Shellfish Immunol. 2014, 40, 304–318. [Google Scholar] [CrossRef]
- Knaut, H.; Werz, C.; Geisler, R.; Nusslein-Volhard, C. A zebrafish homologue of the chemokine receptor Cxcr4 is a germ-cell guidance receptor. Nature 2003, 421, 279–282. [Google Scholar] [CrossRef]
- Boldajipour, B.; Mahabaleshwar, H.; Kardash, E.; Reichman-Fried, M.; Blaser, H.; Minina, S.; Wilson, D.; Xu, Q.; Raz, E. Control of chemokine-guided cell migration by ligand sequestration. Cell 2008, 132, 463–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, M.J.; Szadkowski, E.; Wendel, J.F. Homoeolog expression bias and expression level dominance in allopolyploid cotton. Heredity 2013, 110, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Pala, I.; Coelho, M.M.; Schartl, M. Dosage compensation by gene-copy silencing in a triploid hybrid fish. Curr. Biol. 2008, 18, 1344–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Liu, D.; Wang, X.; Ji, C.; Cheng, F.; Liu, B.; Hu, Z.; Chen, S.; Pental, D.; Ju, Y. The genome sequence of allopolyploid Brassica juncea and analysis of differential homoeolog gene expression influencing selection. Nat. Genet. 2016, 48, 1225–1232. [Google Scholar] [CrossRef] [PubMed]
- Konrad, F.M.; Reutershan, J. CXCR2 in Acute Lung Injury. Mediat. Inflamm. 2012, 2012, 740987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, S.F.; Huising, M.O.; Stakauskas, R.; Forlenza, M.; Lidy Verburg-van Kemenade, B.M.; Buchmann, K.; Nielsen, M.E.; Wiegertjes, G.F. Real-time gene expression analysis in carp (Cyprinus carpio L.) skin: Inflammatory responses to injury mimicking infection with ectoparasites. Dev. Comp. Immunol. 2007, 31, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Devalaraja, R.M.; Nanney, L.B.; Du, J.; Qian, Q.; Yu, Y.; Devalaraja, M.N.; Richmond, A. Delayed wound healing in CXCR2 knockout mice. J. Investig. Derm. 2000, 115, 234–244. [Google Scholar] [CrossRef] [Green Version]
- Yellowley, C. CXCL12/CXCR4 signaling and other recruitment and homing pathways in fracture repair. Bonekey Rep. 2013, 2, 300. [Google Scholar] [CrossRef] [Green Version]
- Knerlich-Lukoschus, F.; von der Ropp-Brenner, B.; Lucius, R.; Mehdorn, H.M.; Held-Feindt, J. Spatiotemporal CCR1, CCL3(MIP-1α), CXCR4, CXCL12(SDF-1α) expression patterns in a rat spinal cord injury model of posttraumatic neuropathic pain: Laboratory investigation. J. Neurosurg. 2011, 14, 583–597. [Google Scholar] [CrossRef]
- Liu, W.; Jin, Y.; Zhang, W.; Xiang, Y.; Jia, P.; Yi, M.; Jia, K. MiR-202-5p Inhibits RIG-I-dependent innate immune responses to RGNNV infection by targeting TRIM25 to mediate RIG-I ubiquitination. Viruses 2020, 12, 261. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. Omics 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhang, H.; Xiang, Y.; Jia, K.; Luo, M.; Yi, M. A novel germline and somatic cell expression of two sexual differentiation genes, Dmrt1 and Foxl2 in marbled goby (Oxyeleotris marmorata). Aquaculture 2020, 516, 734619. [Google Scholar] [CrossRef]



| Name | N-Glycosylation Sites | Count |
|---|---|---|
| BsCXCR1a | N21FTY, N25DSE (N-terminus), N191SSQ (ECL2) | 3 |
| BsCXCR1b | N13DTS, N19LTY (N-terminus), N185NSD, N200ASK (ECL2) N351GTV (C-terminus) | 5 |
| BsCXCR2 | N13TSR, N29SSY, N34TSF (N-terminus), N163LSS (ICL2), N209LTG (ECL2), N229MTV (TM6), N309VTQ (ECL3), N370TSV (C-terminus) | 8 |
| BsCXCR3a1 | N23DTW, N31YTV (N-terminus), N90WSV (ICL1), N287ITL (TM6), N301NTC (ECL3) | 5 |
| BsCXCR3a2 | N16GSY (N-terminus), N118VSF (TM3), N138ITR (ICL2) N262TTL (TM6) | 4 |
| BsCXCR4a | N14DTG (N-terminus) | 1 |
| BsCXCR4b | N13SSE, N17LTD (N-terminus) | 2 |
| BsCXCR5 | N6YTY, N13LTD (N-terminus), N178SSH (ECL2), N273ASC (ECL3), N338QSS (C-terminus) | 5 |
| BsCXCR6 | N11YTQ, N20DST, N29SSD (N-terminus), N366EST (C-terminus) | 4 |
| BsCXCR7 | N21ISE, N28IST (N-terminus), N42RSA, N334RTY, N363LSE (C-terminus) | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, M.; Zhang, H.; Mo, C.; Li, W.; Zhang, W.; Jia, K.; Liu, W.; Yi, M. The CXC Chemokine Receptors in Four-Eyed Sleeper (Bostrychus sinensis) and Their Involvement in Responding to Skin Injury. Int. J. Mol. Sci. 2021, 22, 10022. https://doi.org/10.3390/ijms221810022
Dong M, Zhang H, Mo C, Li W, Zhang W, Jia K, Liu W, Yi M. The CXC Chemokine Receptors in Four-Eyed Sleeper (Bostrychus sinensis) and Their Involvement in Responding to Skin Injury. International Journal of Molecular Sciences. 2021; 22(18):10022. https://doi.org/10.3390/ijms221810022
Chicago/Turabian StyleDong, Mengdan, Hong Zhang, Chengyu Mo, Wenjing Li, Wanwan Zhang, Kuntong Jia, Wei Liu, and Meisheng Yi. 2021. "The CXC Chemokine Receptors in Four-Eyed Sleeper (Bostrychus sinensis) and Their Involvement in Responding to Skin Injury" International Journal of Molecular Sciences 22, no. 18: 10022. https://doi.org/10.3390/ijms221810022
APA StyleDong, M., Zhang, H., Mo, C., Li, W., Zhang, W., Jia, K., Liu, W., & Yi, M. (2021). The CXC Chemokine Receptors in Four-Eyed Sleeper (Bostrychus sinensis) and Their Involvement in Responding to Skin Injury. International Journal of Molecular Sciences, 22(18), 10022. https://doi.org/10.3390/ijms221810022