
CD4+ T-Cell Plasticity in Non-Infectious Retinal Inflammatory Disease

Abstract
:
1. Introduction
2. Evidence of CD4+ T Cell Subsets in Monophasic and Relapsing EAU Models
3. Role of Th17/Th1 Cells in EAU
4. Evidence of Th17/Th1 Cells in Uveitis
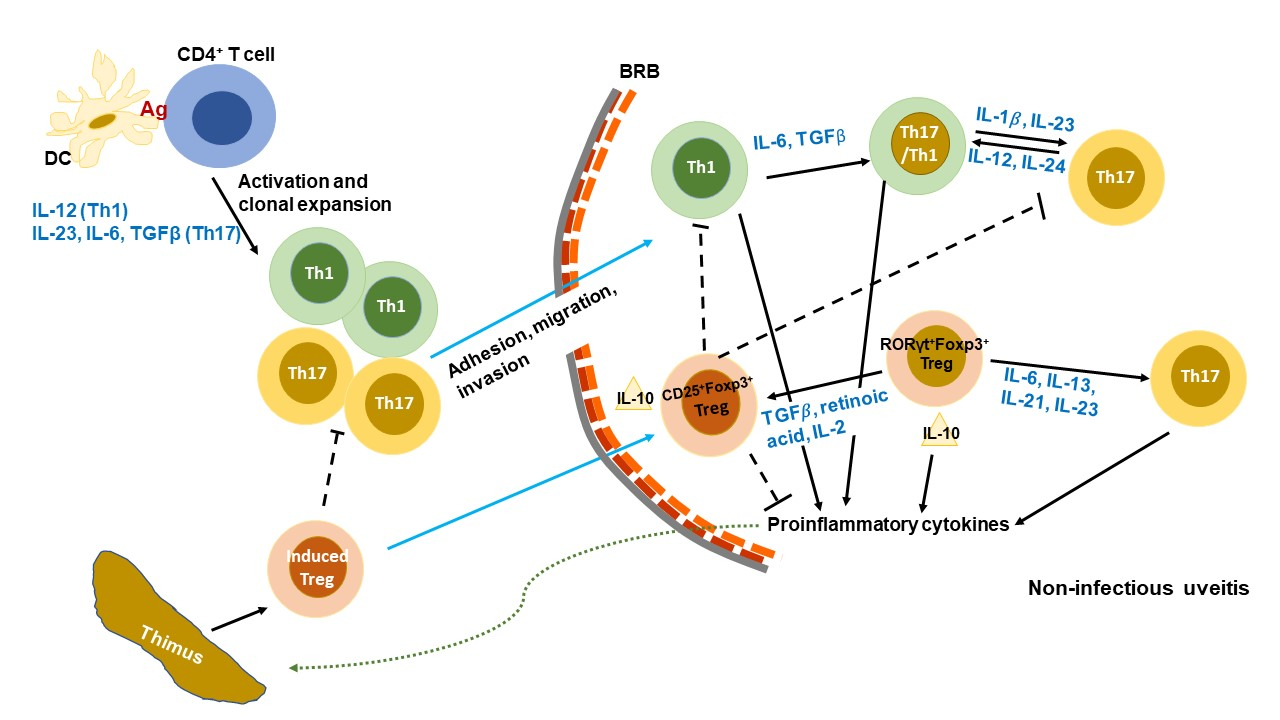
5. Importance of Treg/Th17 Cells in Autoimmune Disease and EAU
6. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| antigen presenting cells | APCs |
| non-infectious uveitis | NIU |
| blood-retinal barrier | BRB |
| interleukin-17 | IL-17 |
| interferon-γ | IFNγ |
| interleukin-10 | IL-10 |
| complete Freund’s adjuvant | CFA |
| experimental autoimmune uveitis | EAU |
| killed mycobacterial antigen | mTB |
| pertussis toxin | PTX |
References
- Tomkins-Netzer, O.; Talat, L.; Bar, A.; Lula, A.; Taylor, S.R.; Joshi, L.; Lightman, S. Long-Term Clinical Outcome and Causes of Vision Loss in Patients with Uveitis. Ophthalmology 2014, 121, 2387–2392. [Google Scholar] [CrossRef] [PubMed]
- García-Aparicio, A.; de Yébenes, M.J.G.; Otón, T.; Muñoz-Fernández, S. Prevalence and Incidence of Uveitis: A Systematic Review and Meta-analysis. Ophthalmic Epidemiol. 2021, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Forrester, J.V.; Kuffova, L.; Dick, A.D. Autoimmunity, Autoinflammation, and Infection in Uveitis. Am. J. Ophthalmol. 2018, 189, 77–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenbaum, J.T.; Dick, A.D. The Eyes Have it: A Rheumatologist’s View of Uveitis. Arthritis Rheumatol. 2018, 70, 1533–1543. [Google Scholar] [CrossRef]
- Wakefield, D.; Li, Q.; McCluskey, P.; Nussenblatt, R.B.; Chan, C.-C. Immunohistochemical localization of T lymphocytes and macrophages and expression of interferon gamma and defensin in uveitis. Ocul. Immunol. Inflamm. 1994, 2, 153–159. [Google Scholar] [CrossRef]
- Luger, D.; Silver, P.B.; Tang, J.; Cua, D.; Chen, Z.; Iwakura, Y.; Bowman, E.P.; Sgambellone, N.M.; Chan, C.-C.; Caspi, R.R. Either a Th17 or a Th1 effector response can drive autoimmunity: Conditions of disease induction affect dominant effector category. J. Exp. Med. 2008, 205, 799–810. [Google Scholar] [CrossRef]
- Caspi, R.R. Understanding Autoimmune Uveitis through Animal Models the Friedenwald Lecture. Investig. Opthalmol. Vis. Sci. 2011, 52, 1873–1879. [Google Scholar] [CrossRef]
- Yoshimura, T.; Sonoda, K.-H.; Miyazaki, Y.; Iwakura, Y.; Ishibashi, T.; Yoshimura, A.; Yoshida, H. Differential roles for IFN- and IL-17 in experimental autoimmune uveoretinitis. Int. Immunol. 2007, 20, 209–214. [Google Scholar] [CrossRef] [Green Version]
- Merrell, A.J.; Stanger, B.Z. Adult cell plasticity in vivo: De-differentiation and transdifferentiation are back in style. Nat. Rev. Mol. Cell Biol. 2016, 17, 413–425. [Google Scholar] [CrossRef]
- DuPage, M.; Bluestone, J.A. Harnessing the plasticity of CD4+ T cells to treat immune-mediated disease. Nat. Rev. Immunol. 2016, 16, 149–163. [Google Scholar] [CrossRef] [Green Version]
- Basdeo, S.A.; Cluxton, D.; Sulaimani, J.; Moran, B.; Canavan, M.; Orr, C.; Veale, D.; Fearon, U.; Fletcher, J.M. Ex-Th17 (Nonclassical Th1) Cells Are Functionally Distinct from Classical Th1 and Th17 Cells and Are Not Constrained by Regulatory T Cells. J. Immunol. 2017, 198, 2249–2259. [Google Scholar] [CrossRef]
- Panzer, M.; Sitte, S.; Wirth, S.; Drexler, I.; Sparwasser, T.; Voehringer, D. Rapid In Vivo Conversion of Effector T Cells into Th2 Cells during Helminth Infection. J. Immunol. 2011, 188, 615–623. [Google Scholar] [CrossRef] [Green Version]
- Blatner, N.R.; Mulcahy, M.F.; Dennis, K.L.; Scholtens, D.; Bentrem, D.J.; Phillips, J.D.; Ham, S.; Sandall, B.P.; Khan, M.W.; Mahvi, D.M.; et al. Expression of ROR t Marks a Pathogenic Regulatory T Cell Subset in Human Colon Cancer. Sci. Transl. Med. 2012, 4, 164ra159. [Google Scholar] [CrossRef] [Green Version]
- Caza, T.; Landas, S. Functional and Phenotypic Plasticity of CD4+T Cell Subsets. BioMed Res. Int. 2015, 2015, 521957. [Google Scholar] [CrossRef] [Green Version]
- Sallusto, F.; Cassotta, A.; Hoces, D.; Foglierini, M.; Lanzavecchia, A. Do Memory CD4 T Cells Keep Their Cell-Type Programming: Plasticity versus Fate Commitment? T-Cell Heterogeneity, Plasticity, and Selection in Humans. Cold Spring Harb. Perspect. Biol. 2017, 10, a029421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, R.K.; Caspi, R.R. Rodent Models of Experimental Autoimmune Uveitis. Autoimmunity 2004, 102, 395–420. [Google Scholar] [CrossRef] [Green Version]
- Caspi, R.R. Experimental Autoimmune Uveoretinitis in the Rat and Mouse. Curr. Protoc. Immunol. 2003, 53, 15.6.1–15.6.20. [Google Scholar] [CrossRef]
- Agrawal, R.; Barathi, V.A.; Iwata, D.; Bansal, S. Experimental autoimmune uveitis and other animal models of uveitis: An update. Indian J. Ophthalmol. 2015, 63, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Qian, H.; Horai, R.; Chan, C.-C.; Caspi, R. Mouse Models of Experimental Autoimmune Uveitis: Comparative Analysis of Adjuvant-Induced vs Spontaneous Models of Uveitis. Curr. Mol. Med. 2015, 15, 550–557. [Google Scholar] [CrossRef]
- Chen, J.; Caspi, R.R. Clinical and Functional Evaluation of Ocular Inflammatory Disease Using the Model of Experimental Autoimmune Uveitis. In Immunological Tolerance; Humana Press: New York, NY, USA, 2019; Volume 1899, pp. 211–227. [Google Scholar] [CrossRef]
- Diedrichs-Möhring, M.; Kaufmann, U.; Wildner, G. The immunopathogenesis of chronic and relapsing autoimmune uveitis—Lessons from experimental rat models. Prog. Retin. Eye Res. 2018, 65, 107–126. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, U.; Diedrichs-Möhring, M.; Wildner, G. Dynamics of Intraocular IFN-γ, IL-17 and IL-10-Producing Cell Populations during Relapsing and Monophasic Rat Experimental Autoimmune Uveitis. PLoS ONE 2012, 7, e49008. [Google Scholar] [CrossRef]
- Pepple, K.L.; Wilson, L.; Van Gelder, R.N. Comparison of Aqueous and Vitreous Lymphocyte Populations From Two Rat Models of Experimental Uveitis. Investig. Opthalmol. Vis. Sci. 2018, 59, 2504–2511. [Google Scholar] [CrossRef]
- Fan, N.-W.; Li, J.; Mittal, S.K.; Foulsham, W.; Elbasiony, E.; Huckfeldt, R.M.; Chauhan, S.K.; Chen, Y. Characterization of Clinical and Immune Responses in an Experimental Chronic Autoimmune Uveitis Model. Am. J. Pathol. 2020, 191, 425–437. [Google Scholar] [CrossRef]
- Tang, J.; Zhu, W.; Silver, P.B.; Su, S.-B.; Chan, C.-C.; Caspi, R.R. Autoimmune uveitis elicited with antigen-pulsed dendritic cells has a distinct clinical signature and is driven by unique effector mechanisms: Initial encounter with autoantigen defines disease phenotype. J. Immunol. 2007, 178, 5578–5587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horai, R.; Caspi, R.R. Microbiome and Autoimmune Uveitis. Front. Immunol. 2019, 10, 232. [Google Scholar] [CrossRef] [Green Version]
- Horai, R.; Silver, P.B.; Chen, J.; Agarwal, R.K.; Chong, W.P.; Jittayasothorn, Y.; Mattapallil, M.J.; Nguyen, S.; Natarajan, K.; Villasmil, R.; et al. Breakdown of immune privilege and spontaneous autoimmunity in mice expressing a transgenic T cell receptor specific for a retinal autoantigen. J. Autoimmun. 2013, 44, 21–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horai, R.; Chong, W.; Zhou, R.; Chen, J.; Silver, P.; Agarwal, R.; Caspi, R. Spontaneous Ocular Autoimmunity in Mice Expressing a Transgenic T Cell Receptor Specific to Retina: A Tool to Dissect Mechanisms of Uveitis. Curr. Mol. Med. 2015, 15, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Egwuagu, C.E.; Charukamnoetkanok, P.; Gery, I. Thymic expression of autoantigens correlates with resistance to autoimmune disease. J. Immunol. 1997, 159, 3109–3112. [Google Scholar] [PubMed]
- Heng, J.S.; Hackett, S.F.; Stein-O’Brien, G.L.; Winer, B.; Williams, J.; Goff, L.A.; Nathans, J. Comprehensive analysis of a mouse model of spontaneous uveoretinitis using single-cell RNA sequencing. Proc. Natl. Acad. Sci. USA 2019, 116, 26734–26744. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wan, F.; Song, J.; Tang, K.; Zheng, F.; Guo, J.; Guo, D.; Bi, H. Imbalance Between Th17 Cells and Regulatory T Cells During Monophasic Experimental Autoimmune Uveitis. Inflammation 2015, 39, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Amadi-Obi, A.; Yu, C.-R.; Liu, X.; Mahdi, R.M.; Clarke, G.L.; Nussenblatt, R.B.; Gery, I.; Lee, Y.S.; E Egwuagu, C. TH17 cells contribute to uveitis and scleritis and are expanded by IL-2 and inhibited by IL-27/STAT. Nat. Med. 2007, 13, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Yang, P.; Du, L.; Zhou, H.; Ren, X.; Kijlstra, A. Contribution of CD4+CD25+T Cells to the Regression Phase of Experimental Autoimmune Uveoretinitis. Investig. Opthalmol. Vis. Sci. 2010, 51, 383–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyu, C.; Bing, S.J.; Wandu, W.S.; Xu, B.; Shi, G.; Hinshaw, S.J.; Lobera, M.; Caspi, R.R.; Lu, L.; Yang, J.; et al. TMP778, a selective inhibitor of RORγt, suppresses experimental autoimmune uveitis development, but affects both Th17 and Th1 cell populations. Eur. J. Immunol. 2018, 48, 1810–1816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silver, P.; Horai, R.; Chen, J.; Jittayasothorn, Y.; Chan, C.-C.; Villasmil, R.; Kesen, M.R.; Caspi, R.R. Retina-Specific T Regulatory Cells Bring About Resolution and Maintain Remission of Autoimmune Uveitis. J. Immunol. 2015, 194, 3011–3019. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Eskandarpour, M.; Gondrand, A.; Zhang, X.; Gu, R.; Galatowicz, G.; Lightman, S.L.; Calder, V.L. Functionally distinct IFN-γ + IL-17A + Th cells in experimental autoimmune uveitis: T-cell heterogeneity, migration, and steroid response. Eur. J. Immunol. 2020, 50, 1941–1951. [Google Scholar] [CrossRef]
- Zou, Y.; Hu, X.; Schewitz-Bowers, L.P.; Stimpson, M.; Miao, L.; Ge, X.; Yang, L.; Li, Y.; Bible, P.W.; Wen, X.; et al. The DNA Methylation Inhibitor Zebularine Controls CD4+ T Cell Mediated Intraocular Inflammation. Front. Immunol. 2019, 10, 1950. [Google Scholar] [CrossRef] [Green Version]
- Maggi, L.; Santarlasci, V.; Capone, M.; Rossi, M.C.; Querci, V.; Mazzoni, A.; Cimaz, R.; De Palma, R.; Liotta, F.; Maggi, E.; et al. Distinctive features of classic and nonclassic (Th17 derived) human Th1 cells. Eur. J. Immunol. 2012, 42, 3180–3188. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Turner, H.; Maynard, C.L.; Oliver, J.R.; Chen, D.; Elson, C.O.; Weaver, C.T. Late Developmental Plasticity in the T Helper 17 Lineage. Immunity 2009, 30, 92–107. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Rodriguez, E.V.; Napolitani, G.; Lanzavecchia, A.; Sallusto, F. Interleukins 1β and 6 but not transforming growth factor-β are essential for the differentiation of interleukin 17–producing human T helper cells. Nat. Immunol. 2007, 8, 942–949. [Google Scholar] [CrossRef]
- Zielinski, C.E.; Mele, F.; Aschenbrenner, D.; Jarrossay, D.; Ronchi, F.; Gattorno, M.; Monticelli, S.; Lanzavecchia, A.; Sallusto, F. Pathogen-induced human TH17 cells produce IFN-γ or IL-10 and are regulated by IL-1β. Nature 2012, 484, 514–518. [Google Scholar] [CrossRef] [PubMed]
- Cosmi, L.; Liotta, F.; Maggi, E.; Romagnani, S.; Annunziato, F. Th17 and Non-Classic Th1 Cells in Chronic Inflammatory Disorders: Two Sides of the Same Coin. Int. Arch. Allergy Immunol. 2014, 164, 171–177. [Google Scholar] [CrossRef]
- Maggi, L.; Capone, M.; Giudici, F.; Santarlasci, V.; Querci, V.; Liotta, F.; Ficari, F.; Maggi, E.; Tonelli, F.; Annunziato, F.; et al. CD4+CD161+ T Lymphocytes Infiltrate Crohn’s Disease-Associated Perianal Fistulas and Are Reduced by Anti-TNF-a Local Therapy. Int. Arch. Allergy Immunol. 2013, 161, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Mazzoni, A.; Maggi, L.; Liotta, F.; Cosmi, L.; Annunziato, F. Biological and clinical significance of T helper 17 cell plasticity. Immunology 2019, 158, 287–295. [Google Scholar] [CrossRef] [Green Version]
- Mazzoni, A.; Maggi, L.; Siracusa, F.; Ramazzotti, M.; Rossi, M.C.; Santarlasci, V.; Montaini, G.; Capone, M.; Rossettini, B.; De Palma, R.; et al. Eomes controls the development of Th17-derived (non-classic) Th1 cells during chronic inflammation. Eur. J. Immunol. 2018, 49, 79–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stadhouders, R.; Lubberts, E.; Hendriks, R.W. A cellular and molecular view of T helper 17 cell plasticity in autoimmunity. J. Autoimmun. 2018, 87, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Leipe, J.; Pirronello, F.; Klose, A.; Schulze-Koops, H.; Skapenko, A. Increased plasticity of non-classic Th1 cells toward the Th17 phenotype. Mod. Rheumatol. 2019, 30, 930–936. [Google Scholar] [CrossRef] [PubMed]
- Geginat, J.; Paroni, M.; Kastirr, I.; Larghi, P.; Pagani, M.; Abrignani, S. Reverse plasticity: TGF-β and IL-6 induce Th1-to-Th17-cell transdifferentiation in the gut. Eur. J. Immunol. 2016, 46, 2306–2310. [Google Scholar] [CrossRef]
- Yan, J.; Wang, R.; Horng, T. mTOR Is Key to T Cell Transdifferentiation. Cell Metab. 2019, 29, 241–242. [Google Scholar] [CrossRef] [Green Version]
- Maggi, L.; Cimaz, R.; Capone, M.; Santarlasci, V.; Querci, V.; Simonini, G.; Nencini, F.; Liotta, F.; Romagnani, S.; Maggi, E.; et al. Brief Report: Etanercept Inhibits the Tumor Necrosis Factor α-Driven Shift of Th17 Lymphocytes Toward a Nonclassic Th1 Phenotype in Juvenile Idiopathic Arthritis. Arthritis Rheumatol. 2014, 66, 1372–1377. [Google Scholar] [CrossRef]
- Ma, Q.; Li, D.; Nurieva, R.; Patenia, R.; Bassett, R.; Cao, W.; Alekseev, A.M.; He, H.; Molldrem, J.J.; Kroll, M.H.; et al. Reduced Graft-versus-Host Disease in C3-Deficient Mice Is Associated with Decreased Donor Th1/Th17 Differentiation. Biol. Blood Marrow Transplant. 2012, 18, 1174–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, J.; Takai, K.; Fujiwara, N.; Arimitsu, N.; Ueda, Y.; Wakisaka, S.; Yoshikawa, H.; Kaneko, F.; Suzuki, T.; Suzuki, N. Excessive CD4+ T cells co-expressing interleukin-17 and interferon-γ in patients with Behçet’s disease. Clin. Exp. Immunol. 2011, 168, 68–74. [Google Scholar] [CrossRef]
- Hirota, K.; Duarte, J.H.; Veldhoen, M.; Hornsby, E.; Li, Y.; Cua, D.J.; Ahlfors, H.; Wilhelm, C.; Tolaini, M.; Menzel, U.; et al. Fate mapping of IL-17-producing T cells in inflammatory responses. Nat. Immunol. 2011, 12, 255–263. [Google Scholar] [CrossRef]
- Ramesh, R.; Kozhaya, L.; McKevitt, K.; Djuretic, I.M.; Carlson, T.J.; Quintero, M.A.; McCauley, J.L.; Abreu, M.T.; Unutmaz, D.; Sundrud, M.S. Pro-inflammatory human Th17 cells selectively express P-glycoprotein and are refractory to glucocorticoids. J. Exp. Med. 2014, 211, 89–104. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Chauhan, S.K.; Shao, C.; Omoto, M.; Inomata, T.; Dana, R. IFN-γ–Expressing Th17 Cells Are Required for Development of Severe Ocular Surface Autoimmunity. J. Immunol. 2017, 199, 1163–1169. [Google Scholar] [CrossRef] [Green Version]
- Fan, N.-W.; Dohlman, T.H.; Foulsham, W.; McSoley, M.; Singh, R.; Chen, Y.; Dana, R. The role of Th17 immunity in chronic ocular surface disorders. Ocul. Surf. 2020, 19, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Reinert-Hartwall, L.; Honkanen, J.; Salo, H.M.; Nieminen, J.; Luopajärvi, K.; Härkönen, T.; Veijola, R.; Simell, O.; Ilonen, J.; Peet, A.; et al. Th1/Th17 Plasticity Is a Marker of Advanced β Cell Autoimmunity and Impaired Glucose Tolerance in Humans. J. Immunol. 2014, 194, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Harbour, S.N.; Maynard, C.L.; Zindl, C.L.; Schoeb, T.R.; Weaver, C.T. Th17 cells give rise to Th1 cells that are required for the pathogenesis of colitis. Proc. Natl. Acad. Sci. USA 2015, 112, 7061–7066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciofani, M.; Madar, A.; Galan, C.; Sellars, M.; Mace, K.; Pauli, F.; Agarwal, A.; Huang, W.; Parkurst, C.N.; Muratet, M.; et al. A Validated Regulatory Network for Th17 Cell Specification. Cell 2012, 151, 289–303. [Google Scholar] [CrossRef] [Green Version]
- Chong, W.P.; Mattapallil, M.J.; Raychaudhuri, K.; Bing, S.J.; Wu, S.; Zhong, Y.; Wang, W.; Chen, Z.; Silver, P.B.; Jittayasothorn, Y.; et al. The Cytokine IL-17A Limits Th17 Pathogenicity via a Negative Feedback Loop Driven by Autocrine Induction of IL-24. Immunity 2020, 53, 384–397.e5. [Google Scholar] [CrossRef]
- Ronchi, F.; Basso, C.; Preite, S.; Reboldi, A.; Baumjohann, D.; Perlini, L.; Lanzavecchia, A.; Sallusto, F. Experimental priming of encephalitogenic Th1/Th17 cells requires pertussis toxin-driven IL-1β production by myeloid cells. Nat. Commun. 2016, 7, 11541. [Google Scholar] [CrossRef]
- Nishihara, H.; Soldati, S.; Mossu, A.; Rosito, M.; Rudolph, H.; Muller, W.A.; Latorre, D.; Sallusto, F.; Sospedra, M.; Martin, R.; et al. Human CD4+ T cell subsets differ in their abilities to cross endothelial and epithelial brain barriers in vitro. Fluids Barriers CNS 2020, 17, 1–18. [Google Scholar] [CrossRef]
- Kim, J.; Park, J.A.; Lee, E.Y.; Lee, Y.J.; Song, Y.W.; Lee, E.B. Imbalance of Th17 to Th1 cells in Behcet’s disease. Clin. Exp. Rheumatol. 2010, 28, S16–S19. [Google Scholar] [PubMed]
- Cetin, E.A.; Cosan, F.; Cefle, A.; Deniz, G. IL-22-secreting Th22 and IFN-γ-secreting Th17 cells in Behçet’s disease. Mod. Rheumatol. 2013, 24, 802–807. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, Y.; Lepzien, R.; Kullberg, S.; Eklund, A.; Smed-Sörensen, A.; Grunewald, J. Expanded lung T-bet+RORγT+ CD4+ T-cells in sarcoidosis patients with a favourable disease phenotype. Eur. Respir. J. 2016, 48, 484–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, R.M.; Zhang, X.; Sampson, R.; Ehrenstein, M.; Nguyen, D.X.; Chaudhry, M.; Mein, C.; Mahmud, N.; Galatowicz, G.; Tomkins-Netzer, O.; et al. Clinical Remission of Sight-Threatening Non-Infectious Uveitis Is Characterized by an Upregulation of Peripheral T-Regulatory Cell Polarized Towards T-bet and TIGIT. Front. Immunol. 2018, 9, 907. [Google Scholar] [CrossRef] [Green Version]
- Walscheid, K.; Neekamp, L.; Heiligenhaus, A.; Weinhage, T.; Heinz, C.; Foell, D. Increased Circulating Proinflammatory T Lymphocytes in Children with Different Forms of Anterior Uveitis: Results from a Pilot Study. Ocul. Immunol. Inflamm. 2018, 27, 788–797. [Google Scholar] [CrossRef]
- Schewitz-Bowers, L.P.; Lait, P.J.P.; Copland, D.A.; Chen, P.; Wu, W.; Dhanda, A.D.; Vistica, B.P.; Williams, E.L.; Liu, B.; Jawad, S.; et al. Glucocorticoid-resistant Th17 cells are selectively attenuated by cyclosporine A. Proc. Natl. Acad. Sci. USA 2015, 112, 4080–4085. [Google Scholar] [CrossRef] [Green Version]
- Maggi, L.; Mazzoni, A.; Cimaz, R.; Liotta, F.; Annunziato, F.; Cosmi, L. Th17 and Th1 Lymphocytes in Oligoarticular Juvenile Idiopathic Arthritis. Front. Immunol. 2019, 10, 450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hori, S.; Nomura, T.; Sakaguchi, S. Control of Regulatory T Cell Development by the Transcription Factor Foxp. Science 2003, 299, 1057–1061. [Google Scholar] [CrossRef] [Green Version]
- Roncador, G.; Brown, P.J.; Maestre, L.; Hue, S.; Martinez-Torrecuadrada, J.L.; Ling, K.-L.; Pratap, S.; Toms, C.; Fox, B.C.; Cerundolo, V.; et al. Analysis of FOXP3 protein expression in human CD4+CD25+ regulatory T cells at the single-cell level. Eur. J. Immunol. 2005, 35, 1681–1691. [Google Scholar] [CrossRef]
- Santamaria, J.C.; Borelli, A.; Irla, M. Regulatory T Cell Heterogeneity in the Thymus: Impact on Their Functional Activities. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Sonmez, C.; Yucel, A.A.; Yesil, T.H.; Kucuk, H.; Sezgin, B.; Mercan, R.; Yucel, A.E.; Demirel, G.Y. Correlation between IL-17A/F, IL-23, IL-35 and IL-12/-23 (p40) levels in peripheral blood lymphocyte cultures and disease activity in Behcet’s patients. Clin. Rheumatol. 2018, 37, 2797–2804. [Google Scholar] [CrossRef]
- Kawakami, R.; Kitagawa, Y.; Chen, K.Y.; Arai, M.; Ohara, D.; Nakamura, Y.; Yasuda, K.; Osaki, M.; Mikami, N.; Lareau, C.A.; et al. Distinct Foxp3 enhancer elements coordinate development, maintenance, and function of regulatory T cells. Immunity 2021, 54, 947–961.e8. [Google Scholar] [CrossRef] [PubMed]
- Foulsham, W.; Marmalidou, A.; Amouzegar, A.; Coco, G.; Chen, Y.; Dana, R. Review: The function of regulatory T cells at the ocular surface. Ocul. Surf. 2017, 15, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.J. The relationship between TIGIT+ regulatory T cells and autoimmune disease. Int. Immunopharmacol. 2020, 83, 106378. [Google Scholar] [CrossRef]
- Muhammad, F.; Wang, D.; McDonald, T.; Walsh, M.; Drenen, K.; Montieth, A.; Foster, C.S.; Lee, D.J. TIGIT+ A2Ar-Dependent anti-uveitic Treg cells are a novel subset of Tregs associated with resolution of autoimmune uveitis. J. Autoimmun. 2020, 111, 102441. [Google Scholar] [CrossRef]
- Chen, L.; Yang, P.; Zhou, H.; He, H.; Ren, X.; Chi, W.; Wang, L.; Kijlstra, A. Diminished Frequency and Function of CD4+CD25highRegulatory T Cells Associated with Active Uveitis in Vogt-Koyanagi-Harada Syndrome. Investig. Opthalmology Vis. Sci. 2008, 49, 3475–3482. [Google Scholar] [CrossRef] [Green Version]
- Ruggieri, S.; Frassanito, M.A.; Dammacco, R.; Guerriero, S. TregLymphocytes in Autoimmune Uveitis. Ocul. Immunol. Inflamm. 2012, 20, 255–261. [Google Scholar] [CrossRef]
- Bing, S.J.; Shemesh, I.; Chong, W.P.; Horai, R.; Jittayasothorn, Y.; Silver, P.B.; Sredni, B.; Caspi, R.R. AS101 ameliorates experimental autoimmune uveitis by regulating Th1 and Th17 responses and inducing Treg cells. J. Autoimmun. 2019, 100, 52–61. [Google Scholar] [CrossRef]
- Zhuang, Z.; Wang, Y.; Zhu, G.; Gu, Y.; Mao, L.; Hong, M.; Li, Y.; Zheng, M. Imbalance of Th17/Treg cells in pathogenesis of patients with human leukocyte antigen B27 associated acute anterior uveitis. Sci. Rep. 2017, 7, 40414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Chen, S.; Sun, D.; Li, X.; Wei, R.; Li, X.; Nian, H. miR-223-3p promotes autoreactive Th17 cell responses in experimental autoimmune uveitis (EAU) by inhibiting transcription factor FOXO3 expression. FASEB J. 2019, 33, 13951–13965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horwitz, D.A. Identity of mysterious CD4+CD25-Foxp3+ cells in SLE. Arthritis Res. Ther. 2010, 12, 101–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Żabińska, M.; Krajewska, M.; Kościelska-Kasprzak, K.; Jakuszko, K.; Bartoszek, D.; Myszka-Kozłowska, M.; Klinger, M. CD4+CD25+CD127− and CD4+CD25+Foxp3+ Regulatory T Cell Subsets in Mediating Autoimmune Reactivity in Systemic Lupus Erythematosus Patients. Arch. Immunol. Ther. Exp. 2016, 64, 399–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, S.G.; Wang, J.H.; Gray, J.D.; Soucier, H.; Horwitz, D.A. Natural and Induced CD4+CD25+ Cells Educate CD4+CD25− Cells to Develop Suppressive Activity: The Role of IL-2, TGF-β, and IL-10. J. Immunol. 2004, 172, 5213–5221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komatsu, N.; Mariotti-Ferrandiz, E.; Wang, Y.; Malissen, B.; Waldmann, H.; Hori, S. Heterogeneity of natural Foxp3+T cells: A committed regulatory T-cell lineage and an uncommitted minor population retaining plasticity. Proc. Natl. Acad. Sci. USA 2009, 106, 1903–1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komatsu, N.; Okamoto, K.; Sawa, S.; Nakashima, T.; Oh-Hora, M.; Kodama, T.; Tanaka, S.; Bluestone, J.A.; Takayanagi, H. Pathogenic conversion of Foxp3+ T cells into TH17 cells in autoimmune arthritis. Nat. Med. 2013, 20, 62–68. [Google Scholar] [CrossRef]
- Hovhannisyan, Z.; Treatman, J.; Littman, D.R.; Mayer, L. Characterization of Interleukin-17–Producing Regulatory T Cells in Inflamed Intestinal Mucosa from Patients With Inflammatory Bowel Diseases. Gastroenterology 2011, 140, 957–965. [Google Scholar] [CrossRef] [Green Version]
- Ueno, A.; Jijon, H.; Chan, R.; Ford, K.; Hirota, C.; Kaplan, G.G.; Beck, P.L.; Iacucci, M.; Gasia, M.F.; Barkema, H.W.; et al. Increased Prevalence of Circulating Novel IL-17 Secreting Foxp3 Expressing CD4+ T Cells and Defective Suppressive Function of Circulating Foxp3+ Regulatory Cells Support Plasticity Between Th17 and Regulatory T Cells in Inflammatory Bowel Disease Patients. Inflamm. Bowel Dis. 2013, 19, 2522–2534. [Google Scholar] [CrossRef]
- Chen, J.; Ye, H.; Xiao, W.; Mao, Y.; Ai, S.; Chen, R.; Lian, X.; Shi, L.; Wang, X.; Bi, S.; et al. Increased Dysfunctional and Plastic Regulatory T Cells in Idiopathic Orbital Inflammation. Front. Immunol. 2021, 12, 1612. [Google Scholar] [CrossRef]
- Coursey, T.G.; Bian, F.; Zaheer, M.; Pflugfelder, S.C.; Volpe, E.A.; De Paiva, C.S. Age-related spontaneous lacrimal keratoconjunctivitis is accompanied by dysfunctional T regulatory cells. Mucosal Immunol. 2016, 10, 743–756. [Google Scholar] [CrossRef]

{kind=link}
| Mouse Model | Method of Induction | Duration of Retinal Inflammation | Patterns of Intraocular T Cells |
|---|---|---|---|
| Monophasic Lewis rat EAU [8,31] | R16 in CFA and mTB H37Ra. PTX ip. | Effector phase: 9–18 dpi Peak phase: 14 dpi | Th17: Highest levels at peak disease, declining thereafter Treg: Levels increase at peak disease, remaining high throughout [31] Th17/Th1 and Th17/Treg: ND |
| Monophasic Lewis rat EAU [22] | PDSAg in CFA and mTB H37Ra. | Effector phase: 11–21 dpi Onset: 11–13 dpi Peak phase: 13–16 dpi | Th1 and Th17: levels remain stable throughout FoxP3+ Treg: peak at resolution Th17/Th1: Levels increase from onset, peaking at resolution phase Th17/Treg, Th1/Treg: not detected throughout |
| Relapsing Lewis rat EAU [22] | R14 in CFA and mTB H37Ra. | Effector phase: 7–23 dpi Onset: 7–9 dpi Peak phase: 9–13 dpi Relapse: 23 dpi | Th1: Gradually increase from onset, peaking at resolution and relapse stage Th17: Highest at onset, then gradually declining. FoxP3+ Treg: peak at resolution phase Th17/Th1: stable at 10% throughout Th17/Treg, Th1/Treg: not detected throughout |
| Monophasic and relapsing classic EAU in B10. RIII & B10.A mice [6] | IRBP1–20 in CFA, mTB H37Ra, +/− PTX | Onset: ~10–12 dpi Peak: 14–21 dpi Retinal atrophy phase starts 4–5 weeks pi | Th1: Dominant at peak and disease relapse [32] Th17: Dominant at induction phase Treg: Increase at peak disease and remain at high levels thereafter [33] Th17/Th1: Comprises 10% of CD4+ T cells at peak disease [34] |
| Monophasic and relapsing classic mouse EAU in C57/Bl6 [6,32] | IRBP161–180 in CFA, mTb H37Ra, PTX | Effective phase: 7–28 dpi Onset: ~10–12 dpi Peak: 14–21 dpi Retinal atrophy phase starts 4–5 weeks pi | Th1: Dominant at peak disease and during relapse [32]. Th17: Dominant at induction phase Treg: Appears at peak disease and increases thereafter [35] Th17/Th1: 5–10% at peak disease [36,37]. Treg/Th17 cells: ND |
| Chronic EAU in B10.RIII mice [24] | IRBP1–20 in CFA, mTb H37Ra, PTX | Effector phase: 14 dpi Peak and plateau: 10 wk pi | Peak phase: memory CD44hi Th17 cells dominate with minimal Th1 cells. Treg: NDTh17/Th1: less than 1% at 12 wk pi Treg/Th17: ND |
| Chronic EAU in C57/Bl6 mice [24] | IRBP1–20 in CFA, mTb H37Ra, PTX | Effector phase: 21 dpi Peak and plateau: week 7 pi | Peak disease: memory CD44hi Th17 cells dominate, with fewer than 1% Th1 cell involvement. Treg: appears at 14 dpi, gradually increasing thereafter [35] Th17/Th1: less than 1% at 12 wk pi Treg/Th17: ND |
| Relapsing uveitogenic DC induced-EAU [25] | Infusion of IRBP-pulsed mature DC (sc) to B10.RIII mice | Inconsistent results. Onset: 12–14 dpi. Remains active for 3 months. | Th1 and Th2 responses dominate at peak disease Th17 response is minimal Treg: ND Th17/Th1 and Th17/Treg cells: ND |
| IRBP-specific (R161H) B10.RIII TCR Transgenic mice [27,28] | The model generates high proportion of effector CD4+ T cells specific to IRBP161–180 | Onset: 5–6 wk. Peak: 8–10 wk. Remains active for 4 months. | Th1 and Th17: both pathogenic, with Th1 cells being dominant cell type. Both polarised Th1 and Th17 cells are susceptible to EAU. Th17/Th1: presented < 1% at peak disease. Treg: Less Treg were detected intraocularly in R161H mice than the wild type EAU and they were generated extrathymically. |
| AIRE−/− EAU [30] | Spontaneous model of EAU due to central tolerance | Effective phase: 5–14 weeks | Th1 cells dominant and a milder Th17 response Th17/Th1 and T17/Treg cells: ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-H.; Lightman, S.; Calder, V.L. CD4+ T-Cell Plasticity in Non-Infectious Retinal Inflammatory Disease. Int. J. Mol. Sci. 2021, 22, 9584. https://doi.org/10.3390/ijms22179584
Chen Y-H, Lightman S, Calder VL. CD4+ T-Cell Plasticity in Non-Infectious Retinal Inflammatory Disease. International Journal of Molecular Sciences. 2021; 22(17):9584. https://doi.org/10.3390/ijms22179584
Chicago/Turabian StyleChen, Yi-Hsing, Sue Lightman, and Virginia L. Calder. 2021. "CD4+ T-Cell Plasticity in Non-Infectious Retinal Inflammatory Disease" International Journal of Molecular Sciences 22, no. 17: 9584. https://doi.org/10.3390/ijms22179584
APA StyleChen, Y.-H., Lightman, S., & Calder, V. L. (2021). CD4+ T-Cell Plasticity in Non-Infectious Retinal Inflammatory Disease. International Journal of Molecular Sciences, 22(17), 9584. https://doi.org/10.3390/ijms22179584