
Therapeutic Effects of Aripiprazole in the 5xFAD Alzheimer’s Disease Mouse Model

 ,
,
Abstract
1. Introduction
2. Results
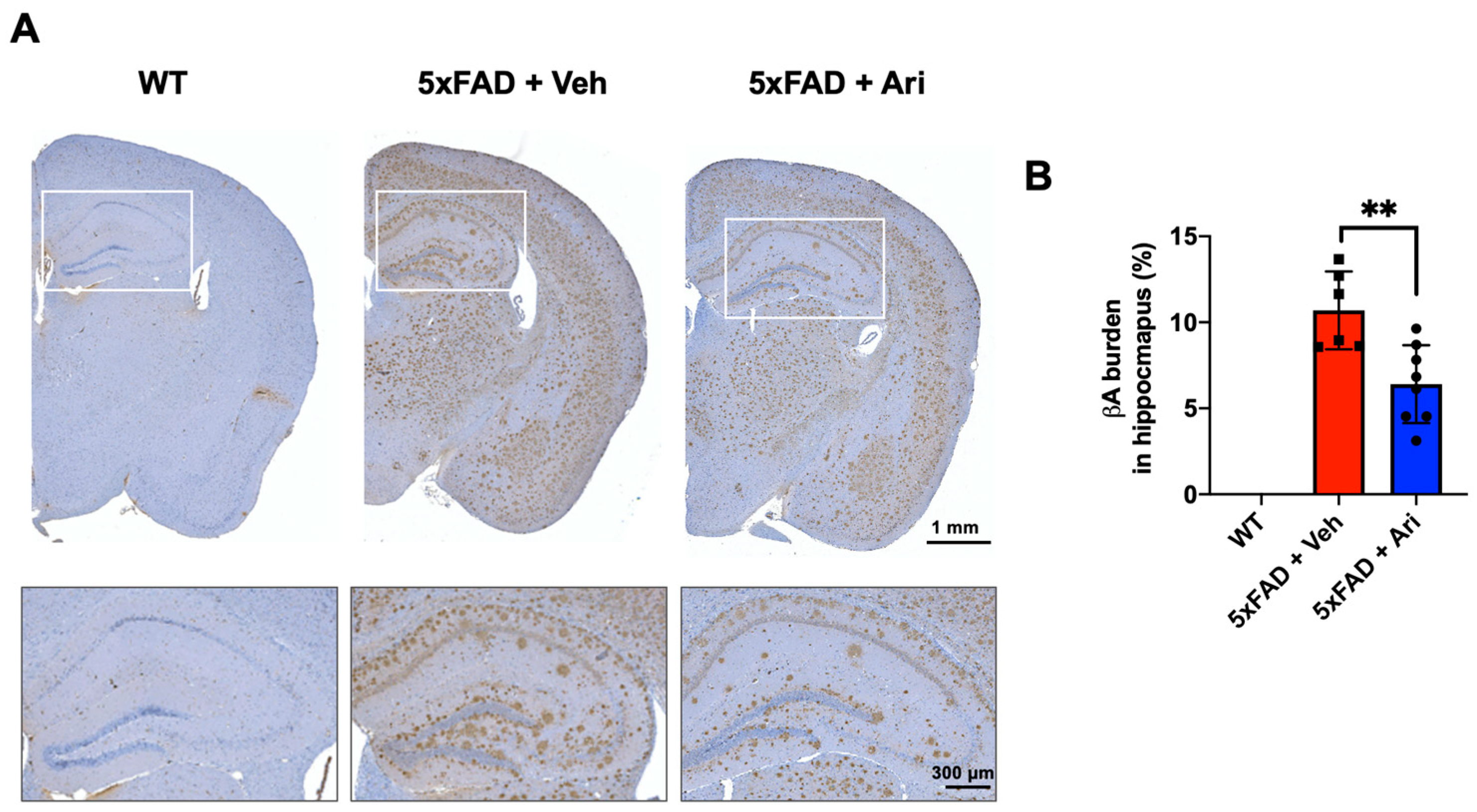
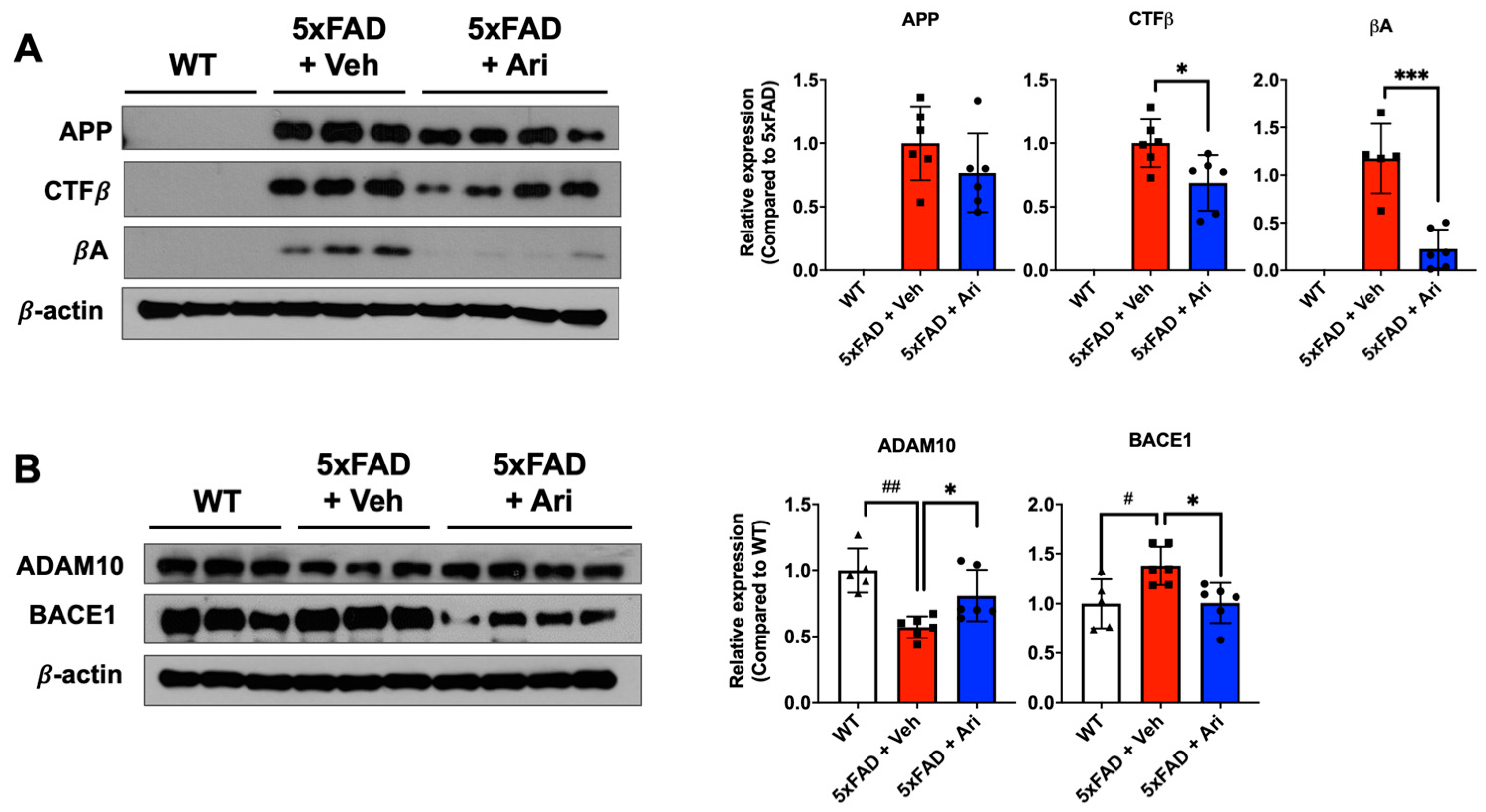
2.1. Aripiprazole Effectively Inhibited βA Deposition in the Brain of 5xFAD AD Mice
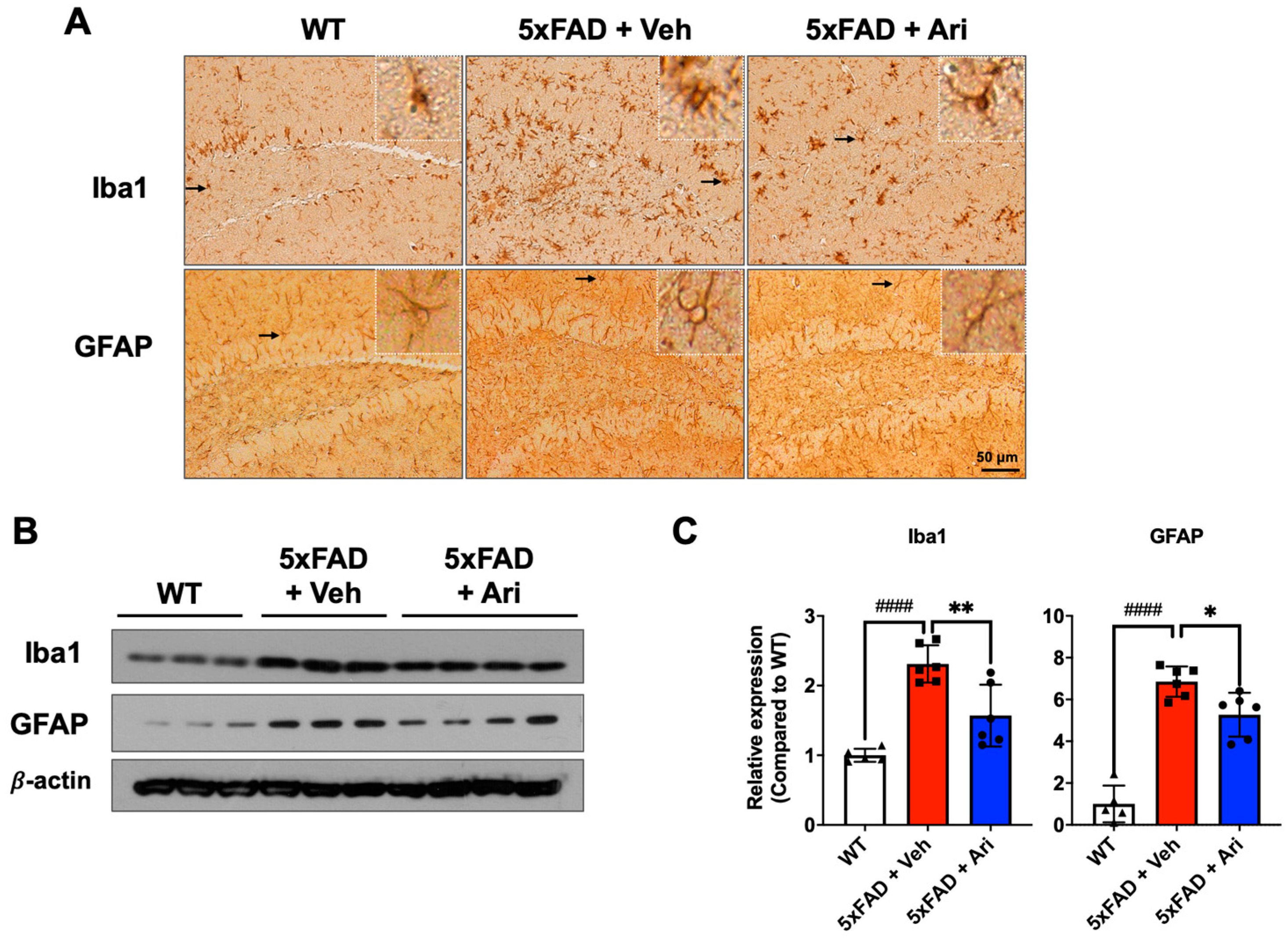
2.2. Aripiprazole Suppressed Neuroinflammation in 5xFAD AD Mice
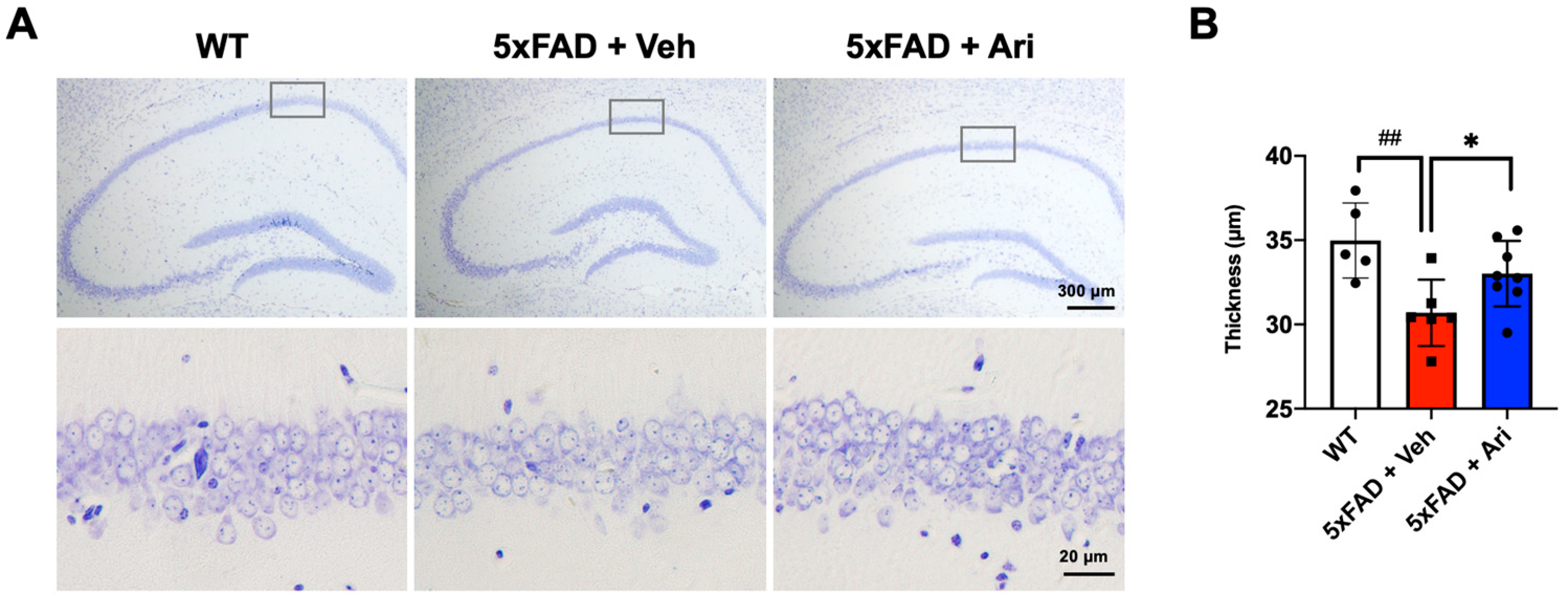
2.3. Aripiprazole Prevented Neuronal Loss in the Hippocampus of 5xFAD Mice
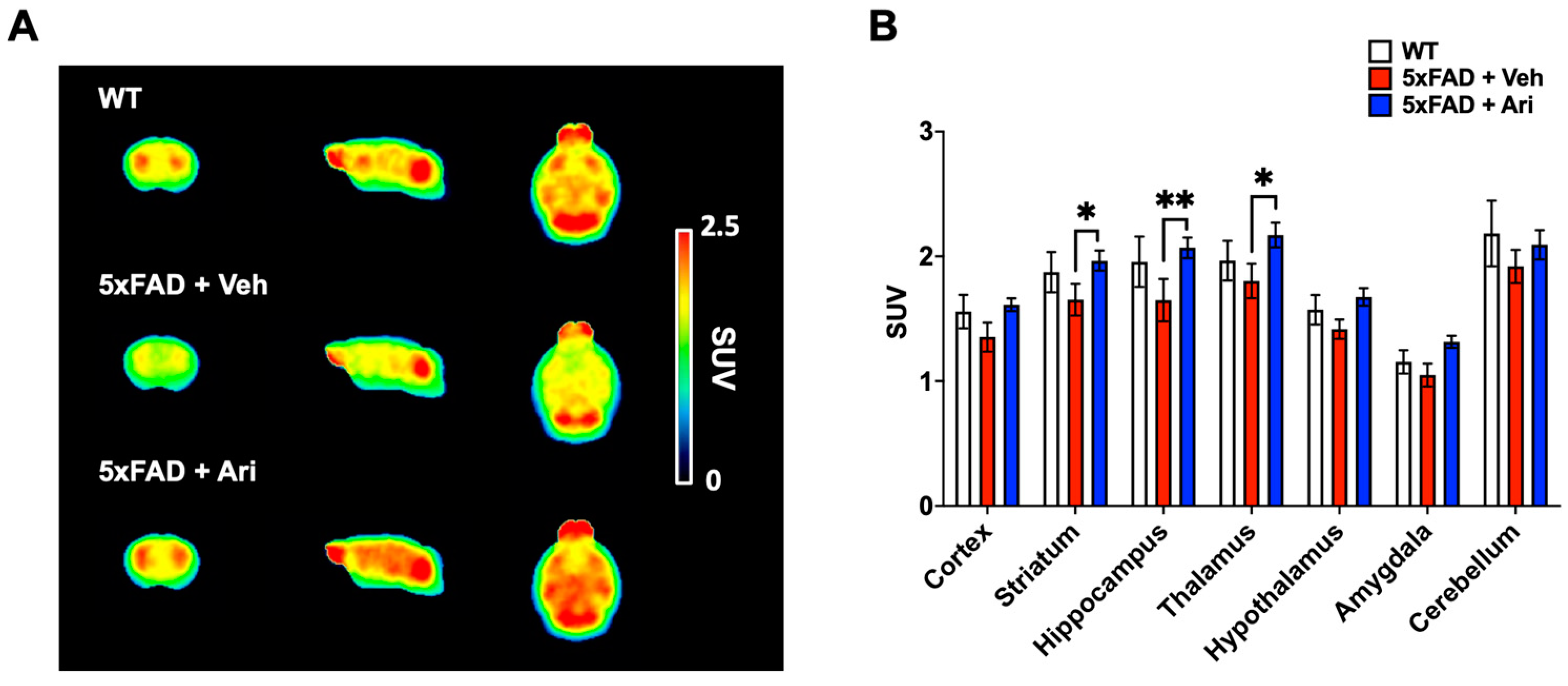
2.4. Aripiprazole Effectively Prevented the Decline of Cerebral Glucose Metabolism in 5xFAD Mice
3. Discussion
4. Materials and Methods
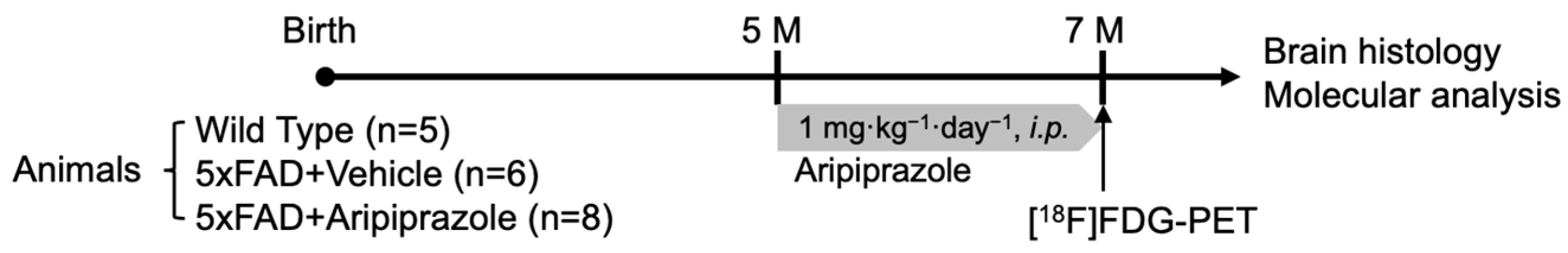
4.1. Animals and Drug Administration
4.2. Immunohistochemistry (IHC)
4.3. Cresyl Violet Staining
4.4. Western Blotting
4.5. Positron Emission Tomography (PET) Scans
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Christina, P. World Alzheimer’s Report; Alzheimer’s Disease International: London, UK, 2018; Volume 2018, pp. 1–48. [Google Scholar]
- Jack, C.R., Jr.; Knopman, D.S.; Jagust, W.J.; Petersen, R.C.; Weiner, M.W.; Aisen, P.S.; Shaw, L.M.; Vemuri, P.; Wiste, H.J.; Weigand, S.D.; et al. Tracking pathophysiological processes in Alzheimer’s disease: An updated hypothetical model of dynamic biomarkers. Lancet Neurol. 2013, 12, 207–216. [Google Scholar] [CrossRef]
- Khan, A.; Corbett, A.; Ballard, C. Emerging treatments for Alzheimer’s disease for non-amyloid and non-tau targets. Expert Rev. Neurother. 2017, 17, 683–695. [Google Scholar] [CrossRef]
- Citron, M. Strategies for disease modification in Alzheimer’s disease. Nat. Rev. Neurosci. 2004, 5, 677–685. [Google Scholar] [CrossRef]
- Selkoe, D.J.; Hardy, J. The amyloid hypothesis of Alzheimer’s disease at 25 years. EMBO Mol. Med. 2016, 8, 595–608. [Google Scholar] [CrossRef]
- Cummings, J.L.; Morstorf, T.; Zhong, K. Alzheimer’s disease drug-development pipeline: Few candidates, frequent failures. Alzheimers Res. Ther. 2014, 6, 37. [Google Scholar] [CrossRef]
- Knopman, D.S. Lowering of amyloid-beta by beta-secretase inhibitors—Some informative failures. N. Engl. J. Med. 2019, 380, 1476–1478. [Google Scholar] [CrossRef]
- Rafii, M.S.; Aisen, P.S. The search for Alzheimer disease therapeutics—Same targets, better trials? Nat. Rev. Neurol. 2020, 16, 597–598. [Google Scholar] [CrossRef]
- Ballard, C.; Aarsland, D.; Cummings, J.; O’Brien, J.; Mills, R.; Molinuevo, J.L.; Fladby, T.; Williams, G.; Doherty, P.; Corbett, A.; et al. Drug repositioning and repurposing for Alzheimer disease. Nat. Rev. Neurol. 2020, 16, 661–673. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Zhu, S.; Liu, X.; Zhou, Y.; Nussinov, R.; Cheng, F. deepDR: A network-based deep learning approach to in silico drug repositioning. Bioinformatics 2019, 35, 5191–5198. [Google Scholar] [CrossRef] [PubMed]
- Kohen, I.; Lester, P.E.; Lam, S. Antipsychotic treatments for the elderly: Efficacy and safety of aripiprazole. Neuropsychiatr. Dis. Treat. 2010, 6, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Pae, C.U. A review of the safety and tolerability of aripiprazole. Expert Opin. Drug Saf. 2009, 8, 373–386. [Google Scholar] [CrossRef]
- Kirino, E. Efficacy and safety of aripiprazole in child and adolescent patients. Eur. Child Adolesc. Psychiatry 2012, 21, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Pae, C.U.; Forbes, A.; Patkar, A.A. Aripiprazole as adjunctive therapy for patients with major depressive disorder: Overview and implications of clinical trial data. CNS Drugs 2011, 25, 109–127. [Google Scholar] [CrossRef]
- Streim, J.E.; Porsteinsson, A.P.; Breder, C.D.; Swanink, R.; Marcus, R.; McQuade, R.; Carson, W.H. A randomized, double-blind, placebo-controlled study of aripiprazole for the treatment of psychosis in nursing home patients with Alzheimer disease. Am. J. Geriatr. Psychiatry 2008, 16, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, M.; Hasebe, S.; Kawamoto, N.; Shiba, T.; Yamaguchi, T.; Kikuta, M.; Shuto, M.; Ogita, K. Beneficial in vivo effect of aripiprazole on neuronal regeneration following neuronal loss in the dentate gyrus: Evaluation using a mouse model of trimethyltin-induced neuronal loss/self-repair in the dentate gyrus. J. Pharm. Sci. 2014, 124, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Besagar, S.; Radabaugh, H.L.; Bleimeister, I.H.; Meyer, E.A.; Niesman, P.J.; Cheng, J.P.; Bondi, C.O.; Kline, A.E. Aripiprazole and environmental enrichment independently improve functional outcome after cortical impact injury in adult male rats, but their combination does not yield additional benefits. Exp. Neurol. 2019, 314, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Mutlu, A.; Mutlu, O.; Ulak, G.; Akar, F.; Kaya, H.; Erden, F.; Tanyeri, P. Superior effects of quetiapine compared with aripiprazole and iloperidone on MK-801-induced olfactory memory impairment in female mice. Biomed. Rep. 2017, 6, 567–570. [Google Scholar] [CrossRef][Green Version]
- Son, Y.; Jeong, Y.J.; Kwon, J.H.; Choi, H.D.; Pack, J.K.; Kim, N.; Lee, Y.S.; Lee, H.J. 1950 MHz radiofrequency electromagnetic fields do not aggravate memory deficits in 5xFAD mice. Bioelectromagnetics 2016, 37, 391–399. [Google Scholar] [CrossRef]
- Lee, M.; Lee, H.J.; Jeong, Y.J.; Oh, S.J.; Kang, K.J.; Han, S.J.; Nam, K.R.; Lee, Y.J.; Lee, K.C.; Ryu, Y.H.; et al. Age dependency of mGluR5 availability in 5xFAD mice measured by PET. Neurobiol. Aging 2019, 84, 208–216. [Google Scholar] [CrossRef]
- Postina, R.; Schroeder, A.; Dewachter, I.; Bohl, J.; Schmitt, U.; Kojro, E.; Prinzen, C.; Endres, K.; Hiemke, C.; Blessing, M.; et al. A disintegrin-metalloproteinase prevents amyloid plaque formation and hippocampal defects in an Alzheimer disease mouse model. J. Clin. Investig. 2004, 113, 1456–1464. [Google Scholar] [CrossRef]
- Picada, J.N.; Dos Santos Bde, J.; Celso, F.; Monteiro, J.D.; Da Rosa, K.M.; Camacho, L.R.; Vieira, L.R.; Freitas, T.M.; Da Silva, T.G.; Pontes, V.M.; et al. Neurobehavioral and genotoxic parameters of antipsychotic agent aripiprazole in mice. Acta Pharm. Sin. 2011, 32, 1225–1232. [Google Scholar] [CrossRef][Green Version]
- Heo, H.J.; Park, S.Y.; Lee, Y.S.; Shin, H.K.; Hong, K.W.; Kim, C.D. Combination therapy with cilostazol, aripiprazole, and donepezil protects neuronal cells from beta-amyloid neurotoxicity through synergistically enhanced SIRT1 expression. Korean J. Physiol. Pharm. 2020, 24, 299–310. [Google Scholar] [CrossRef]
- Kato, T.; Mizoguchi, Y.; Monji, A.; Horikawa, H.; Suzuki, S.O.; Seki, Y.; Iwaki, T.; Hashioka, S.; Kanba, S. Inhibitory effects of aripiprazole on interferon-gamma-induced microglial activation via intracellular Ca2+ regulation in vitro. J. Neurochem. 2008, 106, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Racki, V.; Marcelic, M.; Stimac, I.; Petric, D.; Kucic, N. Effects of haloperidol, risperidone, and aripiprazole on the immunometabolic properties of BV-2 microglial cells. Int. J. Mol. Sci. 2021, 22, 4399. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Fukuyama, H.; Senda, M.; Ishii, K.; Maeda, K.; Yamamoto, Y.; Ouchi, Y.; Ishii, K.; Okumura, A.; Fujiwara, K.; et al. Prediction of outcomes in mild cognitive impairment by using 18F-FDG-PET: A multicenter study. J. Alzheimers Dis. 2015, 45, 543–552. [Google Scholar] [CrossRef]
- Zhang, S.; Han, D.; Tan, X.; Feng, J.; Guo, Y.; Ding, Y. Diagnostic accuracy of 18 F-FDG and 11 C-PIB-PET for prediction of short-term conversion to Alzheimer’s disease in subjects with mild cognitive impairment. Int. J. Clin. Pract. 2012, 66, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Bohnen, N.I.; Djang, D.S.; Herholz, K.; Anzai, Y.; Minoshima, S. Effectiveness and safety of 18F-FDG PET in the evaluation of dementia: A review of the recent literature. J. Nucl. Med. 2012, 53, 59–71. [Google Scholar] [CrossRef]
- Bolmont, T.; Haiss, F.; Eicke, D.; Radde, R.; Mathis, C.A.; Klunk, W.E.; Kohsaka, S.; Jucker, M.; Calhoun, M.E. Dynamics of the microglial/amyloid interaction indicate a role in plaque maintenance. J. Neurosci. 2008, 28, 4283–4292. [Google Scholar] [CrossRef]
- Takkinen, J.S.; López-Picón, F.R.; Al Majidi, R.; Eskola, O.; Krzyczmonik, A.; Keller, T.; Löyttyniemi, E.; Solin, O.; Rinne, J.O.; Haaparanta-Solin, M. Brain energy metabolism and neuroinflammation in ageing APP/PS1-21 mice using longitudinal (18)F-FDG and (18)F-DPA-714 PET imaging. J. Cereb. Blood Flow Metab. 2017, 37, 2870–2882. [Google Scholar] [CrossRef]
- Krstic, D.; Madhusudan, A.; Doehner, J.; Vogel, P.; Notter, T.; Imhof, C.; Manalastas, A.; Hilfiker, M.; Pfister, S.; Schwerdel, C.; et al. Systemic immune challenges trigger and drive Alzheimer-like neuropathology in mice. J. Neuroinflammation 2012, 9, 151. [Google Scholar] [CrossRef]
- Oakley, H.; Cole, S.L.; Logan, S.; Maus, E.; Shao, P.; Craft, J.; Guillozet-Bongaarts, A.; Ohno, M.; Disterhoft, J.; Van Eldik, L.; et al. Intraneuronal beta-amyloid aggregates, neurodegeneration, and neuron loss in transgenic mice with five familial Alzheimer’s disease mutations: Potential factors in amyloid plaque formation. J. Neurosci. 2006, 26, 10129–10140. [Google Scholar] [CrossRef] [PubMed]
- Sadleir, K.R.; Eimer, W.A.; Cole, S.L.; Vassar, R. Aβ reduction in BACE1 heterozygous null 5XFAD mice is associated with transgenic APP level. Mol. Neurodegener. 2015, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Sadleir, K.R.; Popovic, J.; Vassar, R. ER stress is not elevated in the 5XFAD mouse model of Alzheimer’s disease. J. Biol. Chem. 2018, 293, 18434–18443. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Brain Uptake Value (SUV) | ||||||
|---|---|---|---|---|---|---|---|
| Cortex | Striatum | Hippocampus | Thalamus | Hypothalamus | Amygdala | Cerebellum | |
| WT | 1.56 ± 0.13 | 1.87 ± 0.16 | 1.96 ± 0.20 | 1.97 ± 0.16 | 1.57 ± 0.12 | 1.15 ± 0.09 | 2.18 ± 0.26 |
| 5xFAD + Veh | 1.35 ± 0.12 | 1.65 ± 0.13 | 1.65 ± 0.17 | 1.80 ± 0.14 | 1.42 ± 0.08 | 1.05 ± 0.09 | 1.92 ± 0.13 |
| 5xFAD + Ari | 1.61 ± 0.05 | 1.97 * ± 0.08 | 2.07 ** ± 0.08 | 2.17 * ± 0.10 | 1.67 ± 0.07 | 1.32 ± 0.05 | 2.09 ± 0.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, Y.J.; Son, Y.; Park, H.-J.; Oh, S.J.; Choi, J.Y.; Ko, Y.-G.; Lee, H.-J. Therapeutic Effects of Aripiprazole in the 5xFAD Alzheimer’s Disease Mouse Model. Int. J. Mol. Sci. 2021, 22, 9374. https://doi.org/10.3390/ijms22179374
Jeong YJ, Son Y, Park H-J, Oh SJ, Choi JY, Ko Y-G, Lee H-J. Therapeutic Effects of Aripiprazole in the 5xFAD Alzheimer’s Disease Mouse Model. International Journal of Molecular Sciences. 2021; 22(17):9374. https://doi.org/10.3390/ijms22179374
Chicago/Turabian StyleJeong, Ye Ji, Yeonghoon Son, Hye-Jin Park, Se Jong Oh, Jae Yong Choi, Young-Gyu Ko, and Hae-June Lee. 2021. "Therapeutic Effects of Aripiprazole in the 5xFAD Alzheimer’s Disease Mouse Model" International Journal of Molecular Sciences 22, no. 17: 9374. https://doi.org/10.3390/ijms22179374
APA StyleJeong, Y. J., Son, Y., Park, H.-J., Oh, S. J., Choi, J. Y., Ko, Y.-G., & Lee, H.-J. (2021). Therapeutic Effects of Aripiprazole in the 5xFAD Alzheimer’s Disease Mouse Model. International Journal of Molecular Sciences, 22(17), 9374. https://doi.org/10.3390/ijms22179374